
Abstract
Marine turtles were common in the subtropical Upper Cretaceous epi-continental seas that once washed the coasts of the ancient European archipelago. But unlike its contemporaneous faunas from North America, in Europe no taxon surpassed the 1.5 m shell-length. Here, the remains of a new large marine turtle, Leviathanochelys aenigmatica gen. et sp. nov., from the Middle Campanian of the Southern Pyrenees are described. Anatomical and histological evidence concur in identifying the specimen as a basal chelonioid. The new taxon autapomorphically differs from other marine turtles by possessing an additional process on the anteromedial side of the pelvis, and an acetabulum directed strongly ventrally. Based on the pelvis size, it is likely that Leviathanochelys was as large as Archelon, thus becoming one of the largest marine turtles found to ever exist. The large body size of the new taxon could have evolved as a response to the unique habitat conditions of the European Cretaceous archipelago seas. The presence of the accessory pubic process further suggests the occurrence of an additional insertion point of the Musculus rectus abdominis, which together with the paleohistologic evidences support the hypothesis that the new taxon had an open marine pelagic lifestyle.
Similar content being viewed by others

Introduction
Pan-Chelonioidea is a monophyletic clade of cryptodiran Testudines (if considering Protostegidae at the base of the superfamily) that comprises both extinct and extant marine turtles, including the largest turtles that have ever sailed the seas such as the protostegids Archelon or Protostega. The clade is characterized by having several anatomical adaptations to marine lifestyle, such as the modification of the autopodials in paddle-like limbs1,2, cranial modification for exceeding-salt removal3, and the reduction of the shell ossification, together with the development of fontanelles in both carapace and plastron4.
According to the most recent analyses5, the superfamily Chelonioidea only includes Dermochelyidae and Cheloniidae, while the extinct Toxochelyidae and Ctenochelyidae represent stem chelonioids. Anyhow, all of these groups emerged during the Early Cretaceous and peaked in diversity during the Latest Cretaceous4,6,7,8. On the other hand, the phylogenetically controversial protostegids6 experienced a rapid radiation during the Early Cretaceous, becoming globally widespread, ecologically specialized and taxonomically diverse during said period9,10, but become extinct at the end of the Mesozoic era. It is worth noting that gigantic forms mainly occurred during the Campanian in the Western Interior Sea of North America11, to the exception of the Moroccan Ocepechelon12, the Jordanian Gigantatypus13, and material from Eastern Europe referred to Protostega gigas14. The achievement of such large body size might be likely reflecting some unique habitat conditions or the selection towards large sizes by predatory pressure15,16.
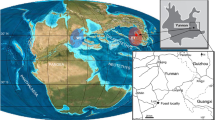
Although European marine deposits have yielded a relatively good fossil record of Late Cretaceous chelonioids17,18, the reports of large taxa are extremely rare19,20,21,22,23. With up to 1.5 m of shell-length, Allopleuron hoffmanni is, to date, the largest Late Cretaceous chelonioid in Europe24,25. Here, we report a new gigantic basal chelonioid, represented by a posterior region of the carapace and a partial pelvic girdle from the Middle Campanian marine deposits of Southern Pyrenees (Fig. 1). Despite the fragmentary nature of the specimen, its inferred body size rivals with that of Archelon, confirming the occurrence of colossal marine turtles in the Late Cretaceous European seas.

Geographic and geological situation of Cal Torrades. The Cal Torrades fossil locality location, respect: (a) the Iberian Peninsula; and (b) the Eastern Pyrenees. (c) Simplified geological map including the locality (white star). (d) Field capture of Cal Torrades outcrop, marking with the star the location of the fossil remains: (e) pelvis; and (f) ilium. (g) Locality stratigraphic column with the geological materials and fossil remains. Modified from Costantino and Angelini26, Vidal27 and free access digital maps of the Institut de Cartografia i Geologia de Catalunya (ICGC; http://www.icc.cat/vissir3/).
Geological setting
During the last 20 years, our knowledge on the Campanian–Maastrichtian vertebrates from the Southern Pyrenees has increased significantly28,29. However, this is mainly restricted to terrestrial environments, and therefore little is known about the marine tetrapods that once inhabited the marine realm of this part of the Iberian Peninsula. Subsequently, here we describe the first locality yielding marine vertebrate remains from the end-Cretaceous of the Southern Pyrenees.
Discovered on July 2016, the locality of Cal Torrades is placed between the Serra d’Aubenç and the Serra de Carreu, outcropping near the small villages of Pera-Rua and Valldarques (Alt Urgell County, Southern Pyrenees; Fig. 1a–e). From a geological viewpoint, Cal Torrades is situated within the lower part of the Perles Formation, whose main lithologic components consist of marls and marly limestones30,31. The lower part of the Perles Formation contains abundant invertebrate macro (—e.g. echinoderms, sponges, lamellibranch bivalves) and microfossils (hyaline benthonic and planktonic foraminifera32, the analyses of which –mainly the benthonic foraminifera assemblage (see33)—allow to establish the age of the new fossil locality as Middle Campanian.
The stratigraphic section at Cal Torrades consists of an alternation between grey marls and sandstone levels (Fig. 1c–e). The locality rests upon a carbonated sandstone bar of 5 m in thickness, whose uppermost part is the base of the studied section (Fig. 1d,e). It contains abundant invertebrate fossil remains such as well-preserved ostreids, pectinid bivalves, hexactinellid sponges, echinoderms, horizontal burrows and ramified bryozoans. The chelonioid remains were found 20 cm over the top of the sandstone base (Fig. 1c–g). A nearly uniform sequence of marls is developed above the sandstone layer. Overall, the stratigraphic section can be defined as slight coarsening-upward sequence (Fig. 1e). The occurrence of a well-developed marly sequence, locally alternated with fine sandstone levels, likely indicates a low energetic offshore depositional environment. On the other hand, the presence of a thick sandstone bar with abundant organisms at the base of the stratigraphic section, rather suggests a relatively low water column. The combination of geological and fossiliferous evidence allows inferring a transgressive depositional sequence, from a near shore environment to a complete offshore environment.
Although a more detailed study is required, preliminary interpretations suggest that the sedimentological sequence at Cal Torrades may represent one of the last transgressive pulses, before the regressive sequence represented by the Upper Cretaceous Gresos d’Areny Formation29 in the Southern Pyrenees. The benthonic macroforaminifera assemblage recovered in the strata immediately above to the Perles Formation suggests a bathymetric depth between 20 and 80 m34,35,36. This interpretation concurs with the absence of high-energy sedimentary structures at the outcrop, suggesting that the depositional environment would be, at least, under the storm-affectation level. Thereby, it is most likely that the sedimentological sequence at the Cal Torrades locality was established in a marine mid-ramp environment, as well as its lateral equivalent formations (e.g. Terradets Formation; see32,35).
Results
Systematic palaeontology
Testudines Batsch, 1788.
Cryptodira Cope, 1868.
Chelonioidea Baur, 1893.
Leviathanochelys aenigmatica gen. et sp. nov.
urn:lsid:zoobank.org:act:28CDDCDB-AC31-45B6-98B1-448E3282A041.
urn:lsid:zoobank.org:act:2B8F389C-6437-450F-812C-E7CD8EFF07A2.
Etymology
The generic name is composed of the following words: Leviathan, in reference to the Biblical marine beast, in allusion to the body size of the new species; and chelys, Latinized name from the ancient Greek χέλυς (“khélūs” meaning turtle in feminine gender). The specific nomination aenigmatica, Latinized adjective from the Greek noun αἴνιγμα (“aínigma” meaning enigma, conundrum or riddle) is in reference to the peculiar anatomical characteristics of its pelvis and carapace.
Holotype
MCD9884. Posterior portion of the carapace including the neural plates 5–8, both left and right fragmentary costals 5–8 and a putative vertebral centra, nearly unidentifiable (MCD9884a); and a partial pelvic girdle, including: the left pubis (MCD9884b); right pubis (MCD9884c); left ischium (MCD9884d); right ischium (MCD9884e); left ilium (MCD9884f); and right ilium (MCD9884g).
Type locality and age
Cal Torrades, Coll de Nargó (Lleida Province, Catalonia, North-eastern Spain). Lower part of the Perles Formation, Middle Campanian, Upper Cretaceous35.
Diagnosis
Large-sized basal chelonioid defined by the following and unique combination of characters: reduction of the costal plates ossification without a sutural contact between costals and peripherals; carapacial plate margins (costals 5–8 and neurals 5–8) finely sutured; hexagonal/octagonal neural plates, longer than wide, that prevent the costals 6–7 from contacting one another; posterior costal plates that are rectangular-shaped, much wider than long; oval articular area of the ilium, located near the lateral margin of the right costal 8; H-shaped pelvis; enlarged and flat lateral pubic process; conspicuously ornamented, textured surface surrounding the acetabular region; extremely elongated iliac neck; and the absence of carapacial scute sulci, keels, or ornamentation on the external part of the carapace, and absence of the ilium’s posterior notch. Leviathanochelys aenigmatica is further diagnosed by having two autapomorphic characters as follows: accessory process on the anteromedial margin of the pubis; and acetabulum strongly ventrolaterally directed.
Nomenclatural acts
The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:8288E740-AE81-4F71-8931-86A730182034. The online version of this work is archived and available from the following digital repositories: PubMed Central and CLOCKSS.
Description
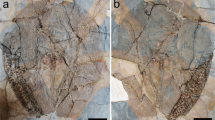
Only a posterior fragment of the carapace is preserved (Fig. 2a,b), consisting of the remains of the neurals 5–8, and both right and left costals 5–8. The smooth dorsal surface of the carapace is slightly convex, whereas the ventral one is almost flat. No epidermal scute marks are present (Fig. 2a), and there is no evidence of keel or medial shallow depression along the medial axis of the preserved neural or costal plates (Fig. 2a). The left portion of the carapace is the most complete, being their lateral edge slightly sinuous and smooth. The preserved costal plates are subrectangular, much wider mediolaterally than long anteroposteriorly, and finely sutured (Fig. 2a). Their distal edges are slightly sinuous and smooth, which indicates that these plates would have had the lateral extensions rod-shaped to join with the peripheral plates. Viscerally, the right portion of the costal 8 preserves an oval concavity to anchor the ilium to the carapace by ligaments (Fig. 2b, Fig. S1a). The neural series only preserves four elements (Fig. 2a,b): Neural 5 only preserves its posteriormost part, neural 6 is the largest plate and is octagonal-shaped, while neural 7 is hexagonal with short sides in front, and neural 8 is represented by its anteriormost part. It is noteworthy that the neural 7 is highly reduced posteriorly, which would indicate that the total number of elements of the neural series would be equal to or less than nine. In either case, both neurals 6–7 prevent the costals 6–7 from contacting one another along the midline (Fig. 2a,b). Remains of the thoracic vertebrae attachments can be discerned viscerally in both neural plates 6–7 (Fig. 2a,b), highlighting extremely crushed vertebral centra preserved over the neural 7.

Shell and pelvic girdle elements of Leviathanochelys aenigmatica gen. et sp. nov. (a) Dorsal view of MCD9884 with the elements disposed as they were discovered, remarking in white the preserved carapace portion (MCD9884a). (b) Visceral view of the carapace with a superimposed interpretation of the shell elements. (c) Dorsal view of the preserved pelvic girdle without the carapace, and (d) ventral view of the same element with the carapace. Asterisk marks indicate the location of the autapomorphic accessory pubic process. Details of the accessory pubic process in (e) dorsal and (f) ventral view. (g) Close up view of the posteromedial part of the pubes, in ventral (upper picture) and posterior view (lower picture), preserving part of the thyroid fossae separated by a thick bone structure (black arrow). (h) Ventral view of the left acetabulum, illustrating the limits between the pelvic bones. (i) Detail of the outer ornamented surface of the ilium. (j) Histological section of the costal 8 (MCD9884.1), showing a cancellous bone zone between the highly vascularized internal and external cortices. Abbreviations: (ac) Acetabulum; (app) Accessory Pubic Process; (cb) cancellous bone; (co) costal plate; (eco) External Cortex; (ico) Internal Cortex; (il) Ilium; (ils) ilium insertion scar; (isc) Ischium; (il) Ilium; (ne) neural plate; (pb) Pubis; (pbb) pubic bridge; (tf) Thyroid fossa.
As for the preserved pelvic bones (Fig. 2c,d,h, Fig. S1b–f), they form a nearly flat and H-shaped pelvic girdle (Fig. 2c,d). Both pubes are almost complete but lacking most of the anteromedial and posteromedial processes. These bones are flat, smooth, and completely fused to each other. Because of the fragmentary nature of the pelvis, it is not possible to accurately evaluate the expansion of the anteromedial pubic process. The lateral pubic process is flat, square-shaped, and prominent (Fig. 2c–f, Fig. S1b): it extends anterolaterally being deflected about 50º from the sagittal plane of the pubic symphysis (Fig. 2c). A pubic accessory process is located between the lateral and medial pubic processes, which is slightly protruding anteriorly (Fig. 2e,f): it shows a striated pattern on its surface and is slightly convex dorsally and ventrally.
Due to the absence of most of the posterior margins of the pubes, it is not possible to evaluate with confidence if the thyroid fenestra was completely separated (Fig. 2g). However, it is certain that an expanded pubioischiadic bridge would have divided the thyroid fenestrae, at least partially along its medial plane, given that the area for accommodating such process is thicker than the surrounding lateral areas (Fig. 1g). The acetabular contour is oval-shaped and slightly constricted anteroposteriorly (Fig. 2d, Fig. S1b). The acetabulum concavity is completely directed ventrally, and slightly tilted laterally (Fig. 2d). Both the lateral and medial external surfaces surrounding the acetabular region are strongly ornamented with irregular anastomosed ridges. There is no posterior notch in the acetabulum.
Both left and right ilia are partially preserved, but given the fragmentary nature of the right ilium, the following description is mainly based on the best-preserved left ilium (Fig. 2h, Fig. S1c–e). It preserves most of the acetabular region and the iliac neck, which is elongated, and when complete, would have reached an anteroposterior length greater than two times the anteroposterior acetabulum’s length. Ventrally, the medial margin of the iliac neck is nearly straight, while the lateral one is convex, proximally straight, and distally deflected posteriorly. The iliac neck slightly bends dorsomedially (Fig. 2h, Fig. S1c–f). Moreover, its medial, lateral and ventral external surfaces are strongly sculptured with anastomosing anteroposteriorly-oriented ridges (Fig. 2i).
Both ischia are poorly preserved. In fact, only fragments of both left and right ischia, which contribute to the posteromedial region of the acetabulum, are available. According to the preserved graphic documents, it can be stated that the ischium contributed significantly to the acetabulum, and that its main body was likely projected medially (Fig. S1f).
Phylogenetic relationships
The parsimony analyses resulted in 20 most parsimonious trees of 1647 steps in length, with a Consistency Index of 0.250 and a Retention Index of 0.686 (Fig. 3). The Strict Consensus topology recovered Leviathanochelys aenigmatica as the sister taxon of the basal chelonioid Allopleuron hoffmanni (Fig. 3). Nonetheless, it is noteworthy that the only common synapomorphy grouping Leviathanochelys and Allopleuron is “the attachment of the pelvis to shell by ligaments, instead of a strong sutural contact” (ch. 318:0)37. However, this feature is highly plesiomorphic since it represents the typical condition for all Testudinata, to the exception of Pleurodira, Proterochersidae and, maybe also Australochelyidae38,39.

Phylogenetic relationship of Leviathanochelys aenigmatica gen. et sp. nov. Simplified phylogenetic hypothesis of the relationship of Leviathanochelys aenigmatica within Pan-Chelonioidea based of 20 MPT with 1647 steps according to the Strict Consensus topology. Number under main branching nodes correlate with Bremer support values. Taxa are illustrated according to their time-range occurrence, but not to the time-divergence of the nodes which are tentatively placed according to fossil record evidences.
The identification of L. aenigmatica as a pan-chelonioid is supported by the absence of contact between costal and peripheral plates (ch. 212:1), while the recovery of Leviathanochelys and Allopleuron as basal members of the Chelonioidea superfamily is established by: the absence of discernible carapacial scutes (ch. 188)37; and the presence of a partially or completely separated thyroid fenestra (ch. 319)37, a feature that is also shared with Peritresius martini and Erquelinnesia gosseleti taxa.
Despite being out of the scope of the present study to deeply analyse all the phylogenetic relationships of the recovered topology, it is worth noting that our phylogenetic results located Protostegidae as stem Chelonioidea. As previously mentioned, the phylogenetic position of this group of Cretaceous marine turtles is a matter of an intense debate5, nonetheless, our results concur with some of the most recent studies37. On the other hand, Ctenochelyidae is recovered as members of Chelonioidea, being its location more inclusive than in other recent phylogenetic studies5,11.
Discussion
Comparison and taxonomic affinity
The discovery of Leviathanochelys aenigmatica in the Middle Campanian marine deposits of the Cal Torrades locality (Southern Pyrenees) represents the first indisputable gigantic bodied chelonioid ever found in Western Europe. The described specimen clearly belongs to a sea turtle characterized by lacking distinct sculpture of the carapace surface, a clear reduction of the ossification of carapace (e.g. costal plates), carapace and pelvis attached to each other by ligaments, and an H-shaped pelvis. Although the entire neural series cannot be evaluated in L. aenigmatica, the preserved neural 6 is octagonal-shaped, which differs from the coffin-like neural plates (hexagonal with shorter sides anteriorly) of Toxochelys latiremis (taken herein as a representative of Toxochelyidae family40; Fig. S2). Moreover, almost all pelvic features exhibited by the genus Toxochelys are different from those observed in Leviathanochelys (Fig. S3), ruling out any possible relationship between these two taxa.
Leviathanochelys shows a conspicuous ornamentation at the external surface surrounding the acetabular region—a bone texture that has been previously correlated with highly vascularized articular surfaces—that is considered characteristic of derived protostegids and dermochelyids41. However, this trait might be a plesiomorphic feature, as it seems to be present in Oertelia gigantea42 as well. Further, the possibility of the new specimen belonging to Dermochelyidae is also discarded, as dermochelyids tend to reduce their shell ossification3,41 and, consequently, the size of both neural and costal plates. The described specimen from Cal Torrades has no evidence of epidermal scutes in the preserved neural and costal plates, a condition shared with the protostegids Archelon9 and Protostega43,44,45, and the chelonioid Allopleuron25, and which further contrasts with the rest of cheloniids. Among protostegids, L. aenigmatica resembles both Archelon and Protostega genera in having a partially divided thyroid fenestrae, as well as an enlarged and flat lateral pubic process; the latter process, which terminates in square-like shape in Archelon (Fig. 2e, Fig. S3) and is fan-shaped in Protostega43,44,45. Unlike Protostega and Archelon, the pubes of L. aenigmatica are strongly fused to each other; however, this could correspond to an advanced ontogenetic stage, as the degree of pubic fusion changes throughout ontogeny, and reflects different developmental stages of the specimens46. Particularly, the extremely elongated iliac neck9,47 displayed by L. aenigmatica only resembles the condition of Archelon. Nonetheless, the absence of continuous or intermittent keels on neurals, and of serrated margins on the carapacial plates2,44,48 in Leviathanochelys rules out a referral to Archelon or Protostega genera.
According to our phylogenetic results, L. aenigmatica is recovered as a sister taxa of Allopleuron hoffmanni, both being considered basal members of the superfamily Chelonioidea (Fig. 3). However, the two taxa differ from each other in several carapace and pelvic features. Firstly, the lateral pubic process of Leviathanochelys is much more developed and more laterally directed (about 50° regarding the axial plane) than that of the Centro-European taxon25 (35° regarding the axial plane; see Fig. S3). Furthermore, the partially divided thyroid fenestrae of Leviathanochelys are smaller than those of Allopleuron, and the almost ventrally directed acetabulum of Leviathanochelys contrasts with the more ventro-laterally directed one of Allopleuron25. Finally, the Leviathanochelys iliac neck is extremely elongated, with an anteroposterior length at least two times longer than the acetabulum’s length (Fig. S1f), being twice as long as the ratio in Allopleuron.
From the aforementioned, it must be highlighted that Leviathanochelys aenigmatica has at least two potential autapomorphic features, which have not yet been recognized or mentioned in any extinct or extant chelonioid taxa: acetabulum strongly ventrolaterally directed, and accessory process on the anteromedial margin of the pubis. In summary, based on all the current data available, the new taxon presented here shares multiple features with the members of the superfamily Chelonioidea and the phylogenetic analysis recovers Allopleuron hoffmanni as the sister taxa of Leviathanochelys aenigmatica. In any case, the identification of Leviathanochelys aenigmatica as a new taxa is indisputable, and provides invaluable insight on the evolutionary history of chelonioids as a whole.
Body size and palaeobiological inferences
Leviathanochelys aenigmatica stands out among other pan-chelonioids for its colossal body size proportions, which can be inferred from the large size of its pelvis. The maximum width of the pelvis of Leviathanochelys was established at 889 mm (Table S1), which is slightly larger than that of Archelon (810 mm wide45). The anteroposterior length of the pubis of Leviathanochelys (395 mm) is about 119.4% larger than that of Protostega (180 mm44), and only 14% smaller than that of Archelon (460 mm45). Despite there not being any allometric correlation between the pelvis size and the total body length in marine turtles, current data suggest that Leviathanochelys could be as large as Archelon48, achieving a body length of up to 3.74 m.
Aside from being the largest marine turtle ever discovered in Europe, and one of the largest worldwide, the finding of Leviathanochelys strongly suggests that gigantism in marine turtles was acquired independently, by different lineages over time. The exact evolutionary processes that favoured the acquisition of the larger body size observed in Leviathanochelys aenigmatica remain unknown. However, it has been evidenced that the body size of extant marine turtles is related to a combination of environmental factors (i.e. predation pressure, competitive release, temperature) and their ecology (e.g. migration capacity, etc.)16.
The identification of Leviathanochelys as a marine turtle is further supported on the base of the diploe structures exhibited in the histological sample from the left costal plate 8 (see Supplementary Information; Fig. 2j, Fig. S5), which is characteristic of the main lifestyle of testudines8,49,50,51. The degree of organization and transition between the cancellous bone to the external and internal cortex observed in Leviathanochelys somehow resembles that of Archelon49, suggesting a similar lifestyle for both taxa, i.e. an open water pelagic marine lifestyle (see Supplementary Information).
One of the most remarkable anatomic features of Leviathanochelys aenigmatica is the occurrence of an accessory process on the anterior side of the pelvis (Fig. 2c–f). The existence of rugosities and striations around this structure indicate the presence of muscular insertions. Given that this feature is not reported in any other extinct or extant marine turtle, its presence may respond to a very specific function involving the pelvic girdle. Due to its anatomical location and biomechanical function, the pubis is the ankle point of several major muscles involved in the hind-limb mobility and the stabilization of the shell (carapace and plastron1,52,53; Fig. S6). Among them, two muscles have their origin in the anterior part of the pubis: the Musculus rectus abdominis (RA) and the M. puboischiofemoralis54 (PIFI and PIFE). RA originates on the anterior part of the lateral process (Fig. S6); it serves as the main stabilizer of the pelvis and acts as a compressor of the plastron during the expiration-inspiration process54,55. The M. puboischiofemoralis is the largest muscle of the pelvis, and it is divided in the internus (PIFI) and the externus (PIFE) parts. PIFI is the main extensor of the hind limb, while the PIFE is the main adductor of the posterior limb. Both muscles originate near the pubic symphysis and insert on the minor trochanter of the femur1,52,54. Given that the accessory pubic process observed in Leviathanochelys projects anteriorly, and has no signs of muscle scar, this suggests backward extension of muscles; it seems unlikely that it would be related to musculature linked to the mobility of the hind limb. On the contrary, the location and the anterior projection of the pubic accessory process suggest that it was somehow related to the plastron and, in fact, it might represent an adaptation related to the respiratory system.
Although there is still some room for speculation, a plausible hypothesis is that the accessory pubic process could have served Leviathanochelys as an additional insertion point of the RA (Fig. S6). If so, it could have acted as an additional stabilizer of the pelvis or provided additional compression of the plastron. The morphological features as described strongly support the open water pelagic marine life-style interpretation from histological data (Fig. S5). In summary, anatomical, histological, and myological evidence suggest that Leviathanochelys was one of the largest pelagic marine turtles to ever roam the Earth’s oceans.
Conclusions
To date, it was thought that the largest marine turtles to ever sail the oceans, such as the protostegids Archelon and Protostega, were restricted to North America during the latest Cretaceous (Campanian–Maastrichtian). The discovery of the new gigantic and bizarre chelonioid Leviathanochelys aenigmatica from the Middle Campanian marine deposits of the Southern Pyrenees, which rivals in size to Archelon, sheds a light on the diversity of marine turtles and on how the phenomenon of gigantism in these groups was also occurring in Europe. Despite the scarcity and fragmentary nature of the individual, the new evidence not only increases the taxonomic diversity of the Late Cretaceous marine turtle biota in Europe, but also opens a new line of exploration and raises new questions, in order to solve the evolutionary mechanisms and ecological pressures that could have favoured the independent evolution of colossal (> 2.5 m in shell length) marine turtles in multiple lineages, especially during the Late Cretaceous.
Material and methods
Nomenclature and terminology
The anatomical shell nomenclature used throughout the descriptions is based on Zangerl56.
Material and institutional abbreviations
The fossil chelonioid remains from Cal Torrades have been recovered through multiple excavations from 2016 to 202136. Because all the remains were found in association to each other and keeping a certain anatomically consistent position, we infer they belong to a single individual (MCD9884), represented by neurals 5–8 (including a highly damaged vertebral centra preserved over the neural 7), costals 5–8 (MCD9884a), and an almost complete pelvic girdle (MCD9884b–g). The pelvic bones include both left (MCD9884b) and right (MCD9884c) pubes, the left (MCD9884d) and right ischium (MCD9884e), and a partially preserved left (MCD9884f) and very fragmented right (MCD9884g) ilia. MCD Museu de la Conca Dellà, Isona, Catalonia, Spain; ICP Institut Català de Paleontologia Miquel Crusafont, Cerdanyola del Vallès, Catalonia, Spain.
Osteohistological analyses
Palaeohistological analyses were performed on both the carapace and left ilium. The descriptive terminology used in this study follows Scheyer57. Two histological samples were taken from the posterolateral side of the preserved portion of the left costal 8. Following the methodology of Scheyer57, the sectioned planes were on lateral and anteroposterior direction, and therefore preserving the anteroposterior carapace plane. Previous to being mechanically sawed, samples were marked to keep the anatomical orientation of the elements. The thin sections were prepared at the Servei de Laboratori de la Universitat Autònoma de Barcelona (UAB). The bone samples were cut with a Buehler Isomet low-speed saw, followed by a polishing process with carborundum powder over a glass sheet. During the polishing process, the grit particles were decreasing progressively in size, on 600, 800 and 1000 grit. Once finished, the next step for the thin-slide preparation, was to fix the sections to a frosted glass slide using ultraviolet curing glue Loctite 358. Posteriorly, the samples were reduced in thickness using a diamond saw (Buehler, PetroThin) of approximately 80 µm. After that, the thin-sections were passed through multiple graded series of alcohol baths and cleared in Histo-Clear II for 5 min. Lastly, the thin-slides were mounted in a DPX mounting medium. They were observed with a Leica DM 2500 P petrographic microscope under both transmitted and polarized light at the ICP facilities.
Phylogenetic analyses
The phylogenetic analyses performed in the present work are based on the data matrix of Evers et al.37, which is one of the most recent, extensive, and updated phylogenetic datasets on marine turtles, including: 96 taxa, and 355 cranial, shell and postcranial characters. Leviathanochelys aenigmatica was initially coded in Mesquite 3.0458 (See Supplementary File 2), and posteriorly analysed in TNT 1.5 software59. All characters were treated as equally weighted and unordered with the new technology search algorithm of TNT, which enabled tree drifting60 and parsimony ratchet61. The initial level of driven search was set to 30 steps, and the number of times the minimum tree length should be obtained was set to 30 as well. The most parsimonious trees (MPTs) of this analysis were subjected to further tree bisection and reconnection (TBR). TNT was used to calculate absolute Bremer decay indices as a measure of branch support.
Data availability
All data generated or analysed during this study are included in the supplementary information file.
References
Walker, W. F. The locomotor aparatus of testudines. In Biology of the Reptilian, Vol 4, Morphology D (eds Gans, C. & Dawson, W. R.) 1–100 (Academic Press, 1973).
Cadena, E. A. & Parham, J. F. Oldest known marine turtle? A new protostegid from the Lower Cretaceous of Colombia. PaleoBios 32, 1–42 (2015).
Hirayama, R. Oldest known sea turtle. Nature 392, 705–708 (1998).
Weems, R. E. Paleocene turtles from the Aquia and Brightseat Formations, with a discussion of their bearing on sea turtle evolution and phylogeny. Proc. Biol. Soc. Wash. 101, 109–145 (1988).
Joyce, et al. A nomenclature for fossil and living turtles using phylogenetically defined clade names. Swiss J. Palaeontol. 140, 1–45 (2021).
Hirayama, R. Distribution and diversity of Cretaceous chelonioids. In Ancient Marine Reptiles (eds Callaway, J. M. & Nicholls, E. L.) 225–241 (Academic Press, 1997).
Lehman, T. M. & Tomlinson, S. L. Terlinguachelys fischbecki, a new genus and species of sea turtle (Chelonioidea: Protostegidae) from the Upper Cretaceous of Texas. J. Paleontol. 78, 1163–1178 (2004).
Marmi, J. & Luján, À. H. An overview of the threatened phylogenetic diversity of living testudines based on a review of the complex evolutionary history of turtles. In Turtles: Anatomy, Ecology and Conservation (eds Cosgrove, M. J. & Roe, S. A.) 117–150 (Nova Science Publishers, 2012).
Wieland, G. R. Archelon ischyros: A new gigantic Cryptodire Testudinate from the Fort Pierre Cretaceous of South Dakota. Am. J. Sci. 2, 399–412 (1896).
Hooks, G. E. Systematic revision of the Protostegidae, with a redescription of Calcarichelys gemma Zangerl, 1953. J. Vertebr. Paleontol. 18, 85–98 (1998).
Gentry, A. D. The non-protostegid Cretaceous marine turtles of Alabama and their bearing on the evolution of extant chelonioids. The University of Alabama at Birmingham. Doctoral dissertation. 1–282 (2020).
Bardet, N. et al. A giant chelonioid turtle from the Late Cretaceous of Morocco with a suction feeding apparatus unique among tetrapods. PLoS ONE 8, e63586 (2013).
Kaddumi, H. F. A new genus and species of gigantic marine turtles (Chelonioidea: Cheloniidae) from the Maastrichtian of the Harrana Fauna-Jordan. PalArch’s J. Vertebrate Palaeontol. 3, 1–14 (2006).
Danilov, I. G., Obraztsova, E. M., Arkhangelsky, M. S., Ivanov, A. V. & Averianov, A. O. Protostega gigas and other sea turtles from the Campanian of Eastern Europe, Russia. Cretaceous Res. 135, 105196 (2022).
Jaffe, A. L., Slater, G. J. & Alfaro, M. E. The evolution of island gigantism and body size variation in tortoises and turtles. Biol. Lett. 7, 558–561 (2011).
Rodrigues, J. F. M., Olalla-Tárraga, M. Á., Iverson, J. B. & Diniz-Filho, J. A. F. Temperature is the main correlate of the global biogeography of turtle body size. Glob. Ecol. Biogeogr. 27, 429–438 (2017).
Kapuścińska, A. & Machalski, M. Upper Albian chelonioid turtles from Poland. Geobios 48, 385–395 (2015).
Mulder, E., Jagt, J., Kuypers, M., Peeters, H. & Rompen, P. Preliminary observations on the stratigraphic distribution of Late Cretaceous marine and terrestrial reptiles from the Maastrichtian type area (SE Netherlands, NE Belgium). Oryctos. 1, 55–64 (1998).
Fritsch, A. Ueber weitere Funde von Iserosaurus litoralis in Miscellanea Palaeontologica-II. Mesozoica 18–22 (1910).
Nopcsa, F. V. Osteologia Reptilium fossilium et recentium in Fossilium Catalogus, I : Animalia (ed. Diener, C.) 1–291 (Junk, W, 1926)
Kuhn, O. Testudines in Fossilium Catalogus, I: Animalia (ed. Westphal, F.) 1–299 (Gravenhage, 1964).
Karl, H.-V. Übersicht über die fossilen Schildkröten Zentraleuropas (Reptilia, Testudines). Mauritiana (Altenburg). 18, 171–202 (2002).
Kear, B. P., Ekrt, B., Prokop, J. & Georgalis, G. L. Turonian marine amniotes from the Bohemian Cretaceous Basin, Czech Republic. Geol. Mag. 151, 183–198 (2014).
Janssen, R., van Baal, R. R. & Schulp, A. S. On the taphonomy of the late Maastrichtian (Late Cretaceous) marine turtle Allopleuron hoffmanni*. Neth. J. Geosci. 90, 187–196 (2011).
Mulder, E. Comparative osteology, palaeoecology and systematics of the Late Cretaceous turtle Allopleuron hoffmanni (Gray 1831) from the Maastrichtian type area. Pubicaties van het Natuurhistorisch Genootschap in Limburg. 1, 23–92 (2003).
Costantino, D. & Angelini, M. G. Geodetic monitoring applied to a mine area. Appl. Geomatics. 3, 61–74 (2011).
Vidal, A. Troballa d’un os de rèptil a Ca l’Andreu de Valldarques, Coll de Nargó (Alt Urgell). Unpublished report, Servei d’Arqueologia i Paleontologia, Departament de Cultura, Generalitat de Catalunya, 1–17 (2016).
Canudo, J. I. et al. The upper Maastrichtian dinosaur fossil record from the southern Pyrenees and its contribution to the topic of the Cretaceous-Palaeogene mass extinction event. Cretac. Res. 57, 540–551 (2016).
Fondevilla, V. et al. Chronostratigraphic synthesis of the latest Cretaceous dinosaur turnover in south-western Europe. Earth Sci. Rev. 191, 168–189 (2019).
Solé-Sugrañes, L. Gravity and compressive nappes in the Central Southern Pyrenees (Spain). Am. J. Sci. 278, 609–637 (1978).
Solé-Sugrañes, L. Alineaciones y fracturas en el sistema catalán según las imágenes Landsat-1. Tecniterrae. 22, 6–16 (1978).
Caus, E., Rodes, D. & Solé-Gruñanes, LL. Bioestratigrafia y estructura del Cretácico superior de la Vall d'Alinyà (Pirineo oriental, prov. de Lleida). Acta Geológica Hispánica. 23, 107–118 (1988).
Ullastre, J. & Masriera, A. Pedraforca: Estratigrafía y estructura (Pirineo catalán, España). Treballs del Museu de Geologia de Barcelona. 12, 11–52 (2004).
Gräfe, K. U. Late Cretaceous benthic foraminifers from the Basque-Cantabrian basin, northern Spain. J. Iber. Geol. 31, 277–298 (2005).
Villalonga, R. et al. Larger foraminifera and strontium isotope stratigraphy of middle Campanian shallow-water lagoonal facies of the Pyrenean Basin (NE Spain). Facies. 65, 1–23 (2019).
Gil, A. & Castillo, O. Informe de l’actuació preventiva en el jaciment paleontològic de Ca l’Andreu (Cal Torrades) de Valldarques (Coll de Nargó, Alt Urgell) 1–10 (Unpublished Report at Servei d’Arqueologia i Paleontologia, 2021).
Evers, S. W., Barrett, P. M. & Benson, R. B. J. Anatomy of Rhinochelys pulchriceps (Protostegidae) and marine adaptation during the early evolution of chelonioids. PeerJ 5, e6811 (2019).
Sterli, J., de la Fuente, M. S. & Rougier, G. Anatomy and relationships of Palaeochersis talampayensis, a Late Triassic turtle from Argentina. Palaeonto. Abt. A. 281, 1–61 (2007).
Szczygielski, T. & Sulej, T. Revision of the Triassic European turtles Proterochersis and Murrhardtia (Reptilia, Testudinata, Proterochersidae), with the description of new taxa from Poland and Germany. Zool. J. Linn. Soc. 177, 395–427 (2016).
Gentry, A. D. & Ebersole, J. A. The first report of Toxochelys latiremis Cope, 1873 (Testudines: Panchelonioidea) from the early Campanian of Alabama, USA. PaleoBios. 35, 1–10 (2018).
Hirayama, R. & Chitoku, T. Family Dermochelyidae (Superfamily Chelonioidea) from the Upper Cretaceous of North Japan. Trans. Proc. Paleontol. Soc. Japan. New Series. 184, 597–622 (1996).
Karl, H.-V., Biermann, S. & Tichy, G. The oldest true sea turtle of the world, Oertelia gigantea (OERTEL, 1914) n. gen. from the Aptian of Kastendamm near Hanover, Germany. Studia Palaeocheloniologica. 5, 107–128 (2012).
Cope, E. D. On the geology and paleontology of the Cretaceous strata of Kansas. in Preliminary Report on the United States Geological Survey of Montana and portions of adjacent territories. 318–349 (1872).
Wieland, G. R. The osteology of Protostega. Carnegie Inst. 7, 279–304 (1906).
Wieland, G. R. The skull, pelvis, and probable relationships of the huge turtles of the genus Archelon from the Fort Pierre Cretaceous of South Dakota. Am. J. Sci. 9, 237–251 (1900).
Griffin, C. T. et al. Assessing ontogenetic maturity in extinct saurian reptiles. Biol. Rev. 96, 470–525 (2021).
Tong, H., Hirayama, R., Makhoul, E. & Escuillié, F. Rhinochelys (Chelonioidea: Protostegidae) from the Late Cretaceous (Cenomanian) of Nammoura, Lebanon. Atti della Società italiana di scienze naturali e del museo civico di storia naturale di Milano. 147, 113–138 (2006).
Wieland, G. R. Revision of the Protostegidae. Am. J. Sci. 27, 101–130 (1909).
Scheyer, T. M. & Sánchez-Villagra, M. R. Carapace bone histology in the giant pleurodiran turtle Stupendemys geographicus: phylogeny and function. Acta Palaeontol. Pol. 52, 137–154 (2007).
Slater, B. J., Reolid, M., Schouten, R. & Benton, M. J. A new Late Jurassic turtle from Spain: Phylogenetic implications, taphonomy and palaeoecology. Palaeontology. 54, 1393–1414 (2011).
Scheyer, T. M., Danilov, I. G., Sukhanov, V. B. & Syromyatnikova, E. V. The shell bone histology of fossil and extant marine turtles revisited. Biol. J. Lin. Soc. 112, 701–718 (2014).
Zug, G. R. Buoyancy, locomotion, morphology of the pelvic girdle and hindlimb, and systematics of cryptodiran turtles. Miscellaneous Publ. Museum Zool. Univ. Michigan. 142, 1–98 (1971).
Zhu, H. Plastron reduction and associated myology in turtles, and its implications for functional morphology and natural history. Theses, Dissertations and Capstones, Marshall University. 1–292 (2011).
Wyneken, J. The anatomy of sea turtles. U.S. Department of Commerce NOAA Technical Memorandum. NMFS-SEFSC-470, 1–172 (2001).
Carrier, D. R. & Farmer, C. G. The evolution of pelvic aspiration in archosaurs. Paleobiology. 26, 271–293 (2000).
Zangerl, R. The turtle shell. in The biology of the Reptilia. Vol. 1. (eds Gans, C. & Bellasirs, A. D.) 311–319 (Academic Press, 1969).
Scheyer, T. M. Comparative bone histology of the turtle shell (carapace and plastron): implications for turtle systematics, functional morphology and turtle origins. PhD Dissertation, Rheinischen Friedrich-Wilhelms-Universität zu Bonn (Bonn, Germany) (2007).
Maddison, W. P. & Maddison, D. R. Mesquite: A modular system for evolutionary analysis. (2019).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics. 24, 774–786 (2008).
Goloboff, P. A. Analyzing large data sets in reasonable times: Solutions for composite optima. Cladistics. 15, 415–428 (1999).
Nixon, K. C. The parsimony ratchet, a new method for rapid parsimony analysis. Cladistics. 15, 407–414 (1999).
Acknowledgements
The present work has been supported by the Spanish Agencia Estatal de Investigación (CGL2016-76431-P, CGL2016-73230-P, and CGL2017-82654-P) and the Generalitat de Catalunya (CERCA Program). Osteohistological analyses were partially supported by the project ‘Xarxes paleocològiques dels jaciments de dinosaures del Cretaci català’ (‘Paleoecological networks of the dinosaur quarries of the Catalan Cretaceous’) CLT/903/2018/67, funded by the Departament de Cultura of the Generalitat de Catalunya. ÀHL is financially supported through the postdoctoral grant ‘Programa Postdoctoral Beatriu de Pinós de la Secretaria d’Universitats i Recerca del Departament d’Empresa i Coneixement de la Generalitat de Catalunya’ (2019 BP 00154), and the Ministerio de Ciencia e Innovación and the Agencia Estatal de Investigación (Grant I+D+I PID2020-117289GBI00 funded by MCIN/AEI/https://doi.org/10.13039/501100011033/). ASG research is supported by the project VIGEOCULT (PLEC2021-00793) funded by the European Union NextGenerationEU/PRTR. The authors thank the Collections Manager of the Yale Peabody Museum and the institution for granting visual resources for this work, as well as Eduardo Espílez from Fundación Conjunto Paleontológico de Teruel-Dinópolis, for providing key anatomical measures. On the same measure, authors further acknowledge the collaboration of the Servei d’Arqueologia i Paleontologia of the Generalitat de Catalunya in granting permissions for the paleontological excavations. Finally, we thank Kelly A. Vega Pagán (from the ICP) for the linguistic correction of the manuscript.
Author information
Authors and Affiliations
Contributions
O.C-V, À.H.L., and A.S designed the experiment, described the specimen, and performed the phylogenetic analyses. O.C-V. and A.S prepared the figures and performed the histological analysis. O.C-V. and À.G. performed the excavations and file works. All authors discussed the results, wrote, and reviewed the main manuscript text.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Castillo-Visa, O., Luján, À.H., Galobart, À. et al. A gigantic bizarre marine turtle (Testudines: Chelonioidea) from the Middle Campanian (Late Cretaceous) of South-western Europe. Sci Rep 12, 18322 (2022). https://doi.org/10.1038/s41598-022-22619-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-22619-w
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
