
Abstract
The Gumelnița site belongs to the Kodjadermen-Gumelnița-Karanovo VI (KGK VI) communities (c. 4700–3900 cal BC) and comprises the tell-type settlement and its corresponding cemetery. This paper reconstructs the diet and lifeways of the Chalcolithic people in the northeastern Balkans using archaeological remains found at the Gumelnița site (Romania). A multi-bioarchaeological investigation (archaeobotany, zooarchaeology, anthropology) was conducted on vegetal, animal, and human remains, alongside radiocarbon dating and stable isotope analyses (δ13C, δ15N) of humans (n = 33), mammals (n = 38), reptiles (n = 3), fishes (n = 8), freshwater mussels shells (n = 18), and plants (n = 24). According to the results of δ13C and δ15N values and FRUITS, the inhabitants of Gumelnița had a diet based on crops and using natural resources, such as fish, freshwater molluscs and game. Although domestic fauna was occasionally exploited for meat, it had a role in providing secondary products. Crops were heavily manured, and chaff and other crop waste may have been necessary fodder for cattle and sheep. Dogs and pigs fed on human waste, although the diet of the latter is more similar to that of wild boars. Foxes had a diet close to dogs, which may indicate synanthropic behaviour. Radiocarbon dates were calibrated with the percentage of freshwater resources obtained by FRUITS. As a result, the corrected dates for the freshwater reservoir effect (FRE) have a delay of an average of 147 years. According to our data, this agrarian community developed a subsistence strategy under the pressure of some climatic changes that started after 4300 cal BC, corresponding to KGK VI rapid collapse/decline episode tracked recently (that begins around 4350 cal BC). This matching of our data in the two models (climatic and chrono-demographic) allowed us to capture the economic strategies that led to the resilience of those people more than other contemporary KGK VI communities.
Similar content being viewed by others


Introduction
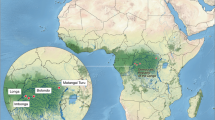
The Kodjadermen–Gumelnița–Karanovo VI (KGK VI) archaeological group reflects the maximum flourishing development of human communities in the Balkans during the Prehistory1,2,3. These people and their material culture features (archaeological artefacts) cover most of the northern Balkans (Fig. 1a) between c. 4700–3900 cal BC. This period is known as Chalcolithic, Copper Age or Eneolithic, representing a distinct period with substantial socio-economic transformations in the target area. Generally, these KGK VI communities occupied a vast geographical area (Fig. 1a) with diverse landscapes and ecosystems, ranging from mountains and hills, to plains, wetlands and costal environments, and from riparian ecosystems to the steppe.

(a) Map showing the location of the Gumelnița site and others mentioned in the text. The maximum spread of KGK VI communities is outlined by the dotted line. This map was created using QGIS 3.20.1-Odense (https://qgis.org). Basemap source: ESRI. (b) Aerial picture showing excavation areas and the location of the graves excavated between 2017 and 2022 (c–e). (c) Graves M1-M8, M11-M16 and C12. (d) Graves M17, M18, M21 and M22. (e) Graves M9 and M10.
KGK VI archaeological and paleogenetic framework
Around 5000 cal BC, in the northern Balkans (Bulgaria and Romania), some fundamental changes took place, such as the rise of tell settlements, the development of the defensive systems—ditches, banks and palisades, the emergence of extramural cemeteries, adoption of some new raw materials—copper, gold, graphite alongside metallurgical innovation. The KGK VI people's lifestyle determined by these innovations and inherent technological progress, led to the development of complex and stratified societies very well reflected by the wealthy graves documented in some extramural cemeteries1,2,3,4.
Following the classic cultural-historical interpretation, most archaeologists still consider that KGK VI originated from the evolution of previous archaeological "cultures" in Bulgaria and Romania (e.g., Boian, Maritsa, Polyanitsa, Karanovo V and Hamangia)4. However, this significant development of KGK VI society could also result from a slow migration process from the Southeast. Recent aDNA analysis demonstrated that both populations (Boian and Gumelnița) have similar genetic signatures due to a common origin in the Northwestern Anatolia from where they arrived in the Balkans5,6,7. This implies that the so-called “cultures” from Northeastern Bulgaria and Southeastern Romania, along with all known innovations, social development, and material cultural progress, were the result of these newcomers, who came with new ideas and managed to occupy the area of the Northern Balkans, a region previously inhabited only sporadically. Newcomers arrived in the northern Balkans around 4901–4751 cal BC, and in the first 300 years, the human population steadily grew, reaching a maximum development and territorial expansion between 4500 and 4400 cal BC; then quickly declined until it vanished from archaeological records around 4150–3800 cal BC8.
From a paleogenomic perspective, this KGK VI people present typical Northwestern Anatolian Neolithic-related ancestry. Also, this population presents some Balkan hunter-gather related ancestry9, along with sporadic evidence of steppe-related ancestry (Varna I and Smyadovo)6, which points towards a genetic admixture between these populations. This interplay of the local, steppe, and western Asian ancestries in Southeastern Europe Chalcolithic people mark the end of the replacement of its local Balkan hunter-gatherers by Anatolian Neolithic farmers, a process that began ~ 8500 years ago7.
This admixture could probably explain some variabilities observed between KGK VI people, at the level of material culture, diet and lifestyle, a situation that could reflects by their different origins, and, consequently, various cultural traditions8.
Gumelnița site
The notoriety of the Gumelnița site is linked to its tell settlement, probably the largest tell settlement of KGK VI people north of the Danube (Fig. S1a). Nearby, the pair cemetery of the KGK VI settlement is documented, along with earlier Neolithic habitation traces (Boian “culture”), and a Cernavodă II cemetery, a Bronze Age settlement (Tei "culture"), and sporadic features corresponding to the first Iron Age and the seventh–eleventh centuries CE10. Only the features assigned to the KGK VI and Boian archaeological groups were radiocarbon dated.
Location
The Gumelnița eponymous site is located in the northern area of the Balkan region (Fig. 1a), in Southeast Romania, on the left bank floodplain of the Danube River, immediately south of the confluence of the Argeş River with a small tributary (Valea Mare)10,11,12,13,14,15.
Excavation biography
The history of this site is linked to the beginnings of the archaeological discipline in Romania. The first archaeological research at this site layed the foundations for the definition of Gumelnița “culture”16,17,18. The mound from “Măgura Gumelnița” or “Măgura Calomfirescu” (the tell settlement) has been mentioned since the nineteenth century, and at the beginning of the twentieth century was the first time investigated archaeologically by Vladimir Dumitrescu (1925). In the interwar period, Dinu V. Rosetti (Bucharest Municipality Museum) and Barbu Ionescu (Oltenița Local Museum) will periodically investigate the site, and in the 60's the excavations will be resumed by a team led by Vladimir Dumitrescu and Silvia Marinescu-Bîlcu. After that, the site would be sporadically investigated by the Oltenița Museum's archaeologists (Călărași County). In 1977, on the terrace (cemetery area) a rescue excavation led to the identification of new graves. Between 2011 and 2012 the site was investigated (partially) again under the umbrella of another rescue excavation. In 2017 our team resumed the archaeological investigation, and since then, research has continued every year except for the lockdown period10,11,13,14,15,16,18.
Tell settlement
The tell mound is located in an erosional remnant from the high terrace of the Danube on the loess deposits11. Although from the ground level, it looks like an elongated mound, the aerial photos show that it has a rectangular shape (Figs. 1b and S1b). The erosional remnant has a length of 272 m and a width of 227 m; the maximum elevation is 38 m.a.s.l, and the area is about 60,000 m2.
Our excavation area is located in the Southeastern corner of the mound (Fig. 1b), not in the top area, where the 2011–2012 rescue trench excavation identified an intense KGK VI habitation.
The archaeological stratigraphy has a thickness of over 4 m (in the top area) and more than 5 m (our excavation), and all anthropic deposits belong to the Gumelnița archaeological signals (A1, A2 and B1 phases or ceramic styles). The absolute chronology of the tell settlement based on the radiocarbon dating of bones, charcoal and seeds (n = 14) is 4800 through 4040 cal BC with a 95.4% probability10 (Table S8).
The KGK VI features identified in our investigations (2017–2021) consist of two burnt and two unburnt houses (features L1, L2, and L3, L4 respectively), some pits (features C1, C2, C3, C5, and C6), and a waste area (feature C4). The investigated features contain different quantities of pottery, flint tools, grinding stones, clay figurines, metal items, and adornments along with animal or plant remains10,11,19. Some of the recovered artefacts (e.g., grinding stones, stone crusher and flint sickle implements) indicate the agricultural activities practised by the Gumelnița people. In addition, there are documented tools related to hunting (e.g., arrowheads and spearheads) and fishing (e.g., clay weights for fishing nets, fishing hooks or harpoons), but also other tools and recipients used for domestic activities (e.g., pots of various dimensions used for cooking, serving or storing the food, stone or flint axes, clay weights for weaving)10,11,19.
Cemetery
The cemetery was identified at the end of the 50's of the 20th century, approximately 200 m East of the tell settlement, when some public works destroyed 7–8 graves. Between 1961 and 1962, in the same area, 5 inhumation graves were examined. In 1977, a rescue excavation led to the identification of other 3 graves. The graves contain skeletons in a crouched position on the left side. Only the individual from grave M3/1962 was in extended dorsal position, a typical burial position for the KGK VI communities that lived on the Black Sea coast (Palazu Mare, Varna I, Durankulak, and Devnja cemeteries)20. In most cases, the individuals had been buried with the head positioned towards east. Our work led to the identification of additional 20 inhumation graves (2017–2022) (Figs. 1c–e, S2, S3 and S4). The skeletons have approximately the same position and orientation as in previously investigated burials in this KGK VI cemetery15,21. Based on the radiocarbon dates performed on human bones (n = 10), the cemetery’s period of use can be estimated to 4545–4179 cal BC (95.4% probability)10. The contemporaneity of the individuals buried in the cemetery is proved by 14C dates obtained for the features investigate on the tell settlement (Table S8).
The grave goods are presented only in a few graves (n = 11) and consist of stone or copper axes, flint blades, bone figurines, copper chisel or shell beads10,11,15,19,22 (Table 1). In some graves, a variety of plant remains were identified (Table S1).
In 3 of the burials (M1, M2, M3) and in C12 pit, human bone fragments which belong to other individuals than those buried in anatomical positions were found. This particular custom fits well in the deviant burials category (addition type) that involves the deliberate incorporation of human bones from another skeleton into a burial of a more or less complete burial25. The situation from Gumelnița cemetery is not a exceptional one, being also documented in the Sultana-Malu Rosu cemetery (Călărași County, Romania) and other KGK VI necropolises26. In the rest of the graves, only single individuals were found buried.
Animal and plant remains
Both plant and animal remains (Tables S1 and S2) from the Chalcolithic period were found on the site10,11,19. Unfortunately, the data regarding faunal and botanical findings from 60's graves are not available (Table 1). Within the mammal remains, domestic animals are the dominant (89.87%), and within them, the most numerous are the cattle (Bos taurus: 41.47%), followed by ovicaprines (Ovis aries/Capra hircus: 32.73%), pigs (Sus domesticus: 9.06%) and dogs (Canis familiaris: 6.61%). Game is 5.54% and is represented by: wild horse (Equus ferus), auroch (Bos primigenius), red deer (Cervus elaphus), roe deer (Capreolus capreolus), wild boar (Sus scrofa), fox (Vulpes vulpes), european badger (Meles meles), hare (Lepus europaeus) and beaver (Castor fiber). The 1966 study27, also concludes that cattle prevail, followed by ovicaprines and pigs, while hunting is poorly represented.
After mammals, molluscs are the animal group with a more significant number of remains. The generus Unio (U. tumidus, U. pictorum, U. crassus and Unio sp.) represents 63.06%, followed by Anodonta sp. (30.07%) and other species (Pseudoanodonta sp., Viviparus sp. Dreissena sp. and Cepaea vindobonensis). The fish with a greater amount of remains are the eurasian carp (Cyprinus carpio) (30.61%) and the catfish (Silurus glanis) (30.61%), followed by the northern pike (Esox lucius) (8.16%). Remains of unidentified species of fishes represent 30.61%. The Reptilia remains belong to the pond turtle (Emys orbicularis). Only four bird bones were identified (3 from small individuals, 1 from a large individual).
Moreover, a large diversity of plants was identified at least at family level10,11, and most of them were recovered from the tell settlement. Most macrobotanical remains were identified at the family level. The cereal remains are the most diverse (and larger batch), with wild and domesticated species. The wild herbaceous cereals species are brome grass (Bromus sp.) and needle grass (Stipa sp.), while the domestic cereals are einkorn wheat (Triticum monococcum), emmer wheat (Triticum dicoccum), barley (naked and hulled) (Hordeum vulgare), and spelt wheat (Triticum cf. spelta).
Pulses are also an important food source documented through both domestic (lentil (Lens culinaris), pea (Pisum sp.) and bitter vetch (Vicia ervilia)), and wild species like vetchling (Lathyrus sp.).
Among the fruits, Prunus sp. stands out because of the numerous fragments of the stone (pyrena) recovered. In some cases it has been possible to reach the species level, thus identifying the cherry plum (Prunus cerasifera). Other wild fruits found were acorn (Quercus robur pedunculiflora), elderberry (Sambucus nigra) and blackberry (Rubus cf. fruticosus). The vegetation signature is completed by some herbaceous plants (Brassica sp., black bindweed (cf. Fallopia convolvulus), fat hen (Chenopodium album), false cleaves (Galium cf. spurium), pale persicaria (Persicaria lapathifolia), common knotgrass (Polygonum aviculare), common sorrel (Rumex acetosa) and water pepper (Persicaria hydropiper)).
The full archaeozoological and archaeobotanical data from the site are described in Lazăr et al.10,19. Tables S1 and S2 indicate the number of remains and the minimum number of individuals.
Stable isotopes
Stable isotopes biogeochemistry as a tool for reconstructing the past
Stable isotope analyses are a standard analytical tool employed in several scientific areas, but also in archaeology in reconstructing components of the dietary system and multiple historical and cultural issues associated with this system28,29. The basic principles of this technique are relatively simple: both carbon and nitrogen occur in nature in two main isotopes, one lighter and more abundant, and the other heavier and scarcer. The ratio of light to heavy isotopes of an element will vary depending on physical–chemical, thermal and biological reactions, which is known as isotope fractionation. This fractionation occurs, for example, between human’s diet and its own isotopic values, being a suitable tool for the reconstruction of the diet of past organisms. The proportion between the light and heavy isotopes of carbon and nitrogen in bone collagen, allows the identification of the origin of the proteins assimilated by the individual, since each large food group has its characteristic isotopic values30,31. In this way, it is possible to differentiate foods of terrestrial origin from marine or freshwater using their carbon isotope ratios, since the terrestrial food chain is based on atmospheric CO2 that plants fix by photosynthesis, while in the aquatic environments carbon derives from carbonate species dissolved in water32,33,34.
In the case of terrestrial plants, we can differentiate C4 plants, which are tropical herbaceous plants among which some are of great interest in human nutrition (millet, corn, sorghum, sugar cane) and have a clearly different carbon isotope signal than C3 plants35,36. For example, the values of δ13C in C3 plants vary between − 30 and − 21‰ with an average of − 26.5 ± 2‰; C4 plants vary between − 15 and − 7‰ with an average of − 12.5 ± 1‰37,38. Among C3 plants, leguminous plants also show a different isotopic signal from the rest, in this case, the nitrogen signal, since they fix N2 directly from air, thanks to structures called mycorrhizae, and not from the nitrates produced by soil bacteria39.
In addition a constant offset in nitrogen isotope values is produced by processes related to the assimilation of food. At each food chain step the nitrogen isotope values increase by 3.5–5‰31,40. Nitrogen isotopes also help to determine whether the food source came from terrestrial or aquatic ecosystems because in the latter, the food web is more complex and nitrogen isotope values are overall higher33. The carbon signal also varies: between a plant and the bone collagen of the herbivore that consumes it, the increase is 5‰; after the primary consumers, each step in the food chain correspond an additional increase in δ13C values of up to 1‰40.
Bone collagen represents 90% of the entire organic fraction of bone, and it is formed from the proteins ingested in the diet. It renews itself very slowly, which makes it possible to reconstruct the diet of humans for at least the last 10 years of their lives30. For other mammals, the turnover rate can be higher, like in dogs, where the rate among young adult dogs is between 6 months and 3 years41.
Conchiolin is the acid-insoluble fraction of the mollusc shell organic matrix and is proteinaceous, so it can be used as a proxy for the soft tissues in isotopic analyses42,43,44. The values obtained in the conchiolin are similar to those of the animal's body42, which represents an approximation of past diets. So far, only marine molluscs have been analysed to study the diets of past humans45,46,47,48.
Carbonized plant materials (combusted under poor oxygen availability) prevent the degradation by microorganisms and their morphological traits are still recognisable. Because of this, wood charcoal and charred seeds and grains are the most common archaeobotanical remains. Studies on modern seeds concluded that there is only a slight change in δ13C and δ15N due to carbonization49. The stable isotopes of the seeds reflect the conditions where the plants grew. The analysis of carbon isotope discrimination (Δ13C) in crop plant remains from archaeological sites is positively correlated with water availability during the period of growth50,51,52. The addition of manuring to the fields can raise the natural δ15N of soils, resulting in crops with higher δ15N than unfertilised soil53.
For the reconstruction of the contribution of the different types of foods in the diet of humans or animals statistical tools have been developed such as Bayesian mixing models54,55. FRUITS (Food Reconstruction Using Isotopic Transferred Signals)56,57 is probably the most used in archaeology and helps to achieve accurate and precise food intake estimates in diet reconstruction studies.
Stable isotopes studies in the KGK VI area
Unfortunately, only a few studies using stable isotope analysis are available in the targeted timespan from the northern Balkans. Varna I and Durankulak cemeteries are the most extensively studied sites in the KGK VI area. The first isotopic results suggested that both populations primarily utilised terrestrial C3-based diets with varying proportions of meat, despite their proximity to the Black Sea58,59, but later a small percentage of marine sourced protein was identified60,61. Mathieson et al.6 published stable isotope data together with ancient DNA and radiocarbon dating from Dzhulyunitsa, Smyadovo, and Sushina sites that belong to the KGK VI area in Bulgaria, but those isotopic results haven’t been interpreted yet. The same happened with the data from Tafani et al.62 from the site of Dridu in Romania.
Regarding domestic animals, cattle were dominant during the Neolithic and also in the KGK VI period63,64, followed by sheep, goats, and pigs. The cattle mortality profile suggests husbandry oriented towards prime meat exploitation (6–24 months) and dairy production (4–9 years)65. The presence of dairy lipid residues in ceramic pots provides direct evidence of this, at least from the sixth millennium BC in the Balkans66,67. For cattle, winter leaf foddering was suggested68. Some of the sheep and cattle from KGK VI show δ13C values compatible with a minor consumption of C4 plants65,69,70, and it has been proposed seasonal exploitation of salt marshes in the Black Sea coastal area.
As for pigs, their hybridisation with wild boar71 makes it necessary to resort to bone metrics to differentiate them. Even so, isotopic interpretation is not easy. However it seems common for wild boars to have lower δ15N values on average, although the distributions of the two overlap72, but the higher contribution of animal protein in pigs' diet suggests that it would be provided by humans and that those pigs would be maintained in the settlement70.
This study aims to reconstruct the diet of the ancient inhabitants of the Gumelnița site and to reconstruct the lifestyle of the KGK VI people that lived here ~ 6000 years ago through the stable isotope analysis of bone collagen, conchiolin extracted from bivalve shells and charred seeds and grains, all recovered from Gumelnița site.
Material and methods
Plants
A total of 24 charred grains/seeds or fragments have been analysed for stable isotopes (Table 2). Grains and seeds analysed here come from our excavations from the years 2017–2020. The domestic plants analysed are emmer wheat (n = 3), einkorn wheat (n = 3), barley (n = 4), lentil (n = 1), pea (n = 2) and bitter vetch (n = 1). The wild seeds and grains analysed are from Prunus sp. (4 fragments coming from 2 different pits), greyish oak (n = 1), blackberry (n = 2) and Bromus sp. (n = 2). All the plants come from pits of the KGK VI period except the bitter vetch which is from a Boian pit10.
To check if there are contaminants in the seeds that are naturally present in the soils and can affect the stable isotope ratios (carbonates, nitrates or humic acids) we used the methods described by Vaiglova et al.77 Due to the small size of the seeds, two samples with the highest weights were chosen (GUM-57 and GUM-61) for testing soil contamination. Samples were crushed and analysed by Fourier Transform Infrared Spectroscopy with Attenuate Total Reflectance (FTIR-ATR) using a Vector 22 from Bruker. Each sample was measured once, the background was subtracted and a baseline correction was performed using OPUS software. The spectra were normalized. The following peaks were observed to test the contamination: 870 and 720 for carbonates; 3300, 1450 and 1085 for nitrates and 3690, 1080 and 1010 for humic acids. Both samples presented a peak in 870 (carbonates contamination), and GUM-61 another one in 1085 (nitrate contamination) (Fig. S5). As carbonates and nitrates were present in the samples, following Vaiglova et al.77, the pre-treatment applied to the sample to remove this contamination was 0.5 M HCl at 80 °C for 30 min (or until effervescence stopped) followed by three rinses in ultrapure Milli-Q water. After this, samples were freeze-dried and analysed for carbon and nitrogen isotope ratios. GUM-57 (now GUM-57-T) and GUM-60 (now GUM-60-T) were again analysed by FTIR-ATR to make sure the contamination had been eliminated. Peaks 870 and 1085 were not present in this new analysis. Figure S5 from Supplementary Materials show the FTIR-ATR spectra (done with Spectragryph 1.2.15) of pre- and post-treatment. Due to the success of the pre-treatment in removing the contaminants, this protocol was used for the rest of the samples. Then samples were freeze-dried for analysis by isotope-ratio mass spectrometry (IRMS).
To correct the values due to the charring effect, we used the corrections of Nitsch et al.73 of − 0.31‰ for the δ15N and − 0.11‰ for the C.
For the calculation of carbon isotope discrimination (Δ13C) in C3 plants, we used the equation from Farquhar et al.51: \(\Delta {}^{13}C=\frac{\delta {}^{13}{C}_{air}-\delta {}^{13}{C}_{p}}{\left(1+\frac{\delta {}^{13}{C}_{p}}{1000}\right)}\), where δ13Cair of the Neolithic period is − 6.5‰78, and δ13Cp is the plant carbon isotope composition.
Water input (WI) represents the rainfall and the irrigation (if applied) during grain filling of barley and wheat grains. For calculations, we used the equations79: Δ13Cbarley = 9.99 + 1.52 × ln(WI) and Δ13Cwheat = 8.50 + 1.78 × ln(WI).
As domestic herbivores eat preferably cereal chaff than grain (which is mainly for human consumption), for calculating the value of the correspondent chaff, offsets of − 2.4‰ of δ15N for both cereals, and − 1.9‰ of δ13C for wheat and − 1.7‰ for barley74,75,76 were used.
Bone collagen
A total of 33 human remains (Table 3) have been selected for isotopic analysis. The information about sex, age, pathologies and grave good can be found in Table 1. From the skeletons excavated during 2017–2022, one sample per individual was selected (except in the case of grave M2, where only M2-I1 was selected) after they were individualized during the anthropological study11,19,23. All of the remains belong to graves with complete skeletons, except the ones from C12, a pit with disarticulated human bones. In this case, we sampled the individuals following the identification of the anthropological study19.
In addition to the graves investigated by us, samples were taken from the human remains recollected during the 60’s on the cemetery (M1/1962, M2/1962 and M3/1962)15 and the tell (M1/1960)24, deposited in the Francisc J. Rainer Institute of Anthropology (Bucharest, Romania). M1/1962, M2/1962 and M3/1962 samples from this old collection come from skeletons in anatomical connection (single graves), for which the bone inventory is recorded. M1/1960-I1 is a child skull discovered in the tell during the excavation of 1960 by Vladimir Dumitrescu. The skull (M1/1960-M1) was found with animal remains and a human vertebra (M1/1960-M2) that belonged to an adult. A synthesis of the new anthropological study done for these human remains excavated in the 60's can be found in Table 1.
Using this sample selection strategy, we tried to cover the entire human population available at Gumelnița site.
23 domestic mammals (6 cattle, 5 dogs, 6 sheep, 1 goat, 5 pigs), 10 wild mammals (1 auroch, 1 beaver, 1 roe deer, 1 red deer, 1 horse, 1 hare, 2 wild boars and 2 red foxes), 2 Sus sp. (due to their fragmentation it was not possible to classify under the wild or the domestic type), 3 reptiles (pond turtle) and 8 fishes (6 catfish and 2 eurasian carps) were selected for collagen extraction (Table 4). The selected bones come from tell features, and from the cemetery (C12 pit). Table S2 contain all zooarchaeological information, including the minimum number of individuals (MNI). Burnt and digested bones are not considered in our study (neither for 14C nor for stable isotopes). Only the human bone of M1-I2 shows some burning traces. Generally, the percentage of the burnt bones in the osteological batch from the Gumelnița site is very small (not more than 1%).
The tell settlement from where animal samples originate comprises a wider range of radiocarbon dates than the cemetery (Table S8), so our isotopic results can reflect variations in human-animal relationship during a larger time interval.
Bones had a standardised pre-treatment for collagen extraction80. An aliquot of bone powder of 5 mg was collected for the sample’s elemental analysis (%C and %N). This analysis allows for assessing the degree of preservation of bone collagen81.
An approximate amount of 250–350 mg of bone powder was employed for collagen purification following Bocherens et al.82 with some in-house improvements. First, the bone powder was demineralized by digestion in 1 M HCl during 20 min. Unwanted organic components were then solubilized by incubation in NaOH 0.125 M during 20 h, at room temperature. Each digestion was followed by a filtration (5μ) to eliminate carbonates, organic contaminants, small fragments of collagen and other organic molecules of low molecular weight. The collagen thus obtained was diluted in mild HCl solution (0.01 M) for 17 h at 100 °C and filtered. The gelatine obtained was freeze-dried to be analysed by IRMS. However, due to the poor results using this method in fish collagen, we tested other variations (see section “Results” of Supplementary Material S1), and, in the end, we employed 500 mg of bone powder and eliminated the NaOH step to extract the collagen.
The bone collagen of RoAMS-1335-110, RoAMS-1336-110 and RoAMS-1337-110 (all cattle remains) were extracted separately10 following Sava et al.83. These samples were analysed directly by IRMS.
Mollusc conchiolin
A total of 17 shell-conchiolin were extracted (3 U. crassus, 4 U. pictorum, 5 U. tumidus and 5 Anodonta sp.), which represent the local freshwater molluscs documented in the site (Fig. S2). The marine shells discovered at Gumelnița (e.g., Spondylus sp., Cardium sp., etc.) are personal ornaments, not remains from cooking or eating.
The conchiolin was obtained by decalcification of the shells following García-Vázquez et al.84 First, the shells were pre-washed with distilled water to remove any remaining soil contaminant. Then the shells were placed in 100 ml beakers. Large shells were broken into fragments. They were immersed in 1 M HCl (except GUM-77 to GUM-80 which were part of a test with 2.5 HCl) till the reaction stopped (CaCO3 + 2HCl → CaCl2 + H2O + CO2). We collected the HCl and placed it in 15 ml centrifuge tubes to remove the supernatant and keep the pellet, which will be the conchiolin. We repeated this process by adding again HCl to the shells to continue de digestion till the reaction stopped and centrifuging the HCl until the shell carbonate was wholly digested. The resultant pellet was then washed with distilled water and centrifuged 3 times. Finally, the pellet was freeze-dried and analysed by IRMS.
To calculate the fractionation between the proteins of the body of the animal and the conchiolin of the shell, we used the offset84: Δ15defatted body-conchiolin = + 0.95‰, Δ13C defatted body-conchiolin = + 0.93‰.
Isotopic measurements
Sample preparation for isotopic analysis was performed at the Stable Isotopes Laboratory of the ArchaeoScience Platform of the University of Bucharest and at the Molecular Palaeontology Lab of the University Institute of Geology (IUX) of the University of A Coruña.
The IRMS analyses were carried out in the Unit of Instrumental Techniques of Analysis (UTIA) of the UDC Research Support Services (SAI). For the stable isotope analysis, approximately 0.5 mg of lyophilised collagen/conchiolin and 0.1 mg (for the smaller seeds) to 5 mg for charred seeds, were loaded into tin capsules and analysed directly on a DeltaV Advantage (ThermoScientific) coupled to a Flash IRMS EA IsoLink CNS (Thermo Scientific) Elemental Analyser Fash IRMS EA Isolink CNS (Thermo Scientific), via a Conflo IV Interface. An analytical precision (reproducibility) was better than 0.2‰ for both carbon and nitrogen. Stable carbon and nitrogen isotope compositions were calibrated relative to Vienna PeeDee Belemnite (VPDB) and atmospheric air (AIR). The following secondary standards were employed: USGS 40 (− 4.52‰), USGS41a (+ 47.55‰), IAEA-N-1 (+ 0.4‰), IAEA-N-2 (+ 20.3‰) and USGS-25 (− 30.4‰) for δ15N, and USGS 40 (− 26.39‰), USGS41a (+ 36.55‰) NBS 22 (− 30.031‰), IAEA-CH-6 (− 10.449‰) and USGS 24 (− 16.049‰) for δ13C. To test for precision acetanilide was employed as an internal standard. Reproducibility resulting in ± 0.15‰ (n = 10). The results are reported using the delta (δ) notation and are expressed in parts per thousand (‰).
Statistics
Statistic calculations and plots were made using RStudio 2022.07.0 + 54885 and Past 4.0586. Plots were edited with Adobe Illustrator CS3.
Radiocarbon dating and freshwater reservoir
A total of 29 14C dates are presented here. Two of them are new and were measured in the labs of CEDAD, University of Lecce (LTL17515A) and Poznań Radiocarbon Laboratory (Poz-52559). The rest were already published10,11,87 (Table S8).
To reconstruct the diet of the population of Gumelnița, we employed the Bayesian mixing model software FRUITS (Food Reconstruction Using Isotopic Transferred Signals)56 (for settings information see Sect. 5.1 of the Supplementary Material S1). The freshwater reservoir effect (FRE) of the Danube Gorges region for a 100% freshwater diet was stabilised at 545 ± 70 14C years88. To correct the human radiocarbon dates for FRE, the percentage of freshwater food in the diet obtained by FRUITS was used to obtain the FRE correction following Sayle et al.89. Radiocarbon dates were calibrated using OxCal 4.490 and the calibration curve from Reimer et al.91 applying the FRE correction only for adult humans.
Results
Charred seeds and grains
The isotopic results of the analysed plants are in Table 2. All the samples were measured in duplicated except GUM-56, GUM-62, GUM-63, GUM-65, GUM-66, GUM-67, GUM-68, GUM-69, GUM-70, GUM-75 and GUM-76. In the case of GUM-73-74, two seeds where measured together.
δ15N values range from 1.3‰ for the pulse bitter vetch, to 13.4‰ for a grain of barley, with a mean of 6.2‰ (± 2.62). δ13C values range from − 28.7‰ for a grain of emmer wheat to − 22.4‰ for another grain of the same species, with a mean of − 24.8‰ (± 1.62). For the domestic cereals, the mean of δ15N values is 6.1‰ (± 2.67) and ranges from 3.7 to 13.4‰ and the mean of δ13C values is − 24.8‰ (± 1.72) with a range of − 28.7 to − 22.4‰. The pulses from Gumelnița period have a mean of 7.4‰ (± 0.58) with a range of 6.9–8.2‰ for δ15N values, and − 23.2‰ (± 0.25) with a range of − 23.4 to − 22.8‰ for the δ13C values.
On average, pulses (14.1) (n = 4) have a higher atomic C:N ratio than the cereals (18.7) (n = 12) due to a higher content of protein, but the range of values overlap (pulses range: 10.6–18.1; cereals range: 11.8–25.1).
Collagen
The isotopic and elemental results obtained from collagen extracted from humans and animals are presented in Tables 3 and 4, respectively. All the samples were measured in duplicated except GUM-4, GUM-7, GUM-17 and GUM-28 which didn’t produce enough collagen for replicates. Elemental analysis in bone powder show %N mean values of 1.4% (± 0.84) with a range of 0.2–3.1. Nitrogen content lower than 0.5% is indicative of poor collagen preservation. Collagen samples with %C higher than 13%, and %N higher than 5%92; with a C:N atomic ratio between 2.9 and 3.693 (where 3.2 is the theoretical value of the collagen94), and an extraction yield higher than 1.5%95 are likely to preserve the original carbon and nitrogen isotopic values.
The majority of human samples meet the quality criteria. Five samples have the %N in bone powder under 0.5% but over 0.3% (GUM-4, GUM-7, GUM-8, GUM-9, and GUM-15). However, the rest of the quality criteria are met. In the case of GUM-4 and GUM-7, only one duplicate measurement could be done, which indicates that the sample had little organic matter preserved, but at the same time the quality of the collagen is acceptable. On average, the δ15N of all adult humans is 10.2‰ (± 0.62) with a range of 9.0–11.5‰, and δ13C is − 20.3‰ (± 0.31) with values ranging from − 21.0 to − 19.6‰. The skeletons identified as females have a mean of 9.8‰ in the δ15N (± 0.45; range: 9.0–10.5‰) and of − 20.3‰ in the δ13C (± 0.31; range: − 21.0 to − 19.8‰). In the case of the skeletons identified as males, they have a mean of δ15N of 10.3‰ (± 0.52; range: 9.5–11.5‰) and of − 20.4‰ (± 0.24; range: − 20.8 to − 20.0‰) for the δ13C. On average, females have slightly lower δ15N values and slightly higher δ13C values than males.
The animal bones with a %N lower than 0.5 in bone powder are GUM-17, GUM-28, GUM-39, GUM-48 and GUM-50. GUM-17 has a yield of 0.9, and GUM-28 of 0.7, so we are going to reject these data based on that. GUM-45, GUM-47, GUM-50 and GUM-52 have the %N in collagen under 5%, and also the %C under 13%. Of these samples, GUM-45 also has a C:N ratio outside the acceptable range. GUM-93 has a C:N ratio greater than 3.6, therefore we did not include these 5 data points.
Domestic mammals have a mean of δ15N values of 7.6‰ (± 1.15; range 5.4–10.5‰) and δ13C values of –19.1‰ (± 1.11; − 21.3 to − 16.9‰), but within this group, there are animals with herbivore and omnivore diets. Domestic herbivores have δ15N values mean of 7.2‰ (± 0.85; range 5.4–8.6‰) and a δ13C values mean of − 18.5‰ (± 0.84; range: − 19.7 to − 16.9‰). Of these, cattle show a lower δ15N values mean (6.9‰ ± 0.67; range: 5.4–7.6‰) and a lower δ13C values mean (− 18.7‰ ± 0.66; range: − 19.7 to − 17.4‰) than the sheep δ15N values (7.5‰ ± 0.99; range: 6.2–8.6‰) and δ13C values mean (− 18.0‰ ± 0.90; range: − 19.3 to − 16.9‰). Wild terrestrial ungulates have a δ15N values mean of 6.3‰ (± 1.14; range: 4.5–7.6‰) and δ13C of − 21.9‰ (± 0.88; range: − 23.2 to − 20.5‰).
Of the domestic omnivores, dogs have higher δ15N values (9.3‰ ± 0.75; range: 8.4–10.5‰) and lower δ13C values mean (− 19.6‰ ± 0.63; range: − 20.6 to − 19.0‰) than pigs δ15N values (7.5‰ ± 0.89; range: 6.5–9.0‰) and δ13C values means (− 20.3‰ ± 0.98; range: − 21.3 to − 18.5‰). These dogs show a diet with a trophic level closer to the humans than the pigs. The δ15N values mean of all suids is 7.4‰ (± 0.87; range: 6.1–9.0‰) and the δ13C values mean − 20.3‰ (± 0.86; range: − 21.3 to − 18.5‰). Wild boars have a lower δ15N values mean (6.7‰ ± 0.53; range 6.1–7.2) and a lower δ13C values mean (− 20.5‰ ± 0.64; range − 21.1 to − 19.8‰) than the pigs. Even though the values are similar, data indicate that pigs have a slightly more animal protein in their diet than the wild boars, which supports the anatomical identification of swine remains. According to these results, pigs' diet could be determined by human management and pigs (but dogs also) used to be fed with human leftovers. In the case of the bones identified and Sus sp. it is not easy to attribute whether they belong to the domestic or the wild species, but their higher δ15N values than the wild boars, suggest they are closer to domestic pigs.
Freshwater-feeding vertebrates (turtles and fishes) have a δ15N values mean of 8.5‰ (± 2.20; range: 5.1–11.2‰) and a δ13C values mean of − 23.2‰ (± 2.15; range: − 26.0 to − 20.4‰). Of these animals, the European carp have the lower δ15N values, as this species is omnivorous, and the catfish the higher values in the δ15N values (10.2‰ ± 0.80; range: 9.2–11.2‰) and δ13C values mean (− 21.5‰ ± 1.51; range: − 24.1 to − 20.4‰). The catfish is the top predator of the Danube and it can also consume terrestrial animals, as suggested by the higher δ13C values. Pond turtles are in an intermediate position (δ15N: 8.4‰ ± 0.55; range: 7.9–8.9‰) (δ13C: − 23.7‰ ± 0.39; range: − 24.1 to − 23.3‰) and are also omnivores.
Conchiolin
Conchiolin results are in Table 5. Data is from García-Vázquez et al.84. Raw values of the conchiolin are transformed into the defatted body because there is an offset between those tissues. The average mean of the values corrected to the defatted body value is 8.5‰ (± 0.74) with a range of 7.2–9.8‰ for the δ15N, and − 25.0‰ (± 2.09) with a range of − 29.0 to − 21.1‰ for the δ13C.
Data modelling
For the FRUITS Bayesian model, we grouped our stable isotope results in terrestrial mammals (domestic and wild), domestic plants (cereals and pulses), fish and shellfish. The results are represented in Fig. 5 (see Table S7 for the numeric values, and figures from S7–S32 for plots of individual results). On average, plant protein represents half of the diet (52%) for this population, followed by freshwater resources (26%) and terrestrial mammals (22%).
The results on the calibration of the radiocarbon dates for the FRE on adult human bones are in Table S8 and are represented in Fig. 7. Due to FRUITS errors transferred to the FRE range, the obtained intervals are wider than the original calibrated dates. Therefore, the difference between calibrated dates with and without FRE is just 147 years.
Discussion
Crop management and the environment of the Gumelnița site
The isotopic values of the plants studied in this paper are represented in a bivariate plot in Fig. 2a. Most domestic plants group in the same area except for two outliers: one in the δ15N (GUM-67), and the other in the δ13C (GUM-54). These two outliers could represent the local conditions of the soil where the plants have grown.

Domestic plants from Gumelnița show medium to high addition of manure, based on Fraser et al.75 (Fig. 2a), especially the pulses. Interestingly, the difference between the domestic pulses (lentil and pea) and the bitter vetch, which has a typical value of a nitrogen-fixing plant96, could indicate that this plant was wild, or if it was cultivated, wasn’t manured. However, N-fixation is inhibited by the presence of inorganic N in the soil97. Thus it may not cause any distinctive signature under manured soils or in alluvial plains96.
Crops are subject to human management, and unlike wild plants, they will mainly reflect human intervention, rather than natural factors. Thus, in each harvest, the soil nutrients assimilated by the plants are removed in the form of biomass. The soils then would be depleted of elements such as nitrogen, so it is necessary to add fertilizer in order to continue harvesting new crops frequently. To avoid this, crop rotation was developed. Nitrogen-fixing plants fix nitrogen from the air, thereby contributing to fertilizing the soil, so it is typical in crop rotation to plant pulses and don’t add any fertilizer that year, but our results don’t support crop rotation with pulses because these plants also have been fertilized.
It would be expected that wild plants would present lower values of δ15N than manured domestic plants, but in our case, except the bitter vetch, all of our charred seed and grains, have δ15N values over 3‰. Due to the tell settlement location in the Danube floodplain, this area possibly had riparian vegetation with high δ15N due to the annual fertilization by the river. The higher values of the blackberry and the Bromus sp. seem to support this. On the terrace and, in general, outside the floodplain, the landscape could be composed of open areas and closed forests. Due to the canopy effect, the oak acorn seems to indicate that it would come from a more closed environment than the Prunus sp. fruits, which present different δ13C values and perhaps belong to different degrees of vegetation coverage. This is also reflected in the isotopic values of wild ungulates.
There's a possibility that cereal crops could be sown in autumn/winter in the floodplain, and harvested in late spring before the Danube floods its banks. These crops should show high δ15N values like GUM-67, but more data would be needed to support this hypothesis. Nor can it be ruled out that the high δ15N values of the legumes come from cultivation in the floodplain.
Our cereal values can be comparable with Neolithic ones from Bulgaria98, where authors also found medium to high contribution of manure in their crops, but the pulses had a low amount of it.
The domestic cereals and pulses from Gumelnița show watering from moderate to well-watered (Fig. 2b), and for GUM-54, this watering was very high. In the case of wild plants, although there are no comparison values established, it does not appear that they lacked water during their growth. The existence of a palaeochannel next to the tell indicates that it could perhaps have been used for irrigation as well as to bring drinking water closer to the settlement. On the assumption that the palaeochannel was used to irrigate the crops, this area closest to the terrace could have been isolated from the Danube flood and it could be where the cultivated fields were located.
Livestock and wild animal resources
The isotopic data of Gumelnița samples are represented in a bivariate plot in Fig. 3. Livestock animals have more positive δ13C values than wild mammals. The δ13C values of the roe deer and the red deer denotes that they come from a more closed environment than the horse, but even this animal has more negative values than sheep and cattle.

Isotopic data of Gumelnița site. Cereals, pulses and chaff are represented by the media and the standard deviation. The δ15N of the forage of wild and domestic herbivores is represented by a dashed line.
The δ15N of the forage is calculated by subtracting 4‰ (as an average of the 3–5‰ range associated with the trophic shift)98. The forage of domestic herbivores is higher (3.2‰) than the wild ones (2.3‰). This may be because they are feeding on cultivated plants such as cereal chaff. In our data, the chaff means calculated from cereal grains (3.7‰) is very similar to the estimated forage. Chaff δ13C mean is − 26.6‰. The cereal grain with less watering (GUM-55) has a chaff value of − 24.3 ‰. The fractionation plant–herbivore is ~ 5‰99, so a herbivore with a 100% diet of this kind of grains, would have a collagen based δ13C value of − 19.3‰. Offset values have not been established for lentil and peas parts, but a small offset in δ15N values of manured Vicia faba between different plant components was found100. The mean of Gumelnița δ13C values in pulses is − 23.2‰, which would give a collagen value for a diet of 100% legumes of − 18.2‰. Feeding with chaff and other crop leftovers can explain part of the δ13C values of cattle and sheep, although a small percentage of C4 plants in the diet is needed to explain all the values.
In Romania, only about 2% of the species are C4101. Roe-deer values from the Cheia site (Hamangia “culture”, first half of the 5th millennium BC, Dobrudja area)69 show a wide variety of ecosystem provenience, and some of them are similar to those of the sheep and cattle values that go towards C4 plants both in Cheia and in our data. In that paper, the authors propose that sheep from Cheia went to feed in the salt marshes on the western coast of the Black Sea where plants of the family Chenopodiaceae are widespread. Analogously to this, the roe deer from Cheia with the most positive values would come from these areas. To test if Gumelnița sheep and cattle with these higher δ13C values fed on the Black Sea coast (a hypothesis that we must consider at least at a theoretical level), a mobility study has to be done because the Gumelnița site is not close to the coast (more than 200 km).
Another possibility with wild C4 plants is the Eragrostio-Polygonion arenastri communities of trampled soil that nowadays exist in the eastern part of the Danube in Romania, characterised by the C4-assimilation syndrome102. Echinochloa crus-galli, or cockspur grass, was found in Gumelnița site (unpublished) and belong to this C4 plants community. This plant is a type of millet native to Eurasia and was domesticated in Japan 4500 year BP ago103. Nowadays can be grown for grain or forage, but also it is mainly found in wet and degraded environments like swamps, ponds or temporarily flooded palustrine wetlands, and seasonally wet habitats104. Therefore, we propose that cockspur grass could have served as food for sheep and cattle that grazed in the vicinity of the site in the floodplain, explaining higher δ13C values of these animals respecting wild herbivores.
Bone fragments from in the tell, show household waste characteristics like traces of cutting, disarticulation, defleshing, burning and traces of carnivorous teeth/swine, etc.11. Dogs’ isotopic values are close to humans, especially GUM-26, which overlaps human distribution. The other 3 data from dogs have a lower δ15N and higher δ13C values and are in the same area (Fig. 3) as the foxes and some of the pigs. This could indicate that those three species could be feeding on human waste, confirming the relation with the teeth marks in the bones. Interestingly, this same isotopic relative position with humans has been found in dogs and foxes of the Bronze Age of the northeastern of the Iberian Peninsula105,106. The earliest synanthropic commensal behaviour of foxes was detected in the Aurignacian, around 42,000 years ago107. This behaviour could explain why dogs and foxes of Gumelnița site have practically the same isotopic values. Hare is one of the potential natural prey of foxes but the δ13C value of the hare from Gumelnița is more than one trophic level apart, which indicates that the fox diet possibly included cultivated plants (cereal grains, pulses), the remains of domestic animals and/or micromammals that fed on crops.
Besides dogs and foxes eating human waste, they were also part of the human diet occasionally or sacrificed for their skin. Cutting marks in dog bones have been found in Gumelnița10 and other KGK VI sites63,64,108,109. Of the two foxes analysed, one was an adult and the other a subadut, and all the dogs older than 6 months, except GUM-29 which was 5–6 months old. This utilisation of the animal bodies could be part of the artificial selection of the domestication process: the dogs that weren’t good enough for work (hunting, herding, etc.) or showed unwanted behaviour could have had this end. This artificial selection process could also control the dog population.
In the case of the suids, they show values corresponding to a gradual increase of animal protein in their diet (tendency to higher δ15N and δ13C). GUM-94 includes the highest amount of terrestrial animal protein in the diet and GUM-41, the lowest, being the difference in δ15N of 2.9‰, almost one trophic level apart. The ecological niche of the wild boar was defined72 for southeast Romania during the Chalcolithic by carbon isotope values always higher than − 22‰ and in most cases higher than − 21‰, which places the wild boar as a species of open environments. In the Gumelnița site the wild boar values are − 21.1‰ and − 19.8‰. Also an enrichment of 1‰ in the wild boar δ15N values (compared with other wild herbivores) was observed72, confirming a more omnivorous diet. But in our case the δ15N is within the variation of wild herbivores, even though the two wild boars analysed are among the suids with lower δ15N of the sampling, suggesting less manured vegetables in the diet. Wild boar diet nowadays is dominated by plants (~ 90%)110, similar to the case of Chalcolithic wild boars from Romania. In the case of the pigs, with different amount of animal protein in the diet, higher δ15N values could be more related to the consumption of domestic animals (and dairy products) and domestic cereals rather than to the freshwater resources or game. Regarding these results, it is not easy to separate wild from domestic pigs, considering their frequent hybridization.
From the freshwater environments, we analysed fishes, molluscs, turtles and a beaver. Of those, the beaver has a semiaquatic life and our value indicates that it fed on C3 plants. The pond turtles are omnivores, whose values are close to one of the catfish and some of the molluscs. Freshwater mussels are omnivores that feed on animal and vegetal particles, living or partially decomposed, present in suspension and on phytobenthic species111. Our data show higher variation in δ13C values due to the different origins of the shells in the sense of habitat, not area. So, the freshwater mussels with less negative δ13C values could be a result of feeding near the river bank or lakeside where there was an influence of terrestrial carbon input112. The rest of the δ13C values variation could be explained by the different eutrophication degrees of the different water bodies, and the most negative values could be explained by the presence of methanogenic bacteria113. Particulate organic matter of the Danube River nowadays has a value of δ15N of 4.88‰114, which is within the trophic level offset of the mean of the freshwater mussels (8.5‰) and their diet.
Regarding the fish, catfish can feed on freshwater fish, molluscs, insects, worms, etc., but also terrestrial animals, birds, and can even predate on other catfish. Our results put them as top predators. All the catfish analysed except GUM-53 show a δ13C value in the middle of freshwater and terrestrial isotopic area, which is consistent with the diet observed in present-day animals. GUM-53 have values that put this individual on a diet based on freshwater animals, in particular, the carp GUM-47 has isotopic values compatible with being the prey type of this catfish. Both GUM-53 and GUM-57 have lower δ15N and δ13C values compared to the other catfish (GUM-53) and to molluscs in the case of GUM-57, which could suggest a different habitat than the other fish.
Human diet
The isotopic values of Gumelnița humans are concentrated, as can be seen in Fig. 3. In Fig. 4a, only human results are represented. The most divergent data are the children. In Fig. 4b, we can observe the differences of the δ15N values in different stages of life. C12-I2 is a 1-year-old baby with the highest δ15N and δ13C values of all the samples. M13, a child between 1 and 3 years old, show a lower δ15N value than C12-I2, and this decrease continues with the rest of the children till the adolescents, which show the lowest δ15N of the non-adults. After this, the values are more homogeneous in the adults. This effect of increasing δ15N values with breastfeeding and decreasing with active growth have been described also in other mammals80,115. During the first months of life, infants have a diet of 100% mothers’ milk, which makes them have a higher trophic level than their mothers. This offset was observed to be about 2–3‰ in humans116,117. From our data, C12-I2 has breastfeeding values, and M13 starts showing a mixture diet of breastmilk and other foods with lower δ15N values. Studies on modern populations suggest that after 6 months of age, the incorporation of more complex foods is required118, which marks the beginning of the weaning process. Nitrogen isotopes are rather insensitive proxies for detecting the start of the weaning process due to bone collagen turnover rates119, so this is why C12-I2 still show that high δ15N value. C12-I2 has an important difference in δ15N values compared with the females. Females have a mean of 9.8‰ and C12-I2 has a value of 14.3‰. The offset is 4.5‰, between the expected trophic level offset (3–5‰)98, but towards the upper limit. Changes in food preference and food consumption during pregnancy120, and the increase of protein requirements during pregnancy and lactation121, make possible that during this period the δ15N values of the female body's blood or milk would be higher, so this effect could be diluted in the δ15N values of the female bones because it reflects the media during the last years of life.

(a) Bivariate plot of δ13C and δ15N of the humans from Gumelnița. Individuals with bold line have flint in their graves. (b) Bivariate plot of age in years vs. δ15N. (c) Bivariate plot of age in years vs.δ13C.
Many typical weaning foods are high in carbohydrates and low in protein122,123, which would produce that difference between C12-I2 and M13. The following children in age group are M3-I2 and M2-I2 of 3 and 4 years, respectively. Again the δ15N values decrease with age but is slower this time. The δ13C values of these children are less negative than the rest of the adults (except C12-I4 and M-12 which show the same value as M3-I2). This can be explained with a diet rich in cereals, vegetables, domestic animal milk and terrestrial meat, and with none or very few foods of freshwater origin. After this, the 4–6 years old child (M1-I1/1960) and the adolescents show similar values with lower δ15N and δ13C as a result of growth physiology and maybe a diet with less protein but now with the inclusion of freshwater resources. M1-I1/1960 show pathologies (criba orbitalia and criba cranii) (Table 1) that could indicate dietary deficiencies or metabolic disorders. Weaning and early dietary transitions through childhood affected the juvenile’s nutritional regime124. Criba orbitalia have been connected to low iron levels due to anemia125 or vitamin (B-12, C) deficiency due to unsanitary living conditions correlated with parasitic infections126. Regarding stable isotopes, opposite results have been found in δ15N values124,125, but these data may indicate that these individuals with criba orbitalia had different diets than the rest. In our case, the difference between M1-I1/1960 and M2-I, could be due to this different diet and related with the pathologies behind criba orbitalia. Human remains are not uncommon intramural21,127, mainly of children127, some of them showing pathologies21, like M1-I1/1960.
On average, the δ15N values of Gumelnița males are slightly higher than in females, but this difference has to be tested statistically. Mann–Whitney test for equal medians of the adults with confirmed sex, in the case of the δ15N, resulted in p = 0.024, which is lower than 0.05, so the null hypothesis of identical distributions for females and males is rejected. For δ13C, was p = 0.445, so for this isotope, they are not significantly different. Doing a hierarchical clustering analysis (Fig. S6) is possible to observe that between the adults there are two main groups (cluster 2.2.1 and cluster 2.2.2) based on δ15N. Cluster 2.2.1 is where most of the men are (and also some of the females and weaned children), and 2.2.2 is formed mainly by females and teenagers, and only one adult male is included.
The dietary intake of adult males is 56 g/day, while in females is 46121. This suggests a higher preference for proteins in males. But during pregnancy and lactation, these requirements increase to 71 g/day121. According to this, it is expected that men could have a higher amount of animal protein in their diet than women. However, it also is expected that women during pregnancy and lactation also had a similar pattern, being nothing of this relating to some social rank or economic status, just human physiology. These differences in preferences towards food with more protein have been observed in a modern population of hunter-gatherers128.
There are 4 graves with flint as grave goods (M8, M9, M14 and M22). All of these skeletons belong to young people (except M9, an adult of unknown age) and are two females and two males. Interestingly their δ13C values are in the middle of the distribution, and these values could correspond with more game resources, indicating that they could be hunters, and this type of work could be carried out by young people.
Only 3 adults are classified as older than 50 years old. This group show a slight decrease in δ15N values compared to the younger groups, but the two males (M11, M6) have higher δ15N values than the female (M16).
The individual with a higher amount of plants in her diet is M16 (Fig. 5), and also has the lowest δ15N among the adults. This woman had a premortem loss of the molars (Fig. S21c), so her diet was under this, and would probably have been dominated by soft foods such as plants and fish. The FRUITS model for this person (Fig. S21) demonstrates this, as the percentage of terrestrial animals in her diet is lower than in the rest of the individuals. M1-I2/1960 (Fig. S29) and M2/1962 (Fig. S31) are among the individuals with a smaller percentage of plants and the adults with higher δ15N, but the model attributes it to freshwater resources, mainly shellfish. C12-I4 also has one of the lower percentages of plants, and the model places this human with the highest percentage of terrestrial mammals.

Percentages of the different food groups estimated by FRUITS.
Looking at the human, game, and domestic δ13C values, it seems more likely that much of the protein assimilated from terrestrial animals came from the game rather than domestic animals. If the Gumelnița settlement was a farming society, the exploitation of domestic animals might be more related to dairy production, exploitation of hair/skin, or even labour power in the fields or for transport. Of the 6 cattle individuals analysed here, two were younger than 1 year, meanings they were used for meat, the same as 5 of the 7 ovicaprines that were between 9 and 12 months old. This indicates that they were indeed consumed, but this would be the result of the fact that to produce milk, cows and sheep have to reproduce, so this consumption would be more occasional, and as a consequence of the milk production process.
To establish if the people's diet from Gumelnița is different (or not) to other communities from the same area and period, we compared our data with those from other Neolithic sites in Romania and Bulgaria (Fig. 6)6,58,62,129 (see Fig. 1a for locations). Thus, Cernica, Dridu, and Dzhulyunitsa overlap with Gumelnița, which means that the inhabitants’ diets could also be similar and that a quarter of the protein in their diets could come from freshwater resources. Employing FRUITS, it was established that Cernica population129 had between 13 and 19% of freshwater fish in their diet, which is a similar result to Gumelnița, but they have not taken into account freshwater mussels that increase the percentage of freshwater resources in the diet. Results shown in Fig. 6, suggest that the amount of protein from freshwater resources was similar between both populations. Cernica, Dridu and Gumelnița sites are all in the north of the Danube, and located close to rivers. Dzhulyunitsa is located south of the Danube, closer to the Balkan Mountains than the Danube, and near a river. Cernica has an older chronology (5355–4940 cal BC129) than Gumelnița, but that only supports the continuity of this type of diet at least since Boian communities.

Situated to the right of the plot are Durankulak and Varna. These two communities are from Hamangia and KGK VI archaeological signals58,59. Varna humans follow a linear relationship59 related to marine fish consumption61. In the case of Durankulak, some individuals overlap with Varna, so they probably had the same diet, but there are others with lower δ15N values but the same range in the δ13C values. These differences in δ15N values can be explained by diets with more percentage of plants, differences in the amount of manuring of the crops, or the consumption of marine molluscs instead of fish by the individuals with lower δ15N.
The KGK VI individuals from Smyadovo and Sushina cemeteries are in the middle of the plot. Smyadovo individuals have very similar δ13C values, but differences in δ15N values, and they overlap with the more negative δ13C values of Varna and Durankulak. Probably they have a more terrestrial diet than the other populations, but more data is needed. In the case of Sushina, we only have 3 data with 3 completely different values that point to different diets: one with more marine food that overlaps with Varna, another with freshwater resources (or game) that overlap with Gumelnița site, and the last one with more terrestrial that overlap with Durankulak. Smyadovo and Sushina are situated in the Balkan Mountains and probably had some communication with Varna or the Black Sea area.
Only one individual from the Gumelnița cemetery (M3/1962) was in the extended dorsal position15, specific for the KGK VI groups from the Black Sea area, also known as “Varna culture”. Considering the isotopic results, this individual has an average diet within the Gumelnița population. If this person was related to the Varna area, he certainly didn’t live there in the last years of his life since his diet is typical of the north of the Danube. A mobility study could clarify this issue.
Isotopic studies (δ13C, δ15N, δ34S) show that hunter-gatherers130,131,132,133,134 from Romania had a higher amount of animal protein in their diet than Neolithic people, especially with a higher amount of freshwater fishes135. The importance of aquatic resources in the diet seems to continue in the Neolithic and Chalcolithic, as we have seen with our results. Depending on the site location, these resources will come from rivers, lakes or the sea.
Reservoir effect
The oldest radiocarbon dates from the cemetery are RoAMS-1335.110 and RoAMS-1624 from a pit (C7) assigned to Boian communities based on the specific archaeological signature (pottery with typical shapes and decorations of the Boian-Vidra ceramic style)10. The next is RoAMS-1616.122, a seed from C6, where pottery from Gumelnița A2 was found10. The youngest of the dates is M16. Whether we use the calibration with or without FRE, all human dates are within the Gumelnița archaeological period (Fig. 7). Interestingly M10 is the older raw human date, but applying the FRE, the older is M3/1962, the only grave where the skeleton is in an extended dorsal position.

Radiocarbon data from the cemetery and the tell calibrated with Oxcal. R indicates that the radiocarbon was calibrated with the FRE (in blue). Mean (empty circle), σ and 2σ are represented in each of the radiocarbon dates.
Burials M15 and M16 were found in the same pit (feature C12) but deposited in overlapping soil layers, and one was placed in front of the other with complementary orientations (Fig. S3e). The difference between them in the mean of the uncalibrated dates is 126, calibrated with no FRE is 136, and with FRE is 72. These differences indicate that they were probably not contemporary, and M16 was buried later, but if the FRE of 540 ± 70 was not correct, M15 could have an earlier date that is masked by higher consumption of freshwater resources. Whatever it is, 2σ distributions of M15 and M16 with FRE overlap more than non-FRE distributions (Fig. 7). Whether we consider the FRE or not, this cemetery was used for at least 300 years. Burial M1-I1/1960 was found in the tell settlement and radiocarbon data of the child fits well into the cemetery 14C dataset. So, it seems that although most of the graves are extramural, KGK VI people still prefer to bury their dead inside the settlement, a funerary tradition that is related to the first Early Neolithic groups in the Balkans.
In Fig. 8, the calibrated human radiocarbon dates with and without FRE and the cattle with available dating are represented in relation to their isotopic values.

δ15N and δ13C vs. radiocarbon dating cal BC (means). Calibrated dates without the FRE are in the filled circles and squares and calibrated data with FRE are in the empty circles. Black line represents the regression of human calibrated dates without the FRE, grey line the regression of human calibrated dates with FRE, and green line the cattle regression.
The resilience of the KGK VI community at Gumelnița site
The human group designed as KGK IV developed within the Atlantic period (6900–3700 cal BC), a warm-moist of Blytt and Sernander’s classification. At the end of this period (c. 4100–3700 cal BC), the climate got cooler and drier136. This transition towards the next period (Subboreal) was set by the 5.9 ka event (Bond event 4)137. This event has also been observed in lipid biomarkers, pollen and speleothems studies138,139 in the Carpathian mountains. During this period, along with fading of many KGK VI tell settlements, other contemporary "cultures" from this part of Europe collapsed too (e.g., Tiszapolgár in Transylvania; the end of Cucuteni A-B and the beginning of B1 phase in Moldova)136. However, in the case of KGK VI, some communities are more resilient than others, and the Gumelnița site is one of those.
In the 300 years covered by the cemetery radiocarbon dates, a decrease in δ15N values can be observed (Fig. 8), but this could be an artefact due to the sex of the individuals (the two youngest dates are females). Besides this, M16 (the youngest date) shows the lowest δ15N value of the adults, probably due to molar loss (Fig. S21c). The precipitation decreased in Romania from 4300 cal BC136, which coincides with the oldest graves (FRE). Aridity increases δ15N values, but the decrease in temperatures, also decreases δ15N values of soils140, producing opposite effects. The domestic plants baseline is not only affected by these climate changes but only by the manuring process, which is an anthropic one. Therefore, a decrease in agricultural activity could be behind these values, which could indicate the decline of the KGK VI civilization. Interestingly, this decrease in precipitation corresponds very well with the KGK VI population dynamics model recently proposed by Popescu et al.8, more precisely with the decline episode of these communities, which started at c. 4350 BC, without a significant recovery, until it vanished toward the end of the 5th millennium BC–early 4th millennium BC.
This situation proves that the Gumelnița settlers were a more resilient community than others, temporarily resisting the shock produced by climatic changes and economic-cultural collapse, which led to the disappearance of many KGK VI tell settlements up to 4200 cal BC8. The fact that the Gumelnița community resisted to the collapse, is also proven by the cemetery's radiocarbon data, which shows that people most likely survived until the beginning of the 4th millennium BC, according to the corrected FRE (Table S8).
Besides the few data on cattle with 14C dates, these results show that δ15N values haven't changed since the Boian period. However, in the case of δ13C values, Gumelnița cattle turned towards those C4 values that were already mentioned. This could be a sign that the pastures of the Gumelnița site area could be less suitable for these animals, and they have to move with the herds at least some part of the year (maybe in summer, as it is the C4 growth period) to areas with wild C4 plants, or the trampled soil ecosystem with C4 plants102, developed near the settlement area during the KGK VI period. On the other hand, four “C4 episodes” were identified over the Balkans141 during the Late Pleistocene. Those episodes were related to dry and cold summers, which don't allow C3 plants to grow, thus supporting C4 development and considering the precipitation decrease in Romania at 4300 cal BC, along with temperature decrease, may have a "C4 episode" here but on a smaller scale, meaning that the percentage of C4 plants was higher during this period.
On the other hand, the fact that naked barley is the most frequent crop in the site, followed by hulled barley and einkorn, indicates another clue regarding Gumelnița people's resilience. Ecologically, barley is well adapted to dry conditions, and it is more susceptible to waterlogged conditions and vulnerable to low temperatures (compared to wheat). Hulled barley had a more significant environmental tolerance than naked barley, and it responded better to environmental changes. Einkorn can grow in cold and dry environments too. This shows a deliberate selection of the cultivated cereals by Gumelnița settlers, apparently species resistant to climate change stress.
Moreover, the documented wild plant species at the Gumelnița site (e.g., Prunus type, followed by blackberry and hazelnut) are not that susceptible to climate change. For example, some Prunus species are adaptable to various conditions, except light. The blackberry is also a very adaptable plant; hazelnuts can resist cold and high temperatures.
Conclusions
The Gumelnița cemetery was used for more than 300 years (4545–4179 cal BC) and corresponded to the period of use of the even settlement (4800–4040 cal BC). If the entire chronological sequence of KGK VI is documented on the tell, the cemetery's radiocarbon data reflects only the field research stage. The human remains buried here reflect the maximum period of development of KGK VI communities (4500–4400 cal BC)8 but also the fast decline period that followed it and marked the collapse of this flourish VI civilization,8. The FRE has been applied in the calibration of radiocarbon dates on humans. Although the range obtained is larger than in the calibrations without the FRE, on average, there is a lapse of 147 years between these calibrations that not affect the archaeological interpretation of the features presented here.
Overall, due to taxonomically diverse material and a consistent chronology from Gumelnița site, the analysis of δ15N and δ13C has allowed us to get a closer look at the people’s way of life of at the Gumelnița site. The first observation is that these people had a terrestrial diet (74%) with a consistent contribution to aquatic resources (26%). Regarding terrestrial resources, we note that domestic plants (52%) play the most crucial role in feeding these people, followed by terrestrial mammals (22%). The diet of the KGK VI population was supplemented with aquatic resources, more specifically shellfish (18%) and fish (8%).
Considering this data, we could conclude that the KGK VI community from Gumelnița was an agrarian society in which half of the protein of the diet came from cereal crops (wheat and barley) and legumes (lentils and peas). The rest of the protein came mainly from freshwater resources (freshwater mussels and fish) and hunting, with meat from livestock being a more occasional resource. Finally, domestic herbivores would be exploited primarily to obtain secondary resources such as dairy products, fur, manure and labour power, an activity not yet demonstrated for the Chalcolithic period in the Eastern Balkans142, but in the Western Balkans143.
However, the diet profile of the Gumelnița population seems to be dominant in northern side of the Danube River, and it has existed already since Boian and Hamangia archaeological groups144, and it is related to the arrival of Anatolian Neolithic farmers in the area6. This situation reflects the perpetuation of culinary traditions, between generations, over several hundred years between these human groups.
The agricultural practices of the KGK VI community from Gumelnița involve manured and irrigated crops, highlighting the use of this manure in legumes, this being the first direct evidence of these processes within KGK VI human groups. Cereal chaff and other crops wastes were used to feed cows and sheep. Also, the dogs and pigs consumed human waste (according to isotopic signature), although the latter had a less carnivorous diet, with different degrees. The foxes appear to have had a diet similar to that of the dogs, indicating possible synanthropic behaviour, a situation which is also for the first time documented in KGK VI area.
Human dietary data does not seem to indicate that there were different classes or ranks within this Chalcolithic society, so it looks pretty egalitarian, although there is a slight difference between the animal protein consumed by men and women, which could be the effect of the particular preferences based on physiological differences. There is no gender difference of grave goods either, which in this society seems quite scarce. In the case of the diet variations on gender classes, we observe some minor differences between men and women, which consist in a slightly higher δ15N in men. Also, for age categories, there is a decrease in δ15N from infants to adolescents determined by lactation and later by active growth. Once adulthood is reached, the diet is homogenous, and is determined by small variations based by own tastes and physiology either due to gender or pregnancy and lactation. Older individuals appear to have a small decrease in δ15N, however, in the case of M16, with premortem loss of molars, her diet would have softer foods such as plants or fish.
On the other hand, slight dietary variations noticed in the human population from the Gumelnița site may reflect the stages of development and decline of the KGK VI civilization. This situation proves that the Gumelnița settlers were a more resilient community than others, temporarily resisting the stress produced by climatic changes and economic-cultural collapse, which led to the disappearance of most KGK VI tell settlements between 4350 cal BC to 4200 cal BC8. The fact that the Gumelnița community resisted better to the collapse was due to human adaptation to the new climate conditions (precipitation decreased after 4300 cal BC136) that led to the abandonment of most of the KGK VI tell settlements after this moment. Thus, being an agrarian community, which had a diet based on plants, as we have shown in this study, the Gumelnița people, to support the crops, used intensive soil fertilization and crop watering alongside the selection of cereal species resistant to climate change stress, as proven by our data. In this way, the community managed to survive for several hundred years more than the other KGK VI settlements, as shown in 14C data from the cemetery (Table S8). However, the KGK VI decline episode was a continuous process spanning almost 550 years (c. 4350–3800 BC)8, and Gumelnița community resilience was only a temporary solution. The socio-economic recovery of it never happened.
Finally, we do not exclude those human dietary variations recorded at the Gumelnița site be the result of particular traditions of different human individuals related to the place of their origin or the paleogenetic background documented for KGK VI (e.g., Anatolian Neolithic farmers, Balkan hunter-gatherers or people with steppe-related ancestry), but this correlation will be made into a further study. Thus, the next steps will be represented by isotopic investigations related to the mobility (δ18O, 87Sr/86Sr) of these individuals, doubled by paleogenetic analyses, in order to increase the accuracy of the lifestyle reconstruction model of the KGK VI human groups.
Data availability
The data sets supporting the conclusions of this article are included within the article and in the Supplementary data file.
References
Bailey, D. W. Balkan Prehistory. (Routledge, 2000). https://doi.org/10.4324/9780203461969.
Chapman, J. A Life in Balkan Archaeology. (Oxbow Books, 2021). https://doi.org/10.2307/j.ctv1wvndcs.
Anthony, D. W. The rise and fall of Old Europe. in The lost world of Old Europe: The Danube Valley, 5000–3500 BC (eds. Anthony, D. W. & Chi, J. Y.) 26–57 (Institute for the Study of the Ancient World at NYU/Princeton University Press, 2010).
Lazăr, C., Mărgărit, M. & Radu, V. Between Dominant Ideologies and Techno-economical Constraints: Spondylus Ornaments from the Balkans in the 5th Millennium BC. in Interchange in Pre- and Protohistory. Case Studies in Iberia, Romania, Turkey and Israel. British Archaeological Reports, International Serie. (eds. Cruz, A. & Gibaja Bao, J. F.) 5–21 (Hadrian Books Ltd, 2018).
Hervella, M. et al. Ancient DNA from South-East Europe reveals different events during early and Middle Neolithic influencing the European Genetic Heritage. PLoS ONE 10, e0128810 (2015).
Mathieson, I. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018).
Lazaridis, I. et al. The genetic history of the Southern Arc: A bridge between West Asia and Europe. Science 377, eabm4247 (2022).
Popescu, G. M. et al. Sine qua non: Inferring Kodjadermen-Gumelniţa-Karanovo VI population dynamics from aggregated probability distributions of radiocarbon dates. Radiocarbon 65, 463–484 (2023).
González-Fortes, G. et al. Paleogenomic Evidence for Multi-generational Mixing between Neolithic Farmers and Mesolithic Hunter-Gatherers in the Lower Danube Basin. Curr. Biol. 27, 1801-1810.e10 (2017).
Lazăr, C. et al. Gumelnița: Research results of the 2018 and 2019 fieldwork seasons. Rev. Cercet. Arheol. şi Numis. 6, 13–100 (2020).
Lazăr, C. et al. Gumelnița: Then and now. The research results of the 2017 fieldwork. Stud. Preist. 14, 119–174 (2017).
Dumitrescu, V. O statuetă neolitică de marmură descoperită la Gumelnița. Stud. şi Cercet. Istor. Veche 14, 139–143 (1963).
Dumitrescu, V. Gumelnița—sondajul stratigrafic din 1960. Stud. şi Cercet. Istor. Veche 17, 51–60 (1966).
Dumitrescu, V. & Marinescu-Bîlcu, S. Noi descoperiri de la Gumelnița. Cult. şi Civilizaţie la Dunărea Jos 16–17, 114–144 (2001).
Lazăr, C. A. Date noi privind unele morminte Gumelnitene. Preist. Dunarii Jos XVI–XVII, 173–183 (2001).
Dumitrescu, V. Fouilles de Gumelnița. Dacia 2, 29–103 (1925).
Dumitrescu, V. Découvertes de Gumelnița. Dacia 1, 325–342 (1924).
Dumitrescu, V. Oameni și Cioburi. Contribuții la istoria contemporană a arheologiei românești. (Muzeul Dunării de Jos, 1993).
Lazăr, C. et al. Gumelnița: research results of the 2021 fieldwork season. Rev. Cercet. Arheol. și Numis. 7, 13–54 (2021).
Lazǎr, C. Rituri şi ritualuri funerare în neoliticul şi eneoliticul din România. (Cetatea de Scaun, 2021).
Lazar, C. et al. The catalogue of the Neolithic and Eneolithic funerary findings from Romania. (Cetatea de Scaun, 2012). https://doi.org/10.13140/2.1.2028.0320.
Șerbănescu, D. Vestigii neolitice descoperite la Ulmeni. Cult. şi Civilizaţie la Dunărea Jos 1, 25–34 (1985).
Vasile, G. Analiza antropologică a scheletelor eneolitice din necropola aşezării eponime de la Gumelniţa (jud. Călăraşi, România). J. Archaeol. Anthropol. Interdiscip. Stud. 1, 43–57 (2019).
Nicolăescu-Plopșor, D. Expertiza antropologică asupra unui craniu de copil descoperit la Gumelnița în 1960. Stud. şi Cercet. Istor. Veche 17, 109–112 (1966).
Chapman, J. ‘Deviant’ burials in the Neolithic and Chalcolithic of Central and South Eastern Europe. in Body parts and bodies whole: changing relations and meanings 30–45 (Oxbow Books, 2010).
Lazăr, C. The eneolithic necropolis from Sultana-Malu Rosu (Romania)—a case study. in Mobility and Transitions in the Holocene. Proceedings of the XVI World Congress of the International Union of Prehistoric and Protohistoric Sciences. BAR International Series 2658. (eds. Oosterbeek, L. & Fidalgo, C.) 67–74 (Archaeopress, 2014).
Necrasov, O. & Haimovici, S. Studiul resturilor de faună neolitică, descoperite în stațiunea Gumelnița. SCIV 17, 101–108 (1966).
Schwarcz, H. P. & Schoeninger, M. J. Stable isotope analyses in human nutritional ecology. Yearb. Phys. Anthropol. 34, 283–321 (1991).
Reitsema, L. J. Beyond diet reconstruction: Stable isotope applications to human physiology, health, and nutrition. Am. J. Hum. Biol. 25, 445–456 (2013).
Tykot, R. H. Stable isotopes and diet: you are what you eat. in Proceedings of the International School of Physics “Enrico Fermi”, Course CLIV (eds. Martini, M., Milazzo, M. & Piacentini, M.) 433–444 (IOS Press, 2004).
Reynard, L. M. & Hedges, R. E. M. Stable hydrogen isotopes of bone collagen in palaeodietary and palaeoenvironmental reconstruction. J. Archaeol. Sci. 35, 1934–1942 (2008).
Chisholm, B., Nelson, D. E. & Schwarcz, H. P. Stable-carbon isotope ratios as a measure of marine versus terestrial protein in anciet diets. Science 216, 1131–1132 (1982).
Schoeninger, M., Deniro, M. J. & Tauber, H. Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220, 1381–1383 (1983).
Webb, E. C. et al. The influence of varying proportions of terrestrial and marine dietary protein on the stable carbon-isotope compositions of pig tissues from a controlled feeding experiment. STAR Sci. Technol. Archaeol. Res. 3, 28–44 (2017).
van der Merwe, N. J. Carbon Isotopes, Photosynthesis, and Archaeology: Different pathways of photosynthesis cause characteristic changes in carbon isotope ratios that make possible the study of prehistoric human diets. Sigma Xi, Sci. Res. Soc. Carbon 70, 596–606 (1982).
Pearcy, R. W. & Ehleringer, J. Comparative ecophysiology of C3 and C4 plants. Plant Cell Environ. 7, 1–13 (1984).
O’Leary, M. H. Carbon isotopes in photosynthesis. Bioscience 38, 328–336 (1988).
O’Leary, M. H. Carbon isotope fractionation in plants. Phytochemistry 20, 553–567 (1981).
Marshall, J. D., Brooks, J. R. & Lajtha, K. Sources of variation in the stable isotopic composition of plants. in Stable isotopes in ecology and environmental Science (eds. Michener, R. & Lajtha, K.) 22–60 (Blackwell Publishing, 2007).
Bocherens, H. & Drucker, D. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: Case studies from recent and ancient terrestrial ecosystems. Int. J. Osteoarchaeol. 13, 46–53 (2003).
Martin, R. B., Burr, D. B., Sharkey, N. A. & Fyhrie, D. P. Skeletal Tissue Mechanics. (Springer New York, 2015). https://doi.org/10.1007/978-1-4939-3002-9.
Watanabe, S., Kodama, M. & Fukuda, M. Nitrogen stable isotope ratio in the manila clam, Ruditapes philippinarum, reflects eutrophication levels in tidal flats. Mar. Pollut. Bull. 58, 1447–1453 (2009).
Delong, M. D. & Thorp, J. H. Mollusc shell periostracum as an alternative to tissue in isotopic studies. Limnol. Oceanogr. Methods 7, 436–441 (2009).
O’Donnell, T. H., Macko, S. A., Chou, J., Davis-Hartten, K. L. & Wehmiller, J. F. Analysis of δ13C, δ15N, and δ34S in organic matter from the biominerals of modern and fossil Mercenaria spp. Org. Geochem. 34, 165–183 (2003).
Arnay-de-la-Rosa, M. et al. Dietary patterns during the early prehispanic settlement in La Gomera (Canary Islands). J. Archaeol. Sci. 36, 1972–1981 (2009).
Arnay-de-la-Rosa, M. et al. Paleodietary analysis of the prehistoric population of the Canary Islands inferred from stable isotopes (carbon, nitrogen and hydrogen) in bone collagen. J. Archaeol. Sci. 37, 1490–1501 (2010).
Bownes, J. M., Ascough, P. L., Cook, G. T., Murray, I. & Bonsall, C. Using stable isotopes and a bayesian mixing model (FRUITS) to investigate diet at the Early Neolithic Site of Carding Mill Bay, Scotland. Radiocarbon 59, 1275–1294 (2017).
Schulting, R. J., MacDonald, R. & Richards, M. P. FRUITS of the sea? A cautionary tale regarding Bayesian modelling of palaeodiets using stable isotope data. Quat. Int. 650, 52-61. https://doi.org/10.1016/j.quaint.2022.02.012 (2023).
DeNiro, M. J. & Hastorf, C. A. Alteration of 15N/14N and 13C/12C ratios of plant matter during the initial stages of diagenesis: Studies utilizing archaeological specimens from Peru. Geochim. Cosmochim. Acta 49, 97–115 (1985).
Farquhar, G. D., Ehleringer, J. R. & Hubick, K. T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 503–537 (1989).
Farquhar, G., O’Leary, M. & Berry, J. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Funct. Plant Biol. 9, 121 (1982).
Araus, J. L. et al. Crop water availability in early agriculture: Evidence from carbon isotope discrimination of seeds from a tenth millennium BP site on the Euphrates. Glob. Chang. Biol. 5, 201–212 (1999).
Bogaard, A., Heaton, T. H. E., Poulton, P. & Merbach, I. The impact of manuring on nitrogen isotope ratios in cereals: Archaeological implications for reconstruction of diet and crop management practices. J. Archaeol. Sci. 34, 335–343 (2007).
Phillips, D. L. et al. Best practices for use of stable isotope mixing models in food-web studies. Can. J. Zool. 92, 823–835 (2014).
Stock, B. C. et al. Analyzing mixing systems using a new generation of Bayesian tracer mixing models. PeerJ 6, e5096 (2018).
Fernandes, R., Millard, A. R., Brabec, M., Nadeau, M.-J. & Grootes, P. Food reconstruction using isotopic transferred signals (FRUITS): A Bayesian model for diet reconstruction. PLoS ONE 9, e87436 (2014).
Fernandes, R., Grootes, P., Nadeau, M.-J. & Nehlich, O. Quantitative diet reconstruction of a Neolithic population using a Bayesian mixing model (FRUITS): The case study of Ostorf (Germany). Am. J. Phys. Anthropol. 158, 325–340 (2015).
Honch, N. V., Higham, T. F. G. G., Chapman, J., Gaydarska, B. & Hedges, R. E. M. M. A palaeodietary investigation of carbon (13C/12C) and nitrogen (15N/14N) in human and faunal bones from the Copper Age cemeteries of Varna I and Durankulak Bulgaria. J. Archaeol. Sci. 33, 1493–1504 (2006).
Honch, N. et al. West Pontic diets: A scientific framework for understanding the Durankulak and Varna I cemeteries, Bulgaria. Interdiscip. Archaeol. 4, 147–162 (2013).
Higham, T., Slavchev, V., Gaydarska, B. & Chapman, J. AMS dating of the late copper age Varna cemetery, Bulgaria. Radiocarbon 60, 493–516 (2018).
Gaydarska, B., Beavan, N. & Slavchev, V. Lifeway interpretations from ancient diet in the Varna cementery. Oxford J. Archaeol. 41, 22–41 (2022).
Tafani, A., Lazăr, C. & Tykot, R. H. Stable isotope data of Neolithic and Eneolithic populations in the Balkans, 6600 to 4000 BC. Data Br. 42, 108114 (2022).
Bălăşescu, A., Moise, D. & Radu, V. The Palaeoeconomy of Gumelniţa Communities on the Territory of Romania. in In Honorem Silvia Marinescu-Bîlcu 167–195 (DAIM P.H., 2005).
Bǎlǎşescu, A. Arheozoologia neo-eneoliticului de pe Valea Teleormanului. (Editura Mega, 2014).
Gillis, R. et al. Sophisticated cattle dairy husbandry at Borduşani-Popină (Romania, fifth millennium BC): The evidence from complementary analysis of mortality profiles and stable isotopes. World Archaeol. 45, 447–472 (2013).
Stojanovski, D. et al. Living off the land: Terrestrial-based diet and dairying in the farming communities of the Neolithic Balkans. PLoS ONE 15, e0237608 (2020).
Evershed, R. P. et al. Dairying, diseases and the evolution of lactase persistence in Europe. Nature 608, 336–345 (2022).
Balasse, M. et al. Early herding at Măgura-Boldul lui Moş Ivănuş (early sixth millennium BC, Romania): Environments and seasonality from stable isotope analysis. Eur. J. Archaeol. 16, 221–246 (2013).
Balasse, M. et al. Cattle and Sheep Herding at Cheia, Romania, at the Turn of the Fifth Millennium cal BC. A View from Stable Isotope Analysis. Proc. Br. Acad. 198, 115–142 (2014).
Balasse, M. et al. Investigating the scale of herding in Chalcolithic pastoral communities settled along the Danube River in the 5th millennium BC: A case study at Borduşani-Popină and Hârşova-tell (Romania). Quat. Int. 436, 29–40 (2017).
Stanc, M. S., Luca, M., Bălășescu, A. & Bejenaru, L. Holocene genetic evolution of Pig (Sus scrofa) on Romanian Territory in a European time and space frame. Diversity 14, 288 (2022).
Balasse, M. et al. Wild, domestic and feral? Investigating the status of suids in the Romanian Gumelniţa (5th mil. cal BC) with biogeochemistry and geometric morphometrics. J. Anthropol. Archaeol. 42, 27–36 (2016).
Nitsch, E. K., Charles, M. & Bogaard, A. Calculating a statistically robust δ13C and δ15N offset for charred cereal and pulse seeds. STAR Sci. Technol. Archaeol. Res. 1, 1–8 (2015).
Vaiglova, P. et al. An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: a new look at Neolithic farming. J. Archaeol. Sci. 42, 201–215 (2014).
Fraser, R. A. et al. Manuring and stable nitrogen isotope ratios in cereals and pulses: Towards a new archaeobotanical approach to the inference of land use and dietary practices. J. Archaeol. Sci. 38, 2790–2804 (2011).
Wallace, M. et al. Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol. 45, 388–409 (2013).
Vaiglova, P., Snoeck, C., Nitsch, E., Bogaard, A. & Lee-Thorp, J. Impact of contamination and pre-treatment on stable carbon and nitrogen isotopic composition of charred plant remains. Rapid Commun. Mass Spectrom. 28, 2497–2510 (2014).
Vaiglova, P. et al. Further insight into Neolithic agricultural management at Kouphovouno, southern Greece: Expanding the isotopic approach. Archaeol. Anthropol. Sci. 12, 43 (2020).
Ferrio, J. P., Araus, J. L., Buxó, R., Voltas, J. & Bort, J. Water management practices and climate in ancient agriculture: Inferences from the stable isotope composition of archaeobotanical remains. Veg. Hist. Archaeobot. 14, 510–517 (2005).
García-Vázquez, A., Pinto-Llona, A. C. & Grandal-d’Anglade, A. Brown bear (Ursus arctos L.) palaeoecology and diet in the Late Pleistocene and Holocene of the NW of the Iberian Peninsula: A study on stable isotopes. Quat. Int. 481, 42–51 (2018).
Bocherens, H., Drucker, D., Billiou, D. & Moussa, I. Une nouvelle approche pour évaluer l’état de conservation de l’os et du collagène pour les mesures isotopiques (datation au radiocarbone, isotopes stables du carbone et de l’azote). Anthropologie. 109, 557–567 (2005).
Bocherens, H. et al. Isotopic biogeochemistry (13C,15N) of fossil vertebrate collagen: Application to the study of a past food web including Neandertal man. J. Hum. Evol. 20, 481–492 (1991).
Sava, T. B. et al. Status report on the sample preparation laboratory for radiocarbon dating at the New Bucharest Roams Center. Radiocarbon 61, 649–658 (2019).
García-Vázquez, A., Radu, V. & Lazăr, C. New method for the isotopic study of ancient conchiolin from archaeological shells of freshwater mussels (Unionoida). 01 December 2022, Prepr. (Version 1) available Res. Sq. (2022). https://doi.org/10.21203/rs.3.rs-2313604/v1.
R Core Team. R: A language and environment for statistical computing. (2020).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 9 (2001).
Bem, C. Noi propuneri pentru o schiţă cronologică a eneoliticului românesc. Pontica 33–34, 25–121 (2000).
Cook, G. T. et al. Problems of dating human bones from the Iron Gates. Antiquity 76, 77–85 (2002).
Sayle, K. L., Hamilton, W. D., Gestsdóttir, H. & Cook, G. T. Modelling Lake Mývatn’s freshwater reservoir effect: Utilisation of the statistical program FRUITS to assist in the re-interpretation of radiocarbon dates from a cemetery at Hofstaðir, north-east Iceland. Quat. Geochronol. 36, 1–11 (2016).
Bronk Ramsey, C. bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Reimer, P. J. et al. The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Ambrose, S. H. Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17, 431–451 (1990).
DeNiro, M. J. Post-mortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317, 806–809 (1985).
Schwarcz, H. P. & Nahal, H. Theoretical and observed C/N ratios in human bone collagen. J. Archaeol. Sci. 131, 105396 (2021).
van Klinken, G. J. Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Archaeol. Sci. 26, 687–695 (1999).
Fiorentino, G., Ferrio, J. P., Bogaard, A., Araus, J. L. & Riehl, S. Stable isotopes in archaeobotanical research. Veg. Hist. Archaeobot. 24, 215–227 (2015).
Shearer, G. & Kohl, D. N2-fixation in field settings: Estimations based on natural 15N abundance. Funct. Plant Biol. 13, 699–756 (1986).
Bogaard, A. et al. Crop manuring and intensive land management by Europe’s first farmers. Proc. Natl. Acad. Sci. 110, 12589–12594 (2013).
Drucker, D. G., Bridault, A., Hobson, K. A., Szuma, E. & Bocherens, H. Can carbon-13 in large herbivores reflect the canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeogr. Palaeoclimatol. Palaeoecol. 266, 69–82 (2008).
Treasure, E. R., Church, M. J. & Gröcke, D. R. The influence of manuring on stable isotopes (δ13C and δ15N) in Celtic bean (Vicia faba L.): Archaeobotanical and palaeodietary implications. Archaeol. Anthropol. Sci. 8, 555–562 (2016).
Pyankov, V. I., Ziegler, H., Akhani, H., Deigele, C. & Lüttge, U. European plants with C4 photosynthesis: Geographical and taxonomic distribution and relations to climate parameters. Bot. J. Linn. Soc. 163, 283–304 (2010).
Carni, A. & Mucina, L. Vegetation of trampled soil dominated by C4 plants in Europe. J. Veg. Sci. 9, 45–56 (1998).
Purugganan, M. D. & Fuller, D. Q. The nature of selection during plant domestication. Nature 457, 843–848 (2009).
Quattrocchi, U. CRC World dictionary of grasses: common names, scientific names, eponyms, synonyms, and etymology (CRC Press, 2006).
Grandal-d’Anglade, A. et al. Dogs and foxes in Early-Middle Bronze Age funerary structures in the northeast of the Iberian Peninsula: human control of canid diet at the sites of Can Roqueta (Barcelona) and Minferri (Lleida). Archaeol. Anthropol. Sci. 11, 3949–3978 (2019).
Albizuri, S. et al. Dogs that ate plants: Changes in the Canine diet during the Late Bronze Age and the First Iron Age in the Northeast Iberian Peninsula. J. World Prehist. 34, 75–119 (2021).
Baumann, C., Bocherens, H., Drucker, D. G. & Conard, N. J. Fox dietary ecology as a tracer of human impact on Pleistocene ecosystems. PLoS ONE 15, e0235692 (2020).
Bălășescu, A. & Radu, V. Paléo-économie animalière et reconstitution de l’environnement. in Société et environnement dans la zone du Bas Danube durant le 5ème millénaire avant notre ère (eds. Carozza, L., Bem, C. & Micu, C.) 385–407 (Editura Universității Al. I. Cuza, 2011).
Bréhard, S. & Bălăşescu, A. What’s behind the tell phenomenon? An archaeozoological approach of Eneolithic sites in Romania. J. Archaeol. Sci. 39, 3167–3183 (2012).
Ballari, S. A. & Barrios-García, M. N. A review of wild boar Sus scrofa diet and factors affecting food selection in native and introduced ranges. Mamm. Rev. 44, 124–134 (2014).
Makhutova, O. N. et al. Feeding spectra of bivalve mollusks Unio and Dreissena from Kanevskoe Reservoir, Ukraine: Are they food competitors or not?. Zool. Stud. 52, 56 (2013).
Vuorio, K., Tarvainen, M. & Sarvala, J. Unionid mussels as stable isotope baseline indicators for long-lived secondary consumers in pelagic food web comparisons. Fund. Appl. Limnol. 169, 237–245 (2007).
Schenk, J. et al. Methane in lakes: Variability in stable carbon isotopic composition and the potential importance of groundwater input. Front. Earth Sci. 9, 722215 (2021).
Bănaru, D., Harmelin-Vivien, M., Gomoiu, M.-T. & Onciu, T.-M. Influence of the Danube River inputs on C and N stable isotope ratios of the Romanian coastal waters and sediment (Black Sea). Mar. Pollut. Bull. 54, 1385–1394 (2007).
Hobson, K. A. & Quirk, T. W. Effect of age and ration on diet-tissue isotopic (Δ13C, Δ15N) discrimination in striped skunks (Mephitis mephitis). Isotopes Environ. Health Stud. 50, 1–7 (2014).
Fogel, M. L., Tuross, N. & Owsley, D. W. Nitrogen isotope tracers of human lactation in modern and archeological populations. in Carnegie Institution 1988–1989, Annual Report Geophysical Laboratory 111–117 (Geophysical Laboratory, Carnegie Institution, 1989).
Fuller, B. T., Fuller, J. L., Harris, D. A. & Hedges, R. E. M. Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am. J. Phys. Anthropol. 129, 279–293 (2006).
Dewey, K. G. & Brown, K. H. Update on technical issues concerning complementary feeding of young children in developing countries and implications for intervention programs. Food Nutr. Bull. 24, 5–28 (2003).
Tsutaya, T. & Yoneda, M. Quantitative reconstruction of weaning ages in archaeological human populations using bone collagen nitrogen isotope ratios and approximate Bayesian computation. PLoS ONE 8, e72327 (2013).
Faas, M. M., Melgert, B. N. & De Vos, P. A brief review on how pregnancy and sex hormones interfere with taste and food intake. Chemosens. Percept. 3, 51–56 (2010).
Dwyer, J. Dietary Reference Intakes (DRIs): Concepts and Implementation. in Encyclopedia of Gastroenterology 613–623 (Elsevier, 2004). https://doi.org/10.1016/B0-12-386860-2/00613-4.
Sellen, D. W. & Smay, D. B. Relationship between subsistence and age at weaning in “preindustrial” societies. Hum. Nat. 12, 47–87 (2001).
WHO. Indicators for assessing infant and young child feeding practices: Part 1 definitions. (World Health Organization, 2008).
Osorio, L. S. Exploring the relationship between nutrition and Cribra Orbitalia: The comparison of dietary stable isotope comparisons of Juveniles from Kuelap, Peru. (University of Central Florida Part, 2022).
Zariſa, G. et al. Cribra orbitalia as a potential indicator of childhood stress: Evidence from paleopathology, stable C, N, and O isotopes, and trace element concentrations in children from a 17th-18th century cemetery in Jekabpils, Latvia. J. Trace Elem. Med. Biol. 38, 131–137 (2016).
Walker, P. L., Bathurst, R. R., Richman, R., Gjerdrum, T. & Andrushko, V. A. The causes of porotic hyperostosis and cribra orbitalia: A reappraisal of the iron-deficiency-anemia hypothesis. Am. J. Phys. Anthropol. 139, 109–125 (2009).
Schuster, C., Kogălniceanu, R. & Morintz, A. The living and the dead: an analysis of the relationship between the two worlds during prehistory at the Lower Danube. (Cetatea de Scaun, 2008).
Berbesque, J. C. & Marlowe, F. W. Sex differences in food preferences of Hadza Hunter-Gatherers. Evol. Psychol. 7, 601–616 (2009).
Stratton, S. et al. The emergence of extramural cemeteries in Neolithic Southeast Europe: A formally modeled chronology for Cernica, Romania. Radiocarbon 61, 319–346 (2019).
Trinkaus, E. et al. Stable isotope evidence for early modern human diet in Southeastern Europe: Peştera cu Oase, Peştera Muierii and Peştera Cioclovina Uscată. Mater. Şi Cercet. Arheol. (serie nouă) V, 5–14 (2009).
Borić, D., Grupe, G., Peters, J. & Mikić, Ž. Is the mesolithic-neolithic subsistence dichotomy real? New stable isotope evidence from the Danube Gorges. Eur. J. Archaeol. 7, 221–248 (2004).
Doboş, A., Soficaru, A., Popescu, A. & Trinkaus, E. Radiocarbon Dating and Faunal Stable Isotopes for the Galeria Principală , Peştera Muierii, Baia de Fier, Gorj County, Romania. Mater. Şi Cercet. Arheol. (serie nouă) V, 15–20 (2009).
Bonsall, C. et al. Interrelationship of age and diet in Romania’s oldest human burial. Naturwissenschaften 99, 321–325 (2012).
Bonsall, C. et al. Radiocarbon and stable isotope evidence of dietary change from the mesolithic to the middle ages in the iron gates: New results from Lepenski Vir. Radiocarbon 46, 293–300 (2004).
Nehlich, O., Borić, D., Stefanović, S. & Richards, M. P. Sulphur isotope evidence for freshwater fish consumption: A case study from the Danube Gorges. SE Eur. J. Archaeol. Sci. 37, 1131–1139 (2010).
Harper, T. K. Demography and climate in Late Eneolithic Ukraine, Moldova, and Romania: Multiproxy evidence and pollen-based regional corroboration. J. Archaeol. Sci. Rep. 23, 973–982 (2019).
Bond, G. et al. A pervasive millennial-scale cycle in north Atlantic Holocene and glacial climates. Science 278, 1257–1266 (1997).
Ramos-Román, M. J. et al. Lipid biomarker (brGDGT)- and pollen-based reconstruction of temperature change during the Middle to Late Holocene transition in the Carpathians. Glob. Planet. Change 215, 103859 (2022).
Drǎguşin, V. et al. Constraining Holocene hydrological changes in the Carpathian-Balkan region using speleothem δ18O and pollen-based temperature reconstructions. Clim. Past 10, 1363–1380 (2014).
Amundson, R. et al. Global patterns of the isotopic composition of soil and plant nitrogen. Global Biogeochem. Cycles 17, 1031 (2003).
Hatté, C. et al. Excursions to C4 vegetation recorded in the Upper Pleistocene loess of Surduk (Northern Serbia): An organic isotope geochemistry study. Clim. Past 9, 1001–1014 (2013).
Bǎlǎşescu, A., Moise, D. & Radu, V. Utilisation des bovins à la traction dans le Chalcolithique de Roumanie: première approche. in Actes du colloque « De l’araire au chariot. Premières tractions animales de l’Europe de l ‘Ouest » 269–273 (Editions Centre Nationale des Recherches Scientifiques, 2006).
Gaastra, J. S., Greenfield, H. J. & Linden, M. V. Gaining traction on cattle exploitation: Zooarchaeological evidence from the Neolithic Western Balkans. Antiquity 92, 1462–1477 (2018).
Bălășescu, A. & Radu, V. Oameni şi animale. Strategii şi resurse la comunităţile preistorice Hamangia şi Boian. (Editura Cetatea de Scaun, 2004).
Acknowledgements
We would like to thank Mădălina Dimache, Valentin Parnic, Adelina Darie, Bogdan Manea, Vlad Bîndea, Cristian Roth and Silviu Cristache for their participation in field excavations, Dan Pîrvulescu and Theodor Zavalaș for logistic and administrative support. Many thanks to Aurora Grandal d’Anglade for giving us access to the Molecular Paleontology Lab of the University Institute of Geology of the University of A Coruña (Spain), and to Viorel Nicu Atudorei (New Mexico University, USA) for proofreading the text. Finally, we dedicate this study to the memory of Professor Silvia Marinescu-Bîlcu (1932-2023), who started more than 60 years ago the first archaeological research in the KGK VI cemetery at Gumelnița site, and permanently supported interdisciplinary research in Romania. Sit tibi terra levis.
Funding
This work of University of Bucharest team was supported by a grant from the Ministry of Research, Innovation, and Digitisation (41PFE/30.12.2021) within PNCDI III. A. García-Vázquez was supported by a Fellowship of Young Researchers (1908/20.01.2022) and a Research Fellowship for Visiting Professors (515/10.01.2022) within the Research Institute of the University of Bucharest (ICUB) under the project “Multi-isotopic approach to the life at the Gumelnița site”. The work of G. Sava and C. Lazar was supported by a grant of the UEFISCDI, project no. 351PED/2020 (CALIB-RO), within PNCDI III. The V. Radu’s and A. Bălășescu’s contribution were supported by a grant from the Ministry of Research, Innovation and Digitization, CNCS/CCCDI-UEFISCDI (PN-III-P4-ID- PCE-2020-2369), within PNCDI III.
Author information
Authors and Affiliations
Contributions
A.G.V. and C.L. conceived and supervised the study. A.G.V. performed stable isotopic analysis. T.I., V.O., M.G., C.C., and C.L. performed archaeological excavation, recording and assembling data. V.R., A.B., G.V., M.G., M.C., T.I., A.G.V., and C.L. performed the sampling. A.B., V.R., M.C., G.V., and M.G. made the archaeobotanical, zooarchaeological, and anthropological investigations. V.O., T.I., C.C., and C.L. processed the archaeological data. G.S. performed the radiocarbon dating. A.G.V. and C.L. wrote the manuscript and the supplementary materials with the input of all other co-authors. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
García-Vázquez, A., Bălășescu, A., Vasile, G. et al. Unravelling the resilience of the KGK VI population from the Gumelnița site (Romania) through stable isotopes. Sci Rep 13, 8499 (2023). https://doi.org/10.1038/s41598-023-35129-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-35129-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
