
Abstract
Medieval southern Italy is typically viewed as a region where political, religious, and cultural systems coexisted and clashed. Written sources often focus on elites and give an image of a hierarchical feudal society supported by a farming economy. We undertook an interdisciplinary study combining historical and archaeological evidence with Bayesian modelling of multi-isotope data from human (n = 134) and faunal (n = 21) skeletal remains to inform on the socioeconomic organisation, cultural practices, and demographics of medieval communities in Capitanata (southern Italy). Isotopic results show significant dietary differences within local populations supportive of marked socioeconomic hierarchies. Bayesian dietary modelling suggested that cereal production, followed by animal management practices, was the economic basis of the region. However, minor consumption of marine fish, potentially associated with Christian practices, revealed intra-regional trade. At the site of Tertiveri, isotope-based clustering and Bayesian spatial modelling identified migrant individuals likely from the Alpine region plus one Muslim individual from the Mediterranean coastline. Our results align with the prevailing image of Medieval southern Italy but they also showcase how Bayesian methods and multi-isotope data can be used to directly inform on the history of local communities and of the legacy that these left.
Similar content being viewed by others

Introduction
The Middle Ages in Italy has been argued to have been marked by political and cultural fragmentation from the collapse of the western Roman empire (476 CE) until the end of the twelfth century, although other scholars note a persistence and reconfiguration of economic and urban networks1,2,3,4. Southern Italy represents a particularly interesting case of how populations in this part of the world responded to the collapse of the western Roman empire and transitioned into the new economic, social and cultural world of the medieval period2,4,5. Showing the strikingly high level of political fragmentation of the region, unlanded aristocrats from Normandy (France) famously sought their fortune in southern Italy during the eleventh century, intervening in local disputes among Lombard princes, Byzantine rulers, the Pope and Muslim pirates4,6,7. Through this, Normans accumulated wealth, land and titles culminating in the establishment of the Kingdom of Sicily in 1130 by the Hauteville family. The Normans imposed a feudal system based on their French experience, although it also included political traits from previous local Byzantine and Muslim forms of government. The Hauteville family was succeeded by the Swabian Hohenstaufen dynasty in 1194, which lasted until 1266, when Charles count of Anjou conquered the kingdom with papal support. While much is known historically about these political upheavals, far less is known in relation to the lived experiences of the multicultural populations navigating this changing social and economic landscape that was to reshape power dynamics in the Mediterranean world. Sites from southern Italy hence provide ideal case studies for exploring how changing political regimes and contact with areas of the eastern Roman empire, Mediterranean islands, and northern Africa altered economic systems and the demographics of local populations2,3,4,8,9,10. Furthermore, they were sites of increasing social differentiation during the medieval period that increasingly revolved around connection to the Christian faith and land ownership7,10,11.
Medieval Capitanata—a region in northern Apulia—has a centuries-long history as a buffer zone between Lombard states and Byzantine Apulia. However, the Norman conquest of the region in the mid-eleventh century led to its pacification and to wide economic transformations12,13,14. This pushed towards a demographic increase and to new food production systems adapted to local geographic features (e.g. fishing in coastal sites, pig husbandry in the plain and sheep/goat husbandry in the Apennines)12. Socio-economic shifts have been inferred through the analysis of the archaeological record, zooarchaeological studies, and limited archaeobotanical evidence on the region12,13,15,16. However, there is little anthropological17 and no biomolecular evidence for late medieval Capitanata. Previous archaeological isotopic studies in the region investigated only the Neolithic period18,19,20. The closest isotopic studies dating to the medieval period are those from Montella (Campania), Apigliano, and Quattro Macine (southern Apulia)21,22.
Overall, the Norman productive system was maintained for the entirety of the late medieval period, with a successive increase of agricultural production and the introduction of ‘masserie’12,15. These were large land estates often under direct administration of the crown. However, in the thirteenth century this region also observed the arrival of a melting-pot of cultures from both Mediterranean and continental regions. This is, for example, the case of Lucera (Fig. 1), a city that hosted the Islamic population that was deported from Sicily by order of the Hohenstaufen king Frederik II4,23,24,25. Tertiveri, one of the sites analysed in this study (Fig. 1), was also donated by the Angevin king Charles II to ‘Abd al- ‘Azīz, a powerful Muslim knight from Lucera23,24. Two individuals found in this site (here sampled and analysed) were buried according to an Islamic rite. Frederik II was also Holy Roman Emperor, but he kept his court in southern Italy and spent most of his time in Apulia25. Hence, diplomats, noblemen, and ecclesiastic personalities were often supposed to travel from northern Italy, Austria, and Germany to Capitanata. Additional evidence of the high-level of cultural admixture involves movements from Provence (France) during the late thirteenth century. Written records attest that the new Angevin king Charles I relocated soldiers to the turbulent Capitanata region to ensure its pacification26. This was followed by donations of lands and the settling of many Provençal individuals with their families20. Thus, late medieval Capitanata presents a high research potential to investigate the impact of migrations and economic systems on past societies.

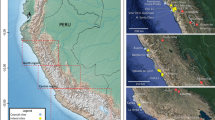
Spatial distribution of medieval sites from the historical region of Capitanata (border division in black) and southern Italy mentioned in this study: (A) Tertiveri; (B) Montecorvino; (C) San Lorenzo in Carminiano; (D) Apigliano; (E) Quattro Macine; (F) Franciscan Friary in Montella; (G) Crepacore Castle; (H) Lucera. With a black font, sites for which new isotopic data is presented. This map was generated using the Interactive map option in IsoMemoApp (https://isomemoapp.com/app/iso-memo-app, Data Search and Spatiotemporal modelling V. 22.12.3.1).
Much of our historical knowledge of medieval southern Italy relies on written sources which often are not representative of a large proportion of past communities, particularly for population segments with a lower socioeconomic status27. Stable isotope analysis, by contrast, has been widely employed in archaeology to reconstruct past human subsistence or spatial mobility, and crop and animal management practices21,22,28,29,30,31. Isotopic ratios can be measured in osteological remains and, depending on the type of tissue analysed (e.g. collagen extracted from bones), information can be obtained for different periods of an individual’s lifetime32,33,34. The most common isotopic proxies employed in archaeological dietary studies are carbon (δ13C) and nitrogen (δ15N) isotopes measured in collagen extracted from bone or dentine. These are primarily informative on dietary protein consumption35,36, whereas δ13C measurements on carbonate from bone or tooth bioapatite inform on the dietary mix of protein, carbohydrates, and lipids. Measurement of the latter will more easily reveal the consumption of C4 plants when the primary source of protein is C3 (or vice-versa). Local environmental and climatic conditions determine the isotopic values of plant and animal food sources37,38. These can also be influenced by agricultural management practices such as irrigation, manuring, penning, etc.36,39,40. Thus, human isotopic values reflect the isotopic values of the food sources and their relative consumption. Human and animal spatial mobility can be investigated using δ18O measurements on carbonate from tooth enamel or bone bioapatite, taking into account potential complications caused by cooking activities, diagenesis and mathematical conversions of human δ18O values into δ18O of ingested water41,42,43,44.
We combined historical and archaeological evidence with novel Bayesian modelling of multi-isotope data (δ13C, δ15N, δ18O) from human (n = 134) and animal (n = 21) skeletal remains to inform on the socioeconomic organisation, cultural practices, and demographics of the medieval communities of Tertiveri, Montecorvino, and San Lorenzo in Carminiano (Capitanata, southern Italy). We were specifically interested in evaluating the implementation of a feudal system at the study sites. Individual isotopic variability was employed as a proxy for socioeconomic hierarchy and/or Christian religious practices. While overall population diets informed on the economic importance of farming and trade activities. The significance of external influences was investigated isotopically through the identification of migrants and the tracking of their places of origin.
Results
Isotopic results, together with other supporting measurement, archaeological, and historical information are given in Supplementary Information file S1 and also deposited in the MATILDA online repository (https://www.doi.org/10.48493/w01v-fe90). The data was also aggregated to the CIMA database that compiles isotopic measurements for medieval archaeological samples45,46.
Bone and tooth samples (human = 134; fauna = 21) areoverall well-preserved in accordance with established parameters for bone collagen preservation47. However, fifteen human samples from Tertiveri show poor preservation with atomic C/N ratios outside the acceptable range. In addition, sample TC100 did not produce sufficient collagen for measurement. These represent 11.9% of the human dataset. Bone carbonate results for samples with bad collagen preservation were also excluded from our analysis48.
Faunal isotopic results
Faunal bulk bone and tooth collagen δ13C and δ15N measurements range from −24.1 to −19.8‰ and from 4.7 to 10.1‰, respectively (Fig. 2a). For δ13C, there is a significant difference between the Montecorvino and Tertiveri animals (mean δ13C: −21.9 ± 0.9‰; −20.7 ± 1.0‰). The sample size is small making the interpretation of the results difficult, but the difference is likely associated with differences in local environmental conditions and/or animal management practices, including a possible ‘canopy effect’ associated with dense forests surrounding Montecorvino40,49. These are mentioned in Angevin documents50 and also attested by palaeo-environmental analysis51. For δ15N values there are no significant differences between the two sites (mean: 6.7 ± 1.9‰ for Montecorvino; 6.8 ± 1.1‰ for Tertiveri). However, the standard deviations for Montecorvino pigs and sheep/goats are relatively large (7.0 ± 1.5‰; 5.8 ± 2.3‰, respectively) when compared to δ15N values measured in the same taxa from two other late medieval sites in southern Apulia, i.e. Apigliano (pigs: 5.3 ± 0.4‰; sheep/goat: 6.1 ± 0.4‰) and Quattro Macine (pigs: 5.8 ± 0.7‰; sheep/goat: 5.4 ± 0.8‰)23 (Fig. 2b). Overall, domestic herbivores from Capitanata display larger δ15N ranges (4.0–8.9‰; mean: 6.2 ± 1.7‰) in comparison to those from southern Apulia (4.1–6.9‰; mean: 5.5 ± 0.9‰). These likely reflect a variety of animal husbandry strategies including intensive husbandry, penning, feeding with plants grown in fields subject to manuring, and transhumance39,40,52.

Isotopic results for archaeological faunal samples. (a) δ13C versus δ15N for fauna from this study; (b) δ13C versus δ15N comparing mean isotopic values for herbivores and omnivores from this study versus those measured in the coeval Apulian sites of Apigliano and Quattro Macine22; (c) δ13Ccollagen versus δ13Ccarbonate for fauna from this study.
The deer remains from San Lorenzo (SLDE1) have a high δ15N collagen value (bone: 7.4‰; tooth: 7.3‰. Figure 2a) when compared to values observed for modern specimens53. Given historical evidence for a royal hunting ground around San Lorenzo in Carminiano54, it is possible that deers had their diet managed by humans, which may have included access to 15N-enriched plants. The bone remains of two tortoises from Tertiveri (δ13C: −22.4‰; δ15N: 6.0‰) and Montecorvino (δ13C: −24.1‰; δ15N: 4.9‰) have low δ13C values suggesting that they fed on freshwater sources55. Tortoises were commonly consumed in medieval Italy and zooarchaeological remains are often found in sites associated with the clergy, since they were considered to be ‘fish’ and not included in the list of proscribed foods by medieval religious fasting rules9,56.
Results of stable carbon isotopes for bone bioapatite in animals range between −14.1‰ and −8.3‰ (mean: −12.1 ± 1.5‰) (Fig. 2c). The highest values for animals are observed for a chicken from Montecorvino (MOCH2 δ13Ccarbonate: −9.1‰; δ13Ccollagen: −21.7‰; δ15N: 10.2‰) and a cattle specimen from Tertiveri (TCCA2 δ13Ccarbonate: −8.3‰; δ13Ccollagen: −20.2‰; δ15N: 8.3‰), which were likely fed with a higher contribution of C4 cereals compared to other animals. However, δ13Ccollagen values from these animals clearly reflect C3 dominated diets. In the case of the chicken (MOCH2), the isotopic difference between δ13Cbioapatite and δ13Ccollagen signals could derive from the combination of a predominant C3 protein source and an overall caloric contribution that included significant C4 components. Such an explanation is more difficult to extend to the cattle specimen (TCCA2), since this animal would only consume plants. A mixed diet of legumes and C3/C4 plants seems unlikely given that the animal has relatively high nitrogen values. Even assuming that this cattle specimen consumed plants from manured fields, which elevates plant δ15N values, a predominant protein source from legumes (typically near 0‰ δ15N values) would be inconsistent with our measurements. Another possibility is that the bioapatite and collagen δ13C signals reflect different lifetime dietary stages. This would imply that bone collagen and bioapatite have different renewal rates as reported previously for other species57. However, these are unknown.
Values for bioapatite and enamel δ18O in fauna from Tertiveri and Montecorvino show a wide range (measured δ18Ocarbonate range: −6.9‰ to −1.6‰; mean: −4.4 ± 1.3‰; calculated δ18Owater range: −10.4‰ to −1.7‰; mean: −6.3 ± 2.1‰) (Fig. 3a). The age and time of death of the animals or the renewal rates of skeletal material are not known. Thus, some variation may be explained by seasonal shifts in water δ18O values. For instance, pigs were commonly slaughtered at the end of the year, whereas sheep/goats were butchered around Easter in accordance with religious practices9,56. Spatial variations in water δ18O values may have also had an impact, since vertical transhumance of ovicaprids and cattle has been recorded for late medieval Capitanata58,59. Although there is a large overlap between the δ18O values for domesticated herbivores and omnivores (Fig. 3b).

Calculated values for ingested water δ18OWater for humans and fauna. (a) histogram showing the distribution of human and faunal δ18OWater values; (b) histogram showing the distribution of domesticated herbivore and omnivore δ18OWater values.
One sheep/goat bone from Montecorvino (MOSG1) shows an unusually high δ18Owater (−1.7‰) value. In addition to spatial and temporal variations in water δ18O values it is possible that this sheep was consuming leaf water with δ18O values higher than meteoritic water60. Ovicaprids are semi-obligate drinkers and obtain most of the water through the plants they consume61. Other sheep/goats from Montecorvino have δ18Owater values between −6.1 and −4.0‰. However, one sheep/goat from Tertiveri (TCSG1) has the lowest δ18Owater value (−7.6‰) among ovicaprids. Multiple factors could explain such variability. Differences in feeding behaviours, including the aforementioned consumption of leaf water, the practice of transhumance at various spatial and temporal scales, plus differences in killing season could explain the wide observed range in δ18Owater values. To resolve this, it is necessary to study animal movements and diets at higher temporal resolutions by undertaking future isotopic measurements on tooth sections61,62.
Human isotopic results and Bayesian modelling
Results for stable carbon and nitrogen isotopic values measured in human bone collagen from the region of Capitanata range from −20.7 to −17.4‰ and from 7.5 to 13.8‰, respectively (Fig. 4a). Mean values are −19.0 ± 0.7‰ for δ13C and 10.0 ± 1.0‰ for δ15N. Human δ13C results measured in bone carbonate range from −14.6 to −5.2‰ (mean: −12.3 ± 1.2‰) (Fig. 4b). Isotopic results for non-adults are presented separately from those of adolescents and adults. Non-adult individuals may include a dietary isotopic signal from the consumption from human milk and/or from food employed during infancy in post-weaning strategies63,64. Figure 4c demonstrates the distribution of δ13C and δ15N values for non-adults in Tertiveri (mean δ13Ccollagen: −18.5 ± 0.8‰; mean δ15N: 10.5 ± 1.5‰; δ13Ccarbonate: −11.9 ± 1.4‰) and Montecorvino (mean δ13Ccollagen: −20.1 ± 0.4‰; mean δ15N: 10.2 ± 1.2‰; δ13Ccarbonate: −12.2 ± 1.0‰). There is no significant difference among age groups for non-adults (Fig. 4c), although neonates present higher δ15N values, which is likely a result of breastfeeding. From San Lorenzo only four adult individuals were analysed (mean δ13Ccollagen: −19.8 ± 0.3‰; mean δ15N: 9.0 ± 0.5‰; δ13Ccarbonate: −12.4 ± 0.4‰), while from Tertiveri 78 adults present an acceptable level of bone preservation (mean δ13Ccollagen: −18.8 ± 0.5‰; mean δ15N: 9.9 ± 0.9‰; δ13Ccarbonate: −12.4 ± 1.3‰) and from Montecorvino tenadults were analysed (mean δ13Ccollagen: −20.0 ± 0.4‰; mean δ15N: 10.2 ± 0.5‰; δ13Ccarbonate: −12.7 ± 0.7‰).

Human isotopic results. (a) δ13Ccollagen versus δ15Ncollagen values for human individuals from this study; (b) δ13Ccollagen versus δ13Ccarbonate values for human individuals from this study (reference dietary lines after Kellner and Schoeninger)65. (c) δ13Ccollagen versus δ15Ncollagen values for human non-adult individuals from this study. Neonates: 0–1 y.; Infants: 1–3 y.; Young Juveniles: 3–7 y.; Old Juveniles: 7–13 y. (d) δ13Ccollagen versus δ15Ncollagen biplot comparing mean isotopic values for adults from this study with those measured in the coeval southern Italian sites of Montella, Apigliano, and Quattro Macine (Torino et al. 2015; Rolandsen et al. 2019)21,22.
The δ13Ccollagen values for Tertiveri are significantly higher than Montecorvino (Mann–Whitney U test: U = 26, n1 = 78, n2 = 10, P < 0.05, two-tailed) and this may be related to differences in the isotopic baseline (e.g. environmental factors such as the ‘canopy effect’ or specific animal management practices such as C3/C4 feeding selection) or to differences in human diets. No significant stable carbon and nitrogen isotopic differences between adult male (mean δ13C: −18.7 ± 0.4‰; mean δ15N: 10.0 ± 0.9‰) and female (mean δ13C: −18.8 ± 0.7‰; mean δ15N: 10.0 ± 1.2‰) individuals were detected at Tertiveri (δ13C: Mann–Whitney U test: U = 800.5, n1 = 36, n2 = 17, P > 0.05, two-tailed; δ15N: Mann–Whitney U test: U = 807.5, n1 = 36, n2 = 17, P > 0.05, two-tailed), whereas at Montecorvino and San Lorenzo the small sample size does not allow for comparisons. When compared with previously published studies for medieval southern Italy, adult individuals from Capitanata show overall higher δ15N values than Montella (mean δ13C: −19.7 ± 0.7‰; mean δ15N: 7.1 ± 1.1‰), Apigliano (mean δ13C: −18.8 ± 0.3‰; mean δ15N: 7.9 ± 0.7‰), and Quattro Macine (mean δ13C: −18.9 ± 0.2‰; mean δ15N: 7.6 ± 0.4‰) (Fig. 4d)21,22. The δ15N values measured in faunal specimens from Capitanata were also higher (Fig. 2b), implying that the isotopic baseline, rather than diet, is the primary factor that accounts for the isotopic difference in adult populations from different sites.
The results for δ13Ccarbonate versus δ13Ccollagen are plotted in Fig. 4b and compared to C3 protein and C4/marine protein reference lines after Kellner and Schoeninger65. This shows that C3 foods were the main protein source although there were minor contributions from C4/marine protein foods for several individuals. The contribution from C4 carbohydrates is also noticeable for several individuals having relatively high δ13Ccarbonate values. One individual (TC65) has particularly high δ13Ccarbonate values (δ13Ccarbonate: −5.2‰; δ13Ccollagen: −18.4‰; δ15N: 10.8‰) suggesting a large consumption of C4 cereals. The source of protein for this individual was predominantly C3 although there could have been a minor contribution from marine resources.
Cluster analysis of adult human δ13Ccollagen and δ15Ncollagen values from Tertiveri revealed two main data clusters (details in "Methods" section) (Fig. 5a). Cluster 1 shows a negative correlation between δ13Ccollagen and δ15Ncollagen values suggesting a mixing line from C3 to C4 protein consumption. In contrast, Cluster 2 shows a roughly positive correlation between δ13Ccollagen and δ15Ncollagen values suggesting a mixing line from C3 protein (lower isotopic values) to marine protein (higher isotopic values) consumption. The former would include C3 animal protein with δ15N values higher than C4 plants (e.g. millet).

Distribution of δ13Ccollagen and δ15Ncollagen values for human adult individuals from Tertiveri. (a) Cluster classification; (b) Archaeological classification.
A different classification of human δ13Ccollagen and δ15Ncollagen values is also offered by archaeological descriptions. This is shown in Fig. 5b (burial locations are outlined in Fig. 6 and more details are given in Supplementary Info File S2). A privileged status is inferred for adult individuals buried in shafts discovered outside the Cathedral and parallel to the lateral walls (n = 41; mean δ13Ccollagen: −18.9 ± 0.4‰; mean δ15N: 10.1 ± 0.8‰; δ13Ccarbonate: −12.5 ± 1.4‰). A group of individuals were also recovered outside the remains of a second church reflecting a wider social stratification (n = 30; mean δ13Ccollagen: −18.6 ± 0.4‰; mean δ15N: 9.7 ± 1.0‰; δ13Ccarbonate: −12.5 ± 0.9‰). Finally, individuals were also discovered in the so-called ‘ordinary’ cemetery around the Cathedral, but only the collagen extracted from seven individuals were sufficiently well preserved for analysis (mean δ13Ccollagen: −19.4 ± 0.8‰; mean δ15N: 9.4 ± 0.5‰; δ13Ccarbonate: −10.9 ± 0.7‰). When the two classifications (i.e. isotopic and archaeological) are compared, we notice that: for Cluster 1 (n = 15) 20% of its individuals are from privileged shafts; 53% from outside a second church; and 27% from the ‘ordinary’ cemetery located outside the Cathedral. Whereas these percentages for Cluster 2 (n = 63) are 60%, 35%, and 5%, respectively. Overall, results suggest a higher consumption of marine resources by higher status individuals whereas lower status individuals would consume a higher proportion of C4 plants, likely millet.

Schematic representation of the relative positions of burial locations at Tertiveri. The figure is not to scale.
For dietary quantitative estimation, we employed the Bayesian mixing model ReSources (an updated version of FRUITS66, details on model settings can be found in Supplementary Information File S3). Seven main food groups were considered for dietary reconstruction (C3 plants, C4 cereals, cattle, ovicaprid, pigs, poultry, and marine resources). Additional Bayesian priors were included as constraints to improve dietary resolution. These were grounded on evidence obtained from archaeofaunal, archaeobotanical, and ethnographic studies plus from written records (Supplementary Information File S3). However, a relatively minor contribution from other resources (e.g. freshwater fish) cannot be excluded.
Bayesian dietary caloric estimates are shown in figure Fig. 7. The magnitude of the uncertainties for these do not allow us to identify clear dietary differences among the sites. Nonetheless, the modelling shows that C3 plants (e.g. wheat, or barley, fruit, vegetables, legumes, and nuts) were the main source of calories for late medieval Capitanata. This is followed by pig products and by ovicaprid products (e.g. mutton, milk, cheese). Consumption of C4 cereals is possible for all sites although the wide credible ranges allow for a near zero dietary contribution. A small contribution from marine fish intake is possible which would indicate trade with coastal settlements. Cattle and poultry products are also estimated to be consumed in very small amounts.

Bayesian estimates of dietary caloric contributions (boxes and whiskers represent 68% and 95% credible ranges, respectively. Horizontal continuous lines represent the mean and dashed horizontal lines the median). Numbers in brackets represent 95% credible ranges. Also included within the graph are numeric estimates of the caloric contributions of protein versus. carbohydrates macronutrients. Estimate for: (a) Montecorvino; (b) San Lorenzo in Carminiano; (c) Tertiveri Cluster 1; (d) Tertiveri Cluster 2.
Human δ18Ocarbonate values range from −13 to −3.2‰ (mean: −5.8 ± 1.6‰) (calculated δ18Owater range: −16.5 to −5.3; mean: −8.4 ± 2.2‰, Fig. 3a). Given the potential for a nursing signal in infants, in the following discussion we only consider adult individuals from Tertiveri and Montecorvino which given their proximity would likely have similar δ18Owater values. For San Lorenzo only four individuals were available for analyses and the site is located at a greater distance from Tertiveri and Montecorvino.
Cluster classification with outlier detection (threshold 5%) was employed to classify the δ18O values for Tertiveri and Montecorvino adult individuals (see "Methods" section) (Fig. 8). Two clusters and four outliers were identified. A total of 57 individuals were assigned to Cluster A (δ18Owater range: −9.8‰ to −6‰; mean: −7.7 ± 0.8‰), whereas five individuals were assigned to Cluster B (δ18Owater range: −14.2‰ to −11.8‰; mean: −12.9 ± 0.9‰). The distribution of calculated human δ18Owater values was compared to local water estimates based on the point values from the Regionalized Cluster-based Water Isotope Prediction model (RCWIP)67. Using these, a Bayesian reference baseline for δ18OWater values across western Europe was generated and the local average for Tertiveri and Montecorvino was estimated (Fig. 9a). This value ( −7.1‰ for a standard error of the mean of 0.3‰) is close to the δ18Owater average for Cluster A which is assumed to represent individuals residing in Tertiveri or Montecorvino. In contrast Cluster B represents incoming individuals. The overall variability for each cluster can be explained by post-mortem alterations to in vivo bone δ18O values due to diagenesis or to changes to local water δ18O values due to cooking/brewing practices41,42,44.

Cluster classification and outlier detection for the distribution of δ18O values for adult individuals from Tertiveri and Montecorvino. Solid green and red lines represent cluster mean values, whereas green and red pointed lines represent a 2-sigma interval. Continuous vertical black line represents the mean δ18OWater value for Tertiveri and Montercorvino ( −7.1‰) determined using Bayesian modelling of RCWIP estimates while dashed vertical black lines represent the standard error of the mean for the estimates (0.3‰) (Terzer et al. 2013)67.

Residence probability maps assigned using the model LocateR. (a) Bayesian modelled δ18Owater isotopic baseline based on RCWIP model. (b) Cluster A (mean value); (c) Cluster B (mean value); (d) TC77; (e) TC82; (f) TC20; (g) TC8. Maps were generated using AverageR and LocateR modelling options from IsoMemoApp (https://isomemoapp.com/app/iso-memo-app, Data Search and Spatiotemporal modelling V. 22.12.3.1). More details in the "Methods" section.
In addition to the above mentioned factors influencing ingested water and post-mortem bone δ18O values it is necessary to note that the bone bioapatite is undergoing renewal during the lifetime of an individual. Thus, the estimated human δ18OWater values are potentially the result of mixed contributions from water sources having different δ18OWater values. The outlier TC20 is placed between clusters A and B and could represent an individual originating from a location outside Tertiveri that moved and resided there for a considerable period of time. It is also possible that clusters A and B themselves include individuals with mixed δ18O values. In the case of cluster A this is another variable to consider on why the average for this cluster is slightly lower than the estimated local RCWIP value.
The model LocateR68 was employed to assign a spatial probability distribution for the place of residence associated with individuals from clusters A and B plus the four outliers. The model compares estimated human δ18OWater values with the spatial distribution of RCWIP values (Fig. 9a). As expected for Cluster A there is a high residence probability assigned to the Tertiveri/Montecorvino region (Fig. 9b) whereas for Cluster B highest probabilities are observed for the alpine region (Fig. 9c).
The four outliers correspond to individuals buried at Tertiveri. Individual TC77 ( −16.4‰) and TC82 ( −16.5‰) have the lowest δ18OWater values within the dataset and high residence probabilities assigned to the Alpine region (Fig. 9d,e). These individuals likely died within a short time of their arrival at Tertiveri. Individual TC20 was also assigned a high probability of residence in the Alpine region although high probabilities are also observed for eastern European regions (Fig. 9f). However, as mentioned above this individual is likely to have a strong mixed signal making the interpretation of LocateR results difficult. Individuals TC77, TC82, and TC20 were all found within the same shaft burial assumed to belong to privileged individuals. The fourth outlier, TC8 ( −5.3‰), corresponds to an adult male individual buried in accordance with an Islamic burial rite. His death likely took place in 1296–1300 CE, when Tertiveri was controlled by a Muslim knight23,69,70. For this individual LocateR shows high residence probabilities for the Mediterranean coastline, including the southern tips of the Italian peninsula and Sicily (Fig. 9g).
Discussion
The main source of calories for humans across the three sites was C3 plants, a food group consisting of plants such as wheat, barley, rye, oat, fruit, nuts, vegetables, and legumes. Historical evidence suggests that these foods were the staple diet for southern Italian populations during the Middle Ages9. In Capitanata, the presence of multiple grain storage buildings (e.g. silos) marks the importance of cereal production as a main economic activity during the late Middle Ages71. However, these sources do not mention C4 cereals as a significant crop. In contrast, our isotope-based estimates revealed the potential intake of a moderate amount of C4 plants across the different sites. This is clearer for Tertiveri where individuals from Cluster 1 are in a mixing line between C3 and C4 protein (Fig. 5). The earliest consumption of C4 cereals in southern Italy (likely millet) was isotopically detected for the Bronze Age72, although C3 plants still dominated diets. A useful characteristic of C4 cereals is that these overall ripen faster than C3 cereals and are more durable73. Millet and sorghum are grown during the spring/early summer periods in southern Italy. This is when transhumance practices from Apulia to the Abruzzo Apennines took place58,59 and so it is possible that the cultivation of C4 crops in Capitanata may have been part of a transhumance economy, whereby shepherds consumed these cereals during their summer journeys across the Apennines.
Bayesian caloric estimates show that pigs were likely the main dietary source of animal products. This is unsurprising, given the accessible prices for pig products during the late Middle Ages in comparison to other animal meats9,74. A pig-based farming economy is often observed in association with demographic growth (e.g. imperial Rome75). Pig husbandry, paired with cereal production and transhumance, became the basis for the farming economy of late medieval Capitanata76. This matches the demographic growth following the onset of Norman rule which would require more intensive agricultural production13.
During the late medieval period there was an increase in transhumance in the region59. Tertiveri and Montecorvino were located in the vicinity of transhumance routes and as a likely result of more intense animal traffic the local consumption of sheep/goat products increased. Bayesian estimates confirm a significant dietary consumption of sheep/goat products, either as meat (mutton) or dairy products, whereas estimates for cattle consumption were small. However, it is known from historical sources that cattle transhumance was also practiced58. During the late medieval period, cattle were usually used for ploughing, whereas their meat had a high market value and was therefore mostly consumed from old animals which could no longer be employed in the fields9,15,74. Our modelling results also indicate that poultry meat and eggs consumption was overall probably small.
The caloric contribution from marine foods was small across the different sites. However, given the distance for investigated sites and the coast, this suggests the presence of intra-regional trade networks connecting Adriatic sites with the Apennines. In Tertiveri, individuals from Cluster 2 (Fig. 5a) had bone collagen isotopic values along a rough mixing line between C3 and marine protein. These contrast with individuals from Cluster 1 where the consumption of C4 cereals was higher although Bayesian estimates do not offer very precise estimates. What is clear is that there is a predominance of high socioeconomic individuals in Cluster 2, in contrast to Cluster 1. Millet was considered a low status food during antiquity while the fish imports were likely to be expensive and more easily accessible to individuals with a higher socioeconomic status8,9.
Included in Cluster 2 is the so-called Tertiveri bishop (TC74)70 showing moderately elevated isotopic values. The consumption of marine foods during the medieval period is often associated with the clergy, given that these represented one of the wealthier strata of society and were more likely to adhere to Catholic fasting rules56. Such rules varied with time, region and monastic order, but in the extreme could imply that terrestrial meat could not be consumed for up to nearly half of the year56. Whilst the so-called bishop does not have the highest isotopic values within the cluster, it is possible that the age of the individual (c. 70–80 years old) may have influenced dietary practices. Moreover, old people were often dispensed from religious fasting56.
Two Muslim individuals (TC8 and TC21) were also found in Tertiveri and show bone collagen isotopic values that do not suggest the consumption of marine foods. Islamic fasting rules forbid the consumption of pork, which within our dataset has distinct isotopic values relatively to the other terrestrial food sources. Individual TC21 is an infant (0–3 years) and their isotopic values likely include a nursing effect63. Individual TC8 is an adult with a diet mainly consisting of cereals (mostly C3 but with a moderate contribution from C4), legumes and ovicaprid products. It should be noticed, however, that from the observed bone collagen isotopic values for TC8 it is not possible to fully exclude pork consumption.
Previous isotopic results for late medieval southern Italy are only available for three sites, i.e. Apigliano and Quattro Macine in southern Apulia22 and the Franciscan monastery of Montella in the Campanian apennines21. The δ15N bone collagen values for the adult population from these sites are considerably lower than those presented in our study. This could be the result of dietary differences and/or differences in food values as a result of varying environmental conditions and farming practices36,37,38,39,40. Manuring can elevate the δ15N value of crops39. As mentioned above, the Capitanata region had an economy based on cereal cultivation plus the zooarchaeological record shows the presence of ovicaprids, cattle, and pigs making the practice of manuring possible. There are no direct isotopic measurements on plants for the studied region or from previous publications. However, herbivores from Capitanata show elevated δ15N values when compared to those from Apigliano and Quattro Macine (the site of Montella had no faunal remains). The same applies to pigs. Similar regional disparities in agricultural practices and environmental conditions were observed in the rest of Italy, Iberia, and England45. This is associated with a high degree of economic and/or political fragmentation in Europe during the Middle Ages.
The distribution of human stable oxygen isotope values confirms the rich historical and cultural heritage of the sites of Tertiveri and Montecorvino. Our study identified buried individuals potentially originating from the Alpine region and from other Mediterranean locations. However, spatial mobility may have also taken place at smaller geographical scales. In this respect, transhumance activities, mentioned above, would require seasonal movements that are not detectable from bulk isotopic measurements due to a lack of temporal resolution61. Late medieval written sources, mainly originating from legislative and administrative records, provide detailed information on regulations applied to transhumance activities, calendar dates for the practice, and rent land prices in Capitanata58. Documents indicate that during the cold season (September–May), herds from the Abruzzo Apennines were grazed in the Tavoliere plain in Capitanata.
Migratory movements from the Alps to Capitanata during the late thirteenth century are described in written records26. These consisted of military expeditions and the settlement of veterans in the region following the Angevin conquest of the Kingdom of Sicily (1266)4. Documents attest that Angevin soldiers were enrolled from Bourgogne and Savoy (south-east France) and detached to the castle of Crepacore in 126926. Once the region was pacified, these soldiers obtained lands nearby Crepacore and likely settled there with their families. A Franco-Provençal dialect similar to those from the French-Swiss border is spoken still today in two communities located less than 15 km from Tertiveri26. Most individuals from Cluster B (C92, TC98, TC99, and TC101) were buried in the so-called ‘ordinary’ cemetery outside Tertiveri Cathedral. There are no clear indications that these were high-status individuals (two were assigned to dietary Cluster 1 and two to Cluster 2 and they could be Angevin soldiers or their family members).
There is no precise chronology for the individuals mentioned above but it is known that this differs from other individuals showing oxygen isotopic values associated with the Alpine region (outliers TC20, TC77, TC82, and individual TC83 from Cluster B). These date to the first half of the thirteenth century and were buried in a privileged burial shaft. During the proposed chronology, the Capitanata region was part of the Kingdom of Sicily and Holy Roman Empire, both ruled by Frederik II Hohenstaufen (1194–1250 CE). It is possible that these individuals had their origins in southern Germany and their presence on the site may be related to military expeditions (e.g. the sixth crusade) or diplomatic and political activities.
Individual TC8 was buried according to an Islamic rite and dated to the last decade of the thirteenth century. In 1296 CE, the bishopric was donated by the King Charles II of Naples to ‘Abd al- ‘Azīz, a Muslim knight from the city of Lucera23. Lucera was a fortified town in Capitanata where Muslims from Sicily were resettled in the 1220 s by Frederik II following religious unrests on the island4,23,24. The Islamic population of Lucera could live according to Muslim laws and cultural habits. They were also heavily involved in trading activities and the wars of the Kingdom. Within this, ‘Abd al- ‘Azīz was one the most influential people and likely was given rule over Tertiveri by King Charles II following his participation in the War of the Vespers (1282–1302) in Sicily23. Tertiveri remained under ‘Abd al- ‘Azīz’s control only until 1300 CE, when it was then donated to a Christian knight. Given the privileged burial and the short chronology, individual TC8 is likely a member of ‘Abd al- ‘Azīz’s clan. His oxygen isotopic values indicate that he likely travelled from coastal regions in the western Mediterranean, including Sicily, or from north African coasts. A possibility involves the participation of this individual within the War of the Vespers in Sicily, alongside ‘Abd al- ‘Azīz’.
Future research avenues could be pursued to further the knowledge presented in this paper. In particular, employing techniques such as radiocarbon dating and Bayesian chronological modelling could allow for a more precise chronology, which in turn could offer a more detailed insight into diachronic trends in the socio-economic strategies and links to changing political regimes in Capitanata. The small skeletal collections available for analysis from Montecorvino and San Lorenzo may be augmented by a recently recovered large skeletal assemblage from Montecorvino, which could offer a more comprehensive view of local lifeways in relation to biological sex and age. Additionally, further refinement of food isotopic values through direct analysis of archaeological materials could potentially lead to more precise Bayesian dietary estimates.
Conclusions
The farming economy of Late medieval Capitanata was based on cereal production and complemented by animal management practices. The rearing of animals, for the most part pigs and ovicaprids, allowed for intensive crop manuring. This practice is apparently absent in other Late medieval sites in southern Italy and may have contributed to a local demographic growth following the establishment of the Norman rule. Predominant cereal production consisted of wheat and barley although millet and/or sorghum were also grown in smaller amounts. The latter may have been linked to transhumance practices. Minor consumption of marine fish, potentially associated to Christian fasting rules, shows the existence of trade routes connecting the sub-Apennine region to the Adriatic coast.
Significant differences in individual diets likely reflect a socioeconomic hierarchy which determined purchase power. Millet and/or sorghum, viewed as lower quality grains, were more affordable than wheat or barley. While animal protein, including marine fish, were costlier food items and more easily accessible to higher status individuals. Among these were likely German elites or Angevin soldiers that moved into the region from Alpine regions during the thirteenth century. One single individual, buried according to an Islamic rite, likely travelled from other western Mediterranean coastal regions (e.g. Sicily).
The traditional image of Late Medieval southern Italy is one where a farming economy sustains a hierarchical feudal system and where a succession of foreign influences conflicted and co-existed. Our multi-isotope Bayesian approach employed in the study of bioarchaeological remains largely corroborates such an image but it also offers a high resolution reconstruction of the socioeconomic, cultural, and demographic dynamics of Late medieval communities. These historical aspects had a formative role that remains visible in the linguistic, religious, culinary, and genetic heritage of southern Italy.
Methods
We carried out bulk collagen and bioapatite stable carbon, nitrogen and oxygen isotope analyses on skeletal samples from the late medieval sites of Tertiveri, Montecorvino and San Lorenzo in Carminiano54,69,70,76. Osteological material was sampled with the permission and collaboration of the Soprintendenza Archeologia, Belle Arti e Paesaggio per le Province di Barletta-Andria-Trani e Foggia. Individuals from Tertiveri are currently stored at the Soprintendenza in Foggia, whereas those from Montecorvino and San Lorenzo are curated by the University of Foggia. A description of the archaeological sites and of a preliminary osteological analysis can be found in Supplementary Information File S2. In total, 134 human individuals and 21 faunal specimens were sampled for stable isotope analysis (Supplementary Information File S1; https://www.doi.org/10.48493/w01v-fe90).
The largest number of human samples was recovered from Tertiveri (113 individuals). This included a so-called ‘bishop’ (TC74) and two individuals buried following an Islamic rite (TC8, TC21). Anthropological analysis from Tertiveri revealed a biological sex ratio of approximately three males to each female. Seven females, ten probable females, fourteen males and twenty-five probable males were identified across the adolescent and adult population. However, it was not possible to determine the biological sex of thirty-seven adolescent and adult individuals due to their fragmented and/or commingled state. The same applied to twenty-one non-adult individuals who did not display clear sexual dimorphism. The site was characterized by a high mortality rate among young adults aged between twenty and forty years old. Also a few elderly individuals over sixty were identified. Non-adults of all age groups were underrepresented, which is a common trait of medieval populations in Italy77. Supplementary Information File S2 provides additional detail on biological sex and age at death distributions in Tertiveri.
The dataset from Montecorvino (n = 17) included four adult females, six adult males, and seven non-adults. This represents the original anthropological core of the site, whereas more recently a larger assemblage was excavated. Therefore, demographic profiles are still under investigation and cannot be determined yet. From San Lorenzo (n = 4), two males, one female, and one undetermined adult could be sampled. Further details on this preliminary osteological analysis from the two sites is presented in Supplementary Information File S2.
For comparability purposes we sought to sample rib bones from all individuals. However, many skeletons found in Tertiveri were disarticulated and some did not have preserved ribs. Turnover rates of skeletal bioapatite and collagen vary with osteological element32. In the case of ribs, the collagen reflects the c. 3–5 years prior to death of an individual. For many osteological elements their turnover rates are not precisely know nor how these vary with the age of an individual. Thus, our dietary and mobility isotopic comparisons may reflect different life stages of the analysed individuals. The skeletal samples from Tertiveri consisted of: fifty ribs, twenty-nine cranium fragments (mostly parietal and frontal parts), twelve fibulae, four metatarsal bones, four vertebrae, three tibiae, two femora, two humeri, two scapulae, one clavicula, one mandibular fragment, one metacarpal bone, and one radius. In some instances, it was possible to sample multiple tissues (e.g. tooth plus bone) from the same human (e.g. TC86) or animal individual (e.g. MOPI1). In Montecorvino (n = 17) and San Lorenzo in Carminiano (n = 4), human ribs were sampled for all individuals.
The faunal dataset is composed by five specimens from Tertiveri, seventeen from Montecorvino, and one from San Lorenzo. The Tertiveri animal dataset consists of bone samples from two cattle, one pig, one ovicaprid, and one tortoise. From Montecorvino, bone fragments from two cattle, two poultry, one horse, four ovicaprids and one tortoise were collected. In addition, five teeth and two bone fragments from five pigs were also included. From San Lorenzo, only one tooth and one bone from the same deer specimen were sampled. Despite potential turnover differences across bone samples and teeth, we assumed that the shorter lifespan for animals should not significantly impact the interpretation of isotopic results.
Samples from Tertiveri were pre-treated at the stable isotope lab of the Max Planck Institute of Geoanthropology (former “for the Science of Human History in Jena”) (MPI-GEA), Germany, whereas samples from Montecorvino and San Lorenzo in Carminiano were pre-treated at the ‘iCONa’ lab at the Università degli studi della Campania ‘Luigi Vanvitelli’ in Caserta, Italy. All stable isotope measurements were carried out at the MPI-GEA. Sample pretreatment and analytical measurements are described in detail in Supplementary Information File 4.
We employed the Bayesian dietary mixing model ReSources (previously FRUITS) to quantitatively reconstruct late medieval diets in Capitanata45,55,66,78. Modelling options are described in Supplementary Information File 3.
Human mobility was investigated using δ18O bone measurements. These were compared with a reference baseline for δ18O values of drinkable water across western Europe. This baseline relies on a dataset of mean δ18OWater annual values from the Regionalized Cluster-based Water Isotope Prediction model (RCWIP)67 from which a smoothed spatial surface for δ18O was created using the Bayesian model AverageR45,79. Probabilistic assignment of residence was generated using the model LocateR45,68. For a direct comparison of human bone and water δ18O values, the values for the former, reported as carbonate measurements relative to the standard VPDB, were re-expressed relative to the VSMOW standard (δ18OVSMOW = δ18OVPDB * 1.03092 + 30.92). Following Chenery et al.80 carbonate values were converted into phosphate (δ18Ophosphate = δ18Ocarbonate * 1.0322−9.6849) and finally into drinkable water following Pollard et al.43 (δ18OWater = δ18Ophosphate * 1.55−33.49). For modelling purposes and in accordance with previous studies, we employed a conservative 2‰ uncertainty for modelled human δ18OWater values which reflects uncertainty introduced by a variety of factors (e.g. cooking, bone diagenesis, etc.)41,42.
For cluster classification of human δ13C and δ15N values we employed the mclust R package81. Mixture estimation relied on the expectation–maximisation algorithm and the optimal number of clusters was identified using the Bayesian Information Criterion (BIC). For cluster classification of human δ18O values we employed the tclust R package82. This gave a cluster assignment similar to mclust but tclust employs a trimming approach83, based on a threshold value, to identify outliers. For our application we employed a trimming threshold value of 5%.
Ethic statement
Archaeological, anthropological, and isotopic analysis of individuals from the sites of Tertiveri, Montecorvino, and San Lorenzo in Carminiano were conducted with the permission and collaboration of the Soprintendenza Archeologia, Belle Arti e Paesaggio per le province di Barletta-Andria-Trani e Foggia.
Data availability
Data that support the findings of this study are available in Supplementary Information file S1 and at https://www.doi.org/10.48493/w01v-fe90.
References
Chavarría Arnau, A. Changes in Scale in the Italian Countryside from Late Antiquity to the Early Middle Ages. in Scale and Scale Change in the Early Middle Ages vol. 6, 121–132 (Brepols Publishers, 2011).
Wickham, C. Framing the Early Middle Ages: Europe and the Mediterranean 400–800 (Oxford University Press, 2006).
Wickham, C. Medieval Europe (Yale University Press, 2016).
Abulafia, D. Italy in the Central Middle Ages 1000–1300 (Oxford University Press, 2004).
Ward-Perkins, B. The Fall of Rome: And the End of Civilization (Oxford University Press, 2006).
Brown, G. S. The Norman Conquest of Southern Italy and Sicily. (McFarland, 2003).
Houben, H. Die Normannen (C. H. Beck, 2012).
Garnsey, P. Food & Society Classical Antiquity (Cambridge University Press, 2008).
Montanari, M. Medieval Tastes. Food, Cooking, and the Table (Columbia Univ Press, 2012).
Sakellariou, E. Southern Italy in the Late Middle Ages: Demographic, Institutional and Economic Change in the Kingdom of Naples, c.1440-c.1530. (Brill, 2011).
Loud, G. A. The Latin Church in Norman Italy (Cambridge University Press, 2007).
Favia, P. Il disegno del territorio e il paesaggio agrario apulo. La Capitanata fra XI e XV secolo alla luce delle ricerche archeologiche. In I paesaggi aperti agropastorali del Mediterraneo: genesi, economie, governo del territorio. Atti del 4° Convegno CRIAT 51–84 (eds Mastrodonato, G. & Salvemini, B.) (Università del Salento, 2020). https://doi.org/10.1285/i9788883051678p51.
Favia, P. Processi di popolamento, configurazioni del paesaggio e tipologie insediative in Capitanata nei passaggi istituzionali dell’XI secolo. In La Capitanata e l’Italia Meridionale nel sec. XI: da Bisanzio ai Normanni, ‘Atti delle II giornate medievali di Capitanata: Apricena 16–17 aprile 2005’ (eds Favia, P. & De Venuto, G.) 103–135 (Edipuglia, 2011).
Favia, P. & Maruotti, M. Caratteri insediativi delle recinzioni e fortificazioni di terra nella Capitanata medievale. Diagnostica archeologica, analisi di superficie, casi di scavo. Archeologia Medievale 40, 91–101 (2013).
De Venuto, G. Allevamento, ambiente ed alimentazione nella Capitanata medievale: archeozoologia e archeologia globale dei paesaggi. (Edipuglia, 2013).
Ruas, M.-P. Cultures et moissons à Fiorentino: étude des semences carbonisées. In Fiorentino ville désertée nel contesto della Capitanata medievale (Ricerche 1982–1993) (eds Calò Mariani, M. S. et al.) 541–565 (École Française de Rome, 2012). https://doi.org/10.1400/209868.
Rubini, M., Dell’Anno, V., Giuliani, R., Favia, P. & Zaio, P. The first probable case of leprosy in Southeast Italy (13th-14th Centuries AD, Montecorvino, Puglia). J. Anthropol. 2012, 1–7 (2012).
Tafuri, M. A. et al. Evidenze isotopiche e paleodieta nel Neolitico Pugliese: verso la globalizzazione?. Studi di Preistoria e Protostoria 4, 357–361 (2017).
Tafuri, M. A. et al. Herding practices in the ditched villages of the Neolithic Tavoliere (Apulia, South-east Italy): A vicious circle? The Isotopic evidence. In Early Farmers: The View from Archaeology and Science (eds Whittle, A. & Bickle, P.) (British Academy, 2014). https://doi.org/10.5871/bacad/9780197265758.003.0008.
Tafuri, M. A. et al. Life and death in Neolithic Southeastern Italy: The strontium isotopic evidence. Int. J. Osteoarchaeol. 26, 1045–1057 (2016).
Torino, M. et al. Convento di San Francesco a Folloni: The function of a Medieval Franciscan Friary seen through the burials. Herit. Sci. 3, 27 (2015).
Rolandsen, G. L., Arthur, P. & Alexander, M. A tale of two villages: Isotopic insight into diet, economy, cultural diversity and agrarian communities in medieval (11th–15th century CE) Apulia, Southern Italy. J. Archaeol. Sci. Rep. 28, 102009 (2019).
Engl, R. ‘Abd al-‘Azīz von Lucera (gest. 1301). Aufstieg und Fall eines muslimischen Ritters im Königreich Sizilien. In Christen und Muslime in der Capitanata im 13. Jahrhundert. Archäologie und Geschichte Vol. 2 (eds Clemens, L. & Matheus, M.) 231–249 (Kliomedia, 2018).
Matheus, M. Lucera, Tertiveri, Bischofsstadt und Bischofssitz. Muslimische Stadt und muslimische Adelsresidenz. Genese eines interdisziplinären Forschungsprojektes zur Geschichte Süditaliens. in Christen und Muslime in der Capitanata im 13. Jahrhundert. Archäologie und Geschichte (eds. Clemens, L. & Matheus, M.) vol. 2 109–130 (Kliomedia, 2018).
Abulafia, D. Frederick II: A Medieval Emperor (Oxford Univ Pr, 1992).
Melillo, M. Briciole francoprovenzali nell’Italia meridionale. Vox Romanica 40, 39–47 (1981).
Halsall, G. The sources and their interpretations. in The New Cambridge Medieval History, Volume 1 c.500 – c.700 (ed. Fouracre, P.) 56–92 (Cambridge University Press, 2005).
Hakenbeck, S. E., Evans, J., Chapman, H. & Fóthi, E. Practising pastoralism in an agricultural environment: An isotopic analysis of the impact of the Hunnic incursions on Pannonian populations. PLoS ONE 12, e0173079 (2017).
Dreslerová, D. et al. Maintaining soil productivity as the key factor in European prehistoric and Medieval farming. J. Archaeol. Sci. Rep. 35, 102633 (2021).
Sirignano, C. et al. Animal husbandry during early and high middle ages in the Basque Country (Spain). Quatern. Int. 346, 138–148 (2014).
Alexander, M. M., Gerrard, C. M., Gutiérrez, A. & Millard, A. R. Diet, society, and economy in late medieval Spain: Stable isotope evidence from Muslims and Christians from Gandía, Valencia. Am. J. Phys. Anthropol. 156, 263–273 (2015).
Fahy, G. E., Deter, C., Pitfield, R., Miszkiewicz, J. J. & Mahoney, P. Bone deep: Variation in stable isotope ratios and histomorphometric measurements of bone remodelling within adult humans. J. Archaeol. Sci. 87, 10–16 (2017).
Hedges, R. E. M., Clement, J. G., Thomas, C. D. L. & O’Connell, T. C. Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements. Am. J. Phys. Anthropol. 133, 808–816 (2007).
Fuller, B. T., Richards, M. P. & Mays, S. A. Stable carbon and nitrogen isotope variations in tooth dentine serial sections from Wharram Percy. J. Archaeol. Sci. 30, 1673–1684 (2003).
Fernandes, R., Nadeau, M.-J. & Grootes, P. M. Macronutrient-based model for dietary carbon routing in bone collagen and bioapatite. Archaeol. Anthropol. Sci. 4, 291–301 (2012).
Lee-Thorp, J. A. On isotopes and old bones. Archaeometry 50, 925–950 (2008).
Hedges, R. E. M., Stevens, R. E. & Richards, M. P. Bone as a stable isotope archive for local climatic information. Quatern. Sci. Rev. 23, 959–965 (2004).
Szpak, P., White, C. D., Longstaffe, F. J., Millaire, J.-F. & Sánchez, V. F. V. Carbon and nitrogen isotopic survey of Northern Peruvian plants: Baselines for paleodietary and paleoecological studies. PLoS ONE 8, e53763 (2013).
Bogaard, A., Heaton, T. H. E., Poulton, P. & Merbach, I. The impact of manuring on nitrogen isotope ratios in cereals: Archaeological implications for reconstruction of diet and crop management practices. J. Archaeol. Sci. 34, 335–343 (2007).
Szpak, P. Complexities of nitrogen isotope biogeochemistry in plant-soil systems: Implications for the study of ancient agricultural and animal management practices. Front. Plant Sci. 5, 288 (2014).
Lightfoot, E. & O’Connell, T. C. On the use of biomineral oxygen isotope data to identify human migrants in the archaeological record: Intra-sample variation, statistical methods and geographical considerations. PLoS ONE 11, e0153850 (2016).
Pederzani, S. & Britton, K. Oxygen isotopes in bioarchaeology: Principles and applications, challenges and opportunities. Earth Sci. Rev. 188, 77–107 (2019).
Pollard, A. M., Pellegrini, M. & Lee-Thorp, J. A. Technical note: Some observations on the conversion of dental enamel δ18Op values to δ18Ow to determine human mobility. Am. J. Phys. Anthropol. 145, 499–504 (2011).
Tuross, N., Reynard, L. M., Harvey, E., Coppa, A. & McCormick, M. Human skeletal development and feeding behavior: The impact on oxygen isotopes. Archaeol. Anthropol. Sci. 9, 1453–1459 (2017).
Cocozza, C., Cirelli, E., Groß, M., Teegen, W.-R. & Fernandes, R. Presenting the compendium isotoporum Medii Aevi, a multi-isotope database for medieval Europe. Sci Data 9, 354 (2022).
Cocozza, C., Cirelli, E., Groß, M., Teegen, W.-R. & Fernandes, R. Compendium Isotoporum Medii Aevi (CIMA). (2021).
DeNiro, M. J. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317, 806–809 (1985).
Fernandes, R. et al. Assessing screening criteria for the radiocarbon dating of bone mineral. Nucl. Instrum. Methods Phys. Res. Sect. B 294, 226–232 (2013).
Bonafini, M., Pellegrini, M., Ditchfield, P. & Pollard, A. M. Investigation of the ‘canopy effect’ in the isotope ecology of temperate woodlands. J. Archaeol. Sci. 40, 3926–3935 (2013).
Savastio, S. Notizie storiche sull’antica città di Montecorvino di Puglia e sul borgo di Serritella. (Industria graf. puteolana, 1939).
Corvino, C. Montecorvino. Il contributo dell’antracologia per la ricostruzione dei paesaggi naturali e dell‟ interazione uomo-ambiente. in Storia e Archeologia globale (ed. Volpe, G.) vol. 1 35–41 (Edipuglia, 2015).
Makarewicz, C. A. Stable isotopes in pastoralist archaeology as indicators of diet, mobility, and animal husbandry practices. In Isotopic Investigations of Pastoralism in Prehistory (eds Ventresca Miller, A. & Makarewicz, C. A.) (Routledge, 2017).
Stevens, R. E., Lister, A. M. & Hedges, R. E. M. Predicting diet, trophic level and palaeoecology from bone stable isotope analysis: A comparative study of five red deer populations. Oecologia 149, 12–21 (2006).
Favia, P. et al. San Lorenzo “in Carminiano” presso Foggia: indagine archeologica su un sito medievale del Tavoliere di Puglia in un contesto di moderna espansione edilizia. In Atti del V Congresso nazionale di archeologia medievale. Foggia-Manfredonia 30 settembre-3 ottobre 2009 (eds Volpe, G. & Favia, P.) 382–391 (All’insegna del giglio, 2009).
Fernandes, R., Grootes, P., Nadeau, M.-J. & Nehlich, O. Quantitative diet reconstruction of a Neolithic population using a Bayesian mixing model (FRUITS): The case study of Ostorf (Germany). Am. J. Phys. Anthropol. 158, 325–340 (2015).
Bynum, C. W. Holy feast and holy fast: The religious significance of food to medieval women (University of California Press, 2000).
Tsutaya, T. & Yoneda, M. Quantitative reconstruction of weaning ages in archaeological human populations using bone collagen nitrogen isotope ratios and approximate Bayesian computation. PLoS ONE 8, e72327 (2013).
Marino, J. A. L’economia pastorale nel Regno di Napoli. (Guida, 1992).
De Venuto, G. Contributo alla ricostruzione dei caratteri dell’allevamento transumante ovino tra Abruzzo e Tavoliere di Puglia in età medievale. in Vie degli Animali, Vie degli Uomini. Transumanza ed altri Spostamenti di Animali nell’Europa Tardoantica e Medievale (eds. Volpe, G., Buglione, A. & De Venuto, G.) 69–81 (Edipuglia, 2010).
Cernusak, L. A. et al. Stable isotopes in leaf water of terrestrial plants. Plant Cell Environ. 39, 1087–1102 (2016).
Makarewicz, C. A. & Pederzani, S. Oxygen (δ18O) and carbon (δ13C) isotopic distinction in sequentially sampled tooth enamel of co-localized wild and domesticated caprines: Complications to establishing seasonality and mobility in herbivores. Palaeogeogr. Palaeoclimatol. Palaeoecol. 485, 1–15 (2017).
Makarewicz, C. A., Arbuckle, B. S. & Öztan, A. Vertical transhumance of sheep and goats identified by intra-tooth sequential carbon (δ13C) and oxygen (δ18O) isotopic analyses: Evidence from Chalcolithic Köşk Höyük, central Turkey. J. Archaeol. Sci. 86, 68–80 (2017).
Schurr, M. R. Using stable nitrogen-isotopes to study weaning behavior in past populations. World Archaeol. 30, 327–342 (1998).
Cocozza, C., Fernandes, R., Ughi, A., Groß, M. & Alexander, M. M. Investigating infant feeding strategies at Roman Bainesse through Bayesian modelling of incremental dentine isotopic data. Int. J. Osteoarchaeol. 31, 429–439 (2021).
Kellner, C. M. & Schoeninger, M. J. A simple carbon isotope model for reconstructing prehistoric human diet. Am. J. Phys. Anthropol. 133, 1112–1127 (2007).
Fernandes, R., Millard, A. R., Brabec, M., Nadeau, M.-J. & Grootes, P. Food reconstruction using isotopic transferred signals (FRUITS): A Bayesian model for diet reconstruction. PLoS ONE 9, e87436 (2014).
Terzer, S., Wassenaar, L. I., Araguás-Araguás, L. J. & Aggarwal, P. K. Global isoscapes for δ18O and δ2H in precipitation: Improved prediction using regionalized climatic regression models. Hydrol. Earth Syst. Sci. 17, 4713–4728 (2013).
Wang, X. et al. The circulation of ancient animal resources across the Yellow River Basin: A preliminary Bayesian re-evaluation of Sr isotope data from the early neolithic to the Western Zhou Dynasty. Front. Ecol. Evol. 9, 16 (2021).
Clemens, L. & Matheus, M. Troia und Tertiveri: Transformationen byzantinischer Bischofssitze in Süditalien. in Menschen, Bilder, Sprache, Dinge. Wege der Kommunikation zwischen Byzanz und dem Westen. 2 Menschen und Worte (eds. Daim, F., Gastgeber, C., Heher, D. & Rapp, C.) 225–234 (Verlag des Römisch-Germanischen Zentralmuseums, 2018).
Teegen, W.-R. Die Toten aus der Bischofsgruft von Tertiveri (Prov. Foggia, Apulien), Ausgrabung 2011. Vorbericht über die anthropologisch-paläopathologischen Untersuchen. in Christen und Muslime in der Capitanata im 13. Jahrhundert. Archäologie und Geschichte (eds. Clemens, L. & Matheus, M.) vol. 2, 275–286 (Kliomedia, 2018).
Favia, P., D’Altilia, L. & Laccone, R. Accumulazione delle risorse e difesa degli spazi signorili nella città di Montecorvino: l’apporto integrato dei dati da indagine aerofotogrammometrica e scavo archeologico. in VIII Congresso nazionale di archeologia medievale. ‘Chiesa del Cristo Flagellato, Matera, 12–15 settembre 2018’ (eds. Sogliani, F., Gargiulo, B., Annunziata, E. & Vitale, V.) 228–231 (All’Insegna del Giglio, 2018).
Tafuri, M. A., Craig, O. E. & Canci, A. Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Am. J. Phys. Anthropol. 139, 146–153 (2009).
Sorghum and millets in human nutrition. (Food and Agriculture Organization of the United Nations, 1995).
Dyer, C. Standards of Living in the Later Middle Ages: Social Change in England c.1200–1520. (Cambridge University Press, 1989). https://doi.org/10.1017/CBO9781139167697.
MacKinnon, M. High on the Hog: Linking zooarchaeological, literary, and artistic data for pig breeds in Roman Italy. Am. J. Archaeol. 105, 649–673 (2001).
Favia, P. et al. Montecorvino: Parabola insediativa di una cittadina dei Monti Dauni fra XI e XVI secolo. in VII Congresso nazionale di archeologia medievale. ‘Palazzo Turrisi, Lecce, 9–12 settembre 2015’ (eds. Arthur, P. & Imperiale, M. L.) 191–196 (All’insegna del giglio, 2015).
Baldoni, M., Nardi, A., De Angelis, F., Rickards, O. & Martínez-Labarga, C. How does diet influence our lives? Evaluating the relationship between isotopic signatures and mortality patterns in Italian Roman imperial and medieval periods. Molecules 26, 3895 (2021).
Sołtysiak, A. & Fernandes, R. Much ado about nothing: assessing the impact of the 4.2 kya event on human subsistence patterns in northern Mesopotamia using stable isotope analysis. Antiquity 95, 1145–1160 (2021).
Cubas, M. et al. Latitudinal gradient in dairy production with the introduction of farming in Atlantic Europe. Nat. Commun. 11, 2036 (2020).
Chenery, C. A., Pashley, V., Lamb, A. L., Sloane, H. J. & Evans, J. A. The oxygen isotope relationship between the phosphate and structural carbonate fractions of human bioapatite. Rapid Commun. Mass Spectrom. 26, 309–319 (2012).
Scrucca, L., Fop, M., Murphy, T. B. & Raftery, A. E. mclust 5: Clustering, classification and density estimation using gaussian finite mixture models. R J. 8, 289 (2016).
Fritz, H., García-Escudero, L. A. & Mayo-Iscar, A. tclust: An R package for a trimming approach to cluster analysis. J. Stat. Softw. 47, 1–26 (2012).
Cuesta-Albertos, J. A., Gordaliza, A. & Matrán, C. Trimmed k-means: An attempt to robustify quantizers. Ann. Stat. 25, 553–576 (1997).
Acknowledgements
We thank Dr. Simona Altieri, Dr. Maria Rosa Di Cicco, Mary Lucas and Erin Scott for technical support in sample pretreatment and measurement. We also thank the Soprintendenza Archeologia, Belle Arti e Paesaggio per le province di Barletta-Andria-Trani e Foggia for granting access to osteological materials from the sites of Tertiveri, Montecorvino and San Lorenzo in Carminiano. The Gerda Henkel Foundation (Düsseldorf, Germany) supported the project "Christen und Muslime im mittelalterlichen Nordapulien. Archäologische Untersuchungen in Tertiveri (Prov. Foggia)" (No. AZ 01/F/15) that included archaeological, anthropological, and isotopic investigations.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
C.C. Co-designed the study, analysed the data, wrote the manuscript and performed the modelling; W.R.T. Co-designed the study, provided for anthropological analysis and interpretation, and co-wrote the manuscript; I.V. Provided for anthropological analysis and interpretation, and co-wrote the manuscript; P.F. Provided for archaeological materials and interpretation, and co-wrote the manuscript; R.G. Provided for archaeological materials and interpretation, and co-wrote the manuscript; I.M.M.: Provided for archaeological materials and interpretation; D.O. Provided for archaeological materials and interpretation; L.C. Provided for archaeological materials and interpretation, and co-wrote the manuscript; M.G. developed modelling apps and co-wrote the manuscript; P.R. Provided for lab capabilities and edited the manuscript; C.L. Provided for lab capabilities and edited the manuscript; R.F. Co-designed the study, supervised the modelling, and edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cocozza, C., Teegen, WR., Vigliarolo, I. et al. A Bayesian multi-proxy contribution to the socioeconomic, political, and cultural history of late medieval Capitanata (southern Italy). Sci Rep 13, 4078 (2023). https://doi.org/10.1038/s41598-023-30706-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-30706-9
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
