
Abstract
Conventional nitrogen removal in wastewater treatment requires a high oxygen and energy input. Anaerobic ammonium oxidation (anammox), the single-step conversion of ammonium and nitrite to nitrogen gas, is a more energy and cost effective alternative applied extensively to sidestream wastewater treatment. It would also be a mainstream treatment option if species diversity and physiology were better understood. Anammox bacteria were enriched up to 80%, 90% and 50% relative abundance, from a single inoculum, under standard enrichment conditions with either stepwise-nitrite and ammonia concentration increases (R1), nitric oxide supplementation (R2), or complex organic carbon from mainstream wastewater (R3), respectively. Candidatus Brocadia caroliniensis predominated in all reactors, but a shift towards Ca. Brocadia sinica occurred at ammonium and nitrite concentrations > 270 mg NH4–N L−1 and 340 mg NO2–N L−1 respectively. With NO present, heterotrophic growth was inhibited, and Ca. Jettenia coexisted with Ca. B. caroliniensis before diminishing as nitrite increased to 160 mg NO2–N L−1. Organic carbon supplementation led to the emergence of heterotrophic communities that coevolved with Ca. B. caroliniensis. Ca. B. caroliniensis and Ca. Jettenia preferentially formed biofilms on surfaces, whereas Ca. Brocadia sinica formed granules in suspension. Our results indicate that multiple anammox bacteria species co-exist and occupy sub-niches in anammox reactors, and that the dominant population can be reversibly shifted by, for example, changing nitrogen load (i.e. high nitrite concentration favors Ca. Brocadia caroliniensis). Speciation has implications for wastewater process design, where the optimum cell immobilization strategy (i.e. carriers vs granules) depends on which species dominates.
Similar content being viewed by others
Introduction
Anaerobic ammonium oxidation (anammox), combined with partial nitritation is applied widely to treat nitrogen-rich and carbon-deficient wastewaters (e.g., sidestream treatment) due to significant energy savings relative to conventional processes. It has also been proposed as a sustainable treatment option for treating municipal wastewaters (i.e. mainstream treatment). Nineteen candidatus anammox bacteria species have been identified in various environments, including suboxic marine zones, coastal sediments, lakes, and wastewater treatment plants. These have been classified into five candidatus genera1,2,3, and while anammox bacteria can colonize diverse natural and engineered systems, different genera rarely coexist in the same habitat4. Differences in growth rates, substrate affinities, sensitivities to inhibitory compounds, preferred growth substrates and differential metabolic pathways, are all thought to contribute to niche specialization1,5,6,7,8,9,10.
Population shifts at species and genus level have been reported in anammox lab-scale reactors under various conditions9,10,11. During scale-up of the first full-scale commercial anammox reactor, the dominant population shifted from Ca. Kunenia stuttgartiensis to Ca. Brocadia anammoxidans, although reasons for this were not provided12. Studies have reported that specific environmental conditions in partial nitritation/anammox (PN/A) reactors can select for single anammox bacteria species only11,13. For instance, Ca. Jettenia moscovienalis2, Ca. B. caroliniensis14, and Ca. B. sinica13 were detected in distinct sidestream reactors treating anaerobic digester liquor, whereas Ca. Brocadia. sp. 40 was identified as the dominant anammox bacteria under mainstream conditions15. Park et al.11 showed that feed composition is more important in anammox bacteria selection than inoculum and reactor configuration. Nonetheless, there is no apparent consensus on which factors select for one anammox bacteria species over another.
Elucidating factors that enrich for specific anammox bacteria species with specific kinetic and physiological properties would potentially enhance process design and performance. Anammox bacteria exist in a range of conditions, such as sidestream and mainstream PN/A systems with high and low ammonium/nitrite concentrations respectively, and many factors are likely involved in species selection. While nitrite (NO2−) is toxic to bacteria, it can also act as an electron acceptor for ammonium oxidation and electron donor for bicarbonate reduction to biomass. It thus applies an anammox bacteria-species selection pressure on the basis of their abilities to utilize and tolerate it. A 50% reduction in anammox activity has been reported in anammox bacteria exposed to NO2- concentrations between 100 and 400 mg N L−116,17,18. While nitric oxide (NO), a potent oxidant produced from nitrite as an intermediate in the anammox biochemical pathway19,20, is toxic, anammox bacteria can tolerate it a higher concentrations than many bacteria21,22. In addition to potential selection pressures from nitrite and NO, the ability to also consume organic substrates (i.e. acetate and propionate) has been found to confer competitive advantages to Ca. B. fulgida and Ca. Anammoxoglobus propionicus respectively, over other species including denitrifers6,7. It remains to be determined whether selection of such ‘facultative chemoorganotrophs’ would be favored in the complex organic carbon milieu present under mainstream conditions. Nonetheless, the anammox bacteria occupy highly specialized niches that are defined by more than just the concentrations of ammonium and nitrite.
The ability to enhance the activity of specific anammox bacteria species with favorable physiological and growth properties is particularly advantageous for starting up and optimizing industrial anammox processes. While established sidestream anammox sludge are commonly used to seed or bio-augment new full-scale installations, inoculating from existing full-scale non-anammox installations to start up anammox reactors is logistically challenging11, and time-consuming owing to the sensitivity of the process to feed composition, oxygen12 and competing microbial species23. High biomass retention is required in anammox reactors due to the slow growth rates of anammox bacteria. This can be achieved by promoting biomass aggregation in biofilm-based anammox reactors24. Biomass in an anammox reactor can self-assemble into flocs in suspension, fixed films on surfaces or carriers, small granules, big granules or some combination of all of these morphologies25. Such aggregates can play functionally different roles within the reactor and even affect nitrogen removal efficiencies26,27. Understanding which factors drive species selection and whether different species assume particular biofilm morphologies could inform process design and control strategies for achieving more stable nitrogen removal under the broad range of operating conditions typically encountered in anammox bioreactors28.
This work aims to explore how anammox community composition, process performance and biofilm morphology are shifted by factors typically encountered in industrial anammox systems (i.e. mainstream compared to sidestream). It was hypothesized that different substrate compositions, simulating (a) organic carbon-deficient wastewaters with N-loads for domestic mainstream and sidestream wastewaters (reactors R1) and with (b) an oxidative stress typically encountered in PN/A systems through the exposure to NO (reactor R2), or (c) domestic strength mainstream wastewater with high COD:N (reactor R3), could select for distinct community, specifically anammox bacteria species. While some of these factors have been previously investigated independently17,29, this study examines these factors on anammox bacteria species selection from the same inoculum. Gaining further insight into how the relative abundance of anammox species can be manipulated will inform N-removal process design, and may contribute to the understanding of niche partitioning in complex microbial/environmental habitats.
Results
Start-up period reduced and N removal activity enhanced with NO supplementation
Anammox bacteria were successfully enriched under all tested enrichment conditions albeit with varying start-up times. Start-up period was the shortest in R2 supplemented with NO and anammox activity was observed within 20 days of inoculation compared to 39 days for R1, operated under standard enrichment conditions (Figs. 1A,B). In the presence of complex organic carbon in R3, anammox activity was only detected after 50 days of operation (Fig. 1C). In both R1 and R2, ammonium and nitrite concentrations were increased to 280 mg N L−1 and 350 mg N L−1, respectively (Fig. 1A,B), above which anammox activity was inhibited. In addition to the shorter start-up period, a shorter hydraulic retention time (HRT) was applied to R2 than R1 due to higher N removal rates of 1200 mg N L−1 day−1 compared to 800 mg N L−1 day−1 under stable operation (Fig. 1A,B). Despite the higher loading rate, the suspended solid concentrations were comparable in both reactors indicating a higher specific N removal activity for R2 than R1. A significantly lower N loading rate of 121 ± 6 mg N L−1 day−1 was achieved in R3. Final effluent ammonium concentration dropped steadily from day 58 to 80, with the reactor displaying stable ammonium removal activity by anammox bacteria henceforth. Residual nitrite in the effluent also decreased gradually from day 60 to 100 along with a decrease in ammonium concentration (Fig. 1C). The average total chemical oxygen demand (TCOD) and soluble chemical oxygen demand (sCOD) in the effluent were 87 ± 9 mg L−1 and 51 ± 8 mg L−1, respectively with an average removal rate of 520 mg L−1 day−1 from day 300 onwards.

Start-up and enrichment of anammox bacteria from activated sludge fed with (A) synthetic waste water with ammonium and nitrite in R1, (B) synthetic waste water with ammonium, nitrite and continuous supply of nitric oxide in R2, and (C) primary effluent supplemented with nitrite in R3. MLVSS ( ), Influent nitrite (NO2,
), Influent nitrite (NO2,  ) and nitrogen loading rates (NLR,
) and nitrogen loading rates (NLR,  ) of each enrichment condition are shown in the upper panel of each graph; with relative abundance of dominant anammox bacteria OTUs affiliated to Ca. B. caroliniensis (
) of each enrichment condition are shown in the upper panel of each graph; with relative abundance of dominant anammox bacteria OTUs affiliated to Ca. B. caroliniensis ( ) and Ca. B. sinica (
) and Ca. B. sinica ( ) and correlated non-anammox bacteria OTUs affiliated to Anaerolineaceae (
) and correlated non-anammox bacteria OTUs affiliated to Anaerolineaceae ( ) and Fimbriimonadia (
) and Fimbriimonadia ( ) are shown in the lower panels of (A,B); Ca. Jettenia (
) are shown in the lower panels of (A,B); Ca. Jettenia ( ) was also detected in R2 (B). Ca. B. caroliniensis, the only dominant anammox bacteria in R3, is shown in (C) along with correlated non-anammox bacteria OTUs affiliated to Comamonadaceae (
) was also detected in R2 (B). Ca. B. caroliniensis, the only dominant anammox bacteria in R3, is shown in (C) along with correlated non-anammox bacteria OTUs affiliated to Comamonadaceae ( ) and Ca. Aquirestis (
) and Ca. Aquirestis ( ) dominating at a different stage. The relative abundance of total anammox bacteria is highlighted as area plot (
) dominating at a different stage. The relative abundance of total anammox bacteria is highlighted as area plot ( ) in each graph. Red dotted lines denote the time points at which anammox biofilm was scraped from the wall of the reactor into suspension. The detailed chemical and microbial community (with OTUs > 5% at any analysed time point) can be found in Supplementary Fig. S1, Supporting Information.
) in each graph. Red dotted lines denote the time points at which anammox biofilm was scraped from the wall of the reactor into suspension. The detailed chemical and microbial community (with OTUs > 5% at any analysed time point) can be found in Supplementary Fig. S1, Supporting Information.
Either high N load or concentration, or both, could shift the dominant anammox bacteria species from Ca. B. caroliniensis to Ca. B. sinica
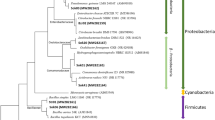
The microbial communities of three reactors were differentiated by day of operation (R = 0.53, p = 0.007) and by the use of different reactors, i.e. R1 vs R2 vs R3 (R = 0.38, p = 0.001). In addition, the R1 and R2 communities at different levels of N showed relatively strong dissimilarity (R = 0.48 and 0.57, respectively, with p = 0.001 for both). Along with the increase in anammox activity, a shift in the functional anammox bacteria was observed in R1 and R2 along with increasing N load, but not in R3 where low N load was maintained. 16S rRNA gene amplicon sequencing showed that anammox bacteria were below the detection limit (< 0.018%) at the start of reactor operation for all three reactors. In R1 and R2, operational taxonomic units (OTUs) annotated to anammox bacteria increased progressively to 80% (day 110) and 90% (day 95) of the relative abundances of OTUs, respectively, at influent nitrite concentration > 200 mg N L-1 (Fig. 1A,B). The microbial communities of R1 and R2 at different levels of N level showed relative strong dissimilarity (R = 0.48 and 0.57, respectively, with p = 0.001 for both). The microbial community at high N load period, on the other hand, was not highly differentiated (R = 0.29, p = 0.003). Despite the increase in relative abundance of multiple OTUs affiliated to anammox bacteria in both R1 and R2, a single OTU annotated to Ca. Brocadia, identified as Ca. B. caroliniensis by clone library analysis (Fig. 2), dominated throughout the first 120 days of reactor operation. Ca. B. caroliniensis increased during enrichment to 50% relative abundance in R1 and R2. However, a further increase in influent ammonium and nitrite concentrations beyond 220 mg N L−1 from day 100 (N loading rate of 500 mg N L−1-day−1 for R1 and 750 mg N L−1-day−1 for R2) resulted in the gradual increase of Ca. Brocadia_2, identified as Ca. B. sinica by clone library analysis (Fig. 2).

Phylogenetic tree based on 16S rRNA sequences of major OTUs (with postfix “*”) and clones (with postfix “**”) from amplicon sequencing and clone library analysis, respectively. Number of identical colonies per total colonies picked is indicated in parentheses, e.g. 34/40 indicated 34 identical colonies per 40 colonies picked. The phylogenetic tree was generated by ARB with SILVA database. Sequences obtained from clone library and amplicon sequencing were inserted into the tree using parsimony insertion tool of ARB. The closest neighbor sequences were selected to generate the final tree with neighbor-joining method with bootstrap of 1000 replications. Dashed line indicates the division of family Ca. Brocadiaceae and genus Ca. Brocadia. Methanosaeta concilii was selected as the outgroup. Only the closest identified sequences were selected to be shown. The scale indicates 0.1 nucleotide change per nucleotide position. Sequence affiliations and relative abundances of major anammox bacteria were consistent between amplicon sequencing and clone library analysis.
A decrease in Ca. B. caroliniensis was also observed (Fig. 1A,B). Beyond 180 days of reactor operation, Ca. B. sinica increased to 33% and 42% in relative abundance in R1 and R2, respectively, while Ca. B. caroliniensis decreased to less than 10% in relative abundance in both reactors. In the presence of organic carbon, Ca. B. caroliniensis was the most dominant anammox taxon throughout the enrichment process in R3 operated at a low N loading rate of 120 mg N L−1 day−1 (Figs. 1C). However, the relative abundance of total anammox bacteria was significantly lower in R3 (~ 50%) than in R1 and R2 (~ 80%), suggesting a more competitive environment for anammox bacteria in the presence of organic carbon. Fluorescent in situ hybridization (FISH) analysis (Fig. 3D–F) on R1 (day 80) and R2 (day 675) further indicated that Ca. B. sinica dominated in those reactors while Ca. B. caroliniensis remained as the only anammox bacteria detected in R3 (day 683). The designed species-specific FISH probes served to observe gradual population shifts during reactor operation in response to changes in the controlling factors.

Light microscopy images of suspended biomass samples showing granular structure in R1 (A) and R2 (B) and floccular structure in R3 (C). FISH analysis carried out on crushed granules to confirm the dominance of Ca. B. sinica (cyan) in R1 (D), and R2 (E) and the prevalence of Ca. B. caroliniensis (magenta) in R3 (F) at the end of Phase III. All other anammox bacteria are in blue. FISH images shown here are representatives of the culture. Scale bars of (A), (B) and (C) indicate 1 mm, while (D), (E) and (F) indicate 1 µm.
While other OTUs affiliated to genus Ca. Brocadia were also detected, their relative abundance was less than 10% (Supplementary Fig. S1). The presence of these Ca. Brocadia OTUs is likely due to different strains of Ca. Brocadia or sequencing errors as only low relative abundances were detected. Aside from Ca. Brocadia, OTUs affiliated to Ca. Jettenia emerged to coexist with Ca. B. caroliniensis only in R2 (with continuous supply of NO), suggesting that the presence of NO may provide a competitive advantage for Ca. Jettenia. However, Ca. Jettenia diminished with increasing N loading rate (i.e. after day 102).
Excess NO availability over nitrite selects for Ca. Jettenia
Ca. Jettenia was only observed in R2 but not in R1 and R3, which could be due to the presence of NO. However, during the enrichment process it was unclear whether the NO effect was due to imposition of an oxidative stress or because it was used as a substrate for ammonium oxidation. This could not be assessed because nitrite was in excess during the enrichment. Hence, to investigate whether NO is consumed, nitrite was systematically depleted in R2 (Phase II) while dosing the same amount of NO (Phase I). After 76 days of nitrite depletion, nitrite was gradually reintroduced in Phase III (Fig. 4).

The effect of nitrite depletion (Phase II) and repletion (Phase III) on the changes in (A) anammox bacteria community of OTUs affiliated to Ca. B. caroliniensis ( ) and Ca. B. sinica (
) and Ca. B. sinica ( ) and Ca. Jettenia (
) and Ca. Jettenia ( ) in suspended (highlighted as “Granules in suspension” on the x-axis) and attached growth biomass (highlighted as “biofilm on the wall” on the x-axis), and the NO consumption rate (NCR,
) in suspended (highlighted as “Granules in suspension” on the x-axis) and attached growth biomass (highlighted as “biofilm on the wall” on the x-axis), and the NO consumption rate (NCR,  ) of R2, fed with synthetic waste water with ammonium, nitrite and continuous supply of nitric oxide. Influent nitrite (NO2,
) of R2, fed with synthetic waste water with ammonium, nitrite and continuous supply of nitric oxide. Influent nitrite (NO2,
 ) was adjusted from normal (Phase I), to depletion (Phase II) and repletion (Phase III). The relative abundance of total anammox bacteria is highlighted as area plot (
) was adjusted from normal (Phase I), to depletion (Phase II) and repletion (Phase III). The relative abundance of total anammox bacteria is highlighted as area plot ( ). FISH images were taken with Ca. B. sinica in cyan and Ca. B. caroliniensis in magenta during Phase I (B) and Phase III (C) from crushed granules.
). FISH images were taken with Ca. B. sinica in cyan and Ca. B. caroliniensis in magenta during Phase I (B) and Phase III (C) from crushed granules.
Reducing the nitrite concentration resulted in a decrease in the relative abundance of Ca. B. sinica in suspension coincident with an increase in NO consumption rate (NCR) while a slight increase was observed for both Ca. B. caroliniensis and Ca. Jettenia in Phase II (Fig. 4). The presence of Ca. B. caroliniensis, absent in Phase I, was also detected through FISH analysis in Phase II (Fig. 4). Batch activity tests conducted during Phase II also showed a decline in the nitrite-dependent ammonium removal rate from 1352 mg N g MLVSS−1 day−1 prior to nitrite depletion (Phase I) to 681 mg N g MLVSS−1 day−1 after nitrite depletion (Phase II). Nevertheless, the overall activity in R2 was still higher than that in R1 even with a specific nitrite-dependent ammonium removal rate of 575 mg N g MLVSS−1 day−1 (Fig. 5). Under normal operation (Phase I), the NO-dependent ammonium oxidation in the absence of nitrite in both the reactors was insignificant, further supporting the hypothesis that nitrite rather than NO is the preferred electron acceptor and ammonium removal cannot be achieved by Ca. B. sinica via direct coupling to NO reduction. In contrast, the ammonium oxidation rates with NO in the absence of nitrite increased more than five times in R2 at 440 mg N g MLVSS−1 day−1 after nitrite depletion in Phase II compared to 33 and 80 mg N g MLVSS−1 day−1 in R1 and R2, respectively in Phase I under normal operation (Fig. 5). This suggests the selection of anammox bacteria species capable of utilizing externally supplied NO to oxidize ammonium.

Batch activity experiments with (i) NH4 + NO2, (ii) NH4 + NO2 + NO, (iii) NH4 + NO in control reactor R1 and experimental reactor R2 (a) before nitrite limitation (phase I), (b) under nitrite limitation (phase II) and (c) after nitrite repletion (phase III).
At the start of experimental Phase III, Ca. B. caroliniensis was found to be in higher relative abundance than Ca. B. sinica in biofilms forming on the wall of the reactor (Fig. 4). Ca. Jettenia also showed a recovery, albeit at low abundance (observed in wall samples) in the absence of nitrite (Fig. 4). While it cannot be confirmed whether this was a consequence of nitrite depletion in Phase II, the relative abundances of Ca. B. caroliniensis and Ca. Jettenia in the biofilm collected from wall, were higher than under normal operation (Phase I in Fig. 6). A clear reversal from Ca. B. caroliniensis to Ca. B. sinica was observed once nitrite was reintroduced between 650 and 680 days (Fig. 4). Similar to that detected in Phase II, the increase in relative abundance of Ca. B. sinica coincided with the recovery of nitrite-dependent ammonium oxidizing activity of 1163 mg N g MLVSS−1 day−1 which is comparable to that in Phase I (1352 mg N g MLVSS−1 day−1) (Fig. 4). In addition, the NO-dependent ammonium oxidizing activity also decreased from 440 (Phase II) to 102 (Phase III) mg N g MLVSS−1 day−1 (Fig. 5), further suggesting that increased NO consumption is likely linked to the increased abundance of Ca. B. caroliniensis or Ca. Jettenia or both. An extended period under nitrite limitation could have further enhanced the recovery of Ca. Jettenia and Ca. B. caroliniensis to outcompete Ca. B. sinica. Nevertheless, this part of the study supports a link between nitrite and NO, species selection and their preferred growth mode.

Distribution of dominant OTUs affiliated to Ca. B. sinica and Ca. B. caroliniensis, along with other anammox bacteria annotated to Brocadiaceae and non-anammox bacteria on the wall (outer layer) and in suspension (inner core) of (A) R1 at day 289 (B), R2 at day 265 (C) and R3 at day 266 (D). The detailed microbial community composition can be found in Supplementary Fig. S3, Supporting Information.
Prevailing anammox bacteria taxa exhibit different biomass morphologies
The predominant anammox bacteria also displayed a distinct preference for attached growth under the various enrichment conditions. Anammox bacteria biomass was present mainly as suspended granules in R1 and R2 while biofilms attached to the reactor surface dominated in R3 (Supplementary Fig. S2). Following the transfer of biofilm from the wall of the reactor into suspension (as indicated by dotted line in Fig. 1A–C), a greater increase in the MLVSS of R3 (around 1.5 g L−1 at day 230) was observed compared to the other two reactors (less than 0.3 g L−1 at days 160, 209, and 258 in R1 and 153, 216, and 253 in R2), suggesting more attached biofilm growth in the primary effluent-fed reactor (R3, Fig. 1C). In addition, following the reintroduction of nitrite into R2 during experimental Phase III, Ca. B. sinica showed a downward trend, more evident in the biofilm samples collected from the walls of the reactor (Fig. 4) than in suspension (p = 0.038). The difference in relative abundance between wall and suspension samples indicates a preference of Ca. B. caroliniensis for attached growth. There was no significant difference between anammox bacteria populations of biomass samples collected from the wall and suspension in R1 and R2, with Ca. B. sinica as the dominant anammox bacteria. However, the relative abundance of predominant Ca. B. caroliniensis was four times higher in the biomass collected from the wall than in suspension for R3 (Fig. 6), further indicating that this species has a tendency towards attached growth (Fig. 6). While granules were found to form in R1 and R2 (Fig. 3A,B), with an average particle size of 1.52 and 1527.9 ± 0.078 μm (Supplementary Table S2) respectively with R = 0.35 (p = 0.006) between the two reactors, the morphology of aggregates in R3 was more floc-like (Fig. 3C), with a particle size of 310.4 ± 0.2 μm (Supplementary Table S2) with R = 0.67 (p = 0.001) compared to the other two reactors. Therefore, it appears that the Ca. B. caroliniensis enriched under domestic wastewater conditions containing organic carbon, cannot support granule maturation and instead forms biofilms on walls, in contrast to Ca. B. sinica which was mainly detected in the granular biomass.
Distinct heterotrophic communities co-developed with specific anammox bacteria taxa
The various enrichment strategies also led to discrete non-anammox bacteria communities in all three reactors, with higher abundances of non-anammox bacteria OTUs in R3 compared to R1 and R2 (Figs S1G, H & I). Supplementation of carbon source increased richness (Supplementary Table S1) in the microbial community in R3 (Chao1: 614 ± 42), but selectively enriched for non-anammox bacteria community (Simpson 1-D: 0.04 ± 0.01) rather than anammox bacteria (Simpson 1-D: 0.75 ± 0.17). Lower richness was observed when nitrite and ammonium were used as main substrates in R1 and R2 (Chao1: 233 ± 31 and 249 ± 32 respectively). Supplementation of NO led to slightly higher richness but lower evenness in R2 than R1. Ca. B. sinica correlated positively with OTUs affiliated to unclassified Fimbriimonadia (phylum Armatimonadetes) and unclassified Anaerolineaceae (phylum Chloroflexi) in R1 (Spearman’s rho > 0.8, p < 0.001). While the relative abundance of non-anammox bacteria was reduced in the presence of NO, the same correlation was nonetheless observed in R2. However, in R3, the non-anammox bacteria community was replaced by OTUs affiliated to unclassified Comamonadaceae (phylum Proteobacteria) and unclassified Bacteroidetes and a negative correlation was observed between unclassified Bacteroidetes and Ca. B. caroliniensis (Spearman’s rho: − 0.7, p < 0.001). Notably, taxa affiliated to the Comamonadaceae family were nearly absent in R1 and R2, while they remained the dominant heterotrophic community in biomass both attached to the reactor wall and in suspension in R3, perhaps suggesting that these taxa have metabolic interactions with Ca. B. caroliniensis and play a role in biofilm formation (Supplementary Fig. S3).
Discussion
N load and/or concentration, and organic carbon availability drive anammox bacteria species selection
Describing factors that drive anammox bacteria niche differentiation is required to select for populations with desired physiological and growth properties. This can enhance process control and operational stability. Anammox bacteria OTUs associated with Ca. Brocadia, Ca. Kuenenia, Ca. Anammoxoglobus, and Ca. Jettenia OTU have all been detected in waste water treatment systems3. Here anammox bacteria populations were selected for from the same activated sludge inoculum. This was achieved by providing different substrates (i.e. ammonium, nitrite, organic carbon and NO) at varying concentrations and loads relevant to mainstream and sidestream PN/A systems. The reproducible enrichment of two key anammox bacteria species, Ca. B. caroliniensis and Ca. B. sinica, was achieved from a single seed activated sludge from the tropics and their preferred ecological niches were described. Ca. B. caroliniensis predominated in all reactors at N loading < 500 mg N L−1 day−1 and lower N concentrations (influent ammonium and nitrite concentration of less than 200 and 250, respectively). An increase in N load and concentration resulted in a succession towards Ca. B. sinica. Ca. B. caroliniensis was also the most competitive species in the presence of organic carbon as shown by their prevalence in R3 under conditions of low N loading.
It is possible that Ca. B. caroliniensis and Ca. B. sinica developed their own niche due to differences in susceptibility to nitrite inhibition since nitrite concentration was increased to raise the N load. A shift in population from Ca. B. caroliniensis to Ca. B. sinica was consistently observed with increasing nitrite concentration beyond 340 mg NO2–N L−1 in the feed (i.e., 170 mg NO2–N L−1 in the reactor). While this is within the range of inhibitory concentrations reported for anammox bacteria of 40 to 400 mg N L−116,17,18,30,31,32, such a shift has not previously been investigated at species level.
An alternative explanation for this shift could be that Ca. B. caroliniensis and Ca. B. sinica have different intrinsic kinetic properties. Using enriched planktonic cells, the nitrite affinity constant and specific growth rate of Ca. B. sinica were determined to be 0.47 mg N L−1 and 0.33 day−1 (corresponding doubling time of 2.1 days)29,33, which is the highest maximum growth rate ever reported for anammox bacteria. This indicates that Ca. B. sinica are r-strategists and would grow at high N loading rates and ammonium and nitrite concentrations33, as observed in our study. However, a similar estimation for Ca. B. caroliniensis is missing. Metagenomic analysis revealed that Ca. B. caroliniensis have multiple copies of nitrite/formate transporters (focA) that provide a competitive advantage at low nitrite concentrations due to a low intrinsic nitrite affinity constant14. This could potentially help them scavenge nitrite from heterotrophic denitrifiers, especially when the competition for nitrite is higher in the presence of organic carbon.
Despite the wide range of environmental conditions applied across the three enrichment reactors, Ca. Brocadia remained the most dominant phylotype throughout the enrichment process, while Ca. Kuenenia and Ca. Anammoxoglobus commonly found in engineered systems were not detected. Although abundances of all anammox bacteria were low in the inoculum, operational conditions, especially the relatively high nitrogen load, contributed to the enrichment of Ca. Brocadia over others34.
NO can select for specific anammox bacteria taxa
NO was provided in R2 to exert oxidative stress and also to potentially select for NO-utilizing anammox bacteria35. While the presence of NO did not appear to suppress the growth of anammox bacteria, it may have provided a competitive advantage to Ca. Jettenia in R2, which increased to a maximum relative abundance of 23% along with Ca. B. caroliniensis at low N loading (Fig. 1). Little is known about the ecological and metabolic drivers of the niche of Ca. Jettenia, probably because they are generally less abundant than other genera of anammox bacteria3. Low nitrite concentrations were shown to encourage the proliferation of Ca. Jettenia over Ca. B. sinica4, consistent with this study. However, Ca. Jettenia was much less abundant than Ca. B. caroliniensis at low nitrite loading rates. Nevertheless, two phylogenetically distant anammox bacteria species, Ca. B. caroliniensis and Ca. Jettenia, were shown to coexist in the same system which supports the previous findings3. However, since microbial community monitoring is based on the 16S rRNA gene only, a bias may be induced when focusing on a single region of the gene. Further validation using a multiple primer sequencing approach could be undertaken to improve the accuracy of quantification 36.
The coevolution of Ca. B. caroliniensis and Ca. Jettenia might also suggest that Ca. B. caroliniensis could utilize NO. A similar nitrite reduction pathway in Ca. B. caroliniensis and Ca. Jettenia was suggested after a nirK homologue was detected from the metagenomic analysis of Ca. B. caroliniensis14,37. The significant increase in NO-dependent ammonium oxidation following nitrite limitation was concomitant with the recovery of Ca. B. caroliniensis and Ca. Jettenia populations. A recent study reported the discovery of nirS in a Ca. Brocadia genome38, however only weak transcripts were found and this observation requires additional validation. It is conceivable that Ca. B. caroliniensis utilize a nirK homologue or a novel nitrite reductase employing the conventional NO-dependent pathway for hydrazine production or possess the ability to switch to a NO-dependent pathway in the absence of nitrite. An alternate pathway for NO production through oxidation of hydroxylamine by a hydroxylamine oxidoreductase (hao)-like protein was indeed detected by Park et al.14 and Irisa et al.39. They proposed that this alternate pathway could potentially be activated under nitrite limitation14. NO was also shown to oxidize ammonium to dinitrogen gas under nitrite limitation in Ca. B. fulgida21 and canonical nirS was absent in its genome40.
In contrast, NO-dependent ammonium oxidation was negligible at significantly higher NO concentrations, further corroborating the assertion that nitrite rather than NO was the preferred electron acceptor and that ammonium removal cannot be achieved by Ca. B. sinica via direct coupling to NO reduction (Fig. 5). While Ca. B. sinica was not completely inhibited in the absence of nitrite, its relative abundance decreased under nitrite limitation as discussed earlier. This supports the study by Oshiki et al.41 demonstrating that Ca. B. sinica does not utilize NO and ammonium for hydrazine synthesis, but instead uses hydroxylamine and ammonium. Shaw et al.42 revealed using 15N-labeling experiments that ammonium was oxidized to dinitrogen via hydroxylamine as intermediate instead of NO in a Ca. Brocadia enrichment culture, with an electrode as electron acceptor. It is noted, however, that it is not possible to assign behaviours to species with absolute confidence in the absence of pure cultures. All anammox bacteria are non-cultivable, and the only option for attributing behaviours to species, and identifying their niches and optimum growth conditions, is phenomenological studies on enrichment reactors. Here, enrichments of between 50 and 80% were achieved, which is high for a population enrichment reactor43; they rank among the highest anammox enrichments that have been achieved in an SBR so far44. An enrichment to 99.5% can be achieved by Percoll density centrifugation45, but the high biomass required hampers resolution of the microbial community at the species level. While membrane bioreactors have been used to enrich planktonic populations of Ca. B. sinica, Ca. Scalindua29 and Ca. K. stuttgartsiensis10, our approach also enriched for species that prefer to grow in biofilms like B. caroliniensis and Ca. Jettenia.
The predominant anammox bacteria species determine biomass retention strategy
High retention of anammox bacteria in reactors is a crucial factor for optimal operation due to the slow growth rate of these bacteria. This can be achieved through biofilm attachment to carriers, formation of granular biomass aggregate and other separation techniques such as membrane filtration to prevent washout of anammox bacteria. The choice of biomass retention in anammox systems may be guided by the growth mode of prevailing anammox bacteria species and the coexisting microbial community under specific operational conditions. In this study, it was demonstrated that the anammox bacteria community and their aggregation states might be distinct under mainstream and sidestream conditions. Ca. B. caroliniensis, likely persisting under mainstream conditions, exhibited a preference for attached biofilm growth. High diversity and abundance of heterotrophic species in R3 was also observed in the presence of complex organic carbon. Particularly, Comamonadaceae remained one of the most abundant heterotrophs in the system both in suspension and in the biofilm. Comamonadaceae are commonly found in biofilm forming communities46,47 suggesting their potential role in assisting biofilm formation. Attached growth of Ca. B. caroliniensis was also observed in a full-scale process treating anaerobic digester liquid supplied with glycerol as the external carbon source14. In this instance, carriers can be used to provide a large surface area to achieve high biomass retention48. Carriers supporting attached biofilm growth can be applied in various configurations, for instance, in rotating biological contactors49, moving bed biofilm reactors50,51 and sequencing batch biofilm reactors52. However, Ca. B. sinica dominated anammox biofilms forming granules (as observed in R1 and R2 operated under side stream conditions), and can be separated physically using a hydrocyclone as with DEMON SBR systems53, lamella separators54 or integrated fixed film activated sludge (IFAS) configurations with a settler55. A particular extracellular protein was found to be highly abundant in the extracellular matrix of the Ca. B. sinica granules which promotes biofilm formation across several length scales due to its ability to phase separate (droplets and gels) and promote adhesion56. This could explain the greater tendency of Ca. B. sinica to self-aggregate (i.e. in the absence of a substratum, unlike Ca. B caroliensis and Ca. Jettenia). Despite the absence of externally supplied organic carbon, heterotrophs belonging to the class Fimbriimonadia (phylum Armatimonadetes) and family Anaerolineaceae (phylum Chloroflexi) proliferated in R1 and R2 to a relative abundance of 10–15%. Gao et al.57 suggested an important role of Anaerolineaceae as cores or carriers for granule formation in anammox sludge and their increase in abundance over time would suggest that they may have supported granulation in both R1 and R2. However, the role of heterotrophic bacteria and their interaction with anammox bacteria cannot be fully uncovered in this study and will require further investigation.
Conclusions
Multiple anammox bacterial species can be enriched from the same activated sludge. Ca. B. caroliniensis dominates at low N loads, both in the presence and absence of organic carbon and under nitrite limitation, and forms attached biofilms; Ca. B. sinica outcompetes it at higher N loads and forms granules; and NO supplementation promotes Ca Jettenia even though it still disappears at high nitrate concentrations. Thus, Ca. B. caroliensis likely dominates in mainstream wastewater anammox processes, where carriers would be the best biofilm retention strategy, Ca B. sinica in sidestream treatments with granules the best retention strategy, and Ca Jettenia is unlikely to be competitive in wastewater treatment systems. Collectively, this study provides insight into understanding the relationship of species selection, growth morphology and process conditions in mainstream and sidestream applications with important implications in process design, control and management of the anammox process at the species level in full scale waste water treatment systems.
Materials and methods
Laboratory-scale sequencing batch reactor set up and operation for anammox bacteria enrichment
Three perspex sequencing batch reactors (SBRs), each with a working volume of 4 L (140 mm internal diameter, 260 mm height) were used here and stirred with a top-mounted impeller (30 mm radius) at 200 rpm. They were seeded with activated sludge from a full-scale water reclamation plant (WRP) performing biological nutrient removal and treating domestic and industrial used waters in Singapore. R1 and R2 were fed with synthetic medium devoid of organic carbon while R3 received primary effluent collected from the WRP once a week comprising complex organic carbon source. Synthetic medium was prepared as (g L−1): KHCO3 1.25, KH2PO4 0.025, CaCl2·6H2O 0.3, MgSO4·7H2O 0.2 and FeSO4·7H2O 0.025 with gradual increase in ammonium (from 30 to 280 mg N L−1) and nitrite concentration (39–350 mg N L−1) and 1.25 mL L−1 of trace mineral solution as described by van de Graaf et al.58. Argon/CO2 (95/5%) was sparged continuously at 25 mL min−1 throughout the anoxic phase to prevent ingress of oxygen in R1 whereas R2 was sparged with both Argon/CO2 and NO with a combined sparging rate of 25 mL min−1 to a final NO gas phase concentration of 400 ppmv to impose oxidative stress. The selected concentration of 400 ppmv was lower than the previously reported tolerance threshold of 600 ppmv for anammox bacteria22. R1 and R2 were operated in cycles of 12 h, each cycle comprised of 5 min of feeding, 108 min of anoxic cycle, 67 min of settling and decanting. Initial ammonium and nitrite concentrations were maintained at 20 mg N L−1 within the reactor for the first seven weeks resulting in a hydraulic retention time (HRT) of 24 h (two litres of synthetic waste water were fed into the reactor each cycle). Upon achievement of 100% ammonium removal, NH4+ and NO2− concentrations in the feed were then increased in steps of 20 mg N L−1. Influent NO2−: NH4+ was maintained at a molar ratio of 1.3 close to theoretical stoichiometry59. HRT was gradually decreased from 24 to 16 h for R1 and from 24 to 12 h for R2, in accordance with the N removal capacity. Thus, R2 was operated at a higher N loading rate compared to R1 due to shorter cycle time concomitant with higher N removal rates in R2. The pH was not controlled in either R1 or R2 and varied between 7.2 and 7.8.
R3 was operated in cycles of 8–12 h, each cycle comprised of 2 h feeding, 5–9 h anoxic phase (depending on the cycle length applied), and 1 h of settling and decanting. Before settling, the reactor was sparged with Argon/CO2 for 5 min to strip out the nitrogen gas produced during the anoxic phase to improve sludge settle-ability. In each feeding period, 2 L of primary effluent supplemented with nitrite was added, resulting in a HRT of 16–24 h. Nitrite was adjusted according to the ammonium concentration at a molar ratio of 2:1 and stored in a chiller at 4 °C to minimize degradation. The nutrient composition of primary effluent was measured after addition of nitrite with average values shown in Supplementary Table S1. Slow feeding of 2 h was applied to minimise oxygen introduction and temperature shock from primary effluent stored in the chiller. The pH of the reactor was not controlled and varied between 7.6 and 8.5 due to denitrification activity. For enrichment purpose, the SRT was not controlled in all three reactors whereby sludge loss only occurred through sampling for nutrient and solids analyses (SRT estimated to be > 20 days).
A heating jacket was connected to maintain the SBR at 35 ± 0.05 °C for R1 and R2 and 33 ± 1 °C for R3. Dissolved oxygen (DO) concentration and pH were continuously monitored using Mettler Toledo InPro6050 DO sensor and Mettler Toledo-InPro 3250i pH sensor, respectively. Samples were collected periodically at the end of the cycle and filtered immediately with 0.2 µm filters for nutrient analyses. Mixed liquor samples were collected in the middle of the anoxic phase for DNA extraction. To determine the microbial composition of the biofilm on the reactor surface, biomass samples from the wall was collected from three random locations after draining the reactor at day 289 for R1, day 265 for R2 and 266 for R3. Both collected suspended and biofilm samples were snap-frozen in liquid nitrogen and stored at − 80 °C until extraction. Biofilm on the surface of the reactor was periodically cleaned to determine the concentration of total mixed liquor suspended solids (MLSS) and the volatile fraction (MLVSS) and the proportion of biomass that was in suspension versus attached growth. Suspended biomass samples were also collected for particle size analysis, and light microscopy imaging after stable enrichment was attained.
NO depletion and repletion experiment
To further validate the effect of NO as a selection pressure for anammox bacteria species selection, R2 was subjected to gradual nitrite depletion and repletion while maintaining availability of NO as an electron acceptor across three experimental phases after stable operation was attained: Phase I -normal operation prior to nitrite depletion, ammonium and nitrite concentration in the feed were 280 and 350 mg N L−1, respectively with continuous supply of NO at 400 ppmv in the gas phase (before day 563); Phase II-nitrite limited operation (day 564–640) whereby nitrite was reduced stepwise from 50 to 0 mg NO2–N L−1 while ammonium and NO were maintained at 50 mg NH4–N L−1 and 400 ppmv, respectively; Phase III (day 640–687) nitrite was gradually reintroduced from 0 to 70 mg NO2–N L−1 with the aforementioned ammonium and NO concentrations in Phase II. Suspended biomass samples were collected twice a week from the mixed liquor throughout the experiment however biofilm attached to the wall were only collected from Phase III due to the limited amount of wall biomass. At each experimental phase, batch activity tests were conducted in triplicate with 80 mg NH4+–N L−1, 100 mg NO2–N L−1 and/or 400 ppmv NO in gas phase under the following conditions with (i) ammonium and nitrite only, (ii) ammonium, nitrite and NO, and (iii) presence of ammonium and NO only. The anammox activity of R1 under normal operation with ammonium and nitrite supplied as substrate served as the control. In all batch activity tests, mixed liquor samples were collected every 30 min and filtered through 0.22 µm Milipore filters for nutrient analysis.
Chemical analysis
All samples collected for nutrient analysis were measured for ammonium, nitrite and nitrate. Ammonium was measured using Hach® kits, nitrate and nitrite were analyzed using ion chromatography (Prominence, Shimadzu). MLSS and MLVSS were analyzed according to the standard methods60. NO was measured in the gas phase using an online chemiluminescence analyzer (Model: 42i, Thermoscientific). Particle size analysis was carried out using laser diffraction particle size analyzer (Model: SALD-MS30, Shimadzu), ANOSIM analysis were carried out on the particle size measurements on samples collected from different reactors.
Suspended biomass was collected from each reactor and subject to size analysis. 1 mL biomass was dispersed on the surface of petri dish, and images were taken by AxioObserver Z1 inverted epifluorescent microscope (Leica, Germany) with brick/seal function. Images were then analyzed with image J61 Analyze particles function.
Microbial community profiling
Genomic DNA was extracted from biomass samples using FastDNA™ SPIN Kit for Soil (MP Biomedicals, USA) with optimization according to Albertsen et al. (2015). Paired-end 16S rRNA gene amplicon sequencing was conducted by DNAsense (http://dnasense.com/) at the Aalborg University (Denmark) with primer set 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) (Caporaso et al. 2011) by Illumina Miseq platform as described in Law et al. (2016). Detailed data analysis can be found in supporting information (SI).
Clone library construction and phylogenetic tree analysis
To further confirm the species level taxonomy identification, four 16S rRNA gene clone libraries were constructed from samples collected on days 74, 280 from R1 and 85, 270 from R2. Sequences obtained from clone library and 16S rRNA amplicon sequencing were used to generate a phylogenetic tree by ARB. Methods for clone library and phylogenetic tree construction are described in detail in SI.
Fluorescent in situ hybridization
Suspended biomass samples were collected and fixed with 4% paraformaldehyde (PFA) overnight. After washing with 1 × phosphate-buffered saline (PBS, 130 mM sodium chloride, 10 mM sodium phosphate buffer, pH 7.2), the biomass samples were stored with 1:1 100% ethanol:1 × PBS at − 20 °C. FISH was performed on crushed biomass samples according to the method described by Daims et al.62 with probes listed in Table 1. The slides were viewed using a Zeiss LSM 780 inverted confocal microscope (Carl Zeiss, Jena, Germany).
Data availability
All raw 16S rRNA amplicon sequences used in this manuscript are available in NCBI under Bioproject PRJNA604076. Contact the corresponding author Thomas Seviour (twseviour@bce.au.dk) to request data from this study.
References
Ali, M. et al. Physiological characterization of anaerobic ammonium oxidizing bacterium Candidatus, “Jettenia caeni”. Environ. Microbiol. 17, 2172–2189 (2015).
Nikolaev, Y. A. et al. Candidatus “Jettenia moscovienalis” sp. nov., a new species of bacteria carrying out anaerobic ammonium oxidation. Microbiology 84, 256–262 (2015).
Sonthiphand, P., Hall, M. W. & Neufeld, J. D. Biogeography of anaerobic ammonia-oxidizing (anammox) bacteria. Front. Microbiol. 5, 399 (2014).
Zhang, L. et al. Microbial competition among anammox bacteria in nitrite-limited bioreactors. Water Res. 125, 249–258 (2017).
Ding, S. et al. Ecological characteristics of anaerobic ammonia oxidizing bacteria. Appl. Microb. Biotechnol. 97, 1841–1849 (2013).
Kartal, B. et al. Candidatus “Anammoxoglobus propionicus” a new propionate oxidizing species of anaerobic ammonium oxidizing bacteria. Syst. Appl. Microbiol. 30, 39–49 (2007).
Kartal, B. et al. Candidatus “Brocadia fulgida”: An autofluorescent anaerobic ammonium oxidizing bacterium. FEMS Microbiol. 63, 46–55 (2008).
Oshiki, M., Shimokawa, M., Fujii, N., Satoh, H. & Okabe, S. Physiological characteristics of the anaerobic ammonium-oxidizing bacterium Candidatus “Brocadia sinica”. Microbiology 157, 1706–1713 (2011).
Park, H., Sundar, S., Ma, Y. & Chandran, K. Differentiation in the microbial ecology and activity of suspended and attached bacteria in a nitritation-anammox process. Biotechnol. Bioeng. 112, 272–279 (2015).
van der Star, W. R. et al. The membrane bioreactor: A novel tool to grow anammox bacteria as free cells. Biotechnol. Bioeng. 101, 286–294 (2008).
Park, H. et al. Impact of inocula and growth mode on the molecular microbial ecology of anaerobic ammonia oxidation (anammox) bioreactor communities. Water Res. 44, 5005–5013 (2010).
der Star, W. R. V. et al. Startup of reactors for anoxic ammonium oxidation: Experiences from the first full-scale anammox reactor in Rotterdam. Water Res. 41, 4149–4163 (2007).
Hu, B. et al. Identification and quantification of anammox bacteria in eight nitrogen removal reactors. Water Res. 44, 5014–5020 (2010).
Park, H., Brotto, A. C., van Loosdrecht, M. C. & Chandran, K. Discovery and metagenomic analysis of an anammox bacterial enrichment related to Candidatus “Brocadia caroliniensis” in a full-scale glycerol-fed nitritation-denitritation separate centrate treatment process. Water Res. 111, 265–273 (2017).
Cao, Y. et al. Mainstream partial nitritation and anammox in a 200,000 m3/day activated sludge process in Singapore: Scale-down by using laboratory fed-batch reactor. Water Sci. Technol. 74, 48–56 (2016).
Kimura, Y., Isaka, K., Kazama, F. & Sumino, T. Effects of nitrite inhibition on anaerobic ammonium oxidation. Appl. Microb. Biotechnol. 86, 359–365 (2010).
Lotti, T., der Star, W. V., Kleerebezem, R., Lubello, C. & Loosdrecht, M. V. The effect of nitrite inhibition on the anammox process. Water Res. 46, 2559–2569 (2012).
Strous, M., Kuenen, J. G. & Jetten, M. S. Key physiology of anaerobic ammonium oxidation. App. Environ. Microb. 65, 3248–3250 (1999).
Kartal, B. et al. Molecular mechanism of anaerobic ammonium oxidation. Nature 479, 127–130 (2011).
Strous, M. et al. Deciphering the evolution and metabolism of an anammox bacterium from a community genome. Nature 440, 790–794 (2006).
Kartal, B. et al. Effect of nitric oxide on anammox bacteria. Appl. Environ. Microb. 76, 6304–6306 (2010).
Schmidt, I. et al. Anaerobic ammonia oxidation in the presence of nitrogen oxides (NOx) by two different lithotrophs. Appl. Environ. Microb. 68, 5351–5357 (2002).
Lackner, S., Terada, A. & Smets, B. F. Heterotrophic activity compromises autotrophic nitrogen removal in membrane-aerated biofilms: Results of a modeling study. Water Res. 42, 1102–1112 (2008).
Strous, M., Heijnen, J., Kuenen, J. & Jetten, M. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms. Appl. Microb. Biotechnol. 50, 589–596 (1998).
Hubaux, N., Wells, G. & Morgenroth, E. Impact of coexistence of flocs and biofilm on performance of combined nitritation-anammox granular sludge reactors. Water Res. 68, 127–139 (2015).
Vlaeminck, S. E. et al. Aggregate size and architecture determine microbial activity balance for one-stage partial nitritation and anammox. Appl. Environ. Microb. 76, 900–909 (2010).
Volcke, E., Picioreanu, C., Baets, B. D. & Loosdrecht, M. V. The granule size distribution in an anammox-based granular sludge reactor affects the conversion—Implications for modeling. Biotechnol. Bioeng. 109, 1629–1636 (2012).
Wang, Q. & He, J. Partnering of anammox and denitrifying bacteria benefits anammox’s recovery from starvation and complete nitrogen removal. Sci. Total Environ. 815, 152696 (2022).
Oshiki, M., Awata, T., Kindaichi, T., Satoh, H. & Okabe, S. Cultivation of planktonic anaerobic ammonium oxidation (anammox) bacteria using membrane bioreactor. Microbes Environ. 28, 436–443 (2013).
Bettazzi, E., Caffaz, S., Vannini, C. & Lubello, C. Nitrite inhibition and intermediates effects on Anammox bacteria: A batch-scale experimental study. Process Biochem. 45, 573–580 (2010).
Dapena-Mora, A. et al. Evaluation of activity and inhibition effects on Anammox process by batch tests based on the nitrogen gas production. Enzyme Microb. Technol. 40, 859–865 (2007).
Fux, C., Marchesi, V., Brunner, I. & Siegrist, H. Anaerobic ammonium oxidation of ammonium-rich waste streams in fixed-bed reactors. Water Sci. Technol. 49, 77–82 (2004).
Zhang, L. et al. Maximum specific growth rate of anammox bacteria revisited. Water Res. 116, 296–303 (2017).
Costa, M. C. M. S. et al. Impact of inocula and operating conditions on the microbial community structure of two anammox reactors. Environ. Technol. 35, 1811–1822 (2014).
Hu, Z., Wessels, H. J. C. T., van Alen, T., Jetten, M. S. M. & Kartal, B. Nitric oxide-dependent anaerobic ammonium oxidation. Nat. Commun. 10, 1244 (2019).
Orschler, L., Agrawal, S. & Lackner, S. On resolving ambiguities in microbial community analysis of partial nitritation anammox reactors. Sci. Rep.-UK. 9, 6954 (2019).
Hu, Z., Speth, D. R., Francoijs, K.-J., Quan, Z.-X., Jetten, M. Metagenome analysis of a complex community reveals the metabolic blueprint of anammox bacterium Candidatus “Jettenia asiatica”. Front. Microbiol. 3 (2012).
Yang, Y. et al. Complex microbial nitrogen-cycling networks in three distinct anammox-inoculated wastewater treatment systems. Water Res. 168, 115142 (2020).
Irisa, T., Hira, D., Furukawa, K. & Fujii, T. Reduction of nitric oxide catalyzed by hydroxylamine oxidoreductase from an anammox bacterium. J. Biosci. Bioeng. 118, 616–621 (2014).
Gori, F., Tringe, S. G., Kartal, B., Machiori, E. & Jetten, M. S. The metagenomic basis of anammox metabolism in Candidatus “Brocadia fulgida”. Biochem. Soc. Trans. 39, 1799–1804 (2011).
Oshiki, M., Ali, M., Shinyako-Hata, K., Satoh, H. & Okabe, S. Hydroxylamine-dependent anaerobic ammonium oxidation (anammox) by Candidatus Brocadia sinica. Environ. Microbiol. 18, 3133–3143 (2016).
Shaw, D. R. et al. Extracellular electron transfer-dependent anaerobic oxidation of ammonium by anammox bacteria. Nat. Commun. 11, 2058 (2020).
Seviour, T. W., Lambert, L. K., Pijuan, M. & Yuan, Z. Selectively inducing the synthesis of a key structural exopolysaccharide in aerobic granules by enriching for Candidatus “Competibacter phosphatis”. Appl. Microbiol. Biotechnol. 92, 1297–1305 (2011).
Strous, M., Heijnen, J. J., Kuenen, J. G. & Jetten, M. S. M. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms. Appl. Microbiol. Biotechnol. 50, 589–596 (1998).
Strous, M. et al. Missing lithotroph identified as new planctomycete. Nature 400, 446–449 (1999).
Bereschenko, L. A., Stams, A. J. M., Euverink, G. J. W. & van Loosdrecht, M. C. M. Biofilm formation on reverse osmosis membranes is initiated and dominated by Sphingomonas spp. Appl. Environ. Microb. 76, 2623–2632 (2010).
Douterelo, I., Sharpe, R. & Boxall, J. Bacterial community dynamics during the early stages of biofilm formation in a chlorinated experimental drinking water distribution system: Implications for drinking water discolouration. J. Appl. Microbiol. 117, 286–301 (2014).
Lemaire, R. et al. Mainstream deammonification with ANITA™ Mox process. Proc. Water Environ. Fed. 2014, 2183–2197 (2014).
Pynaert, K., Smets, B. F., Beheydt, D. & Verstraete, W. Start-up of autotrophic nitrogen removal reactors via sequential biocatalyst addition. Environ. Sci. Technol. 38, 1228–1235 (2004).
Agrawal, S. et al. The role of inoculum and reactor configuration for microbial community composition and dynamics in mainstream partial nitritation anammox reactors. Microbiol. Open. 6, e00456 (2017).
Lackner, S. & Horn, H. Comparing the performance and operation stability of an SBR and MBBR for single-stage nitritation-anammox treating wastewater with high organic load. Environ. Technol. 34, 1319–1328 (2013).
Yu, Y.-C., Gao, D.-W. & Tao, Y. Anammox start-up in sequencing batch biofilm reactors using different inoculating sludge. Appl. Microb. Biotechnol. 97, 6057–6064 (2013).
Wett, B. et al. Syntrophy of aerobic and anaerobic ammonia oxidisers. Water Sci. Technol. 61, 1915–1922 (2010).
Lackner, S. et al. Full-scale partial nitritation/anammox experiences—An application survey. Water Res. 55, 292–303 (2014).
Malovanyy, A., Trela, J. & Plaza, E. Mainstream wastewater treatment in integrated fixed film activated sludge (IFAS) reactor by partial nitritation/anammox process. Bioresour. Technol. 198, 478–487 (2015).
Seviour, T., Wong, L. L., Lu, Y., Mugunthan, S., Yang, Q. H., Segaran, U., Bessarab, I., Liebl, D., Williams, R., Law, Y., Kjelleberg, S. Phase transitions by an abundant protein in the anammox extracellular matrix mediate cell-to-cell aggregation and biofilm formation. mBio. 02052–20 (2020).
Gao, D., Liu, L., Liang, H. & Wu, W.-M. Aerobic granular sludge: Characterization, mechanism of granulation and application to wastewater treatment. Crit. Rev. Biotechnol. 31, 137–152 (2011).
de Graaf, A. A. V., de Bruijn, P., Robertson, L. A., Jetten, M. S. & Kuenen, J. G. Autotrophic growth of anaerobic ammonium-oxidizing micro-organisms in a fluidized bed reactor. Microbiology 142, 2187–2196 (1996).
Kartal, B. et al. How to make a living from anaerobic ammonium oxidation. FEMS Microbiol. Rev. 37, 428–461 (2013).
Eaton, A., Clesceri, L. S., Rice, E. W., Greenberg, A. E., Franson, M. APHA: Standard Methods for the Examination of Water and Wastewater. Centennial Edition. (APHA, AWWA, WEF, 2005).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671–675 (2012).
Daims, H., Nielsen, J. L., Nielsen, P. H., Schleifer, K. H. & Wagner, M. In situ characterization of nitrospira-like nitrite-oxidizing bacteria active in wastewater treatment plants. Appl. Environ. Microb. 67, 5273–5284 (2001).
Acknowledgements
This research was supported by the Singapore National Research Foundation and Ministry of Education under the Research Centre of Excellence Programme, and by a program grant from the National Research Foundation (NRF), project number 1301-IRIS-59. We would like to acknowledge the assistance of Mr. Larry Liew and staff from the Public Utilities Board (PUB) of Singapore for the weekly collection of primary effluent and Mr. Eganathan Kaliyamoorthy for his help in conducting solids analysis.
Author information
Authors and Affiliations
Contributions
T.S., Y.Y.L. and Y.L. wrote the manuscript and designed experiments, R.W., Y.L. and K.A. analysed sequencing data, G.N., T.N. and S.T. maintained and collected experimental data, Y.L. and G.N. analysed the experimental data. All authors contributed to editing the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lu, Y., Natarajan, G., Nguyen, T.Q.N. et al. Controlling anammox speciation and biofilm attachment strategy using N-biotransformation intermediates and organic carbon levels. Sci Rep 12, 21720 (2022). https://doi.org/10.1038/s41598-022-26069-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-26069-2
This article is cited by
-
Mechanisms of microbial co-aggregation in mixed anaerobic cultures
Applied Microbiology and Biotechnology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
