
Abstract
Comprising the oldest unequivocal dinosauromorphs in the fossil record, silesaurs play an important role in the Triassic radiation of dinosaurs. These reptiles provide the main source of information regarding the ancestral body plan of dinosaurs, as well as the basis for biogeographic models. Nevertheless, the co-occurrence of silesaurs and the oldest unequivocal dinosaurs is rare, which hampers reliable ecological inferences. Here we present the first species of silesaur from the oldest unequivocal dinosaur-bearing beds from Brazil. Amanasaurus nesbitti gen. et sp. nov. possesses a unique set of femoral traits among silesaurs, including the oldest occurrence of an anterior trochanter separated by the femoral shaft by a marked cleft. Its femoral length indicates that the new species rivals in size with most coeval dinosaurs. This find challenges the assumption that in faunas where silesaurs and unambiguous dinosaurs co-occurred, silesaurs were relatively smaller. Moreover, the presence of dinosaur-sized silesaurs within ecosystems with lagerpetids, sauropodomorphs and herrerasaurids reinforces the complex scenario regarding the early radiation of Pan-Aves. Silesaurs—independent of their phylogenetic position—persisted during most of the Triassic Period, with its plesiomorphic body size advancing through the dawn of dinosaurs, instead of silesaur lineages decrease in body size through time.
Similar content being viewed by others

Introduction
Silesaurs are part of the wide Triassic radiation of archosaurs1,2. Most silesaurs are characterized by slender limbs and a “beak-like” projection from the anterior tip of the lower jaw3. Whereas these reptiles are present in the fossil record of Middle to Upper Triassic4,5,6,7,8, no records have been reported from Jurassic or younger strata9,10. Silesaurs are particularly interesting because of their close phylogenetic relationships with dinosaurs4,11,12, with several studies placing silesaurs as the closest evolutionary relatives of dinosaurs4,11,13. On the other hand, there are alternative hypotheses where silesaurs are recovered as members of Ornithischia9,10,14,15. This latter scenario supports two main models: (i) silesaurs are part of a monophyletic assemblage (i.e., wide Silesauridae) that is the sister group of “traditional/core ornithischians”14,15,16; or (ii) silesaurs are assembled in low-diversity clades representing a stem group leading to “traditional/core ornithischians”9,10. Despite the competing affinities of silesaurs, these reptiles are key taxa in order to understand the dawn of the avian stem lineage. Silesaurs are the oldest dinosauromorphs reported in the fossil record4,5, providing clues on the ancestral body plan and biogeography of the group. The Middle Triassic occurrences from Brazil, Tanzania, and Zambia support a gondwanan origin of silesaurs4,5, whereas during the Upper Triassic, the group was present in both, Gondwana17,18,19,20 and Laurasia7,21,22. Although restricted to Argentina and Brazil, the fossil record from South America is particularly rich. There are two species from Argentina: Lewisuchus admixtus23, from the early Carnian beds of Chañares Formation; and Ignotosaurus fragilis, from the late Carnian of the Ischigualasto Formation18. Regarding Brazil, there are two species too: Gamatavus antiquus8, from the Ladinian/early Carnian of Santa Maria Formation; and Sacisaurus agudoensis17, from the early Norian of Caturrita Formation. In addition, there is an unnamed silesaur reported from the mid-to-late Carnian beds of Santa Maria Formation24. This unnamed material is remarkable because it comes from the oldest unequivocal dinosaur-bearing beds worldwide25, providing evidence of the co-occurrence of distinct groups of Pan-Aves during this crucial moment. Unfortunately, the scarcity of silesaurs from these beds obscures our understanding of the ecological relationships between these groups. Here, we describe the first silesaur species from Carnian (Upper Triassic) beds from Brazil and discuss its role on the evolutionary history of the group.
Material and methods
Institutional abbreviations
CAPPA/UFSM Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia da Universidade Federal de Santa Maria, São João do Polêsine, Rio Grande do Sul, Brazil; CRILAR-Pv Paleontologıa de Vertebrados, Centro Regional de Investigaciones Cientıficas y Transferencia Tecnologica, Anillaco, Argentina; MNA Museum of Northern Arizona, Flagstaff, USA; NMMNH New Mexico Museum of Natural History and Science, Albuquerque, USA; UFSM Laboratório de Estratigrafia e Paleobiologia, Universidade Federal de Santa Maria, Santa Maria, Rio Grande do Sul, Brazil.
Specimen
The specimens here described are housed at the palaeovertebrates collection of the Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia/Universidade Federal de Santa Maria (CAPPA/UFSM), under the specimen number CAPPA/UFSM 0374 and CAPPA/UFSM 0375.
Phylogenetic analysis
In order to access the phylogenetic affinities of the new silesaur, it was scored in the data matrix of Norman et al.10, which is a modified version of the data matrix published by Müller & Garcia9. This is the most comprehensive dataset regarding silesaurs. Furthermore, we inserted Gamatavus antiquus8, a recently described silesaur from Brazil. Its scoring was performed through first-hand examinations of the holotype (UFSM 11348a, a partial right ilium) and a referred specimen (UFSM 11348b, a partial left femur). Chilesaurus diegosuarezi was removed from the data matrix because of its controversial affinities26. The most parsimonious trees were recovered in the software TNT v. 1.527. All characters received the same weight and characters 4, 13, 18, 25, 63, 82, 84, 87, 89, 109, 142, 166, 174, 175, 184, 186, 190, 201, 203, 205, 209, 212, 225, 235, 236, 239, 250 and 256 were treated as additive (ordered). Euparkeria was used to root the most parsimonious trees, which were constructed using random addition sequence + tree bisection reconnection (TBR), which included 1000 replicates of Wagner trees (with random seed = 0), TBR and branch-swapping (holding 20 trees saved per replicate).
Femoral length estimation criteria
The total femoral length of CAPPA/UFSM 0374 and CAPPA/UFSM 0375 was estimated according to two ordinary least squares linear regressions employing the dataset of Barrett et al.28. This dataset includes measurements of 31 femora of distinct Triassic and Lower Jurassic ornithodirans. The femoral length of CAPPA/UFSM 0374 was estimated using the proximal long axis of the femoral head as the independent variable, whereas the femoral length of CAPPA/UFSM 0375 was estimated using the distal long axis as the independent variable.
Results
Systematic paleontology
Archosauria Cope, 1869
Pan-Aves Gauthier & de Queiroz, 2001
Dinosauromorpha Benton, 1985
Silesauridae Nesbitt et al., 2010
Amanasaurus nesbitti gen. et sp. Nov.
Holotype
CAPPA/UFSM 0374 (Table 1), a proximal portion of a right femur.
Etymology
The genus combines the Tupi word “amana” (= rain) and the Greek “saurus” (= lizard), referring to the Carnian pluvial episode. The specific epithet honors Dr. Sterling J. Nesbitt, a prominent North American paleontologist, for his contribution and studies on silesaurs and Triassic archosaurs.
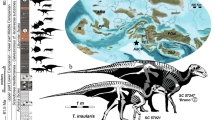
Type locality, age, and horizon
Pivetta site (29°39′37″ S, 53°25′51″ W), between the municipalities of Restinga Sêca and São João do Polêsine, Rio Grande do Sul, Brazil (Fig. 1). Lower portion of the Candelária Sequence29 of the Santa Maria Supersequence30, Paraná Basin. The presence of the rhynchosaur Hyperodapedon places the Pivetta site within the Hyperodapedon Assemblage Zone31, which is considered mid to late Carnian (Late Triassic) in age according to high-precision U–Pb zircon geochronology that indicated a maximum age of 233.23 ± 0.73 Ma32.

Provenance of Amanasaurus nesbitti gen. et sp. nov. (a) Surface distribution of the geologic units in the area depicting the location of the Pivetta site. (b) General view of the Pivetta site. (c) hypothetical reconstruction of the skeleton of Amanasaurus nesbitti gen. et sp. nov. depicting (in orange) the preserved portions. (d) CAPPA/UFSM 0374 (holotype), a proximal portion of a right femur in anterior view. (e) CAPPA/UFSM 0375 (referred specimen), a distal portion of a left femur in anterior view. Figures were generated with GIMP 2.8 (https://www.gimp.org/).
Referred specimen
CAPPA/UFSM 0375 (Table 1), a distal portion of a left femur from an individual slightly larger than the holotype and excavated from the same locality.
Diagnosis
Amanasaurus nesbitti differs from all other known silesaurs with comparable material in (*local autapomorphies): posteromedial tuber of the femoral head reduced to absent; ventral margin of the anteromedial tuber exceeding the femoral head margin; presence of a fossa trochanterica; absence of a raised anterolateral scar; presence of a semi-circular scar on the posterodorsal surface of the femoral head*; cleft between the proximal tip of the anterior trochanter and the femoral shaft (see Supplementary Information for a differential diagnosis).
Description
The femoral head of Amanasaurus nesbitti is well-preserved (Fig. 2a–e). The bone surface preserves fine details and the specimen shows no evidence of sedimentary compression. Similar to other silesaurs, the femoral head is poorly expanded transversely. This condition differing the Am. nesbitti from most dinosaurs and pterosauromorphs33,34. It is triangular in proximal view, with a deep straight groove separating the anterior and posterior surfaces (Fig. 2c). This groove is absent in Lewisuchus admixtus20,34. The medial articular surface is straight, whereas in L. admixtus and Asilisaurus kongwe it is rounded19,35. The anterolateral tuber is well-developed, as well as the anteromedial tuber. The later forms the posteromedial margin of the femoral head (Fig. 2e), delimiting the distalmost extension of the articular surface of the proximal portion of the femoral head, such as in Sacisaurus agudoensis14. Distinct from the other tubers, the posteromedial tuber is poorly developed, lacking a sulcus for ligamentum captis femoris in proximal view. The posteromedial tuber of L. admixtus, As. kongwe and Eucoelophysis baldwini is well-developed19,35, differing from the new specimen. There is a reduced fossa trochanterica (Fig. 2c), resembling As. kongwe35. In L. admixtus it is well-developed19, whereas in other silesaurs it is absent7,14,21,36. The greater trochanter is angled, whereas in lagerpetids it is rounded15,37,38. The specimen lacks the “overhang structure” on the proximal surface, which is reported for some specimens of Silesaurus opolensis39. The typical “notch” between the ventral transition from the femoral head to the femoral shaft is present (Fig. 2a). This differs from the concave emargination that marks the transition in most dinosaurs33. In addition, the medial articular surface of the femoral head bears a transverse scar above the notch (Fig. 2d). In Si. opolensis there is a similar scar that forms the ventral margin of the attachment point of the iliofemoral ligament40. There is a smooth surface on the homologous surface of Sa. agudoensis14. Its surface is reduced in the Am. nesbitti. There is an unusual sub-circular scar on the posterior surface of the femoral heart (Fig. 2e), slightly below the fossa trochanterica. An identical scar was not reported for other silesaurs.

Holotype and referred specimen of Amanasaurus nesbitti gen. et sp. nov. from the Candelária Sequence (mid-to-late Carnian) of the Santa Maria Supersequence, southern Brazil. Holotype (CAPPA/UFSM 0374) in anterior (a), lateral (b), proximal (c), medial (d), and posterior (e) views. Referred specimen (CAPPA/UFSM 0375) in anterior (f), lateral (g), posterior (h), and distal (i) views. alt anterolateral tuber, amt anteromedial tuber, at anterior trochanter, cl cleft, ctfb crista tibiofibularis, dlt dorsolateral trochanter, dltp posterior portion of the dorsolateral trochanter, fo foramen, fot fossa trochanterica, gt greater trochanter, lc lateral condyle, lia linea intermuscularis cranialis, mc medial condyle, ms muscle scar, no notch, pg proximal groove, pof popliteal fossa, scs sub-circular scar. Figures were generated with GIMP 2.8 (https://www.gimp.org/).
The anterior surface of the dorsolateral trochanter is sharp and merges with the shaft well-below the proximal articular surface (Fig. 2a). In contrast, this trochanter is rounded for all the ontogenetic stages sampled for As. kongwe41. A proximodistally oriented scar runs on the lateral surface of this trochanter (Fig. 2b), whereas a transverse scar extends from the trochanter to the posterior margin of the femoral shaft (Fig. 2e). A raised anterolateral scar is absent, whereas it is reported for L. admixtus, As. kongwe, and Si. opolensis19,20,39,41. Indeed, there are faint striations on the homologous surface. The anterior trochanter is finger-like, extending proximodistally. Its proximal tip is separated from the femoral shaft by a cleft (Fig. 2b), such as in several silesaurs, prionodontians, and theropods10,11,14,42. On the other hand, in L. admixtus and As. kongwe the proximal tip merges smoothly with the femoral shaft, lacking the cleft19,20,35. Whereas the proximal tip of the anterior trochanter is not connected to the shaft in the new specimen, it is far less expanded than the wing-like trochanter of several prionodontians10,43,44. The condition of the new specimen is also distinct from the pyramidal anterior trochanter of some theropods42,45. The proximal portion of the linea intermuscularis cranialis rests medial to the anterior trochanter (Fig. 2a). It is absent in Sa. agudoensis14. A foramen pierces the femoral shaft medial to the proximal portion of the cranial intermuscular line (Fig. 2d). The trochanteric shelf is absent, a condition shared with Sa. agudoensis, E. baldwini, Kwanasaurus williamparkeri, Diodorus scytobrachion, and prionodontians7,10,11,14,36.
The referred distal portion of a left femur bears a well-preserved bone surface (Fig. 2f–i). On the other hand, the bone is fragmented. Distinct from lagerpetids15,37, the anterior surface is convex in distal view (Fig. 2i), lacking any evidence of an extensor grove. There is a raised scar extending from the anterior to the lateral surface of the bone (Fig. 2f–h), which is common for dinosauromorphs. The lateral margin of the lateral condyle is rounded in distal view and there is a depression between this condyle and the crista tibiofibularis. The exact size and shape of the crista tibiofibularis and the medial condyle are uncertain. Whereas the popliteal fossa is not entirely preserved, it is proximodistally elongated (Fig. 2h), resembling the condition of most silesaurs and aphanosaurs4,46. A raised scar runs from the surface above the crista tibiofibularis into the popliteal fossa.
Phylogenetic analysis
The heuristic search recovered 1728 most parsimonious trees (MPTs) of 1074 steps each, with a consistency index of 0.298 and a retention index of 0.689. The general topology of the strict consensus tree (Fig. 3a) follows that recovered by Norman et al.10, where silesaurs are nested in low diversity groups in the branch that leads to Prionodontia (i.e., traditional ornithischians). Amanasaurus nesbitti nests as a parapredentatan within Ornithischia in all the MPTs. The new taxon nests in a trichotomy with Ignotosaurus fragilis and Silesarus opolensis, which is supported by a fossa on the ventral surface of postacetabular part of ilium (ch. 174: 1 → 2), iliac lamina two times deeper or more than the acetabulum (ch. 187: 0 → 1), and ligament sulcus of the femoral head does not form a medial excavation in proximal view (ch. 204: 0 → 1). Only the latter character is coded for Am. nesbitti. Following the phylogenetic definition proposed by Nesbitt et al.4, the clade supporting Am. nesbitti, I. fragilis and Si. opolensis receives the name Silesauridae.

Results of the analyzes. (a) Time-calibrated reduced strict consensus tree depicting the phylogenetic position of Amanasaurus nesbitti gen. et sp. nov. (b) Left (reversed) femur of Scutellosaurus lawleri (MNA 175) in lateral view. (c) Left (reversed) femur of Eucoelophysis baldwini (NMMNH P-22298) in lateral view. (d) Right femur of Amanasaurus nesbitti gen. et sp. nov. (CAPPA/UFSM 0374) in lateral view. (e) Right femur of Lewisuchus admixtus (CRILAR-Pv 552) in lateral view. (f) Occurrence of Pan-Aves according the Middle to Upper Triassic Assemblage Zones of Brazil; (g) Plot of log10-transformed proximal long axis of the femur versus log10-transformed femoral length of distinct ornithodirans depicting (purple triangle) the estimated femoral length for the holotype of Amanasaurus nesbitti gen. et sp. nov. (h) Plot of log10-transformed distal long axis of the femur versus log10-transformed femoral length of distinct ornithodirans depicting (purple triangle) the estimated femoral length for the referred specimen of Amanasaurus nesbitti gen. et sp. nov. Figures were generated with GIMP 2.8 (https://www.gimp.org/).
Regarding Gamatavus antiquus, it nests as the basalmost member of Sulcimentisauria, an arrangement supported by the absence of the femoral trochanteric shelf (ch. 215: 0 → 1) and facies articularis antitrochanterica not ventrally descended (ch. 216: 0 → 1). Furthermore, Saltopus elginensis nested as sister taxon of Lewisuchus admixtus and is recovered as an ornithischian for the first time. This result should be taken carefully, however, given the fragmentary and difficult-to-interpret nature of the holotype. In previous iterations of this dataset Sa. elginensis was recovered as an early-diverging saurischian sister to Eodromaeus murphi9 and as a non-dinosaur dinosauromorph15.
Discussion
The holotype of Amanasaurus nesbitti possesses typical traits of silesaurs, such as the presence of a notch between the ventral transition from the femoral head to the femoral shaft and a straight medial articular facet of the proximal portion in proximal view4,11,14,28,36. Therefore, Am. nesbitti can be safely assigned to a silesaur. Although the referred specimen lacks overlapping material with the holotype, the preserved distal portion of the femur resembles that of other silesaurs (e.g., presence of an elongated popliteal fossa4) and lacks typical features of other related groups. So, this specimen is referred to Am. nesbitti on the basis of the geological context and the shared morphology with other silesaurs.
Regarding the general morphology of Am. nesbitti, it bears a unique set of plesiomorphic and apomorphic traits for silesaurs, which is consistent with its phylogenetic and stratigraphic position. It retains a fossa trochanterica, a trait presents in older silesaurs, such as Lewisuchus admixtus and Asilisaurus kongwe19,35. Conversely, the posteromedial tuber is extremely reduced, resembling the condition observed in late diverging forms, such as in Sacisaurus agudoensis14 and Kwanasaurus williamparkeri7. Perhaps, the “transitional” shape of the anterior trochanter comprises one of the most interesting features of the new taxon. The anterior trochanter of dinosaurs and close related groups is usually regarded as the insertion point for the m. iliofemoralis40,47. In early silesaurs (e.g., L. admixtus; As. Kongwe; Gamatavus antiquus), the proximal tip is completely connected to the femoral shaft (Fig. 3e), whereas in late diverging forms (e.g., Eucoelophysis baldwini) its tip is completely separated from the shaft by a marked cleft14 (Fig. 3c). This condition is interpreted as an early stage of the “wing-like” anterior trochanter of prionodontians (Fig. 3b) and provided further support for the ornithischian affinities of silesaurs10,14. In Am. nesbitti, the anterior trochanter is less pronounced than in post-Carnian silesaurs; however, it bears the cleft separating the proximal tip from the femoral shaft (Fig. 3d). The new taxon comprises the oldest silesaur expressing this condition, revealing a Carnian origin for this feature.
The new silesaur provides further support for the presence of silesaurs in the Hyperodapedon Assemblage Zone (AZ) of Brazil. These reptiles are reported for three of the four AZs assigned to Middle and Upper Triassic8,17. The current scenario depicts the total absence of silesaurs and other Pan-Aves solely in the Santacruzodon AZ (Fig. 3f), which is poorly sampled in comparison with other AZs and its geographical distribution is limited29. In addition, the presence of the new silesaur in Carnian beds of Southern Brazil reinforces the co-occurrence of distinct Pan-Aves groups during the initial evolution of dinosaurs (ca. 230 Ma). The new silesaur comes from an outcrop area that yielded lagerpetids, early sauropodomorphs, and herrerasaurids15,16,48. This diversity of Pan-Aves surpasses that of older AZs from Brazil (i.e., Dinodontosaurus AZ and Santacruzodon AZ), being comparable to that of the Riograndia AZ (Fig. 3f). A similar diversity is also reported for the coeval Ischigualasto Formation18, where a silesaur is also reported (i.e., Ignotosaurus fragilis). It is reasonable to conclude that the landscapes that witnessed the early evolution of dinosaurs supported a wide range of avian line archosaurs as well. Moreover, according to the femoral length estimations performed here, Am. nesbitti reached the same size of early sauropodomorphs. The estimated femoral length of the holotype (CAPPA/UFSM 0374) is 121 mm (Fig. 3g), whereas for the referred specimen (CAPPA/UFSM 0375) it is 143 mm (Fig. 3h). For comparison, the femoral length of specimens of the early sauropodomorph Buriolestes schultzi ranges from 89 mm (ULBRA-PVT05649) to 138 mm (ULBRA-PVT28049). These specimens were excavated from correlate strata that are 500 m distant from the Pivetta site. It is the first time that silesaurs rivaling in size with early dinosaurs are recovered from the oldest unequivocal dinosaur-bearing beds, challenging the idea that in faunas where silesaurs and unambiguous dinosaurs co-occurred, silesaurs were relatively smaller28. This discovery reinforces the complex scenario regarding the radiation of Pan-Aves during the Triassic. Surely, the body plan of early diverging forms being surpassed by late diverging dinosaurs does not fit within the current models anymore. Actually, silesaurs –independent of their phylogenetic position– persisted during most of the Triassic Period, with its plesiomorphic body size advancing through the dawn of dinosaurs, instead of silesaur lineages decrease in body size through time.
Data availability
All data generated or analysed during this study are included in this published article and its Supplementary Information files.
Change history
22 August 2024
A Correction to this paper has been published: https://doi.org/10.1038/s41598-024-70440-4
References
Brusatte, S. L., Benton, M. J., Lloyd, G. T., Ruta, M. & Wang, S. C. Macroevolutionary patterns in the evolutionary radiation of archosaurs (Tetrapoda: Diapsida). Earth Environ. Sci. Trans. R. Soc. Edinb. 101, 367–382 (2010).
Benton, M. J., Forth, J. & Langer, M. C. Models for the rise of the dinosaurs. Curr. Biol. 24, 87–95 (2014).
Langer, M. C., Nesbitt, S. J., Bittencourt, J. S. & Irmis, R. B. Non-dinosaurian dinosauromorpha. Geolog. Soc. Lond. Spec. Publ. 379, 157–186 (2013).
Nesbitt, S. J. et al. Ecologically distinct dinosaurian sister-group shows early diversification of Ornithodira. Nature 464, 95–98 (2010).
Peecook, B. R. et al. A new silesaurid from the upper Ntawere Formation of Zambia (middle triassic) demonstrates the rapid diversification of Silesauridae (Avemetatarsalia, Dinosauriformes). J. Vertebr. Paleontol. 33, 1127–1137 (2013).
Peecook, B. R., Steyer, J. S., Tabor, N. J. & Smith, R. M. Updated geology and vertebrate paleontology of the Triassic Ntawere Formation of northeastern Zambia, with special emphasis on the archosauromorphs. J. Vertebr. Paleontol. 37(sup1), 8–38 (2017).
Martz, J. W. & Small, B. J. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7, e7551 (2019).
Pretto, F. A. et al. The oldest South American silesaurid: New remains from the Middle Triassic (Pinheiros-Chiniquá Sequence, Dinodontosaurus Assemblage Zone) increase the time range of silesaurid fossil record in southern Brazil. J. S. Am. Earth Sci. 120, 104039 (2022).
Müller, R. T. & Garcia, M. S. A paraphyletic ‘Silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biol. Let. 16, 20200417 (2020).
Norman, D. B., Baron, M. G., Garcia, M. S. & Müller, R. T. Taxonomic, palaeobiological and evolutionary implications of a phylogenetic hypothesis for Ornithischia (Archosauria: Dinosauria). Zool. J. Linn. Soc. 196, 1273–1309 (2022).
Ezcurra, M. D. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan & Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28, 649–684 (2006).
Langer, M. C., Ezcurra, M. D., Bittencourt, J. S. & Novas, F. E. The origin and early evolution of dinosaurs. Biol. Rev. 85, 55–110 (2010).
Novas, F. E. et al. Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications. J. S. Am. Earth Sci. 110, 103341 (2021).
Langer, M. C. & Ferigolo, J. The Late Triassic dinosauromorph Sacisaurus agudoensis (Caturrita Formation; Rio Grande do Sul, Brazil): Anatomy and affinities. Geolog. Soci. Lond. Spec. Publ. 379, 353–392 (2013).
Cabreira, S. F. et al. A unique Late Triassic dinosauromorph assemblage reveals dinosaur ancestral anatomy and diet. Curr. Biol. 26, 3090–3095 (2016).
Pacheco, C. et al. Gnathovorax cabreirai: A new early dinosaur and the origin and initial radiation of predatory dinosaurs. PeerJ 7, e7963 (2019).
Ferigolo, J. & Langer, M. C. A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone. Hist. Biol. 19, 23–33 (2007).
Martínez, R. N. et al. Vertebrate succession in the Ischigualasto formation. J. Vertebr. Paleontol. 32(Suppl. 1), 10–30 (2013).
Ezcurra, M. D., Nesbitt, S. J., Fiorelli, L. E. & Desojo, J. B. New specimen sheds light on the anatomy and taxonomy of the early Late Triassic dinosauriforms from the Chañares Formation, NW Argentina. Anat. Rec. 303, 1393–1438 (2020).
Agnolín, F., BrissónEgli, F., Ezcurra, M. D., Langer, M. C. & Novas, F. New specimens provide insights into the anatomy of the dinosauriform Lewisuchus admixtus Romer, 1972 from the upper Triassic levels of the Chañares Formation, NW Argentina. Anat. Rec. 35, 1119–1146 (2022).
Dzik, J. A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. J. Vertebr. Paleontol. 23, 556–574 (2003).
Sarıgül, V., Agnolin, F. L. & Chatterjee, S. Description of a multitaxic bone assemblage from the Upper Triassic Post Quarry of Texas (Dockum Group), including a new small basal dinosauriform taxon. Hist. Nat. (Corr.) 8, 5–24 (2018).
Romer, A. S. The Chañares (Argentina) Triassic reptile fauna. XIV. Lewisuchus admixtus, gen. et sp. nov., a further thecodont from the Chañares beds. Breviora 390, 1–13 (1972).
Mestriner, G. et al. Histological analysis of ankylothecodonty in Silesauridae (Archosauria: Dinosauriformes) and its implications for the evolution of dinosaur tooth attachment. Anat. Rec. 305, 393–423 (2022).
Garcia, M. S., Müller, R. T., Da-Rosa, Á. A. & Dias-da-Silva, S. The oldest known cooccurrence of dinosaurs and their closest relatives: A new lagerpetid from a Carnian (Upper Triassic) bed of Brazil with implications for dinosauromorph biostratigraphy, early diversification and biogeography. J. S. Am. Earth Sci. 91, 302–319 (2019).
Baron, M. G. The effect of character and outgroup choice on the phylogenetic position of the Jurassic dinosaur Chilesaurus diegosaurezi. Palaeoworld https://doi.org/10.1016/j.palwor.2022.12.001 (2022).
Goloboff, P. A. & Catalano, S. A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32, 221–238 (2016).
Barrett, P. M., Nesbitt, S. J. & Peecook, B. R. A large-bodied silesaurid from the Lifua Member of the Manda beds (Middle Triassic) of Tanzania and its implications for body-size evolution in Dinosauromorpha. Gondwana Res. 27, 925–931 (2015).
Horn, B. L. D. et al. A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. J. S. Am. Earth Sci. 55, 123–132 (2014).
Zerfass, H. et al. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: A contribution to southwestern Gondwana palaeogeography and palaeoclimate. Sediment. Geol. 161, 85–105 (2003).
Langer, M. C., Ribeiro, A. M., Schultz, C. L. & Ferigolo, J. The continental tetrapod-bearing Triassic of south Brazil. N. M. Mus. Nat. Hist. Sci. Bull. 41, 201–218 (2007).
Langer, M. C., Ramezani, J. & Da-Rosa, Á. A. S. Upb age constraints on dinosaur rise from south Brazil. Gondwana Res. 57, 133–140 (2018).
Nesbitt, S. J. The early evolution of archosaurs: Relationships and the origin of major clades. Bull. Am. Mus. Nat. Hist. 352, 1–292 (2011).
Ezcurra, M. D. et al. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature 588, 445–449 (2020).
Nesbitt, S. J., Langer, M. C. & Ezcurra, M. D. The anatomy of Asilisaurus kongwe, a dinosauriform from the Lifua Member of the Manda Beds (~ Middle Triassic) of Africa. Anat. Rec. 303, 813–873 (2020).
Kammerer, C. F., Nesbitt, S. J. & Shubin, N. H. The first silesaurid dinosauriform from the Late Triassic of Morocco. Acta Palaeontol. Pol. 57, 277–284 (2011).
Irmis, R. B. et al. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317, 358–361 (2007).
Nesbitt, S. J. et al. Hindlimb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. J. Vertebr. Paleontol. 29, 498–516 (2009).
Piechowski, R., Tałanda, M. & Dzik, J. Skeletal variation and ontogeny of the Late Triassic Dinosauriform Silesaurus opolensis. J. Vertebr. Paleontol. 34, 1383–1393 (2014).
Piechowski, R. & Tałanda, M. The locomotor musculature and posture of the early dinosauriform Silesaurus opolensis provides a new look into the evolution of Dinosauromorpha. J. Anat. 236, 1044–1100 (2020).
Griffin, C. T. & Nesbitt, S. J. The femoral ontogeny and long bone histology of the Middle Triassic (? late Anisian) dinosauriform Asilisaurus kongwe and implications for the growth of early dinosaurs. J. Vertebr. Paleontol. 36, e1111224 (2016).
Müller, R. T. A new theropod dinosaur from a peculiar Late Triassic assemblage of southern Brazil. J. S. Am. Earth Sci. 107, 103026 (2021).
Galton, P. M. Notes on the postcranial anatomy of the heterodontosaurid dinosaur Heterodontosaurus tucki, a basal ornithischian from the Lower Jurassic of South Africa. Rev. Paléobiol. 33, 97141 (2014).
Baron, M. G., Norman, D. B. & Barrett, P. M. Postcranial anatomy of Lesothosaurus diagnosticus (Dinosauria: Ornithischia) from the Lower Jurassic of southern Africa: Implications for basal ornithischian taxonomy and systematics. Zool. J. Linn. Soc. 179, 125–168 (2017).
Marsh, A. D. & Rowe, T. B. A comprehensive anatomical and phylogenetic evaluation of Dilophosaurus wetherilli (Dinosauria, Theropoda) with descriptions of new specimens from the Kayenta Formation of northern Arizona. J. Paleontol. 94, 1–103 (2020).
Nesbitt, S. J. et al. The earliest bird-line archosaurs and the assembly of the dinosaur body plan. Nature 544, 484–487 (2017).
Ballell, A., Rayfield, E. J. & Benton, M. J. Walking with early dinosaurs: Appendicular myology of the Late Triassic sauropodomorph Thecodontosaurus antiquus. R. Soc. Open Sci. 9, 211356 (2022).
Müller, R. T. et al. Early evolution of sauropodomorphs: Anatomy and phylogenetic relationships of a remarkably well-preserved dinosaur from the Upper Triassic of southern Brazil. Zool. J. Linn. Soc. 184, 1187–1248 (2018).
Müller, R. T. On the presence and shape of anterolateral scars in the ontogenetic series of femora for two early sauropodomorph dinosaurs from the Upper Triassic of Brazil. Paleontol. Res. 26, 1–7 (2022).
Acknowledgements
We thank Arthur S. Brum and an anonymous reviewer for comments and suggestions that greatly improved this manuscript. We also extend our gratitude to the editor Dawid Surmik for assistance and discussions. We thank the Willi Henning Society, for the gratuity of TNT software. This work was carried out with aid of the Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS 21/2551-0000680-3) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 404095/2021-6 and 303034/2022-0) to Rodrigo Temp Müller and a Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES 88887.608076/2021-00) scholarship for Mauricio Silva Garcia.
Author information
Authors and Affiliations
Contributions
R.T.M. data curation, funding acquisition, formal analysis, investigation, writing. M.S.G. formal analysis, investigation, writing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Müller, R.T., Garcia, M.S. A new silesaurid from Carnian beds of Brazil fills a gap in the radiation of avian line archosaurs. Sci Rep 13, 4981 (2023). https://doi.org/10.1038/s41598-023-32057-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-32057-x
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
