
Abstract
Joint inhumations of adults and children are an intriguing aspect of the shift from collective to single burial rites in third millennium BC Western Eurasia. Here, we revisit two exceptional Beaker period adult–child graves using ancient DNA: Altwies in Luxembourg and Dunstable Downs in Britain. Ancestry modelling and patterns of shared IBD segments between the individuals examined, and contemporary genomes from Central and Northwest Europe, highlight the continental connections of British Beakers. Although simultaneous burials may involve individuals with no social or biological ties, we present evidence that close blood relations played a role in shaping third millennium BC social systems and burial practices, for example a biological mother and her son buried together at Altwies. Extended family, such as a paternal aunt at Dunstable Downs, could also act as ‘substitute parents’ in the grave. Hypotheses are explored to explain such simultaneous inhumations. Whilst intercommunity violence, infectious disease and epidemics may be considered as explanations, they fail to account for both the specific, codified nature of this particular form of inhumation, and its pervasiveness, as evidenced by a representative sample of 131 adult–child graves from 88 sites across Eurasia, all dating to the third and second millennia BC.
Similar content being viewed by others
Introduction
This paper presents new evidence for the genetic relations of individuals from two double graves discovered in Altwies in 20001,2 and in Dunstable Downs in 18873, believed to date from the Beaker period. Three whole genomes at ~ 2X are reported here, as well as target enriched genomic data from one individual. New anthropological analysis and radiocarbon dates are also presented.
Today, the death of a child represents a traumatic event, invoking grief and bereavement. Children's burials are often private, family matters, with community engagement occurring only when tragedy or violence evoke a public response. Past reactions to child death were perhaps more complex and diverse, and in the archaeological record arguably find their clearest expression in surviving traces of funerary rites. Whilst we must be wary of applying modern, Western concepts of parenting and family structure to prehistoric communities, the close bond between child and adult, be they biological parent or social care-giver, appears to find expression through shared burials, perhaps when death overcame both at the same time. At the Mesolithic site of Vedbaek in Denmark, for instance, a newborn was buried with a juvenile woman on a swan’s wing4. Shared burials signal more than emotion; they may also serve, for example, to highlight family status lineage, or the place of children in the descent system.
The biomolecular evidence of close kin relations revealed by this study allows us to explore more fully a form of burial that has long intrigued archaeologists through the specific perspective of biological or family relationships. If the inhabitants can be shown to have been placed in the grave together, as intertwined, fully-fleshed bodies, we need to consider if they died naturally, within hours of each other, or suffered more sinister deaths. Interpretations of these burials as human sacrifices or premature burials were common amongst British antiquarians and archaeologists3,5,6. The German academic tradition postulated that young children rarely survived their mother in prehistoric times7,8,9,10; killing motherless children would have relieved matriarchal societies from the duties of child-care in the absence of marriage institutions8. Even today, shared burials are regarded as a proxy for massacres and ‘raids’11,12 or linked to infectious diseases and even major epidemics, such as the plague13.
Background to the study
The practice of burying adults and children together in later European prehistory is known in German literature as ‘mother–child burials’ (‘Mutter und Kind Bestattungen’10). At the turn of the third millennium BC, societies predominantly buried their dead in single graves14,15, marking a widespread shift from collective to individual inhumation16,17,18,19. For such societies, shared inhumations represent an exception, hence the characterization of adult–child burials as “special” or “deviant burials” in archaeological literature20. Shared burials in which inhumations took place simultaneously must be distinguished from consecutive burials, such as those found in Unterhautzenthal, Austria21, or in Bell Beaker central Iberia, for example Camino del Molino, and La Atalayuela (La Rioja), where large deposits of human bones result from skeletons being pushed aside over time to make space for new inhumations22,23,24, and from intercutting graves, widely found in Beaker cemeteries.
Widespread changes in burial practices have been linked to the advent of pastoral populations from the Pontic-Caspian steppes, or at least their descendants, who admixed in various proportions with European Neolithic farmers and settled as far west as Britain c. 2,500 BC25,26,27,28,29. The discovery of first-generation immigrants to Wessex, such as the ‘Amesbury Archer’, who presumably spent his childhood in Continental Europe, around the western Alps30,31, has helped to rekindle interest in the so-called ‘Beaker folk’32,33,34. The movement of this new population into Northwest Europe has been portrayed as violent and disruptive, notably in popular studies of ancient genetics35,36, whilst monumental landscapes such as Stonehenge appear to have been radically transformed in the wake of their arrival37.
Genetic analysis of two adult–child burials, from Altwies and Dunstable Downs, provides an ideal opportunity to explore the nature of shared burial practices in Beaker-period societies. These graves are incredibly similar in layout, to the extent that the same arrangement of stones can be observed on either side of the skeletons (Fig. 1)2. Were the child and the adult buried simultaneously? Were they biologically related, or was this a social relationship in death? If they were biologically related, what was the degree of relatedness between them? Few Beaker-period graves have produced genetic information about shared burials27. The phenomenon has received more attention in Central Europe, within the context of Late Neolithic to Early Bronze Age societies15,20,38,39,40,41.

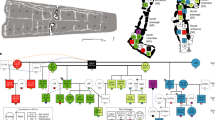
Adult–child graves sampled for this study, Bell Beaker period. (A) Altwies ‘‘Op dem Boesch’’, Luxembourg (photograph: Le Brun-Ricalens et al. 2011: Fig. 7142; Institut National de Recherches Archéologiques); (B) Dunstable Downs, Southern Bedfordshire, United Kingdom (etching: W.G. Smith 18943); (C) inferred pedigrees for the individuals buried in Altwies (left) and Dunstable Downs (right). Key: circle: female; square: male. Colored shapes represent individuals with genetic data and dashed shapes, inferred individuals buried somewhere else.
Results
The adult–child burial of Altwies: archaeological and anthropological context
Two graves of the Bell Beaker culture were discovered in 2000, during rescue excavations at Altwies “Op dem Boesch”, Grand Duchy of Luxembourg, in advance of the construction of Highway A13 near the French border2. They were cut in the slope of a Jurassic plateau, c. 275 m in height, offering panoramic views of the valley of the Moselle river, a tributary of the Rhine43. While the plateau was densely populated in Early Neolithic times, no trace of occupation beside the two graves could be ascribed to the Bell Beaker period (Luxembourg Final Neolithic)44,45. The region between the Saar and the Moselle rivers has yielded over 100 Beaker-period habitation and funerary sites, including those at Sehndorf in Saarland (Germany) and Montenach in Moselle (France), both about 10 km away from Altwies; Beaker graves are usually found isolated or in groups of two to four, more rarely in small necropolises46,47.
New and previously published AMS dates from Altwies, based on human bones and charcoal samples, place the inhumations in the second half of the 3rd millennium BC (Table 1). Grave 1 was probably that of a young adult male, accompanied by a retouched flake, the tip of a stone point and a flint strike-a-light1. This skeleton yielded no DNA due to the poor preservation and incomplete survival of the bones. The woman and child were buried c. 25 m further upslope, in an oval pit, Grave 2, c. 1.85 m in length and 1.45 m in width. Prior to burial, a fire was lit at the bottom of the pit, resulting in a cracked reddish floor and burned walls. The bodies were subsequently deposited alongside artefacts that showed no evidence of burning, including a stone ‘ring’, possibly made of fossilized shell, and a largely complete maritime-style Beaker42.
The skeletons were seemingly placed and posed in the grave simultaneously—the head of the three to four-year-old child resting, for instance, in the right hand of the 25–35-year-old woman2. The woman’s body was placed in a flexed position on the right side, while the child was buried on the left side, the two individuals facing each other in death (Fig. 1A). Some anatomical articulations have been disturbed, suggesting that the grave was not backfilled, but left as an empty cavity, which may have been initially covered by a rigid frame, possibly a timber cap, and a small burial mound2. An examination of the bones failed to determine a cause of death for either individual.
The ‘echinoid burial’ of Dunstable Downs: a 140-year old puzzle
The second burial under examination, the so-called ‘echinoid burial’, recovered from a round barrow on Dunstable Downs, Bedfordshire, is more problematic. In 1887, work to level the damaged barrow revealed human bones. Worthington George Smith (1835–1917), a local antiquary and naturalist, became involved in the retrieval of what he described as the crouched remains of a ‘girl’ and child discovered in a small, shallow grave in the east side of the barrow. Smith subsequently produced a drawing of the grave (Fig. 1B) for his book, Man, the Primeval Savage3.
The soft, fragmentary remains50, were ‘cleaned, dried, and then soaked […] in thin, hot (almost boiling) gelatine’, and dried again (3, 336). Once the bones were hard, they were fitted together with glue dissolved in spirit3,50. After Smith’s death, the remains passed to local naturalist Jannion Steele Elliot, who donated them to the Pritchard Memorial Museum at Bedford Modern School where the burial was displayed in the entrance hall50,51. Today, the remains are held at Luton Cultural Trust store. The use of such antiquarian materials always brings concerns about provenance and reliability, but Smith valued accuracy of representation and was well-respected within the nascent British archaeological community. The history of the remains post-excavation is well documented50.
The barrow, now destroyed, was located on the summit of Dunstable Downs and is designated barrow 8 of a dispersed cemetery of late Neolithic/early Bronze Age date focused on the seven barrows of the Five Knolls barrow cemetery. It lies 0.4 km south-east of Five Knolls52,53. The geology of the Downs is Late Cretaceous chalk, and the plateau is c. 244 m in height, providing panoramic views over the Vale of Aylesbury. The river Ouzel rises close by as a chalk stream. The barrow, originally 3 m high and 14 m in diameter, appears to have been multi-phase; Smith recorded that there was a large central grave, already completely robbed, whilst the grave examined here was one of ‘six or seven’ peripheral, secondary, burials, all located about 0.92 m from the surface. Of these other graves, two were empty, others contained fragmentary human remains, and a third contained a cremation and funerary urn3,52.
The woman was between 18 and 25 years of age; the child was four to eight years old3,52 (supplementary material SI 4). Smith depicted the woman lying on her left side, holding the child, with her head to the north (Fig. 1). Two broken pots were deposited near the woman’s head; a hammerstone, and white quartz pebble near her right hand. Elsewhere in the grave were two other hammerstones, two flint scrapers, and flint flakes. A flint arrowhead was lost during excavation3,54. Numerous fragments of aurochs were also found in the grave55. Dyer identified 45 fragments of undecorated early Bronze Age pottery, five beaker sherds with whipped cord and stabbed decoration, and eight ‘rustic’ beaker sherds from the excavations, but it is unclear whether these are from the grave or barrow52.
Perhaps the most remarkable of all grave finds were fossil echinoids (sea urchins), hence references to the grave as ‘the echinoid burial’. Initially, Smith reported 12 echinoids ‘surrounding the girl’, but ‘on extending the grave’, 91 were reportedly found and a further 200 from the entire barrow3. On balance, it is likely that the fossils came predominantly from the barrow, perhaps occurring naturally, or incorporated in the burial monument as ‘natural’ offerings56. In his drawing of the burial, Smith used the echinoids as a decorative border, which Dyer52 has claimed is unlikely to be accurate. However, such a placement of natural flint nodules and chalk lumps around a body is reported from burials in Yorkshire and Wessex, notably Barrow Clump, Wiltshire, where the crouched inhumation of a child was surrounded by a series of flint nodules (57, 105; 167).
New 14C dates obtained for the adult and child appear to be anomalously young (Table 1), considering the archaeological material associated with the burial, which is Bell Beaker (2500–1800 BC) in appearance and seems to share characteristics with Beaker graves in Wessex and Yorkshire. Garwood58 proposes that large multi-phase mounds, located in dominating landscape locations and containing many different age and gender categories, such as at Dunstable, can be dated to 2150–1800 BC. Given the use of animal-based products for bone preservation, contamination problems were suspected59 and the dates were repeated, at the CEZA Laboratory in Mannheim, using the inner part of the ear bone. Repeating the dating produced dates that are consistent with the adult and child being from the same period, but still about 500 years younger than expected under the assumption that Dunstable Downs and Altwies were contemporaneous (Table 1).
Genetic relationship between the adult and the child
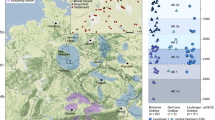
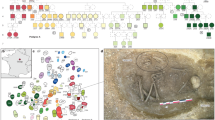
The adult and child were biologically related in both cases. We were able to sequence whole genomes with low contamination estimates (MT < 1%, autosomal < 3%) and establish the degree of relatedness between the adult and child at Altwies and Dunstable Downs using the pairwise mismatch rate and KIN (see method section ‘Genetic Relatedness’). The four individuals are relatively similar on a PCA of modern and ancient Eurasians and show high levels (ALW1 68.2%; ALW2 67.2%; LUT1 59.4%; LUT2 58%) of steppe-related ancestry in a three-population qpAdm model (Fig. 2). The amount of steppe ancestry is consistent with the Beaker expansion in Northwest Europe27,31, a conclusion emphasised by identity by descent (IBD) segments of ≥ 16 cM shared between the four individuals and contemporary genomes from England, the Netherlands, Germany, Bohemia and Poland, ascribed to Bell Beaker and contemporary sites (Fig. 3; supplementary materials, Fig. S2). A particular hotspot emerges in Bohemia, in sites attributed to the Corded Ware, the Bell Beaker and the Únětice cultures27,60. Although intriguing, we caution against overinterpreting these results, without a deeper understanding of the underlying demographic history of these individuals. Furthermore, the possibility of a sampling bias, based on the availability of reference genomes cannot be ruled out. The absence of long runs of homozygosity excludes recent inbreeding. While the two graves look similar archaeologically, we found no evidence at genomic level that the two pairs of individuals were ever in contact. Pigmentation phenotypes for hair, skin, and eyes (SI Table ‘HIrisPlex results’) were produced for all four individuals, and used for the reconstruction in Fig. 4.

(A) PCA of modern and projected ancient West Eurasians and Southwest Asian individuals, highlighting the Altwies and Dunstable Downs individuals; ancient reference individuals are plotted according to the cultural phenomenon or subsistence basis to which they are traditionally assigned (legend in the top right corner); (B) Altwies and Dunstable Downs individuals modelled as mixtures of Western hunter-gatherer (WHG), Aegean–Anatolian farmer, and steppe-related (here represented by Yamnaya) ancestry components with qpAdm.

Location of Altwies “Op dem Boesch” (red dot) and Dunstable Downs (blue dot). Archaeological sites, in which individuals were found to share at least one IBD segment of ≥ 16 cM with our newly reported genomes are shown in the same color. Sharing patterns are further highlighted by a vector line. Relevant third- and second-millennium BC adult–child graves are represented by grey dots. Adult–child graves from the Eurasian steppe belt are not depicted here. For a full list of burials and descriptions, see Table 2 and Supplementary Materials, Fig. S1. Matplotlib Basemap Toolkit 1.3.8 and Python 3.11 were used to produce the map. As map background, a display shaded relief image (from http://www.shadedrelief.com) was used.

The grave of Altwies, left: the bones of the mother and child highlighted (photograph: Le Brun-Ricalens et al. 2011: Fig. 7142; Institut National de Recherches Archéologiques); right: hypothetical reconstruction of the grave based on phenotypic traits partly inferred from the ancient genomes.
Our analyses indicate that the child at Altwies, who was genetically male, was buried alongside his biological mother (Fig. 1). Both individuals shared the same mitochondrial haplogroup, H (Table 1). A difference at two SNPs was observed (12,127 G- > A, 16188C- > T), which is likely the result of post mortem damage and differences in coverage. The boy’s Y haplogroup, R1b, is at high frequency in Bell Beaker males29. The orientation of the woman at Altwies may be regarded as anomalous. In Continental European Beaker communities, women and girls were placed on their right side, with the head at the south, whereas men and boys were buried on their left side, with their head at the north—both sexes thus facing the eastern direction61. Conversely, in Altwies, the woman was placed with her head to the north, facing west. The orientation of the grave thus apparently aligned with the sex of the child.
At Dunstable Downs, both the child and the adult were identified as genetically female. The two individuals were second degree-related on the paternal side (Fig. 1). They belonged respectively to the K1c and N1b mitochondrial haplogroups and therefore had different mothers. Given the pairwise mismatch rates and the age of the adult compared to the child, we can exclude half-sibling and grandmother-granddaughter relationships. The adult is likely to be a paternal aunt. According to Smith, the head of the crouching female child was to the north3. It is not clear if the orientation of the grave was intended for the child or the adult. The orientation of Bell Beaker burials is generally less strict in the south of England, with females lying either on the left side with the head to the north, or on the right side with the head to the east62.
Discussion
A widespread phenomenon in third millennium BC Western Eurasia
In order to shed further light on the burials, we looked for archaeological parallels beyond Altwies and Dunstable Downs. Adult–child burials cut across virtually all major cultural horizons in third millennium BC Western Eurasia, occurring in Bell Beaker, Corded Ware, and Yamnaya cemeteries, as indicated by the representational list of graves in Table 2 and in the Supplementary Materials, and were surprisingly common, even in societies that predominantly practised single inhumation. Child burials of all periods are rare in the archaeological record, not in the least with regard to special burials, grave forms, and artefact associations58. Many children died young and did not receive formal burial. Established archaeological studies of children and childhood emphasise the need to assess children as active cultural and social agents and participants in their societies, in both life and death63.The cultural significance of children in the early Bronze Age is uncertain and the treatment of their physical remains varied greatly58. Double burials containing both adults and children therefore offer us rare glimpses into their contemporary social and cultural importance alongside adults.
Children's graves are generally under-represented in Beaker-period cemeteries from Northern France and Britain64,65,66. Some can be described as mere imitations of adult graves, with miniature vessels, weapons and items of personal adornment57,67. Others were intended for children as well as adults. Shared burials from France, such as those of Achenheim and Léry, consisted of children and babies placed in front or back-to-back with a crouched woman68,69. A three to five-year old child was discovered by the side of an adult man and the cremated remains of another adult in the rectangular pit grave or burial chamber of Hatrize in the Grand Est region of France70. The close relationship between the skeletal remains of the adult and the child implies that both individuals were interred at the same time. Evidence from Early Bronze Age Britain indicates that children, and indeed women, seldom went to the grave alone. Here, multiple burials were the most usual form of burial for both adults and subadults71,72. Between 30 and 40% of child burials from Scotland, Yorkshire, and Wiltshire were buried with an adult, with a bias towards burial with females57,58. At the Beaker cemetery of Craig Tara Holiday Park, Ayrshire, Scotland, for example, every child was buried in a multiple grave containing an adult73.
Looking further afield, shared burials were remarkably frequent in some Corded Ware cemeteries of Central Europe. Up to 44% of graves in two cemeteries from the Tauber Valley, Southern Germany, contained more than one individual11. Intriguingly, shared burials are mainly found outside the area of distribution of regular child graves in Corded Ware Central Europe, suggesting two burial traditions instead of one14. At Eulau, Oechlitz, Lauda-Königshöfen, and Tauberbischofsheim, small clusters of two to five individuals, including adults and children, were buried together in simple pits14,74,75,76. Ancient DNA analyses indicate in this instance close genetic relatedness between some of the adults and children38,39,40,77. The Únětice culture, which succeeds and partly overlaps Corded Ware and Bell Beaker traditions in Central Europe, provides many examples of shared burials, some of which have since produced biomolecular evidence for parent–child inhumations15,20.
The origins of this practice remain unclear. The fact that shared burials in Altwies and Dunstable Downs were associated with individuals with high levels of steppe-related ancestry may indicate the spread of a new burial practice among steppe pastoralists and their descendants, representing innovations in cultural, ritual, and possibly symbolic attitudes and beliefs16,17. Early examples of adult–child burials in pit graves under burial mounds, or kurgans, can be observed in the Yamnaya core area in the Pontic-Caspian steppes78. Double and multiple inhumations of adults and children occur frequently here. The dead are often arranged in theatrical displays of personal affection, such as hugs, kisses, or eye contact, that resemble those observed in Corded Ware and Bell Beaker cemeteries10,14. Furthermore, subadults may have been preferentially buried with an adult in Yamnaya graves78. Looking further East, Central Asian Afanasievo graves, which are linked to the expansion of steppe populations to South Siberia and Mongolia79, featured similar shared burials80.
Transformative attitudes to family identity and social status may have been central to the adoption of these new practices, affecting individual bonds within families, which could have contained both biologically and non-biologically related members, and communities. Increased diversity in burial rites, particularly double and triple burials with infants, and the appearance of child burials with elaborate grave goods suggests that at least in death, children had become increasingly important57,81. Both women and children may have been regarded as of relational importance in death as in life, symbols of fertility or status, or responsible for creating and cementing transgenerational links between and within communities, even linking past and present71. This link may also explain the frequent occurrence of the disarticulated remains of children found buried with adults in the early Bronze Age. This practice has been interpreted by Garwood58 as evidence of children as ‘grave goods’, but equally, the remains of children could have been stored until the death of a socially or ritually appropriate family or community member permitted burial.
Biological and substitute parents
Our study provides the first genetic evidence that Bell Beaker communities in Northwest Europe buried children with their biological mothers and other close biological relatives, apparently in accordance to a specific rite practised throughout the wider ‘Beaker’ world, and indeed beyond. Conversely, Olalde et al. found no close genetic relationship between co-buried individuals at the Bell Beaker necropolis of Camino de las Yeseras, in San Fernando de Henares, Spain27. A young adult woman, who had a typical ancestry of the Iberian Peninsula, was buried atop the skeleton of an infant girl in an artificial cave. The infant was neither her daughter nor a close biological kin, but had a comparatively high amount of steppe ancestry27,82,83. We may speculate that kinship practices were different in the southern domain of the Bell Beaker culture, where collective burial traditions in megalithic tombs and artificial caves continued uninterrupted23. Alternatively, close ties of kinship, and perhaps circumstances of death, needed to be articulated through a specific performative ritual language of side-by-side body posture and rite.
While genetic evidence remains scarce, a variety of kinship patterns are reflected in third millennium BC shared burials. That the adult and child at Dunstable Downs were second-degree related on the paternal side might hint at a patrilineal descent system, in which the paternal aunt could play the role of substitute parent or primary care-giver. At Altwies, the grave’s orientation matched the sex of the boy, not that of his biological mother. Central European Corded Ware graves have previously yielded evidence for ‘nuclear’ families—referring here to biological parents and their offspring38. At Eulau, a single pit grave included the remains of four intertwined skeletons, placed in pairs facing each other; the father and one son were placed with the head to the West, the mother and another son with the head to the North38. The orientation of the children’s skeletons deviated from the Central European Corded Ware tradition, emphasizing the relationship with their parents instead of their biological sex38.
Some of the children in Corded Ware graves were buried with adults who were not their biological parents, making it hard to generalize observations; not all kinship relationships would have been biological. At Eulau, two children, who were probably siblings, were placed by the side of a woman, who had a different mitochondrial haplogroup38. She may have been a paternally-related aunt, a stepmother, or a completely unrelated individual; the exchange of foster children as part of kinship systems, or social and political networks has been proposed84. The Althausen grave in Baden-Württemberg belonged to a ‘patchwork’ or blended family. It consisted of a man and a woman facing each other, with two children placed between them, who were not related to each other40. Burial with a family member or care-giver may have been the preferred rite for children, but in their absence, perhaps any adult may have served, the creation of proxy families or relationships perhaps seen as necessary for the safe passage of the child through death.
The interpretation of parent–child burials
While the causes of death could not be established for the Altwies and Dunstable Downs individuals, the sheer number of actual and potential parent–child burials identified in the third millennium BC (Table 2) raises further questions about the interpretation of this burial form. Did the buried individuals die naturally or were they killed? Birth accidents can be ruled out in all but a few examples, such as the grave of an adult woman and a neonate at the Corded Ware site of Stetten a. d. Donau in Germany85. Natural child deaths should not be discounted; 40% of children in prehistoric societies may have died before the age of five57. However, the simultaneous inhumation of children and adults, who appear to be close relatives or care-givers, is more difficult to explain. Epidemics, for instance Yersinia pestis, common across Eurasia during the Bronze Age86, could theoretically explain the death of multiple individuals at close intervals, but data remain lacking for third millennium BC shared burials.
Interpretations of the mother–child bond in shared burials might also evoke suspicions of violence, sacrifice, or even honour killings. That women and children were frequently the victims of violence in late Neolithic/Early Bronze Age Europe is undisputed. At Eulau, Germany, one of the adult women buried with a child suffered an arrow wound, described as ‘unambiguously fatal’11. The killing of children, for instance by blunt force to the skull, has been documented at several Early Bronze Age sites in Europe, including Schleinbach in Lower Austria, Cezavy Hill in the Czech Republic, and Nord-Trøndelag in Norway87,88,89,90.
Smith3 believed that the child at Dunstable Downs had been ‘buried alive with the mother’. Such willingness to accept the idea of human sacrifice was widespread amongst early prehistorians and indeed, can still be found in literature today. In the process of forming ideas for their new discipline, early prehistorians relied heavily both on Classical writers, such as Herodotus, who described the alleged strangling of retinue members and servants in Scythian barrows (Herodotus, Histories IV, c. 71), and contemporary ethnography, leading them to conceptualise the early Bronze Age peoples of Europe as ‘savages’ or barbarians and adopt sinister or violent explanations. Today, such notions seem fantastical. However, we should not discount the possibility that ritual killing and/or human sacrifice was practised in later prehistoric Europe91.
We should take care not to perceive all adult–child burials as the same. Non-kinship patterns such as relative age, sex, manner of death and social standing, could be just as important as kinship in determining how to handle the death of community members. The burials examined in this article were socially significant events and did not lack care or elaboration; they were furnished as regular graves with beakers and status items. Preparation for the inhumation of the mother and son at Altwies likely spanned two days; the burial pit was cleansed by fire prior to the deposition of the bodies and the artefacts2. Given such care, ‘ad hoc’ interpretations, such as burial following enemy raids, are problematic. Do all shared burials represent the aftermath of raids, when joint burial was offered to victims by survivors? At Eulau, five of the 13 skeletons placed in the four shared burials displayed perimortem injuries, including defence wounds11. These explanations, evoking actual crime scenes, fail to reflect the repetitive and codified nature of shared burial practices in third millennium BC Western Eurasia.
For Garwood58 funerary displays described social and political biographies in individualised if idealised and aesthetically consistent ways. Perhaps the key to understanding the specific examples of ‘parent–child’ burials described here lies not in their potential cause of death, but in the specific, nuanced configuration of the bodies in the grave; two individuals buried in each other’s arms (Fig. 4). Turek92 has observed that one of the most highly symbolic elements within Corded Ware burial rites is the position of the buried person’s arms. This was not related to distinctions between sex and age groups, nor did it appear to relate to grave goods. This positioning continued into the Beaker period and may relate to a social category/identity as yet unclear. The body of a woman, lying as though sleeping, clasping a child in her arms, obviously had a specific meaning to early Bronze Age peoples, a meaning retained across thousands of miles and amongst many diverse and fluid contemporary funerary practices. Whatever it was, it represented something powerful and emotive.
Material and methods
Sample preparation
All laboratory analyses were carried out at the dedicated ancient DNA facilities of the Palaeogenetics Group, Institute of Organismic and Molecular Evolution (iomE) at the Johannes Gutenberg University, Mainz, according to strict ancient DNA protocols to prevent contamination with modern DNA as well as cross-contamination between samples94,95,96, including decontamination of workspace, lab-ware and samples, and processing of negative controls during all steps (sample pulverisation, DNA extraction, library preparation and PCR reactions). All the steps before PCR amplification were conducted in the dedicated ancient DNA facility physically separate from post-PCR areas.
One petrous bone, part of the temporal bone, was collected for each of the four individuals. The dense core of the petrous bone contains the highest amount of endogenous DNA, and is therefore the optimal sampling area of ancient skeletons97.
The petrous bones were decontaminated under ultraviolet light (254 nm) from 2 sides for 45 min per side, then cleaned with a sandblasting machine (P-G 400, Harnisch + Rieth, Winterbach, Germany) to remove the outer bone surface and soil. The densest, non-porous, inner part of the petrous bone was then isolated and cut in small cubes using a disk saw (Marathon N7, SMT), which were irradiated again with ultraviolet light (254 nm) from two sides for 45 min per side and pulverized using a milling machine (MM200, Retsch). Blank milling controls containing hydroxyapatite were processed in parallel with the samples to control contamination.
DNA extraction
The modified Yang et al. (1998) protocol98,99,100, with some additional modifications101,102 described below, was followed for DNA extraction.
The bone powder, 0.10–0.15 g per sample, underwent a pre-lysis by being incubated with 1 ml of EDTA (0.5 M, pH 8) at room temperature for 10 min prior to extraction. The supernatant was discarded after the solution was centrifuged at 10,000 rpm to pellet the powder.
Using 1.8 ml of an extraction buffer made of EDTA (950 µl, 0.5 M, pH8), Tris–HCl (20 µl, 1 M, pH8), N-Lauroylsarcosine (17 µl, 5%), and Proteinase K (13 µl; 20 mg/ml), lysis was carried out on rocking shakers (1400 rpm) at 37 °C for 48 h.
Following the 48-h incubation period, the samples were centrifuged for 10 min at 10,000 rpm to separate the supernatant, which was then transferred to an Amicon Filter (Amicon Ultra-4 30 kDA, 15 ml) and centrifuged for 10 min at 2500 rpm. The DNA was then washed twice with 3 ml 1X Tris–EDTA followed by centrifugation at 2500 rpm for 20 min and removing the flow-through in between. After washing, the extract was concentrated to 100 μl and then purified using the MinElute PCR Purification Kit as directed by the manufacturer, but incubating for 5 min during the elution with 44 μl elution buffer (preheated to 65 °C).
During DNA extraction, blank controls were processed and used in all subsequent analysis stages.
Library preparation
Double-indexed Illumina libraries were created according to the protocol by Kircher et al. (2012)103 with slight modifications. The damage patterns of the DNA fragments were used to show that there was no modern DNA contamination. For whole genome sequencing the extracts were treated with USER™ enzyme prior to library preparation: 5 μl of USER™ enzyme was added to 16.25 μl of DNA extract and the mixture was incubated for 30 min at 37 °C.
The blunt-end repair was performed using the NEBNext End Repair Module (New England Biolabs, Ipswich, Massachusetts, United States): 20 µl of DNA extract (or 16.25 µl + 5 µl of USER™ for non-screening libraries) were mixed with NEBNext End Repair Reaction Buffer (10X, 7 µl), NEBNext End Repair Enzyme Mix (3.5 µl) and nuclease-free water (39.5 µl for screening or 38.25 µl for non-screening libraries; for a final reaction volume of 70 µl) and incubated for 15 min at 25 °C followed by 5 min at 12 °C.
In the adapter ligation step, hybridized adapters P5 and P7 (IDT, Leuven, Belgium)104 were used at a concentration of 0.75 µM.
To add unique and sample-specific index pairs to the library molecules, 3 µl of the fill-in product (total volume: 40 µl) were amplified using the AccuPrime™ Pfx SuperMix (20 µl) in one PCR parallel (final reaction volume: 25 µl; final primer concentration: 200 nM). Double indexing was done following Kircher et al. (2012)103, but with index sequences from the NexteraXT index Kit v2 (Illumina; barcode length 8 bp).
The PCR was carried out in 10–14 cycles; the temperature profile used for the PCR complied with the manufacturer's guidelines, but used an annealing temperature of 60 °C, extending for 30 s during each cycle, and carrying out a final elongation step for 5 min.
Purification during library preparation was conducted using the MinElute PCR Purification Kit (Qiagen, Hilden, Germany), while amplified libraries were purified with the MSB® Spin PCRapace kit (Invitek, Stratec Molecular, Berlin, Germany). Library concentrations were quantified by Qubit Fluorometric quantitation (dsDNA HS assay) and measurement on the Agilent 2100 Bioanalyzer System (High Sensitivity DNA Analysis) was used to estimate fragment length distributions of the libraries.
To confirm the success of the library preparation and to monitor contamination, blank controls, as well as positive controls (nonsense hybrids) of known concentration, were processed during every library step, including PCR amplification. A quantitative analysis of the blank controls revealed no significant contamination in any laboratory stage (pulverisation, DNA extraction, library preparation and amplification). A summary of all labwork and sequencing is reported in SI Table.
Whole-genome sequencing
All four samples were firstly screened for their endogenous DNA preservation using shallow shotgun sequencing. Screening runs were performed on an Illumina MiSeq™ platform at StarSEQ GmbH (Mainz, Germany) in single-end runs with 50 bp read length. After adapter trimming and quality check103, reads were aligned to the human reference genome GRCh37/hg19 using BWA aln105, duplicates were removed and reads were filtered for a minimum length of 30 bp using the MarkDuplicates function from the Picard tools package (picardtools, http://broadinstitute.github.io/picard). The percentage of endogenous DNA was calculated as the ratio of unique aligned reads to the reference genome against the total number of reads after quality filtering. One sample had very low endogenous content (ALW1) at 1.4%, the other three between 20 and 50% (SI Table ‘General Stats’). Post-mortem damage patterns in aligned, length-filtered sequence reads were obtained with the software package MapDamage 2.0106 for all the libraries to evaluate sample authenticity.
For deeper whole genome sequencing, 2–3 DNA extracts and 3–9 libraries of each sample were prepared (as already described above). The libraries were amplified in up to 12 PCR parallels to increase the complexity of the libraries.
The number of cycles for the PCR reactions varied between samples (10–14 cycles) as it was adjusted to the presumed quantity of DNA fragments in the library. After amplification, all parallels of the same library were purified together and quantified as described above. For sequencing, libraries were pooled according to their concentrations measured on Qubit® and taking into account the endogenous DNA content as estimated by MiSeq sequencing and subsequently purified with Agencourt® AMPure® XP beads.
The samples were sequenced on an Illumina NovaSeq6000 (SE, 100 cycles) at the Next Generation Sequencing Platform at the University of Berne, Switzerland.
Target enrichment
Screening showed that one sample (ALW1) had very low endogenous content. This sample was target enriched using the Twist Bioscience ancient DNA Target Enrichment reagent, following the protocol of Rohland et al. (2022)107 with the modifications described below.
Two independent capture reactions were performed for sample ALW1, the first with two independent double-indexed libraries originating from the same DNA extract, and the second using additional libraries from a second DNA extraction resulting in a total of four libraries (see SI Table for details).
The double-indexed libraries were pooled and concentrated to obtain 1500 ng (instead of 1 library of 1000 ng) that are then reconstituted in 7 µl of universal blockers and 5 µl blocker solution. Following the indication of Rohland et al. (2002)107, we mixed 1 µl of Twist Ancient DNA probes (Twist custom probe panel number: TE-94002772, Part Number: 104298) and 0.167 µl of the MT probes (Part Number: 102040 with 5 µl of hybridization mix. The probes were heated for 2 min at 95 °C and subsequently cooled for 5 min at 4 °C. After heating libraries and blockers for 5 min at 95 °C, both solutions were left at room temperature for 5 min. The probes mix (6.167 µl) was added to the 12 µl library and blocker, mixed, overlaid with 30 µl hybridization enhancer and incubated at 62 °C in a thermal cycler for 16 h. We used 300 µl streptavidin beads and bound the targets for 30 min at room temperature on a shaker (600 rpm). The beads were then washed four times with two different wash buffers; three were stringent washes at 49 °C. Only 50% of the bead slurry (the rest was frozen and stored) was then amplified with 25 µl Equinox Library Amp Mix in 12 cycles with 2.5 µl of the provided primers (ILMN). The amplified capture products were then purified with 1.8 × DNA purification beads and eluted in 32 µl Elution buffer.
The two capture reactions were then sequenced on an Illumina NovaSeq6000 (SE, 100 cycles) at the Next Generation Sequencing Platform at the University of Berne, Switzerland, and afterwards, the data were merged.
Bioinformatics pipeline
Residual adapters were removed with trimmomatic 0.36108 and sequences shorter than 30 bp were discarded prior to aligning the reads to the human reference genome (GRCh37/hg19) with bwa mem109. During conversion to BAM format using samtools110 only reads with a mapping quality ≥ 30 (-q 30) were kept. PCR duplicates were removed from the alignment with sambamba markdup111 and the remaining reads were realigned around known SNPs and InDels using GATK 3.6112. Potential contamination was assessed based on reads aligning to the mitochondrial genome using contamMix 1.0.9113 and for autosomal reads using ContamLD114, together with ANGSD115 for X-chromosomal reads in ALW2. Since the ANGSD estimates suggested contamination levels below 1%, we used the ContamLD estimates for ALW2 for correcting the estimates for the remaining genomes. Genetic sexing of each individual was done following Skoglund et al. (2013)116.
Variant call and phasing/imputation
Variant detection was done with ATLAS117. After estimating deamination patterns for sequenced libraries separately, the sequencing error was estimated jointly based on a set of sites, known to be ultra-conserved among multiple mammalian species, following the description at the bitbucket repository:
https://bitbucket.org/wegmannlab/atlas/wiki/Home.
The “majorityBase” function was used for sites overlapping the 1240 K capture regions described in Mathieson et al. (2015)118 and the mitochondrial genome to generate pseudo-haploid genotype calls. Diploid calls for a set of sites determined to be bi-allelic and present in at least two individuals in the 1000 Genomes data-set119 were produced with the “MLE” call-function. To further minimise potential artefacts introduced by PMD, we ignored the first and last 2 bp of each read during calling.
Phasing/imputation of the genomes was performed by running glimpse 1120 following the provided instructions and using default parameters with the 1000 genomes data as reference119.
Uniparental markers
MtDNA- and Y-chromosomal haplotypes were determined with HaploGrep3121, and Y-leaf122 with the ISOGG 2018 tree (International Society of Genetic Genealogy) respectively.
Patterns of variation
PCA, qpAdm
Explorative genomic similarity analysis was performed by PCA using LASER v.2.04123 following Hofmanová et al.102 projecting BAM files onto a reference space of modern Eurasian populations124. Modern reference populations used: Southern European (Italian North/South, Sicilian, Spanish/- North, Canary Islander, Maltese, Greek), Basque, Sardinian, Cypriot, Central European (Albanian, Bulgarian, Romanian, Hungarian, Croatian, Czech, German, French), Eastern European (Russian, Ukrainian, Belarussian, Polish, Sorb), Mordovian, Eastern Baltic and Finnish (Estonian, Lithuanian, Finnish), British Isles (English, Orcadian, Scottish, Irish/-Ulster, Shetlander), Scandinavian (Icelandic, Norwegian), Caucasian (Georgian, North Ossetian, Abkhasian, Chechen, Adygei, Lezgin, Kumyk, Balkar), West Asian (Turkish, Armenian), Iranian/-Bandari, and Near Eastern (Palestinian, Druze, Jordanian). Ancient reference individuals are given in SI Table ‘PCA references’ (annotation from the Allen Ancient DNA Resource125).
f4 admixture proportions were computed with qpAdm from ADMIXTOOLS126 using default parameters and on pseudohaploid calls at the positions overlapping with the human origin/Illumina capture. Outgroups for qpAdm were individuals with labels Han.SDG, Karitiana.SDG, Mbuti.SDG, and Papuan.SDG127 and ancient genomes Russia_MA1_HG.SG, Ethiopia_4500BP_published.SG, Belgium_UP_GoyetQ116_1_published, Russia_Ust_Ishim.DG, Spain_ElMiron, all retrieved from the Allen Ancient DNA Resource v50.0125.
IBD and ROH analyses
Screening for runs of homozygosity was done for each individual using hapROH128. Shared haploid IBD segments between our newly reported genomes and a set of previously published individuals (for a detailed list see Supplementary Table 7) were determined with ancIBD129 focussing on sites in the 1240 K capture array118.
Phenotypic inference
The HIrisPlex-S webtool130 was used to predict pigmentation phenotypes for hair, skin, and eyes for each sample. Results were used for the reconstruction in Fig. 4.
Genetic relatedness
Genetic relatedness was assessed by computing pairwise-relatedness coefficient r for autosomal and X-chromosomal SNPs following Fowler et al. (2022)131 with a set of contemporary genomes from previously published studies (see SI Table for further details) in addition to running KIN132. Directionality of the inferred relatedness was resolved by comparing autosomal and X-chromosomal r-values, in addition to the assessment of IBD patterns estimated by KIN.
Anthropological analyses
Detailed anthropological analysis of the Beaker-period skeletons recovered in Altwies took place in 2000 and is reported in Toussaint et al.2. Anthropological examination of the two individuals from Dunstable downs was carried out to detect the presence of any pathological or traumatic lesions that could have led to their death. The age at death was estimated for the subadult through the analysis of dental eruption and the development of various skeletal parts and the degree of fusion of ossification centres133,134. The adults’ age-at-death was estimated using the degenerative changes due to ageing in the skeleton, in particular dental wear and ectocranial suture obliteration135,136,137. Paleopathological examination and trauma detection was carried out through macroscopical examination of the skeletons. The results of the analysis are reported in the Supplementary Materials.
Radiocarbon dating
New radiocarbon dates were obtained for three of the individuals sampled (ALW2, LUT1, LUT2). In addition, a distal phalanx of the left thumb of the adult at Altwies (ALW1) was dated in 2000 to 2198–1947 calBC at 2σ (Beta-145714: 3680 ± 40 BP1). For new samples, the analyses were conducted at the Curt-Engelhorn-Zentrum Archäometrie gGmbH (Mannheim, Germany). The collagen was extracted from fragments of the petrous bones used for aDNA analysis. All dates were calibrated in OxCal v4.4.4, using the IntCal20 calibration curve48,49. The date obtained for the child at Altwies, 2135–1950 calBC (ALW2: MAMS-56230: 3656 ± 21 BP) is consistent with the date of the mother (ALW1: R_Combine value of 2135–1955 calBC at 2σ, X2-Test: df = 1 T = 0.3(5% 3.8)).
For Dunstable Downs, the adult and child initially produced dates that were internally inconsistent at 2σ, despite the second-degree kinship relationship between them. The dates were respectively 1666–1500 calBC (LUT1: MAMS-58671: 3299 ± 32 BP) and 1386–1201 calBC (LUT2: MAMS-58672: 3021 ± 21 BP) (Table 1). Contamination issues were suspected, due to the use by Smith of gelatin and ‘shellac dissolved in spirit’ to glue together the bones (55, 319). Shellac is a bioadhesive polymer, a solution of melted lac, the resinous excretion of the Lac insect (Coccus or Carteria lacca), formerly used extensively in archaeological conservation138. Re-dating of the samples in Mannheim, removing the bone surface before pretreatment, appears to have eliminated the age discrepancy between the two individuals. The adult and child are now respectively dated to 1613–1453 calBC (LUT1: MAMS-61441: 3263 ± 27 BP) and 1502–1417 calBC (LUT2: MAMS-61442: 3182 ± 26 BP) (Table 1). It is unclear if the gelatin only contaminated the surface of bones or also soaked into the material, but in any case the treatment could have produced artificially younger dates59. The dates should therefore be treated with caution. In addition, MAMS-61441 shows a very low collagen preservation (0.4%).
Data availability
Sequencing data are available in FASTQ and BAM format at the European Nucleotide Archive under the accession number PRJEB65118. VCFs and raw outputs from ancIBD used to make Fig. 3 are available at: https://irods-web.zdv.uni-mainz.de/irods-rest/rest/fileContents/zdv/project/palaeogenetik/IBDres.tar.gz?ticket=0O2nNMMnBbyN4KL.
References
Le Brun-Ricalens, F., Hauzeur, A., Toussaint, M., Jost, C. & Valotteau, F. Les deux sépultures campaniformes d’Altwies, Op dem Boesch (Grand-Duché de Luxembourg): matériel archéologique et contexte régional. Bull. Soc. Préhist. Luxembourgeoise 23, 285–300 (2001).
Toussaint, M., Le Brun-Ricalens, F. & Hauzeur, A. Les deux sépultures campaniformes d’Altwies, Op dem Boesch (Grand-Duché de Luxembourg): méthodologie, données anthropologiques préliminaires et essai de caractérisation des pratiques sépulcrales. Bulletin de la Société préhistorique luxembourgeoise 23–24, 249–284 (2001).
Smith, W. G. Man, the Primeval Savage: His Haunts and Relics from the Hill-tops of Bedfordshire to Blackwall. (E. Stanford, 1894).
Stutz, L. N. The Importance of ‘Getting It Right:’ Tracing Anxiety in Mesolithic Burial Rituals. in The Archaeology of Anxiety: The Materiality of Anxiousness, Worry, and Fear (eds. Fleisher, J. & Norman, N.) 21–40 (Springer New York, 2016).
Greenwell, W. British Barrows: A Record of the Examination of Sepulchral Mounds in Various Parts of England. (Clarendon Press, 1877).
Burgess, C. The age of Stonehenge. (Dent, 1980).
Wilke, G. Mutter und Kind. Ein Beitrag zur Frage des Mutterrechts. Mannus 21, 26–51(1929).
Wilke, G. Nochmals Mutter und Kind. Mannus 22, 358–360 (1930).
Fischer, U. Die Gräber der Steinzeit im Saalegebiet. (Walter de Gruyter & Co., 1956).
Häusler, A. Zum Verhältnis von Männern, Frauen und Kindern in Gräbern der Steinzeit. Arbeits und Forschungsberichte zur Sächsischen Bodendenkmalpflege 14(15), 25–73 (1966).
Meyer, C. et al. The Eulau eulogy: Bioarchaeological interpretation of lethal violence in Corded Ware multiple burials from Saxony-Anhalt, Germany. J. Anthropol. Archaeol. 28, 412–423 (2009).
Schroeder, H. et al. Unraveling ancestry, kinship, and violence in a Late Neolithic mass grave. Proc. Natl. Acad. Sci. U. S. A. 116, 10705–10710 (2019).
Swali, P. et al. Yersinia pestis genomes reveal plague in Britain 4,000 years ago. Nat. Commun. 14, 2930 (2023).
Seregély, T. Kleine Bekannte auf weiter Flur? Ein Überblick zu Kindern in der Kultur mit Schnurkeramik. in Lebenswelten von Kindern und Frauen in der Vormoderne : Archäologische und anthropologische Forschungen in memoriam Brigitte Lohrke 299–315 (curach bhán publications, 2015).
Rebay-Salisbury, K. 3. Personal Relationships between Co-buried Individuals in the Central European Early Bronze Age. AmS-Skrifter 26, 35–48 (2018).
Harrison, R. & Heyd, V. The transformation of Europe in the third millennium BC: The example of ‘Le Petit-chasseur I + III’ (Sion, Valais, Switzerland). Praehist. Z. 82, 129–214 (2007).
Heyd, V. Families, prestige goods, warriors & complex societies: beaker groups of the 3rd millennium cal BC along the upper & middle danube. Proc. Prehist. Soc. 73, 327–379 (2007).
Salanova, L. Les sépultures campaniformes: lecture sociale. in Le Chalcolithique et la construction des inégalités. 1. Le continent européen (ed. Guilaine, J.) 213–228 (Errance, 2007).
Furholt, M. Mobility and social change: understanding the european neolithic period after the archaeogenetic revolution. J. Archaeol. Res. 29, 481–535 (2021).
Knipper, C. et al. A distinct section of the Early Bronze Age society? Stable isotope investigations of burials in settlement pits and multiple inhumations of the Únětice culture in central Germany. Am. J. Phys. Anthropol. 159, 496–516 (2016).
Rebay-Salisbury, K. Bronze Age Beginnings: The Conceptualization of Motherhood in Prehistoric Europe. in Motherhood in Antiquity (eds. Cooper, D. & Phelan, C.) 169–196 (Springer International Publishing, 2017).
Fernández-Crespo, T., Ordoño, J., Barandiarán, I., Andrés, M. T. & Schulting, R. J. The Bell Beaker multiple burial pit of La Atalayuela (La Rioja, Spain): stable isotope insights into diet, identity and mortuary practices in Chalcolithic Iberia. Archaeol. Anthropol. Sci. 11, 3733–3749 (2019).
Liesau von Lettow-Vorbeck, C., Ríos, P. & Blasco, C. Bell Beakers in central Iberia: keeping the ancestors’ memory alive. Préhistoires méditerr. (2020) https://doi.org/10.4000/pm.2206.
Díaz-Navarro, S., Haber Uriarte, M., Tejedor-Rodríguez, C. & Lomba Maurandi, J. Emphasising the community: demographic composition of an exceptional tomb—the Chalcolithic burial site of Camino del Molino, Caravaca de la Cruz. Murcia. Archaeol. Anthropol. Sci. 15, 140 (2023).
Allentoft, M. E. et al. Population genomics of Bronze Age Eurasia. Nature 522, 167–172 (2015).
Haak, W. et al. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 522, 207–211 (2015).
Olalde, I. et al. The Beaker phenomenon and the genomic transformation of northwest Europe. Nature 555, 190–196 (2018).
Olalde, I. et al. The genomic history of the Iberian Peninsula over the past 8000 years. Science 363, 1230–1234 (2019).
Haak, W. et al. 5 The Corded Ware Complex in Europe in Light of Current Archaeogenetic and Environmental Evidence. in The Indo-European Puzzle Revisited: Integrating Archaeology, Genetics, and Linguistics (eds. Kristiansen, K., Kroonen, G. & Willerslev, E.) 63–80 (Cambridge University Press, 2023).
Evans, J. A., Chenery, C. A. & Montgomery, J. A summary of strontium and oxygen isotope variation in archaeological human tooth enamel excavated from Britain. J. Anal. At. Spectrom. 27, 754–764 (2012).
Patterson, N. et al. Large-scale migration into Britain during the Middle to Late Bronze Age. Nature 601, 588–594 (2022).
Fitzpatrick, A. P. The Amesbury Archer and the Boscombe Bowmen: Early Bell Beaker burials at Boscombe Down, Amesbury, Wiltshire, Great Britain: Excavations at Boscombe Down, volume 1. (Wessex Archaeology, 2013).
Pearson, M. P. et al. The Beaker people: isotopes, mobility and diet in prehistoric Britain. vol. 7 (Oxbow, 2019).
Armit, I. & Reich, D. The return of the Beaker folk? Rethinking migration and population change in British prehistory. Antiquity 95, 1464–1477 (2021).
Lemercier, O. Le guerrier dans l’Europe du 3e millénaire avant notre ère. L'arc et le poignard dans les sépultures individuelles campaniformes. in L’armement et l'image du guerrier dans les sociétés anciennes: de l'objet à la tombe (eds. Baray, L., Honegger, M. & Dias-Merinho, M.-H.) 121–165 (Ed. universitaires, 2009).
Reich, D. Who We Are and How We Got Here: Ancient DNA and the new science of the human past. (Oxford University Press, 2018).
Parker Pearson, M. Stonehenge: Exploring the Greatest Stone Age Mystery. (Simon and Schuster, 2012).
Haak, W. et al. Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age. Proc. Natl. Acad. Sci. U. S. A. 105, 18226–18231 (2008).
Meyer, C., Ganslmeier, R., Dresely, V. & Alt, K. W. New approaches to the reconstruction of kinship and social structure based on bioarchaeological analysis of Neolithic multiple and collective graves. in Theoretical and Methodological Considerations in Central European Neolithic Archaeology (eds. Kolář, J. & Trampota, F.) 11–23 (Bar International Series vol. 2325, 2012).
Mittnik, A. et al. Kinship-based social inequality in Bronze Age Europe. Science 366, 731–734 (2019).
Linderholm, A. et al. Corded Ware cultural complexity uncovered using genomic and isotopic analysis from south-eastern Poland. Sci. Rep. 10, 6885 (2020).
Le Brun-Ricalens, F., Toussaint, M. & Valotteau, F. Les sépultures campaniformes d’Altwies-‘Op Dem Boesch’ (Grand-Duché de Luxembourg). in Les sépultures individuelles campaniformes en France (eds. Salanova, L. & Tchérémissinoff, Y.) vol. XLIe supplément 115–123 (CNRS Editions, 2011).
Jost, C., Le Brun-Ricalens, F., Hauzeur, A. & Ziesaire, P. Altwies, Op dem Boesch (Grand-Duché de Luxembourg): contexte géographique et historique des recherches. Bulletin de la Société Préhistorique Luxembourgeoise 23, 153–161 (2001).
Jost, C., Hauzeur, A., Toussaint, M. & Le Brun-Ricalens, F. Altwies ‘Op dem Boesch’, Grand-Duché de Luxembourg. Occupation du Rubané final et du Campaniforme en bordure de plateau. Bulletin de la Société Préhistorique Française 98, 551–554 (2001).
Hauzeur, A. & Jost, C. Une occupation rubanée particulière à Altwies, Op dem Boesch (Grand-Duché de Luxembourg). Bulletin de la Société préhistorique luxembourgeoise 23–24, 209–239 (2001).
Fritsch, T. Gräber der Glockenbecherkultur von Sehndorf. in Archäologischen Untersuchungen im Trassenverlauf der Bundes-autobahn A8 im Landkreis Merzig-Wadern (ed. Miron, A.) 281–290 (Landesdenkmalamt im Ministerium für Umwelt, 2000).
Blouet, V. et al. Le troisième millénaire entre la Sarre et la Meuse française. in Préhistoire de l’Europe du Nord-Ouest: mobilité, climats et identités culturelles. 28e congrès préhistorique de France. Vol. 3. 321–344 (Société Préhistorique Française, 2016).
Reimer, P. J. et al. The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Bronk Ramsey, C. OxCal v4.4.4. Retrieved from https://c14.arch.ox.ac.uk (2021).
Dyer, J. F. Worthington George Smith. in Worthington George Smith and Other Case Studies (ed. Bell, P.) 141–176 (Bedfordshire Historical Record Society, 1978).
Leeming, P. 'Also found…(not illustrated)…’: the curious case of the missing magical fossils. in The Materiality of Magic. An artifactual investigation into ritual practices and popular beliefs (eds. Houlbrook, C. & Armitage, N.) 15–22 (Oxbow Books, 2015).
Dyer, J. The five knolls and associated barrows at dunstable. Bedfordshire. Bedfordshire Archaeological Journal 19, 25–29 (1991).
Kaye, D. Topographical Survey and Observation Recording Report: Five Knolls Dunstable Downs Bedfordshire. (KDK Archaeology, 2013).
Smith, W. G. Early Man. in The Victoria History of the County of Bedford. Volume 1 (ed. Page, W.) 145–174 (A. Constable, 1904).
Smith, W. G. Skeleton of brachycephalic celt. Nature 43, 319–320 (1891).
Brück, J. & Jones, A. M. Finding objects, making persons: Fossils in British Early Bronze Age burials. in Relational identities and other-than-human agency in archaeology (eds. Harrison-Buck, E. & Hendon, J. A.) 237–262 (University of Colorado Press, 2018).
McLaren, D. P. Funerary rites afforded to children in earlier Bronze Age Britain: case studies from Scotland, Yorkshire and Wessex. (The University of Edinburgh, 2011).
Garwood, P. Vital resources, ideal images and virtual lives: children in Early Bronze Age funerary ritual. in Children, Childhood and Society (eds. Crawford, S. & Shepherd, G.) 63–82 (Bar Publishing, 2007).
Prüfer, K. et al. A genome sequence from a modern human skull over 45,000 years old from Zlatý kůň in Czechia. Nat. Ecol. Evol. 5, 820–825 (2021).
Papac, L. et al. Dynamic changes in genomic and social structures in third millennium BCE central Europe. Sci. Adv. 7, (2021).
Gaydarska, B. et al. To Gender or not To Gender? Exploring Gender Variations through Time and Space. Eur. J. Archaeol. 1–28 (2023).
Heise, M. E. Heads North or East? A Re-Examination of Beaker Burials in Britain. (University of Edinburgh, 2016).
Lillehammer, G. Archaeology of children. Arqueología de la infancia. Complutum 21, 15–45 (2010).
Petersen, F. Traditions of multiple burial in Later Neolithic and Early Bronze Age England. Archaeol. J. 129, 22–55 (1972).
Case, H. Où en sont les Campaniformes de l’autre côté de la Manche?. Bulletin de la Société Préhistorique Française 95, 403–411 (1998).
Salanova, L. Behind the warriors: Bell Beakers and identities in Atlantic Europe (third millennium BC). in Celtic from the West 3: Atlantic Europe in the Metal Ages—questions of shared language 13–39 (Oxbow Books, 2016).
Herrero-Corral, A. M., Garrido-Pena, R. & Flores Fernández, R. The inheritors: Bell Beaker children’s tombs in Iberia and their social context (2500–2000 cal BC). Journal of Mediterranean Archaeology 32.1, 63–87 (2019).
Ulrich, H. Ein Zonenbechergrab von Achenheim im Elsass. Germania: Anzeiger der Römisch-Germanischen Kommission des Deutschen Archäologischen Instituts 26, 175–177 (1942).
Mantel, E. Les sépultures des Petits Prés et du Chemin des Vignes à Léry (Eure): Étude archéologique. Gall. préhistoire 33, 185–192 (1991).
Lefebvre, A., Franck, J., Veber, C. & Duval, H. Les sépultures individuelles campaniformes en Lorraine: l’exemple de Pouilly (Moselle) et d'Hatrize (Meurthe-et-Moselle). in Les sépultures individuelles campaniformes en France (eds. Salanova, L. & Tchérémissinoff, Y.) Gallia Préhistoire, vol. 41 supplément, 97–113 (CNRS Éditions, 2011).
Bloxam, A. The other Beaker people: funerary diversity in Britain c.2450–1950 BC. (UCL, 2020).
Haughton, M. Seeing children in prehistory: a view from bronze age Ireland. Cambridge Archaeol. J. 31, 363–378 (2021).
Haughton, M. Narrativizing Difference in Earlier Bronze Age Society: a comparative analysis of age and gender ideologies in the burials of Ireland and Scotland. (Jesus College, Cambridge, 2020). https://doi.org/10.17863/CAM.53751.
Dresely, V. Schnurkeramik und Schnurkeramiker im Taubertal. Forschungen und Berichte zur Vor- und Frühgeschichte in Baden-Württemberg vol. 81 (Landesamt für Denkmalpflege im Regierungspräsidium Stuttgart – K. Theiss, 2004).
Menninger, M. Die schnurkeramischen Bestattungen von Lauda-Königshofen. Steinzeitliche Hirtennomaden im Taubertal? (Universität Tübingen, 2008).
Fröhlich, M. & Becker, M. Die endneolithische Mehrfachbestattung von Oechlitz, Saalekreis – Eine gemeinsame Grablege der Schnurkeramik- und Glockenbecherkultur. in Neue Gleise auf alten Wegen II Jüdendorf bis Gröbers Band II (eds. Meller, H. & Becker, M.) Archäologie in Sachsen-Anhalt, Sonderband 26/II, 308–314 (Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt – Landesmuseum für Vorgeschichte, 2017).
Haak, W. et al. Die schnurkeramischen Familiengräber von Eulau–ein außergewöhnlicher Fund und seine interdisziplinäre Bewertung. Tagungen des Landesmuseums für Vorgeschichte Halle (Saale) 4, 80–89 (2010).
Häusler, A. Die Gräber der älteren Ockergrabkultur zwischen Dnepr und Karpaten. (De Gruyter, 1976).
Narasimhan, V. M. et al. The formation of human populations in South and Central Asia. Science 365, (2019).
Honeychurch, W. et al. The earliest herders of East Asia: Examining Afanasievo entry to Central Mongolia. Archaeol. Res. Asia 26, 100264 (2021).
Sosna, D. Social differentiation in the Late Copper Age and the Early Bronze Age in South Moravia (Czech Republic). (The Florida State University, 2007).
Blasco, C., von Lettow-Vorbeck, C. L. & Mendoza, P. R. El registro funerario campaniforme de la Región de Madrid reflejo de una sociedad plural y compleja. in ¡Un brindis por el príncipe!: el vaso campaniforme en el interior de la Península Ibérica (2500–2000 A.C.) : Museo Arqueológico Regional, Comunidad de Madrid : exposición, del 9 de abril al 29 de septiembre, 2019 247–277 (Museo Arqueológico Regional, 2019).
Soriano, I., Herrero-Corral, A. M., Garrido-Pena, R. & Majó, T. Sex/gender system and social hierarchization in Bell Beaker burials from Iberia. J. Anthropol. Archaeol. 64, 101335 (2021).
Sjögren, K.-G. et al. Kinship and social organization in Copper Age Europe. A cross-disciplinary analysis of archaeology, DNA, isotopes, and anthropology from two Bell Beaker cemeteries. PLoS One 15, e0241278 (2020).
Wahl, J., Dehn, R. & Kokabi, M. Eine Doppelbestattung der Schnurkeramik aus Stetten an der Donau. Lkr. Tuttlingen. Fundberichte 15, 175–211 (1990).
Rasmussen, S. et al. Early divergent strains of Yersinia pestis in Eurasia 5,000 years ago. Cell 163, 571–582 (2015).
Jelínek, J. A. N. Human sacrifice and rituals in Bronze and Iron Ages: the state of art. Anthropologie 28, 121–128 (1990).
Fyllingen, H. Society and violence in the early bronze age: an analysis of human skeletons from Nord-Trøndelag Norway. Norwegian Archaeol. Rev. 36, 27–43 (2003).
Pany-Kucera, D. & Spannagl-Steiner, M. Social Relations, Deprivation and Violence at Schleinbach, Lower Austria. Insights from an Interdisciplinary Analysis of the Early Bronze Age Human Remains. Archaeologia Austriaca 104, 13–52 (2020).
Rebay-Salisbury, K. et al. Child murder in the Early Bronze Age: proteomic sex identification of a cold case from Schleinbach. Austria. Archaeol. Anthropol. Sci. 12, 265 (2020).
Parker Pearson, M. The practice of human sacrifice. BBC History: Ancient (2012).
Turek, J. Being a Beaker child. The position of children in Late Eneolithic society. In Memoriam Jan Rulf 424–438 (2000).
Martin, R. & Saller, K. Lehrbuch der Anthropologie I-II. Gustav Fischer: Stuttgart, Germany (1957).
Bollongino, R. et al. 2000 years of parallel societies in Stone Age Central Europe. Science 342, 479–481 (2013).
Bramanti, B. et al. Genetic discontinuity between local hunter-gatherers and central Europe’s first farmers. Science 326, 137–140 (2009).
Scheu, A. et al. The genetic prehistory of domesticated cattle from their origin to the spread across Europe. BMC Genet. 16, 54 (2015).
Parker, C. et al. A systematic investigation of human DNA preservation in medieval skeletons. Sci. Rep. 10, 18225 (2020).
Yang, D. Y., Eng, B., Waye, J. S., Dudar, J. C. & Saunders, S. R. Technical note: improved DNA extraction from ancient bones using silica-based spin columns. Am. J. Phys. Anthropol. 105, 539–543 (1998).
Gamba, C. et al. Genome flux and stasis in a five millennium transect of European prehistory. Nat. Commun. 5, 5257 (2014).
MacHugh, D. E., Edwards, C. J., Bailey, J. F., Bancroft, D. R. & Bradley, D. G. The extraction and analysis of ancient DNA from bone and teeth: a survey of current methodologies. Anc. Biomol. 3, 81–102 (2000).
Marchi, N. et al. The genomic origins of the world’s first farmers. Cell 185, 1842-1859.e18 (2022).
Hofmanová, Z. et al. Early farmers from across Europe directly descended from Neolithic Aegeans. Proc. Natl. Acad. Sci. U. S. A. 113, 6886–6891 (2016).
Kircher, M., Sawyer, S. & Meyer, M. Double indexing overcomes inaccuracies in multiplex sequencing on the Illumina platform. Nucleic Acids Res. 40, e3 (2012).
Meyer, M. & Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, db.prot5448 (2010).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Jónsson, H., Ginolhac, A., Schubert, M., Johnson, P. L. F. & Orlando, L. mapDamage2.0: fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 29, 1682–1684 (2013).
Rohland, N. et al. Three assays for in-solution enrichment of ancient human DNA at more than a million SNPs. Genome Res. 32, 2068–2078 (2022).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv [q-bio.GN] (2013).
Li, H. et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Tarasov, A., Vilella, A. J., Cuppen, E., Nijman, I. J. & Prins, P. Sambamba: fast processing of NGS alignment formats. Bioinformatics 31, 2032–2034 (2015).
McKenna, A. et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303 (2010).
Fu, Q. et al. A revised timescale for human evolution based on ancient mitochondrial genomes. Curr. Biol. 23, 553–559 (2013).
Nakatsuka, N. et al. ContamLD: estimation of ancient nuclear DNA contamination using breakdown of linkage disequilibrium. Genome Biol. 21, 199 (2020).
Korneliussen, T. S., Albrechtsen, A. & Nielsen, R. ANGSD: analysis of next generation sequencing data. BMC Bioinf. 15, 356 (2014).
Skoglund, P., Storå, J., Götherström, A. & Jakobsson, M. Accurate sex identification of ancient human remains using DNA shotgun sequencing. J. Archaeol. Sci. 40, 4477–4482 (2013).
Link, V. et al. ATLAS: Analysis tools for low-depth and ancient samples. bioRxiv 105346 (2017) https://doi.org/10.1101/105346.
Mathieson, I. et al. Genome-wide patterns of selection in 230 ancient Eurasians. Nature 528, 499–503 (2015).
1000 Genomes Project Consortium et al. A global reference for human genetic variation. Nature 526, 68–74 (2015).
Rubinacci, S., Ribeiro, D. M., Hofmeister, R. J. & Delaneau, O. Efficient phasing and imputation of low-coverage sequencing data using large reference panels. Nat. Genet. 53, 120–126 (2021).
Weissensteiner, H. et al. HaploGrep 2: mitochondrial haplogroup classification in the era of high-throughput sequencing. Nucleic Acids Res. 44, W58-63 (2016).
Ralf, A., Montiel González, D., Zhong, K. & Kayser, M. Yleaf: Software for Human Y-Chromosomal Haplogroup Inference from Next-Generation Sequencing Data. Mol. Biol. Evol. 35, 1291–1294 (2018).
Wang, C., Zhan, X., Liang, L., Abecasis, G. R. & Lin, X. Improved ancestry estimation for both genotyping and sequencing data using projection procrustes analysis and genotype imputation. Am. J. Hum. Genet. 96, 926–937 (2015).
Lazaridis, I. et al. Genomic insights into the origin of farming in the ancient Near East. Nature 536, 419–424 (2016).
Mallick, S. et al. The Allen Ancient DNA Resource (AADR): A curated compendium of ancient human genomes. bioRxiv (2023) https://doi.org/10.1101/2023.04.06.535797.
Patterson, N. et al. Ancient Admixture in Human History. Genetics 192, 1065–1093 (2012).
Bergström, A. et al. Insights into human genetic variation and population history from 929 diverse genomes. Science 367, (2020).
Ringbauer, H., Novembre, J. & Steinrücken, M. Parental relatedness through time revealed by runs of homozygosity in ancient DNA. Nat. Commun. 12, 5425 (2021).
Ringbauer, H. et al. ancIBD - Screening for identity by descent segments in human ancient DNA. bioRxiv (2023) https://doi.org/10.1101/2023.03.08.531671.
Chaitanya, L. et al. The HIrisPlex-S system for eye, hair and skin colour prediction from DNA: Introduction and forensic developmental validation. Forensic Sci. Int. Genet. 35, 123–135 (2018).
Fowler, C. et al. A high-resolution picture of kinship practices in an Early Neolithic tomb. Nature 601, 584–587 (2022).
Popli, D., Peyrégne, S. & Peter, B. M. KIN: a method to infer relatedness from low-coverage ancient DNA. Genome Biol. 24, 10 (2023).
Ubelaker, D. H. Human Skeletal Remains. Excavation, Analysis, Interpretation (Taraxacum, 1989).
Scheuer, L. & Black, S. The Juvenile Skeleton. (Elsevier, 2004).
Brothwell, D. R. Digging up bones : the excavation, treatment, and study of human skeletal remains. (Cornell University Press, 1981).
Lovejoy, C. O. Dental wear in the Libben population: Its functional pattern and role in the determination of adult skeletal age at death. Am. J. Phys. Anthropol. 68, 47–56 (1985).
Meindl, R. S. & Lovejoy, C. O. Ectocranial suture closure: A revised method for the determination of skeletal age at death based on the lateral-anterior sutures. Am. J. Phys. Anthropol. 68, 57–66 (1985).
Koob, S. P. The removal of aged shellac adhesive from ceramics. Stud. Conserv. 24, 134–135 (1979).
Acknowledgements
This research was funded by the Institut National de Recherches Archéologiques (INRA), Luxembourg, and by the German Research Foundation (DFG), project number 466680522 (‘CO-MOVE’), awarded to Maxime Brami. Nicoletta Zedda thanks the University of Ferrara (grant for the project of Fondazione CRUI ‘Go for It’) and Barbara Bramanti for their support. Yoan Diekmann was funded by the ERC Advanced Grant 788616 (‘YMPACT’), awarded to Volker Heyd. Laura Winkelbach was supported by the DFG, project number BU 1403/19-1. We are grateful to the Luton Cultural Trust for the permission to sample the bones of Dunstable Downs. We thank Richard Harrison, Florian Helmecke, Volker Heyd, Mykyta Ivanov, Joseph Maran, Mike Parker-Pearson, Bianca Preda-Bălănică, Katharina Rebay-Salisbury, Stephen Shennan and Piotr Włodarczak for helpful information about the graves. Thanks also to Paul Majewski for help with the anthropological analyses. Parts of this research were conducted using the supercomputer MOGON2 at Johannes Gutenberg University Mainz (hpc.uni-mainz.de). Finally, we would like to thank the editors and two anonymous referees for their valuable feedback.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Conceptualisation: M.B. Methodology: N.Z., J.Bl., Y.D., M.B. Software: N.Z., J.Bl., Y.D. Validation: N.Z., J.Bl., J.Bu. Investigation: N.Z., K.M., J.Bl., Y.D., M.B. Resources: L.W., E.N., L.B., F.V., F.L.-R., J.Bu. Data curation: N.Z., K.M., J.Bl., Y.D., A.V.G., M.K., K.K., A.-L.T., M.B. Visualisation: J.Bl., Y.D., K.K., A.-L.T., M.B. Supervision: L.W., J.Bl., F.L.-R., J.Bu., M.B. Project administration: M.B. Funding acquisition: M.B., F.L.-R. Writing-original draft: M.B., N.Z., K.M., J.Bl. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zedda, N., Meheux, K., Blöcher, J. et al. Biological and substitute parents in Beaker period adult–child graves. Sci Rep 13, 18765 (2023). https://doi.org/10.1038/s41598-023-45612-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-45612-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
