
Abstract
Theria represent an extant clade that comprises placental and marsupial mammals. Here we report on the discovery of a new Late Cretaceous mammal from southern Patagonia, Patagomaia chainko gen. et sp. nov., represented by hindlimb and pelvic elements with unambiguous therian features. We estimate Patagomaia chainko attained a body mass of 14 kg, which is considerably greater than the 5 kg maximum body mass of coeval Laurasian therians. This new discovery demonstrates that Gondwanan therian mammals acquired large body size by the Late Cretaceous, preceding their Laurasian relatives, which remained small-bodied until the beginning of the Cenozoic. Patagomaia supports the view that the Southern Hemisphere was a cradle for the evolution of modern mammalian clades, alongside non-therian extinct groups such as meridiolestidans, gondwanatherians and monotremes.
Similar content being viewed by others


Introduction
Mesozoic mammals are usually regarded as small-sized, insectivore, shrew-like creatures of nocturnal habits1,2,3,4. This traditional view has recently changed thanks to discoveries that notably increased the ecological and morphological disparity of Mesozoic mammals, including swimming, burrowing and gliding taxa5,6,7,8,9,10, as well as dog-sized forms11,12,13. Among them, the fossil record of Late Cretaceous therian mammals from the Laurasian continents includes a wide array of small-bodied shrew-like insectivores of therian affinities1,2. Because the fossil record of modern-line mammals is notably rich in the Northern Hemisphere, it has been usually argued that the early evolution and origin of therians occurred exclusively on Laurasian landmasses, with therian mammals being rare—if not entirely absent—in Gondwana14,15,16,17,18. However, some molecular analyses19,20,21,22,23,24,25,26 in conjunction with sparce paleontological data27,28,29,30,31,32,33,34,35 are lending support to the view that some therian lineages, at least, evolved and diversified in the Southern Hemisphere in Late Mesozoic times.
Here we report on a therian mammal from the Mesozoic of Patagonia. Present discovery is of high relevance, because it supports the idea that southern landmasses constituted an important theatre for the early evolution of modern mammals. It is also indicative of the persistence of wide gaps in the fossil record of the Southern Hemisphere, up to now representing about 5% of the global fossil record of Mesozoic mammals33.
Class Mammalia Linnaeus 1758
Subclass Theria Parker and Haswell, 1897
Patagomaia nov. gen.
Generic diagnosis
Patagomaia is a large mammal (~ 14 kg) distinguished by the following unique combination of character states: fused acetabulum with a complete rim lacking a dorsal emargination; femur with subspherical head having a well-defined fovea capitis; femur head separated from the rest of the bone by a well-defined and medially tilted neck; lesser trochanter of femur small and located on the posteromedial surface of the shaft; distal end of femur with nearly symmetrical distal condyles and reduced epicondyles. Patagomaia further differs from other Mesozoic mammals in having the autapomorphic condition of a thick, well-defined, and obliquely oriented intercondylar ridge delimiting a deep fossa at the distal end of the femur.
Type species
Patagomaia chainko sp. nov.
Etymology
Patago, from Patagonia; maia, mother in Greek.
Patagomaia chainko sp. nov.
Zoobank registration: urn:lsid:zoobank.org:pub:39766F43-6876-4460-9830-D80646C312CC
Holotype
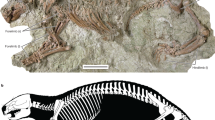
MPM-PV-23365 (Museo Padre Molina, Río Gallegos, Santa Cruz, Argentina), associated postcranial remains including the distal end of the left ulna, two fragments of the preacetabular wing of the left ilium, acetabular region of the left hemipelvis, fragment of the ischial blade, proximal end of the right femur; distal end of the left femur, proximal end of the left tibia, and other indeterminate bone fragments (Fig. 1; Supplementary Figs. 1, 3, 4, 6, 7).

Images of Patagomaia chainko holotype remains, MPM-PV-23365 References: (a) fragments of the left pelvis; (b) silhouette and skeletal scheme with details of the preserved bones; (c) distal end of the left ulna; (d) proximal end of the right femur and distal end of the left femur; e, proximal half of the left tibia. Scale bar: 20 mm.
Referred material
MPM-PV-23366, partial left acetabulum and ischium (see Supplementary Fig. 2); incomplete right femoral shaft (see Supplementary Fig. 5).
Etymology
The species name is derived from the Aonikenk language: chaink, large and ko, bone.
Diagnosis
The same as for genus by monotypy.
Type locality and age
La Anita Farm, Santa Cruz, Argentina. The remains were collected at a new site (S 50° 30′ 39.888″, W 72° 33′ 18.035″) geographically close to and at the same stratigraphic level as the Isasicursor 2 site (Chorrillo Formation, lower Maastrichtian, Upper Cretaceous)36,37. Both the holotype and referred specimens were found within an area of about 20 × 30 m. In close proximity, fossil wood and indeterminate hadrosaurid remains were found.
Description
The preacetabular wing of the ilium has a well-defined acetabular ridge that separates the dorsal gluteal fossa from the ventral iliac fossa, as in other early diverging therians38,39,40. There is a thickened surface for the origin of the m. rectus femoris anterior to the acetabulum, similar to some placental mammals but in contrast to other therians41, non-therian mammals and some early diverging therians (e.g., multituberculates Akidolestes, Henkelotherium, Vincelestes, Ukhaatherium)41,42,43 in which there is a distinct preacetabular tubercle (see Supplementary Information 1; Supplementary Fig. 8).
The pubis, ischium, and ilium of Patagomaia are strongly fused in the acetabular region, in sharp contrast with the unfused condition in many non-therian mammaliaforms (e.g., morganucodontids, dryolestoids, docodontans, multituberculates, eutriconodonts)7,11,42,43,44,45,46,47.
In Patagomaia, the acetabulum is deep and subspherical, and it is surrounded by a complete rim lacking the dorsal emargination that characterizes non-therian mammaliaforms (e.g., docodontans, multituberculates, Vincelestes; see Supplementary Figs. 1, 2, 11, 16)7,45. The acetabular rim shows a prominent anterior wall, corresponding with a subspherical femoral head. The lunate surface of the acetabulum is wide, with a deep acetabular fossa posteroventrally continuous with a narrow acetabular notch. Although the iliopubic eminence is represented only by its base, it appears to have been robust, as in monotremes and some therians42, but different from many other Mesozoic taxa (e.g., Adalatherium, Vincelestes; see Supplementary Fig. 11).
The femoral head of Patagomaia is clearly offset from the rest of the bone, unlike non-therian mammals in which it is close to the main axis of the femur (e.g., monotremes, Akidolestes, Vincelestes)42. The femoral neck is constricted and medially oriented, forming an angle of about 55° with the main axis of femur, whereas non-therian mammals lack a constricted femoral neck (e.g., monotremes, Akidolestes, Vincelestes)42. The trochanteric fossa is well-defined, wide, and distally bounded by a crescent-shaped crest for the insertion of the m. quadratus femoris39,40, in contrast to the shallow poorly defined fossa in non-therian mammals (e.g., Akidolestes, Henkelotherium, Vincelestes)42,43. The greater trochanter is robust, anteroposteriorly broad, and projected posteriorly; it reaches the same level as the femoral head proximally, resembling many therians39,48,49 but unlike non-therian mammals (e.g., monotremes, multituberculates, Henkelotherium, Vincelestes)42,43,44,45,50,51. The lesser trochanter of Patagomaia is relatively small and posteroventrally displaced. A thick crest extends posterodistally from the base of the greater trochanter. Such a crest is usually associated with the presence of a prominent third trochanter, suggesting that this trochanter was also developed in Patagomaia but is broken off and missing from the holotype (Supplementary Figs. 3 and 11).
The proximal end of the femur of Patagomaia lacks diagnostic features of multituberculates, such as the presence of a post-trochanteric fossa, a prominent greater trochanter that extends proximally beyond the femoral head and is separated from it by a deep incisure, a plate-like lesser trochanter at the confluence of the greater trochanter and neck that strongly protrudes ventrally, or the presence of a subtrochanteric tubercle45,50,51.
The posterior position of the lesser trochanter gives the femur of Patagomaia a subquadrangular contour in proximal view, as in therian mammals (e.g., Argyrolagus, Ukhaatherium, leptictids, Meniscotherium, Microgale)41,49,52. This condition differs from most non-therian mammaliaforms (e.g., morganucodontans, docodontans, Henkelotherium, Vincelestes, Necrolestes)43,53,54 in which the greater trochanter, femoral head, and lesser trochanter are almost aligned (see Supplementary Fig. 9).
The distal end of the femur of Patagomaia is transversely narrow, with the distal condyles proximo-distally low and nearly symmetrical in size and shape, in contrast with most non-therian mammals (e.g., morganucodontans, monotremes, Haldanodon; Docofossor, Necrolestes)44,53,54,55 in which the distal condyles are markedly asymmetrical (see Supplementary Fig. 10). The femoral condyles of Patagomaia are separated posteriorly by a deep but narrow intercondylar groove. There is a thick and prominent intercondylar ridge that is obliquely oriented and is distally delimited by a pit-like concavity (see Supplementary Fig. 4). This condition is unknown in most other mammals, being observed in only a few extant therians (e.g., Arctictis, Hystrix, Meles; see Supplementary Fig. 12). In Patagomaia, the patellar surface is represented by a shallow concavity delimited by low crests.
The tibia of Patagomaia shows symmetrical proximal facets for articulation with the femoral condyles, in contrast to the asymmetrical articular facets of non-therian mammaliaforms (e.g., morganucodontans, docodontans, Akidolestes, Vincelestes; see Supplementary Figs. 6)7,42,54. A deep and well-defined fossa extends along the posterior surface of the shaft of the tibia, a trait only seen in some eutherians (e.g., leptictids56, periptychids57, carnivores; see Supplementary Fig. 13). The anterior tibial tubercle is low and extends distally as a crest.
Available elements show that they belong to a medium-sized mammal, comparable in size to a the canid Lycalopex culpaeus. Following methodologies described in the SI, we estimated a body mass of ~ 14 kg for the holotype specimen of Patagomaia chainko (Supplementary Information 4; Supplementary Table 1; Supplementary Fig. 16). The histology of the femur and tibia of Patagomaia reveals that the cortex is composed of parallel-fibered and lamellar bone tissue and sparse/moderate density of longitudinal canals, closely resembling some Mesozoic eutherians (e.g., Barunlestes, Zalambdalestes)58. The presence of a well-defined External Fundamental System indicates the somatic maturity of the holotype of Patagomaia chainko (Fig. 2; Supplementary Information 2). This suggests that the size and body mass estimated here for the holotype of Patagomaia chainko likely represents the maximum ones that this animal could have reached.

Simplified time-calibrated cladogram showing the phylogenetic affinities of Patagomaia chainko, geographic location and paleohistological images. The simplified cladogram shows our interpretation unifying the three analyses conducted (see Supplementary Information 5). Map showing the fossil locality. The specimen here reported was recovered at the new site located at S 50° 30′ 39.888″ and W 72° 33′ 18.035″, close to the Isasicursor 2 site (marked with a red star) in the Maastrichtian Chorillo Formation. Transverse section of the femur (left) showing the External Fundamental System (white arrowheads); and tibia (right) in polarized light with lambda compensator. ER, Erosion room; PFB, parallel-fibered bone tissue; TB, trabecular bone; VC, vascular canal. Scale bar: 0.75 mm.
Discussion
To analyse the phylogenetic affinities of Patagomaia chainko, it was scored into three different comprehensive mammalian data matrices13,59,60. The incomplete nature of the holotype specimen, plus the fact that most of these datasets heavely rest on skull and teeth information, we were able to score just a small number of characters (less than 2.5%) for Patagomaia. However, the analyses consistently recover Patagomaia within Theria (Supplementary Information 5; Supplementary Figs. 17, 18 and 19). Synapomorphic features of Theria7,44,47,52 present in Patagomaia are: acetabulum completely fused and devoid of a dorsal emargination; femur with well-differentiated and medially-oriented head and neck; lesser trochanter small and posteroventrally displaced; distal end of femur with symmetrical articular condyles and reduced epicondyles; proximal end of tibia with symmetrical articular facets (in agreement with the condition of distal femoral condyles). This set of characters clearly distinguishes Patagomaia from non-therian mammals of the Southern Hemisphere, such as meridiolestidans (i.e., Necrolestes), multituberculates50, and gondwanatherians (i.e., Adalatherium)13.
It is worth noting that, although Patagomaia reveals therian affinities, it differs from Paleogene South American representatives of this clade (e.g., notoungulates, litopterns, astrapotherians, xenarthrans, and sparassodonts) in having a shallow patellar groove delimited by poorly defined crests that do not form a deep trochlea. In sum, Patagomaia does not exhibit morphological features that may ally it with any of the mammalian clades (i.e., Gondwanatheria, Dryolestoidea, Marsupialia, Ungulata) frequently recorded in Cretaceous and early Paleogene beds from South America. The prevailing view has been that the early evolutionary radiation of therian mammals occurred in the Northern Hemisphere15,16,17 although it has overlooked the occurrences of therians and stem-therians in the Cretaceous of Africa, Madagascar, India and South America27,28,29,30,31,32,33. In this context, Patagomaia constitutes an important addition to the meagre record of Mesozoic therians from Gondwana and indicates that some stages of early therian evolution occurred in the Southern Hemisphere19,20,21,22,23,24,25,26.
We estimate that the body mass of the holotype individual of Patagomaia ranged between 2.6 and 26 kg, with an average estimate of approximately 14 kg (a value obtained using the mean of 14 different regressions; see Materials and Methods and Supplementary Information 4). Even the smallest estimates would place it among the larger Mesozoic mammals, while the average and higher estimates exceed by far those of the largest Mesozoic mammals previously known: the Early Cretaceous Chinese eutriconodont Repenomamus (approximately 10 kg), and the Late Cretaceous gondwanatherian Vintana (8.9 kg). This would make Patagomaia the largest known Mesozoic mammal.
Most mammalian taxa recorded in Campanian–Maastrichtian assemblages of the Northern Hemisphere have an estimated body mass below 100 g, and only 1% of them are estimated to have reached a body mass of 1 kg61. In sharp contrast, at least 17 valid mammalian taxa (mainly belonging to Monotremata, Gondwanatheria and Meridiolestida) are known from Patagonian faunal assemblages, eight of which surpass 1 kg in body mass (see Supplementary Information 4). Similarly, large-bodied (much larger than 1 kg) mammals such as Vintana sertichi and Adalatherium hui are known from the latest Cretaceous of Madagascar53. This suggests that the evolution of larger body size among Gondwanan mammalian taxa began prior to the end-Cretaceous mass extinction event, thus preceding their Laurasian relatives in reaching large body size by at least 5 million years.
This new discovery demonstrates that Late Cretaceous mammalian faunas from South America were taxonomically diverse, not only including gondwanatherians, dryolestoids and monotremes, but also early therians. Patagomaia also reveals that the evolution of large body size among Late Cretaceous mammals was more complex than previously understood.
Materials and methods
Description
We follow the terminology of postcranial anatomy and myology used by Muizon38, Argot39,40,62,63, Fostowicz-Frelik64, Gambaryan et al.65 and Warburton et al.66. Paleohistological methods are explained in the Supplementary Information 3.
Body mass estimation
Estimates of the body mass of Patagomaia were made by taking measurements of the postcranial remains, using regressions that have already been used in other fossil mammals. Measurements were taken with a digital caliper. In addition, other regressions, based on cranial and dental measurements already published by other authors, were used to calculate the body mass of other mammals from the Cretaceous of South America. The averages of all regressions were calculated and compared with the results obtained in Patagomaia. The methodology used for the body mass estimations is explained in the Supplementary Information 4.
Phylogenetic analyses
Three data matrices were used with the aim of testing the phylogenetic position of Patagomaia chainko among mammals. We chose these data sets because they are very complete and updated matrices, and each one is focused on different groups of Mesozoic mammals (see Supplementary Information 5).
Data availability
The datasets analyzed during the current study are included in this published article (and its Supplementary Information file).
References
Bakker, R. T. Dinosaur physiology and the origin of mammals. Evolution 25, 636–658 (1971).
Hopson, J. A. Endothermy, small size, and the origin of mammalian reproduction. Am. Nat. 107, 446–452 (1973).
Kielan-Jaworowska, Z., Cifelli, R. L. & Luo, Z. X. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure (Columbia University Press, New York, 2004).
O’Leary, M. A. et al. The placental mammal ancestor and the post–K-Pg radiation of placentals. Science 339, 662–667 (2013).
Ji, Q., Luo, Z. X., Yuan, C. X. & Tabrum, A. R. A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science 311, 1123–1127 (2006).
Luo, Z. X. & Wible, J. R. A Late Jurassic digging mammal and early mammalian diversification. Science 308, 103–107 (2005).
Luo, Z. X. et al. Evolutionary development in basal mammaliaforms as revealed by a docodontan. Science 347, 760–764 (2015).
Meng, Q. J. et al. An arboreal docodont from the Jurassic and mammaliaform ecological diversification. Science 347, 764–768 (2015).
Meng, Q. J. et al. New gliding mammaliaforms from the Jurassic. Nature 548, 291–296 (2017).
Luo, Z. X. et al. New evidence for mammaliaform ear evolution and feeding adaptation in a Jurassic ecosystem. Nature 548, 326–329 (2017).
Hu, Y., Meng, J., Wang, Y. & Li, C. Large Mesozoic mammals fed on young dinosaurs. Nature 433, 149–152 (2005).
Krause, D. W. et al. First cranial remains of a gondwanatherian mammal reveal remarkable mosaicism. Nature 515, 512–517 (2014).
Krause, D. W. et al. Skeleton of a Cretaceous mammal from Madagascar reflects long-term insularity. Nature 581, 421–427 (2020).
Bonaparte, J. F. Approach to the significance of the Late Cretaceous mammals of South America. Berliner geowiss Abh E 13, 1–44 (1994).
Archibald, J. D. Timing and biogeography of the eutherian radiation: Fossils and molecules compared. Mol. Phylogenet. Evol. 28, 350–359 (2003).
Hunter, J. P. & Janis, C. M. Spiny Norman in the Garden of Eden? Dispersal and early biogeography of Placentalia. J. Mammal. Evol. 13, 89–123 (2006).
Wible, J. R., Rougier, G. W., Novacek, M. J. & Asher, R. J. Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary. Nature 447, 1003–1006 (2007).
Rougier, G. W., Martinelli, A. G. & Forasiepi, A. M. Mesozoic Mammals from South America and Their Forerunners (Springer Nature, Berlin, 2021).
Springer, M. S., Murphy, W. J., Eizirik, E. & O’Brien, S. J. Placental mammal diversification and the Cretaceous-Tertiary boundary. PNAS 100, 1056–1061 (2003).
Springer, M. S., Meredith, R. W., Janecka, J. E. & Murphy, W. J. The historical biogeography of Mammalia. Philos. Trans. R. Soc. B: Biol. Sci. 366, 2478–2502 (2011).
Murphy, W. J. et al. Resolution of the early placental mammal radiation using Bayesian phylogenetics. Science 294, 2348–2351 (2001).
Madsen, O. et al. Parallel adaptive radiations in two major clades of placental mammals. Nature 409, 610–614 (2001).
Scally, M. et al. Molecular evidence for the major clades of placental mammals. J. Mammal. Evol. 8, 239–277 (2001).
Upham, N. S., Esselstyn, J. A. & Jetz, W. Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLOS Biol. 17, e3000494. https://doi.org/10.1371/journal.pbio.3000494 (2019).
Beck, R. M. & Baillie, C. Improvements in the fossil record may largely resolve current conflicts between morphological and molecular estimates of mammal phylogeny. Proc. R. Soc. B 285(1893), 20181632 (2018).
Upham, N. S., Esselstyn, J. A. & Jetz, W. Molecules and fossils tell distinct yet complementary stories of mammal diversification. Curr. Biol. 31, 4195–4206 (2021).
Prasad, G. V., Jaeger, J. J., Sahni, A., Gheerbrant, E. & Khajuria, C. K. Eutherian mammals from the upper Cretaceous (Maastrichtian) intertrappean beds of Naskal, Andhra Pradesh, India. J. Vert. Paleontol. 14, 260–277 (1994).
Prasad, G. V. et al. First mammal evidence from the Late Cretaceous of India for biotic dispersal between India and Africa at the KT transition. CR Palevol. 9, 63–71 (2010).
Sigogneau-Russell, D., Hooker, J. J. & Ensom, P. C. The oldest tribosphenic mammal from Laurasia (Purbeck Limestone Group, Berriasian, Cretaceous, UK) and its bearing on the ‘dual origin’of Tribosphenida. C. R. Acad. Sci. S II A Earth Planet. Sci. 333, 141–147 (2001).
Krause, D. W. Fossil molar from a Madagascan marsupial. Nature 412, 497–498 (2001).
Averianov, A. O., Archibald, J. D. & Martin, T. Placental nature of the alleged marsupial from the Cretaceous of Madagascar. Acta Palaeontol. Pol. 48, 149–151 (2003).
Rana, R. S. & Wilson, G. P. New Late Cretaceous mammals from the Intertrappean beds of Rangapur, India and paleobiogeographic framework. Acta Palaeontol. Pol. 48, 331–348 (2003).
Goswami, A. et al. A radiation of arboreal basal eutherian mammals beginning in the Late Cretaceous of India. PNAS 108, 16333–16338 (2011).
Chimento, N., Agnolín, F. L. & Martinelli, A. G. Mesozoic mammals from South America: Implications for understanding early mammalian faunas from Gondwana. Contr. Mus. Argent. Cienc. Nat. 6, 199–209 (2016).
Castro, M. C. et al. A Late Cretaceous mammal from Brazil and the first radioisotopic age for the Bauru Group. R. Soc. Open Sci. 5, 180482 (2018).
Moyano-Paz, D. et al. The uppermost Cretaceous continental deposits at the southern end of Patagonia, the Chorrillo Formation case study (Austral-Magallanes Basin): Sedimentology, fossil content and regional implications. Cretaceous Res. 130, 105059 (2022).
Novas, F. et al. Paleontological discoveries in the Chorrillo Formation (upper Campanian-lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina. Rev. Mus Argent. Cienc. Nat. 21, 217–293 (2019).
Muizon, C. D. Mayulestes ferox, a borhyaenoid (Metatheria, Mammalia) from the early Palaeocene of Bolivia. Phylogenetic and paleobiologic implications. Geodiversitas 20, 19–142 (1998).
Argot, C. Functional-adaptive analysis of the hindlimb anatomy of extant marsupials and the paleobiology of the Paleocene marsupials Mayulestes ferox and Pucadelphys andinus. J. Morphol. 253, 76–108 (2002).
Argot, C. Postcranial analysis of a carnivoran-like archaic ungulate: The case of Arctocyon primaevus (Arctocyonidae, Mammalia) from the late Paleocene of France. J. Mammal. Evol. 20, 83–114 (2013).
Horovitz, I. Postcranial skeleton of Ukhaatherium nessovi (Eutheria, Mammalia) from the late Cretaceous of Mongolia. J. Vert. Paleontol. 23, 857–868 (2003).
Chen, M. & Luo, Z. X. Postcranial skeleton of the Cretaceous mammal Akidolestes cifellii and its locomotor adaptations. J. Mammal. Evol. 20, 159–189 (2013).
Jäger, K. R. K., Luo, Z. X. & Martin, T. Postcranial skeleton of Henkelotherium guimarotae (Cladotheria, Mammalia) and locomotor adaptation. J. Mammal. Evol. 27, 349–372 (2020).
Jenkins, F. A. Jr. & Parrington, F. R. The postcranial skeletons of the Triassic mammals Eozostrodon, Megazostrodon and Erythrotherium. Philos. Trans. R. Soc. Lond. B Biol. Sci. 273, 387–431 (1976).
Krause, D. W. & Jenkins, F. A. The postcranial skeleton of North American multituberculates. Bull. Mus. Comp. Zool. 150, 199–246 (1983).
Hurum, J. H. & Kielan-Jaworowska, Z. Postcranial skeleton of a Cretaceous multituberculate mammal Catopsbaatar. Acta Palaeontol. Pol. 53, 545–566 (2008).
Panciroli, E. et al. New species of mammaliaform and the cranium of Borealestes (Mammaliformes: Docodonta) from the Middle Jurassic of the British Isles. Zool. J. Linnean Soc. 192, 1323–1362 (2021).
Candela, A. M. & Picasso, M. B. Functional anatomy of the limbs of Erethizontidae (Rodentia, Caviomorpha): Indicators of locomotor behavior in Miocene porcupines. J. Morphol. 269, 552–593 (2008).
Abello, M. A. & Candela, A. M. Paleobiology of Argyrolagus (Marsupialia, Argyrolagidae): An astonishing case of bipedalism among South American mammals. J. Mammal. Evol. 27, 419–444 (2020).
Kielan-Jaworowska, Z. & Gambaryan, P. P. Postcranial anatomy and habits of Asian multituberculate mammals. Foss. Strata 36, 1–92 (1994).
Krause, D. W., Hoffmann, S. & Werning, S. First postcranial remains of Multituberculata (Allotheria, Mammalia) from Gondwana. Cretaceous Res. 80, 91–100 (2017).
Chester, S. G. B., Sargis, E. J., Szalay, F. S., Archibald, J. D. & Averianov, A. O. Therian femora from the Late Cretaceous of Uzbekistan. Acta Palaeontol. Pol. 57, 53–64 (2012).
Gambaryan, P. P. & Averianov, A. O. Femur of a morganucodontid mammal from the Middle Jurassic of Central Russia. Acta Palaeontol. Pol. 46, 99–112 (2001).
Martin, T. Postcranial anatomy of Haldanodon exspectatus (Mammalia, Docodonta) from the Late Jurassic (Kimmeridgian) of Portugal and its bearing for mammalian evolution. Zool. J. Linnean Soc. 145, 219–248 (2005).
Luo, Z. X. & Wible, J. R. A Late Jurassic digging mammal and early mammalian diversification. Science 308, 103–107 (2005).
Rose, K. D. Postcranial skeleton of Eocene Leptictidae (Mammalia), and its implications for behavior and relationships. J. Vert. Paleontol. 19, 355–372 (1999).
Shelley, S. L., Williamson, T. E. & Brusatte, S. L. The osteology of Periptychus carinidens: A robust, ungulate-like placental mammal (Mammalia: Periptychidae) from the Paleocene of North America. PLOS ONE 13, e0200132 (2018).
Chinsamy, A. & Hurum, J. H. Bone microstructure and growth patterns of early mammals. Acta Palaeontol. Pol. 51, 325–338 (2006).
Martinelli, A. G. et al. New cladotherian mammal from southern Chile and the evolution of mesungulatid meridiolestidans at the dusk of the Mesozoic era. Sci. Rep. 11, 7594 (2021).
Zhou, C. F., Bhullar, B. A. S., Neander, A. I., Martin, T. & Luo, Z. X. New Jurassic mammaliaform sheds light on early evolution of mammal-like hyoid bones. Science 365, 276–279 (2019).
Grossnickle, D. M., Smith, S. M. & Wilson, G. P. Untangling the multiple ecological radiations of early mammals. Trends Ecol. Evol. 34, 936–949 (2019).
Argot, C. Functional adaptations of the postcranial skeleton of two Miocene borhyaenoids (Mammalia, Metatheria), Borhyaena and Prothylacinus, from South America. Palaeontology 46, 1213–1267 (2003).
Argot, C. Evolution of South American mammalian predators (Borhyaenoidea): Anatomical and palaeobiological implications. Zool. J. Linnean Soc. 140, 487–521 (2004).
Fostowicz-Frelik, Ł. The hind limb skeleton and cursorial adaptations of the Plio-Pleistocene rabbit Hypolagus beremendensis. Acta Palaeontol. Pol. 52, 447–476 (2007).
Gambaryan, P. P., Aristov, A. A., Dixon, J. M. & Zubtsova, G. Y. Peculiarities of the hind limb musculature in monotremes: An anatomical description and functional approach. Russian J. Theriol. 1, 1–36 (2002).
Warburton, N. M., Yakovleff, M. & Malric, A. Anatomical adaptations of the hind limb musculature of tree-kangaroos for arboreal locomotion (Marsupialia: Macropodinae). Aust. J. Zool. 60, 246–258 (2012).
Acknowledgements
The present paper is the result of the joint Argentine-Japanese explorations carried out in March 2020, March 2022, and February 2023. We thank the late Coleman Burke (New York) for his encouragement and financial assistance to carry out the first field explorations to La Anita farm. Dr. Yoshihiro Hayashi, former Director General, National Museum of Nature and Science, Japan, for his support of the project by funding a major part of the expedition from the internal grant of the museum. Special thanks are due to Santiago Miner (LACEV-MACN), who kindly helped with the reconstruction of 3D images of Patagomaia, Kai R.K. Jäger (Section Palaeontology, Institute of Geosciences, Rheinische Friedrich-Wilhelms-Universität Bonn, Germany) for providing 3D models of the Henkelotherium, and David Krause (Denver Museum of Nature and Science, Colorado) for sharing 3D models of Adalatherium. We thank M. Martinelli and M. Ezcurra (MACN-PV), Pablo Teta and Sergio Lucero (MACN-Ma), for allowing access to fossil specimens under their care. The specimen of Patagomaia was collected by G.E. Lo Coco (LACEV-MACN). The authors would like to thank Oscar Canto and Carla Almazán (Secretaría de Cultura) for supporting our projects and explorations in Santa Cruz. Special thanks are also due to Federico Braun for allowing access to his property and F. Echeverría, D. Fraser and A. Prieto for the hospitality and their valuable geographic knowledge of the Anita territories. Authors would also like to thank the entire field crew, including C. Sakata, C. Miyamae, H. Kamei, M.P. Isasi, F. Brissón-Egli, A. Moreno Rodríguez, G. Lio, S. Miner, G. Muñoz, J. De Pasqua, C. Thompson, D. Piazza, G. Lo Coco, A. Misantone, G. Stoll, S. Rozadilla, A.M. Aranciaga Rolando, M.J. Motta, G. Álvarez-Herrera, and R. Álvarez-Nogueira for their assistance and logistics during fieldwork. We especially thank Mirta González and Dr Victoria Sánchez (MACN-Icn) for their assistance in the use of the petrographic microscope. We also are grateful to Roberto Pujana (MACN, BA Pb) for the preparation of the thin-sections. We also thank the geologists D. Moyano-Paz, M. Coronel, D.G. Poiré, M.S. Raigemborn and S. Lizzoli, and paleobotanist E. Vera for their valuable comments on the stratigraphy and regional geology of the Chorrillo Formation. We would like to express our sincere thanks to Benjamin Breenden, for his reading and contributions to the better writing of the manuscript. Thanks also to Elsa Panciroli, Robin Beck, Guillermo Rougier and an anonymous reviewer, for their enlightening comments that greatly improved the quality of the manuscript.
Author information
Authors and Affiliations
Contributions
N.R.C., F.L.A. and F.E.N. conceptualized the study. N.R.C. phylogenetic analyses. J.G.M. paleohistological study. F.E.N., M.M. and T.T. acquired fundings. N.R.C., F.L.A. and J.G.M. made the figures. All authors contributed to the writing and general revision of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chimento, N.R., Agnolín, F.L., García-Marsà, J. et al. A large therian mammal from the Late Cretaceous of South America. Sci Rep 14, 2854 (2024). https://doi.org/10.1038/s41598-024-53156-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-53156-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
