
Abstract
The lakeshore site of La Marmotta is one of the most important Early Neolithic sites of Mediterranean Europe. The site is famous for the exceptional preservation of organic materials, including numerous wooden artefacts related to navigation, agriculture, textile production, and basketry. This article presents interdisciplinary research on three of the most complete and well-preserved sickles recovered from the site, yet unpublished. All the components of the tools are analysed: the stone inserts, the wooden haft and the adhesive substances used to fix the stones inside the haft. Our innovative methodology combines use-wear and microtexture analysis of stone tools through confocal microscopy, taxonomical and technological analysis of wood, gas chromatography–mass spectrometry analysis of the adhesive substances, and pollen, non-pollen palynomorphs, and phytolith analysis of the remains incorporated within the adhesive. This multiproxy approach provides a significant insight into the life of these tools, from their production to their use and abandonment, providing evidence of the species of harvested plants and the conditions of the field during the harvesting.
Similar content being viewed by others


Introduction
Harvesting techniques are a fundamental element of both past and modern farming technologies. Specific tools were used for harvesting cereals even prior to cereal domestication. At the submerged site of Ohalo II, located on the southwest shore of the Sea of Galilee in Israel, during the Late Glacial Maximum (ca. 23,000 cal BP), flaked stone tools, probably hafted into a sickle handle, were used to gather barley and wheat1,2. Direct evidence of the use of a sickle for harvesting wild cereals in the Early Natufian was found at the site of Wadi Hammeh 27, in northwest Jordan, where a complete antler sickle, dated back to 14,000 cal BP, was recovered3. Fragments of sickle hafts carved into ungulate bones have also been found at the Natufian sites of Kebara Cave and El Wad (Mount Carmel, Israel), Hayonim Cave (western Galilee, Israel), and Azraq 18 (eastern Jordan)4,5,6,7. During the course of Pre-Pottery Neolithic (PPNA and PPNB), the presence of sickle-blades is one of the most characterising elements of the archaeological assemblages8,9,10, and their frequency in lithic collections increases considerably between 11,000 BP and 9000 cal BP11. Sickles underwent several changes, gradually shifting from straight to curved hafts during the Middle and Late PPNB12, as shown by the remains of a sickle found at Tell Halula in the middle Euphrates valley (Syria) (ca. 9500–9000 cal BP)13.
As from ca. 8600 cal BP onwards14 farming began to spread into the Mediterranean basin, Neolithic groups brought domesticated crops to new regions together with the set of technologies and knowledge necessary to cultivate, harvest, store, process and consume them. Harvesting tools were definitely one fundamental part of this package, an important functional category, but probably also charged with aesthetic and identity values15,16. In the Central and Western Mediterranean, the first Neolithic occupations were associated with Impresso-Cardial Ware17,18,19,20. Those early communities brought the so-called Neolithic package; consisting of plants and animals domesticated in the Near East, along with specific architectural technologies, ceramic styles, ornaments and flaked and polished tool technologies.
Unfortunately, in most of the archaeological contexts perishable materials are not preserved and little information on the morphology, size and use of sickles is therefore available. Stone inserts are often all that remain of those complex instruments21,22,23,24. Research on stone tools indicates that adaptations were made to the harvesting toolkit during the migration process. While in the Near East, sickle-blade production was characterised by an increasing technological investment in their manufacture, from the Natufian to PN, early Mediterranean sickle inserts were, in general, largely produced through a relatively simple and not particularly specialised flaking process. Throughout the migration process, harvesting technologies were adapted to different economic, technological and social conditions25. Despite that, given the paucity of archaeological evidence, information about Neolithic harvesting practises is still fragmentary, largely relying only on data from stone tools and archaeobotanical records11,25.
For this reason, an invaluable source of information is represented by those sites characterised by a good preservation of organic remains (i.e. submerged, lakeshore or lake dwelling sites). Through the study of their exceptional archaeological record, our perception of Neolithic societies can change profoundly. This is the case of a lakeshore site, excavated between 1992 and 2006 in the Lake of Bracciano, near Rome (Italy): La Marmotta26,27,28. Among the extraordinary materials recovered from the excavations (i.e. five dugout canoes and numerous objects related to navigation, basketry and all kinds of wooden crafts), 52 wooden sickles were brought to light. These finds, including both complete and fragmentary specimens, represent the largest collection of Neolithic sickles in the entire Mediterranean Basin. Until now, La Marmotta sickles had never been published except for a few drawings and photos, and their potential remained unexploited28. However, harvesting tools can inform us about several aspects of Neolithic farming. The adoption of a specific harvesting method is a response to technical, economic and cultural determinants. For example, harvesting techniques are often adapted to the type of cultivated plants and the products sought (e.g., whole plants, whole ears only, individual grains, leaves only, etc.), but other factors influence as well, for example the production scale and the size of the harvested fields. In addition, age- and gender-specific aspects can also play an important role, as for example tool’s weight and shape may be suited for specific ages or sexes29. This knowledge, which was culturally transmitted from generation to generation, enabled Neolithic groups to successfully colonise new territories30,31,32.
In this research, we address the study of the three most complete and extraordinarily well-preserved La Marmotta sickles. The three sickles display considerable typological variations between each other in the shape of the handle and the size of the cutting edge. They are therefore representative of the diversity of sickles recovered at the site.
This study has been conducted through an innovative interdisciplinary approach. All the components of the tools are analysed: the stone inserts, the wooden haft and the adhesive substances used to fix the stones inside the haft. The current study followed an integrated analytical approach combining: (1) use-wear and microtexture analysis of stone tools through confocal microscopy, (2) taxonomical and technological analysis of wood, (3) gas chromatography–mass spectrometry (GC–MS) analysis of the adhesive substances, and (4) pollen, non-pollen palynomorphs (NPP), and phytolith analysis of the remains incorporated within the adhesive. Such an integrated approach allowed us not only to reconstruct the manufacture, use and management of the tools, but also to characterise more precisely the type of cultivated cereals and the conditions of the field during the harvesting.
Materials and methods
Materials
The archaeological excavations
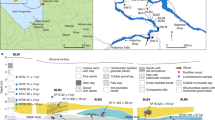
The early Neolithic site of La Marmotta is located under the waters of Lake Bracciano (Anguillara Sabazia, Rome, Italy). It has been excavated under the direction of M. A. Fugazzola Delpino (Soprintendenza Speciale, Museo Nazionale Preistorico Etnografico ‘L. Pigorini’, today Museo delle Civiltà)26,27. The archaeological deposit is located approximately 300 m from the modern shoreline, submerged at a depth of 11 m (8 m of water and 3 m of sediment), which has permitted exceptional conservation of the organic remains. The site covers an area of about 2 hectares, of which approximately 25% has been excavated (Fig. 1a). The stratigraphic sequence is composed of three contiguous layers (Layer II, Layer I, Layer ‘Chiocciolaio’), without any sterile layer or interruption between them33. Layer II corresponds to the beginning of the settlement. The pottery assemblage belongs to the Tyrrhenian facies of the Impressed Ware Culture. Layer I represents the time of the most intense occupation of the village. The pottery is here dominated by Cardial and Painted Ware. The last phase of Layer I, also called Layer ‘Chiocciolaio’, represents the abandonment of the settlement26. The radiocarbon dates from the wooden posts obtained so far are distributed over a time interval between 7840 and 7575 cal BP (post 213, square A2) and 7425–7010 cal BP (post 21, square B29)34. Similar dates have recently been obtained by dating two waterlogged poppy seed microsamples (7580–7420 and 7570–7420 cal BP)35. In the framework of the current research, six samples of charred caryopsides of emmer wheat (Triticum dicoccum) were dated (Table 1). Results confirm the data previously obtained; charred seeds from Layer II are dated between 7560 and 7330, while Layer I is dated between 7570 and 7165 cal BP. It might be speculated whether some of the dated samples (CNA-5289.1.2 and CNA-5291.1.1) were intrusive in their respective layers, but only with a larger series of dates will it be possible to define the chronology of the two main layers better.

(a) Geographical position of La Marmotta. Photograph of the subaquatic excavation. Photo from Museo delle Civiltà (MuCiv); (b) position of the sickles within the two excavated layers (Layer I and II).
The artefacts
Sickle No. 23001 was found in the year 2000, in square A179 of Level II (Fig. 1b). The sickle is complete and still conserves nine stone inserts. The handle measures 8.5 (L) × 1.8 (W) × 1.2 cm (T) and is characterised by a hinged termination. The cutting edge measures 18.5 (L) × 4.5 (W) × 1.2 (T) cm, has a curved shape and on the inner side it was carved to create a space for placing the stone inserts, which measure between 0.9 and 1 cm in width. The tool was finely finished with smooth wooden surfaces and a decorative motif along the external ridge, characterised by a series of regularly distributed grooves (Fig. 2).

Sickle No. 44297 with macro and microscopic view of the lithic inserts (44297-1 to 44297-8). Micrographs have been taken at both 100 × and 200 × for each insert.
Sickle No. 44297 was found in 2005, in square A507 of Layer II (Fig. 1b). The complete sickle conserves eight stone inserts. The handle measures 18 (L) × 2 (W) × 1.6 cm (T) and is characterised by a ‘T’ shaped termination. The cutting edge measures 18.2 (L) × 4.2 (W) × 1.2 (T) cm, and has a curved shape. An incision between 1.0 and 0.7 cm in width was carved in it. It is characterised by smooth wooden surfaces (Fig. 3).

Sickle No. 7186 with macro and microscopic view of the lithic inserts (7186-1 to 7186-3). Micrographs have been taken at both 100 × and 200 × for each insert.
The third sickle, No. 7186, was found in 1996, in square D147 of Layer I (Fig. 1b). It conserves only three complete stone inserts. A fragment of the fourth insert is still visible inside the carved groove, but it is not possible to analyse it being too fragmentary. The handle measures 9 (L) × 5.9 (W) × 1.3 cm (T) and is characterised by an arched hand guard. The cutting edge measures 16.5 (L) × 3 (W) × 1.1 (T) cm, and has a markedly curved shape, with an incision of 1.0–1.3 cm width (Fig. 4).

Sickle No. 23001 with macro and microscopic view of the lithic inserts (23001-1 to 23001-9). Micrographs have been taken at both 100 × and 200 × for each insert.
All wooden artefacts were treated at the Laboratorio di Conservazione e Restauro at the Museo delle Civiltà (Rome, Italy). After excavation, for their conservation, artefacts were cleaned with demineralized water, and then treated with polyethylene glycol (PEG 400 and PM 4000) at different proportions (10%, 20%, 40%, and 80%) during a gradual restoration process that included cold and warm phases.
To obtain precise measurements and to avoid further manipulation of the artefacts, we built 3D digital models of the three sickles (Supplementary Information S1). We acquired sub-millimetric 3D geometric/colour data using a structured light scanner (model Breuckmann Smartscan-3D duo with colour cameras and sensors for inter-changeable 90, 150, 200 and 450 mm fields of view). The 3D capture equipment, in conjunction with modelling software (Rapidform/Geomagic and Meshlab), allowed the reconstruction of high fidelity (sub-millimetric) digital models of the sickles. Such models present a high level of geometric detail and they scale to the exact real size of the artefacts. We subsequently downgraded the raw, larger models (about one million polygons and half a million colour pixels on each mesh) to different levels of detail. These derivative models reflect different research aims (e. g., in most cases, models for geometric measurement do not require colour information; etc.). Models in 3D-PDF format for their visualisation and measurement have been included in supplementary information (Supplementary Information S8–S10).
Methods
Experimental practice
In order to build an experimental interpretative framework, different harvesting experiments have been conducted. Three experimental sickles have been made; two of them with wood, Quercus sp., and one using antler. Experimental adhesive to fix stone inserts has been produced with a mix of pine resin (Pinus sylvestris), crushed pine charcoal, and beeswax. Stone inserts were made using different varieties of fine-grained tertiary flints. Three different experimental cereal fields were harvested: Triticum aestivum ‘Gentil rosso’ wheat36 in Tuscany (Italy) (15 h), Hordeum vulgare in Soria (Spain) (8 h), and Triticum monococcum in Provence-Alpes-Côte d'Azur (France) (10 h)37. Two experimental tools have been used for reaping common reeds (Phragmites australis) (2 h) and two tools to cut wild grasses (Festuca sp. and Juncus sp.) (1 h).
In addition, four other sickles were used to test low versus high height harvesting. Two sickles were used to harvest Triticum dicoccum in Asturias (Spain) (both for 6 h), and two others were used to harvest Triticum spelta also in Asturias (Spain) (one sickle used for 12 h and the other for 16 h). Therefore, the same cereal was harvested high with one sickle, near the spikelet, and low with the other, near the ground. More detailed information on the experimental practice is to be found in Supplementary Information S2.
Use-wear and microtexture analysis through confocal microscopy
Moulds of the lithic inserts fixed within the sickle handles have been made by using a two-component silicone paste (Provil Novo Light Fast by Heraeus Kulzer GmbH) that is commonly used for this goal in use-wear studies38. Prior to moulding, lithic surfaces were cleaned repeatedly with water, acetone, and alcohol and in order to remove superficial impurities, as well as to remove the polyethylene glycol used to restore the tools. Moulds were analysed at 100 × and 200 × magnifications using a Leica DM2500 M optical microscope for micro polish characterization with reflected light. Micrographs were taken with a Leica DFC420 C digital camera.
Experimental stone inserts were first cleaned with soapy water, using Derquim LM 01. Successively, inserts were immerged in a 10% H2O2 solution for 20 min’ bath in an ultrasonic tank to remove grease.
Polished areas from both archaeological and experimental inserts were scanned with the Sensofar Plu Neox blue light scanning confocal microscope, using a 20 × (0.45 NA) objective, with a spatial sampling of 0.83 μm, optical resolution of 0.31 μm, vertical resolution of 20 nm and a z-step interval of 1 μm. From 6 to 17 areas of 650 × 500 μm were scanned for each lithic insert. Afterwards, samples of 200 × 200 μm were selected from each area. Sampled areas were later processed with the SensoMAP Standard v.8 from Digital Surf in order to extract ISO 25178 texture parameters (Supplementary Information S11). Quadratic discriminant function analysis was used to treat extracted parameters, building a predictive model for group membership following a methodology previously described by Ibáñez and colleagues39. More detailed information on microtexture analysis is to be found in Supplementary Information S3.
Analysis of woody material: botanical identification
Taxonomic identification of the wood species was accomplished by observing the three anatomical planes of the wood (transversal, radial longitudinal and tangential longitudinal). The samples were obtained by extracting thin laminas from each of three planes of the wood with a cutting tool. The samples extracted must be of a size appropriate for the identification of species, but not too large as to damage the object40,41,42. The thin sections were observed with an optic microscope equipped with objectives with 4, 10, 20 and 50 × magnification (Leica DM 750 M at the Instituto Patagónico de Ciencias Sociales y Humanas: IPCSH-CONICET, Puerto Madryn, Argentina) and compared with a specialized atlas43. The treatment with polyethylene glycol has not impeded the taxonomic study a posteriori, as the microanatomy of the wood can still be observed, allowing taxonomic identification.
Organic residue extraction, GC and GC–MS analyses
Molecular analyses were conducted at the ORA Laboratory of the University of Tübingen (Germany) to investigate the organic residues which were used as hafting glue. The extraction and treatment of organic substances were carried out using the same method described elsewhere43 as well as gas chromatography (GC) and GC-Mass Spectrometry (GC–MS) analyses45. Briefly, ∼10 mg of residue was ground and solvent-extracted (Dichloromethane) for 30 min by ultrasonication to target lipid compounds. Each extract was trimethylsilylated by adding N,O-bis(trimethylsilyl)trifluoroacetamide (BSTFA, 50 μL) and a catalytic reagent (pyridin, 4 μL). Then, the trimethylsilylated samples were analysed by GC and GC–MS. More detailed information to be found in Supplementary Text S4.
Pollen and non-pollen palynomorph analysis
In total, seven samples of mastic were taken from the three sickles under investigation for palynological analyses. In order to preserve as much as possible the integrity of these unique and extraordinarily well-preserved artefacts, the amount sampled was very small, averaging 101 mg per sample (between 74 and 141 mg). The procedure to extract palynomorphs followed a protocol proven successful in several previous studies46,47. After dissolving in cold toluene for 24 h, the material was washed by centrifuging in the decreasing toluene-ethanol series and then in the increasing ethanol-deionised water series until hydration was complete. Standard acetolysis, carried out for 5 min at 100 °C48 and filtration through a 250 μm steel mesh were carried out. When a large amount of microcharcoal and mineral particles occurred, these were removed by “swirling” technique and dissolution with cold HF 40% for 24 h and hot HCl 10%49. The obtained residue was diluted to a known volume (0.5 mL) to enable calculation of absolute pollen frequency through sampling of 50–100 μL with a micropipette. Transmitted-light optical microscopy analysis was carried out at 400 × -600 × magnifications using a Leitz Diaplan, an Olympus BHS and a Vickers ML 1300 compound microscopes. Pollen and non-pollen palynomorph atlases and keys, reference collections and digital image archives were used for palynomorph determination50,51,52,53,54.
Phytolith analysis
Six samples were extracted from adhesive material for phytolith analyses, two samples for each sickle. Sickle 23001 was sampled from both an internal and external area, both near lithic 23001-6. Sickle 44297 was sampled in the mesial part, one external sample near lithic 44297-5, and one internal near lithic 44297-7. Sickle 7186 was sampled in the mesial part, both from the internal area near the broken lithic insert (Supplementary Materials S5). The methods used are similar to those described by Katz and colleagues55. A weighed aliquot of ca. 40 mg of air-dried and ashed sample (500 °C for 4 h) was treated with 50 µl of a volume solution of 6 N HCl and 450 µl 2.4 g/ml sodium polytungstate [Na6(H2W12O40)·H2O]. Microscope slides were mounted with 50 µl of material. Samples were examined at 200 × and 400 × magnification with an Olympus Bx43 optical microscope in the Archaeobotany Laboratory, Autonomous University of Barcelona. Morphological identification was based on standard literature56,57,58,59 and modern plant reference collections60,61,62,63,64. The terms used followed the International Code for Phytolith Nomenclature v. 2.065.
Results
Use-wear and microtexture analysis through confocal microscopy
Microscopic observation of the silicone impressions through reflected-light microscopy identified well-preserved use-wear traces. The appearance of the traces closely matches the experimental traces obtained from plant harvesting (Figs. 2, 3, 4). In order to quantitatively determine whether sickle inserts had been used to cut domesticated cereals or other plants, glossy areas were studied by texture analysis of 3D images obtained through confocal microscopy. Five inserts were excluded from the analysis, two because they were made on coarse-grained flints (23001-2, and 23001-8), which could affect the classification, and three because they almost showed no use-wear traces (44297-1, 44297-2, and 44297-4). The results obtained for the remaining tools show that 14 of the 15 analysed inserts were classified as having harvested domesticated cereals (Supplementary Information S3), while only one insert was classified as having cut reeds. This latter insert (23001-9) shows very lightly developed wear and was probably added to the sickle in a later stage. As observed experimentally, the inserts placed at the distal end of the sickle are the ones most easily lost during harvesting as they are more exposed and, therefore, more likely to become detached after accidental impact with the ground, stones, etc.
The different degree of trace development reveals the existence of specific management strategies. While different degrees of trace developments can occur in the same sickle, depending on the position of each insert, in the case of sickle No. 44297 it is clear that the inserts were placed in the shaft at different times. The first two inserts (44297-1, 44297-2) were almost unused, while the third one shows a very well-developed gloss (44297-3). This suggests that the first two inserts had been replaced soon before the abandonment of the sickle, while the other inserts had been used for a considerably longer period. Similarly, the lithic 44297-4 is a residual insert from previous uses, and did not possess a functional purpose in the actual configuration of the sickle. Therefore, inserts were most likely not changed all at once, but at different times, possibly due to accidental loss of inserts during harvesting.
As can be seen from the micrographs, use-wear traces on the archaeological specimens display rather smooth polish, with a slightly pitted appearance and thin striations. This is clear especially in the case of inserts with well-developed gloss (23001-1, 23001-5, 7186-2, 7186-3, and 44297-3). The presence of striations and abrasions is usually associated with low height harvesting, because of the occasional contact of the lithic inserts with ground particles66. This has been confirmed by the quantitative analysis of experimental and archaeological use-wear traces, as 93.3% of the analysed sub-areas on the La Marmotta sickles are classed together with experimental tools used for low height harvesting, at the base of the stem39 (Supplementary Information S3).
Analysis of woody material: botanical identification
The taxonomical analysis of the three sickles evidenced the use of two woody species as raw material (Fig. 5). Two of them were made with wood from deciduous oak (Quercus sp.) (No. 44297 and No. 23001) and one with wood from Rosaceae/Maloideae (No. 7186). It should be noted that the taxonomic analysis of other types of artefacts at La Marmotta, also has equally shown the use of deciduous Quercus sp. in their fabrication (publication in prep.). However, the use of Rosaceae/Maloideae had not been identified at the site for other types of artefacts, connected with sailing or hunting, for instance. Despite that, the use of deciduous oak wood is well-documented at the site overall67,68. Although Rosaceae/Maloideae has not been identified as a raw material for other types of artefacts, this taxon had been documented at La Marmotta. According to Fugazzola’s studies, the inhabitants of the village knew several fruit tree species, including crab apples26,67.

(A) Sample from the transversal section of deciduous Quercus sp.; (B) sample from the tangential section of deciduous Quercus sp.; (C) sample from the radial section of deciduous Quercus sp.; (D) sample from the transversal section of Rosaceae/Maloideae; (E) sample from the tangential section of Rosaceae/Maloideae; (F) sample from the radial section of Rosaceae/Maloideae.
GC–MS analysis
The identification of characteristic diterpenes markers from the abietane and pimarane families in the adhesive samples indicates the use of Pinaceae resin(s)69,70,71 to haft the three sickles from La Marmotta. Dehydroabietic, abietic, isopimaric and sandaracopimaric acids were detected in all of them (Supplementary Information S4). In addition, the presence of Seco- dehydroabietic (α and β) and pimaric acid as main pimarate suggest a pine origin69 for the resin on sickle 23001.
Pollen and non-pollen palynomorph analysis
Mastic was sampled both from the outer surface, in the gaps between lithics, and at the depth of a few millimetres, inside the hafting grooves, in order to assess potential qualitative and quantitative differences in the palynological content at different depths. The pollen absolute frequency was variable (between 6501 and 133,929 grains/g), and usually higher in the inner samples compared with the outer ones. Each sample, mostly containing a large amount of well-preserved pollen, has allowed counting, on average, more than 780 grains (from 271 to 1481). Figure 6 show percentage values of individual arboreal and shrubby taxa (AP), herbaceous taxa (NAP) and NPPs, as well as cumulative data for plant categories, which are relevant in terms of ecology and anthropogenic impact.

(a) Pollen percentage diagram of Arboreal Pollen (AP), Non-Arboreal Pollen (NAP) taxa, and non-pollen palynomorphs (NPPs) in the adhesive on three sickles from La Marmotta. NPP values are expressed as percentages of total pollen; (b) pollen percentage diagram of selected categories of ecological and anthropogenic interest in the adhesive on three sickles from La Marmotta.
The first two sickles (No. 7186 and 44297) suggest a very similar palynological scenario. Among the arboreal taxa (AP between 29.5 and 39.1%), oak (Quercus t. caducifolia) (6.4–23.5%) prevails, with a sporadic occurrence of other elements in the mesophilous deciduous oakwood, including Carpinus betulus, Ulmus, Ostrya/Carpinus orientalis, Corylus, Fraxinus ornus-type and Vitis. Typically, Mediterranean evergreen species, such as Quercus ilex-type, Olea-type and Pistacia, play only a minor role in this assemblage.
Among the hydrophilous arboreal species Alnus glutinosa-type deserves a mention, followed by Salix and Populus. More widely documented are grains of a number of taxa probably common in the riparian and aquatic habitats of the site, such as Parnassia, Alisma plantago aquatica-type, Typha, Callitriche, Potamogeton, Sparganium and Nymphaea alba-type. Potamogeton, Sparganium and Nymphaea have been documented also from the analysis of macroremains (Supplementary Information S7).
Wild grasses occur in particularly high percentages, which reach identical values in the adhesive from the two sickles, in both cases with higher values in the superficial levels of the mastic (between 22.0 and 23.1%) than in deep samples (between 11.4 and 11.6%). Analogous considerations apply to the herbaceous anthropogenic taxa typical of cultivated and ruderal areas (Cirsium-type, Cichorieae minor-type, Plantago lanceolata-type, Urticaceae, Galium, Papaver rhoeas gr., Chenopodiaceae, Rumex-type. and Polygonum aviculare-type)72,73.
In order to separate different cereals, the biometric data of each individual pollen grain have been evaluated, measuring the average diameter and the pore annulus. The examination, in spite of some limitations that cannot always avoid equivocal determinations51,52,74,75,76,77, have allowed identification of both Triticum-type (d > 50 μm and annulus > 10 μm) and Hordeum-type (d = 40–50 μm and annulus 8–10 μm), occurring with values of 5.5–10.6% and 4.4–8.6%, respectively. Generally, percentages of Cerealia are highly relevant in all samples (between 15.4 and 26.0%). Both cereals often occurred also as pollen aggregates.
The results obtained from the three mastic samples of sickle No. 23001, one taken from the surface and two from within the adhesive, appear entirely different and atypical compared with the two previous artefacts. Arboreal taxa occur in minimal amounts (AP between 2.7 and 4.7%), among which Alnus glutinosa-type prevails (values between 1.6 and 2.4%), along with equally low percentages of Cerealia (between 0.2 and 0.9%) and anthropogenic indicators (averaging 0.8%), whereas grains of swampy herbaceous plants occur in constantly high percentages, notably Oenanthe-type (averaging 78.1%), and to a lesser extent Alisma plantago-aquatica-type (averaging 13.8%).
Based on preliminary observations made exclusively using optical microscopy and concerning the main morphological and biometric characteristics of 50 grains of Oenanthe-type [P = 33.8-(39.4)-48.4 μm; E = 14.2-(19.4)-22.4 μm; P/E = 1.7-(2.0)-2.5 μm; PAI = 0.22-(0.30)-0.39 μm], and according to the data reported in lietrature78, their attribution to Oenanthe aquatica group and to Oenanthe peucedanifolia group should be considered valid.
Most of samples were devoid of NPPs, with the exception of No. 23001 and 7186 (the samples from the external surfaces). No. 23001 included algal remains (Pediastrum), whereas No. 7186 contained five fungal taxa (Alternaria, Apiosordaria-type, Chaetomium-type, Bactrodesmium, Diporotheca webbiae), the first of which is by far dominant. Due to their size (c. 50–55 μm), the typically club-shaped spores of Alternaria can be identified as conidia of A. tenuissima79, while the morphology of Bactrodesmium spores points to conidia of B. obovatum or B. abruptum80.
Phytolith analysis
Phytoliths were recorded in all the adhesive samples in different amounts (ranging from 84,000 to 470,000 phytoliths per 1 g of material Supplementary Table S5). In addition, other siliceous microfossils such as diatoms and sponge spicules were also noted in the phytolith slides (Fig. 7). Their presence in association with phytolith abundance points towards general good preservation conditions of silica assemblages.

Photomicrographs of phytoliths and other siliceous microfossils identified in the samples (200 × or 400 ×). (a) Sponge spicule; (b) articulated spheroid phytoliths from dicotyledonous plants; (c) multicelled elongate phytoliths from grasses.
Most of the phytoliths from inner samples derived from dicotyledonous plants, mainly from wood/bark with an average of about 60% (Supplementary Table S5). The dominant morphotypes were spheroids, displayed either singly and in articulated groups (Fig. 7). Spheroid phytoliths occur in a range of plants including woody dicotyledonous (in the sense of non-monocot angiosperms) and some herbaceous monocots64, but in particular, ornate surface textures (rugose or scabrate) are produced in variable amounts by woody taxa from Mediterranean areas, such as (Pinus sp.) and oak (Quercus sp.)59,64. The phytolith records are consistent with biomarker signatures pointing to a pine origin of the adhesive, and therefore also with the sickle wood raw material (deciduous Quercus sp. in samples 44,297 and 23,001).
Of particular note is the presence in all the samples of phytoliths from the culms and leaves of Pooideae grasses (Fig. 7c), which are common in wetlands and include cereals such as wheat and barley reported in the site’s macrobotanical records, including hulled barley (Hordeum distichum), emmer (Triticum dicoccum), einkorn (Triticum monococcum) and free-threshing wheat (Triticum turgidum or Triticum durum)81.
Discussion
This study presents new data for three of the most complete sickles in the Early Mediterranean Neolithic, dated between ca. 7560 and 7165 cal BP. Combining use-wear, archaeobotanical, and residue analysis it has been possible to gain insight into the earliest evidence for harvesting practises in the Central Mediterranean Neolithic, but also into the environment around the site and the cereals cultivated.
The three sickles are rather small, between ca. 36 and 24 cm of maximum length, with a cutting edge between 18 and 16 cm long. This size is similar to the so-called Karanovo antler sickles from Bulgaria, dated between ca. 8200–7500 cal BP, whose cutting edge is between 24 and 14 cm long82. Identical specimens have been recovered at the sites of Kovačevo83, also in Bulgaria, and Hacılar VI84 in Turkey. In comparison with historical and ethnographic iron sickles, it seems that Neolithic harvesting tools were smaller and adapted to cutting a small number of plants with each stroke85,86. The length of the blade is, indeed, directly related to the quantity of plants to be harvested29. For example, traditional sickles employed in Eurasia during the eighteenth–nineteenth centuries generally had a blade of about 30–40 cm or more87.
The ligneous raw material used for their manufacture (deciduous Quercus sp. and Rosaceae/Maloideae) were not exclusively employed for sickles. Both species were widely used at La Marmotta for a diversity of wood crafts. On one hand, the frequent use of deciduous Quercus sp. wood at the site may be related to the frequency of the taxon in the local vegetation during the Neolithic. Pollen records from the nearby Lago di Vico and Lagaccione (Lago di Bolsena), further to the north, show that abundance of deciduous Quercus sp. was a key feature of the mid-Holocene vegetation88. On the other hand, the wood of Maloideae would have been relatively easy to acquire during forest clearings. The wood of the different species of the Maloideae is hard and characterised by close, compact grain that makes it ideal for carving and engraving. At other prehistoric sites, the wood of Maloideae has been used for artefacts such as axe hafts, spear shafts and tool handles89,90,91, while at La Draga site, in north-eastern Iberia, the use of Rosaceae/Maloideae as a raw material has only been documented in the remains of charcoal associated with the use of firewood92.
All sickles show a very specific design and particular attention was paid to their shaping and finishing. As experimentally tested, the arched hand guard of sickle No. 7186 enables a tighter control over the tool during harvesting. Still puzzling is the ‘T’ shape termination of sickle No. 44297 and more detailed experimental and use-wear studies are necessary. In addition, it would also be important to determine whether the heterogeneity in the use of ligneous raw materials reflects differences in the type of haft (i.e., hinged handles, handles with hand guard, etc.). Ongoing archaeobotanical studies, based on techno-morphometric analyses, will be able to assess whether the taxonomic selection was connected to the morphology of the different sickles.
Stone inserts are mainly made on blade fragments, as visible from Figs. 2, 3 and 4. Their typological and morphometric characteristics are like the rest of the sickle inserts found in the flaked stone assemblage of La Marmotta (Supplementary Information S6). Truncations are dominant, average sizes are 29–20 mm in length, 13–10 mm in width, 3–2 mm in thickness. Based on a macroscopic observation, a provenance from Gargano promontory can be suggested for some of the stone inserts included in the wooden sickles (e.g. 44297-5). However, regional varieties of Apenninian chert types are the most used raw-materials93,94. The flaked stone assemblage recovered at La Marmotta is made on these same raw materials, indicating that the site was integrated in regional and supraregional networks of chert circulation. Obsidian provenance95 data confirm that the site was embedded in a far-reaching network of raw-material distribution.
The use of Pinaceae resin for hafting is confirmed both by GC–MS analysis and by the two microfossil records (pollen and phytoliths). Pinaceae resin is not common in archaeological contexts, and it probably represents the first evidence of its use as adhesive substance during the Neolithic. The only other known case of the use of Pinaceae resin for hafting is from two Palaeolithic cave sites in South Italy96. Mediterranean pine was available in the site surroundings, as well as documented by pollen analysis from neighbouring lakes88, pointing to a local or regional procurement. This local supply strategy for adhesive materials seems to confirm practices previously documented in Central Italy in the eighth millennium cal BP. Indeed, local bitumen was used for the same function in the region between the Adriatic Sea and the Apennine Mountains97,98. Nevertheless, and although rarely documented, the use of exogenous materials to produce adhesives has recently been identified for the Mediterranean Neolithic. This is the case of birch bark, whose use is documented over a large area from Catalonia to the Apennines43,98,99.
The analysis of Neolithic mastic from the site of La Marmotta has shown the capability of this material to trap a remarkable amount of pollen grains, which appeared to be in an excellent state of preservation46. Therefore, this material has the potential to provide new research insights and data on the vegetation scenario around a site, the cultivated taxa and the local synanthropic flora.
The results obtained from the first samples (No. 7186 and 44297), rich in individual or grouped Triticum-type and Hordeum-type grains, further confirm the reaping function of these tools. Indeed, it is widely known that cereal pollen has a tendency to stick to the caryopses, in particular underneath the glumes of hulled wheat until spike maturation occurs, and that these grains can be found in high numbers within the harvest and even in the by-products of cereal grinding75,100,101,102,103. In contrast, the exceptionally high concentration of Oenanthe-type pollen in the third sickle (No. 23001) is unusual and unexpected. The genus Oenanthe includes hydro-hygrophilous plants in wet meadows, swamps and lakeshores, and it is still present in the Tyrrhenian area of central Italy. Due to the content of oenanthotoxins in the fruits and roots, several species of Oenanthe genus possess pharmacological properties. These substances have active principles similar to the cicutoxin of Cicuta virosa, with a neuro-enterotoxic effect. Some species, when consumed fresh and even in small quantities, seem to be able to induce a mental state similar to drunkenness, but can also cause nausea, vomiting and convulsions; but they are harmless when consumed dry104,105,106. Oenanthe plants have long been used as medicinal plants in Asia107. In an eighteenth century herbarium, however, it is named as Cicuta palustre. The caption near the specimen explains that, due to its toxicity, it is “…stirps ipsa venenata infamis” (known for its poisonous properties)108.
This anomalous find lacks an easy interpretation at the moment and can be added to the list of ‘unusual’ taxa identified at La Marmotta, so far known through the macrofossil record, such as Papaver somniferum, Carthamus lanatus and Silybum marianum, which are species with officinal and medical properties. In this sense, in agreement with Rottoli109,110, it cannot be ruled out that the first Neolithic communities settled on the shores of Lake Bracciano managed floral resources in various ways, and not only for strictly nutritional purposes. Aside from cereal crops (Hordeum disticum, Triticum dicoccum, Triticum monococcum, Triticum durum/turgidum) and legumes (Pisum sativum, Lens culinaris, Lathyrus cicera/sativus, Vicia cf. sativa, Vicia cf. ervilia, Vicia cf. faba) documented in the macrobotanical records, a number of officinal plants may have been harvested around the lakeshore village of La Marmotta for medicinal, tanning and textile purposes, and also for various craft works, the exact ethnobotanical significance of which it is difficult to unveil. The potential psychoactive effects linked to the “drunkenness” effect provoked by the active principles of Oenanthe may have been part of these medical or ritual uses.
Pollen results for sickles No. 7186 and 44297 are in agreement with data provided by the quantitative analysis use-wear traces. Use-wear traces are grouped with experiments harvesting domesticated cereals (Hordeum vulgare, Triticum monococcum and Triticum aestivum). Nevertheless, the high percentage of Oenanthe-type pollen on sickle No. 23001 is not fully reflected by the use-wear traces on the inserts of this same tool, which is grouped with the experimental harvesting of domesticated cereals. This discordancy between pollen and use-wear may be a consequence of two different factors:
-
(a)
it is possible that the accumulation of Oenanthe pollen is the result of a last, single harvesting, while the use-wear pattern represents the accumulation of several previous, repeated, harvesting episodes. The presence of a minor component of Cerealia-type pollen in the adhesive from No. 23001 (Fig. 6) and of phytolith morphotypes derived from the leaves and stems of grasses of the Pooids (Supplementary Information S5) are consistent with the hypothesis that sickle No. 23001 was used for harvesting cereals at some point. In addition, it is worth remembering that insert 23001-9, likely the last one to be added to the sickle given the low degree of use-wear development, is classed together with experimental tools used for cutting reeds, plants growing in moist, swampy areas, as also many plants of the Oenanthe genus. Therefore, the use-wear traces on 23001-9 may represent the outcome of a last harvesting episode in which the sickle was used for cutting non-cereal plants, such as plants of the Oenanthe genus, but other also plants in moist, swampy areas. Among the macrobotanical record, for example, fragments of the rhizome of Phragmite australis are abundant (Supplementary Information S7). It is also remarkable that sickle No. 23001 is the only one among all the La Marmotta wooden sickles with a decoration on its back, and a special function for this tool may be imagined.
-
(b)
another possibility is that the presence of pollen from aquatic plants is due to the depositional conditions from which tools have been recovered. The presence of moist habitats is well indicated by the abundance of aquatic pollen grains and is paralleled by the occurrence of Pediastrum, a green alga colonising eutrophic water pools111, of Diporotheca webbiae, a fungus parasitic on marsh ferns112, and of Bactrodesmium, a fungal genus thriving especially on submerged wood. In addition, the presence of diatoms and sponges also confirms that siliceous remains occurring in freshwater habitats were embedded within the resin.
Plant micro remains (phytoliths, pollen, NPPs) trapped into the adhesive also contribute to a reconstruction of the environment near the harvested fields. The palynological record indicates the presence of a Mediterranean deciduous forest in the surroundings, but also with species typical of aquatic habitats. The presence among NPPs of Bactrodesmium obovatum and B. abruptum, both saprophytic on arboreal taxa such as Quercus, Fraxinus, Acer, Fagus, Alnus, Betula, Populus79, also points to forested areas in the immediate vicinity of the field, as fungal spores have a limited spatial dispersal compared to pollen grains. Interesting is the presence of spores of Apiosordaria-type and Chaetomium-type, which although occurring in very low amounts, are often associated with herbivore dung and herding activity113. The faunal record from La Marmotta is based on domesticates, mainly goat and sheep, along with cattle, pigs and dogs, as well as wild animals including red deer, red fox, birds, reptiles and fish26,27,28.
Wild grasses are incorporated in the adhesive as well, particularly in sickles No. 7186 and No. 44297; in both cases with higher values in the superficial levels of the mastic (between 22.0 and 23.1%) than in deep samples (between 11.4 and 11.6%). Analogous considerations apply to the herbaceous anthropogenic taxa typical of cultivated and ruderal areas (Cirsium-type, Cichorieae minor-type, Plantago lanceolata-type, Urticaceae, Galium, Papaver rhoeas gr., Chenopodiaceae, Rumex-type and Polygonum aviculare-type)72,73. These data suggest that the harvested cereal fields were characterized by a greater diversity of weeds than inferred from the on-site macrobotanical record at La Marmotta (Supplementary Information S7). Harvested crops probably underwent several cleaning stages before being stored at the site.
The low height harvesting documented through the qualitative and quantitative study of use-wear traces confirms that the entire plant was cut and exploited, not only the ears. Cereal straw was probably part of the materials used for building housing structures and different crafts at La Marmotta. Nevertheless, despite the abundant and exceptionally preserved archaeobotanical record at the site, a full and detailed study of basketry, textiles, ropes and other aggregates of vegetal fibres is still to be addressed. Preliminary analysis of the charred seed assemblage indicates that the percentage of culms within the carpological assemblage is very low. However, as mentioned above, this might be due to crop cleaning activities carried out outside the village.
Through the analysis of microremains trapped in the adhesive it has been possible to obtain a better picture of the floral scenario around the dwelling area and the crop fields, to date investigated through more common approaches (e.g., palynological and macrobotanical records from archaeological sediments). Nevertheless, some caution is necessary when interpreting results, as adhesives have undergone a series of manipulations over time: from their hot preparation, obtained mixing several ingredients of different origin (pine resin, charcoal powder, etc.), to their use for hafting lithics inside the groove cut into the sickle and their exposure to the external environment during agricultural activities, and after that, to depositional and post-depositional agents. Moreover, these tools may have had a long lifetime, perhaps extending over a few decades, and they may have been periodically fixed, due to inevitable wearing-out of lithic blades and consequent replacement, to maintain efficiency of use. Evidence for this has also been provided by experimental studies (Supplementary Information S2), which have shown that these sickles need frequent mastic replacement during the use, in order to keep the blades in place and prevent them from detaching and moving. This action can easily transport organic material towards the inner layers of the mastic, which is eventually trapped in the longitudinal groove of the tool. Therefore, also based on experimental research, it is reasonable to hypothesize that most of the residues observed in the adhesive were incorporated during harvesting activities. The sickles trapped inside and on their surfaces a wide range of microparticles of botanical origin, including pollen grains, fungal spores, microalgae, fragment of leaf epidermis and other tissues, as well as microorganisms and dust particles.
Conclusions
The wooden sickles from the submerged site of La Marmotta are some of the most impressive archaeological finds associated with the expansion of the Neolithic into the Mediterranean. This contribution provides significant insight into the life of these tools, from their production to their use.
The multiproxy approach carried out integrates, for the first-time, use-wear and microtexture analysis of stone tools, with taxonomical analysis of wood, GC–MS analysis of the adhesive substances, along with direct microfossil evidence from pollen, NPP and phytoliths. It has considerably improved our knowledge of Early Neolithic harvesting and farming systems. Sickles are emblematic tools whose manufacture and use are embedded in a larger economic system including the procurement and crafting of exogenous and local raw materials, plant food processing and production, as well as possibly medicinal and psychoactive behaviours.
Data availability
All data generated or analysed during this study are included in this article as supplementary information files.
References
Snir, A. et al. The origin of cultivation and proto-weeds, long before Neolithic farming. PLoS ONE 10(7), e0131422. https://doi.org/10.1371/journal.pone.0131422 (2015).
Groman-Yaroslavski, I., Weiss, E. & Nadel, D. Composite sickles and cereal harvesting methods at 23,000-years-old Ohalo II Israel. PLoS ONE 11(11), e0167151. https://doi.org/10.1371/journal.pone.0167151 (2016).
Edwards, P. C. A 14000 year-old hunter-gatherer’s toolkit. Antiquity 81(314), 865–876. https://doi.org/10.1017/S0003598X0009596X (2007).
Le Dosseur, G. Bone Objects in the Southern Levant from the Thirteenth to the Fourth Millennia. Bulletin du Centre de recherche français à Jérusalem 12, 111–125 (2003).
Garrard, A., & Yazbeck, C. The Natufian of Moghr el-Ahwal in the Qadisha valley, northern Lebanon. in Natufian Foragers in the Levant. International Monographs in Prehistory (eds. Bar-Yosef, O. & Valla, F. R.). 17–47. (Michigan, Ann Arbor, 2013).
Belfer-Cohen, A. The Natufian in the Levant. Annu. Rev. Anthropol. 20, 167–186. https://doi.org/10.1146/annurev.an.20.100191.001123 (1991).
Stordeur, D. Le Natoufien et son évolution à travers les artefacts en os in Natufian Foragers in the Levant. International Monographs in Prehistory (eds. Bar-Yosef, O. & Valla, F. R.). 457–482. (Michigan, Ann Arbor, 2013).
Rosen, S. A. Lithics after the Stone Age: a handbook of stone tools from the Levant. (Rowman Altamira, 1997).
Anderson, P. C. Prehistory of agriculture: new experimental and ethnographic approaches. (Cotsen Institute of Archaeology Press, 1999).
Ibáñez, J. J., González Urquijo, J. E., & Rodríguez, A. The evolution of technology during the PPN in the Middle euphrates. A view from use wear analysis of lithic tools. in Systèmes techniques et communautés du Néolithique Préceramique au Proche Orient. Technical Systems and Near Eastern PPN Communities (eds. Astruc, L., Binder, D. & Briois, F.) 153–165 (Editions APDCA, 2007).
Maeda, O., Lucas, L., Silva, F., Tanno, K. I. & Fuller, D. Q. Narrowing the harvest: Increasing sickle investment and the rise of domesticated cereal agriculture in the Fertile Crescent. Quatern. Sci. Rev. 145, 226–237. https://doi.org/10.1016/j.quascirev.2016.05.032 (2016).
Pichon, F. Exploitation of the cereals during the Pre-pottery Neolithic of Dja’de-el-Mughara: Preliminary results of the functional study of the glossy blades. Quatern. Int. 427, 138–151. https://doi.org/10.1016/j.quaint.2016.01.064 (2017).
Borrell, F., & Molist, M. Projectile Points, Sickle Blades and Glossed Points. Tools and Hafting Systems at Tell Halula (Syria) during the 8th millennium cal. BC Paléorient, 33(2), 59–77 (2007). https://doi.org/10.2307/41496812.
Douka, K., Efstratiou, N., Hald, M., Henriksen, P. & Karetsou, A. Dating Knossos and the arrival of the earliest Neolithic in the southern Aegean. Antiquity 91(356), 304–321. https://doi.org/10.15184/aqy.2017.29 (2017).
Perlès, C. From the Near East to Greece: Let's reverse the focus. Cultural elements that didn't transfer. in How did farming reach Europe? (ed. Lichter, C.) 275–290 (Istanbul, Ege Yayınları, 2005).
Gijn A.L. van & Wentink K. The role of flint in mediating identities: The microscopic evidence. in Mobilty, meaning & transformations of things, shifting contexts of material culture through time and space. (eds. Hahn, H.P. & Weiss, H.) 120–132 (Oxford, Oxbow Books, 2013).
Guilaine, J. The neolithic transition: From the Eastern to the Western Mediterranean. in Times of Neolithic Transition along the Western Mediterranenn. (eds. O., García-Puchol & D. C., Salazar-García) 15–31 (New York, Springer, 2017). https://doi.org/10.1007/978-3-319-52939-4_2.
Forenbaher, S. & Miracle, P. T. The spread of farming in the Eastern Adriatic. Antiquity 79(305), 514–528 (2005).
Gabriele, M. et al. Long-distance mobility in the North-Western Mediterranean during the Neolithic transition using high resolution pottery sourcing. J. Archaeol. Sci. Rep. 28, 102050. https://doi.org/10.1016/j.jasrep.2019.102050 (2019).
Manen, C., Perrin, T., Guilaine, J., Bouby, L., Bréhard, S., Briois, F., Durand, F., Marinval, P. & Vigne, J. D. The Neolithic transition in the western Mediterranean: A complex and non-linear diffusion process—the radiocarbon record revisited. Radiocarbon 61(2), 531–571 (2019). https://doi.org/10.1017/RDC.2018.98
Ibáñez, J. J., Clemente Conte, I., Gassin, B., Gibaja, J. F., Gonzáles Urquijo, J. E., Márquez, B., Philibert, S., Rodríguez Rodríguez, A. Harvesting technology during the Neolithic in south-west Europe. in Prehistoric technology 40 years later: functional studies and the Russian legacy (eds. Longo L. & Skakun, N.) 183–95 (Oxford, Archaeopress, 2008).
Gibaja, J. F., Ibáñez, J. J., González Urquijo, J. E. Neolithic Sickles in the Iberian Peninsula. in Exploring and Explaining Diversity in Agricultural Technology, EARTH 2 (eds. van Gijn, A., Whittaker, P. & Anderson, P.) 112–118 (Oxford, Oxbow Books, 2014).
Mazzucco, N., Capuzzo, G., Petrinelli-Pannocchia, C., Ibáñez, J. J., Gibaja, J. F. Harvesting tools and the spread of the Neolithic into the Central-Western Mediterranean area. Quat. Int. 470(Part B), 511–528 (2018). https://doi.org/10.1016/j.quaint.2017.04.018.
Mazzucco, N., Guilbeau, D., Kačar, S., Podrug, E., Forenbaher, S., Radić, D., Moore, A. T. M. The time is ripe for a change. The evolution of harvesting technologies in Central Dalmatia during the Neolithic period (6th millennium cal BC). J. Anthropol. Archaeol. 51, 88–103 (2018). https://doi.org/10.1016/j.jaa.2018.06.003
Mazzucco, N. et al. Migration, adaptation, innovation: The spread of Neolithic harvesting technologies in the Mediterranean. PLoS ONE 15(4), e0232455. https://doi.org/10.1371/journal.pone.0232455 (2020).
Fugazzola Delpino, M. A., D’Eugenio, G. & Pessina, A. “La Marmotta” (Anguillara Sabazia, RM): Scavi 1989—un abitato perilacustre di età Neolitica. Bull. Paletnol. Ital. 84, 181–315 (1993).
Fugazzola Delpino, M. A., Pessina, A. Le village néolithique submergé de La Marmotta (lac de Bracciano, Rome). in Le Néolithique du Nord-Ouest méditerranéen (ed. Vaquer, J.) 35–38 (Société préhistorique française, Paris, 1999)
Fugazzola Delpino, M. A. La Marmotta. in Le ceramiche impresse nel Neolitico antico. Italia e Mediterraneo (eds. Fugazzola, M.A., Pessina, A. & Tiné, V) 373–395 (Istituto Poligrafico e Zecca dello Stato, Roma, 2002).
Grantham, G. L. faucille et la faux. Études rurales 151–152, 103–131 (1999).
Sigaut, F. Identification des techniques de récolte des graines alimentaires. J. Agric. Trad. Bot. Appl. 25(3), 145–161 (1978).
Anderson, P. C., Sigaut, F. Introduction: reasons for variability in harvesting techniques and tools. in Exploring and Explaining Diversity in Agricultural Technology, EARTH 2 (eds. van Gijn, A., Whittaker, P. & Anderson, P.) 85–93 (Oxford, Oxbow Books, 2014).
Halstead, P. Two oxen ahead: Pre-mechanized farming in the Mediterranean (John Wiley & Sons, 2014).
Fugazzola Delpino, M. A. & Mineo, M. La piroga neolitica di Bracciano (La Marmotta 1). Bull. Paletnol. Ital. 86, 197–266 (1995).
Fugazzola Delpino, M. A., Tinazzi, O. Dati di cronologia da un villaggio del Neolitico Antico. Le indagini dendrocronologiche condotte sui legni de La Marmotta (lago di Bracciano-Roma). in Miscellanea in ricordo di Francesco Nicosia, 1–10 (Studia Erudita, Fabrizio Serra Editore, 2010).
Salavert, A. et al. Direct dating reveals the early history of opium poppy in western Europe. Sci. Rep. 10, 20263. https://doi.org/10.1038/s41598-020-76924-3 (2020).
Ghiselli, L. et al. Nutritional characteristics of ancient Tuscan varieties of Triticum aestivum L. Ital. J. Agron. 11(4), 237–245 (2016).
Pichon, F. Une moisson expérimentale de céréales, Séranon (août 2016), ArchéOrient - Le Blog, 14 octobre2016, (2016). https://archeorient.hypotheses.org/6667.
Banks, W. E. & Kay, M. High-resolution casts for lithic use-wear analysis. Lithic Technol. 28(1), 27–34. https://doi.org/10.1080/01977261.2003.11721000 (2003).
Ibáñez, J. J., Anderson, P. C., Gonzalez-Urquijo, J. & Gibaja, J. Cereal cultivation and domestication as shown by microtexture analysis of sickle gloss through confocal microscopy. J. Archaeol. Sci. 73, 62–81. https://doi.org/10.1016/j.jas.2016.07.011 (2016).
Caruso Fermé, L. Modalidades de adquisición y uso del material leñoso entre grupos cazadores-recolectores patagónicos (Argentina). Métodos y técnicas de estudios del material leñoso arqueológico. PhD Dissertation (Universidad Autónoma de Barcelona, Barcelona, 2012).
Caruso Fermé, L., Clemente, I., Civalero, M.T. A use-wear analysis of wood technology of patagonian hunter-gatherers. The case of Cerro Casa de Piedra 7, Argentina. J. Archaeol. Sci. 15, 315–321 (2015). https://doi.org/10.1016/j.jas.2015.03.015.
Caruso Fermé, L., Aschero, C. Manufacturing and use of the wooden artifacts. A use-wear analysis of wood technology in hunter-gatherer groups (Cerro Casa de Piedra 7 site, Argentina). J. Archaeol. Sci. 31, 102291 (2020). https://doi.org/10.1016/j.quaint.2020.10.067.
Schweingruber, F. H. Anatomy of European wood: An atlas for the identification of European trees, shrubs and dwarf shrubs (Paul Haupt, 1990).
Rageot, M. et al. Birch bark tar production: Experimental and biomolecular approaches to the study of a common and widely used prehistoric adhesive. J. Archaeol. Method Theory 26, 276–312. https://doi.org/10.1007/s10816-018-9372-4 (2019).
Rageot, M. et al. New insights into Early Celtic consumption practices: Organic residue analyses of local and imported pottery from Vix-Mont Lassois. PLoS ONE 14(6), e0218001. https://doi.org/10.1371/journal.pone.0218001 (2019).
Arobba, D., Caramiello, R., Martino, G. P. Analisi paleobotaniche di resine dal relitto navale romano del Golfo Dianese. Rivista di Studi Liguri, LXIII-LXIV: 339–355 (1999).
Marshall, D. M. Archaeological pollen: extraction from ancient resins. The American Association of Stratigraphic Palynologists. Prog. and Abstr., 38th Ann. Mtg., 34 (2005).
Berglund, B. E., Ralska-Jasiewiczowa, M. Pollen analysis and pollen diagrams. in Handbook of Holocene Palaeoecology and Palaeohydrology. (eds. Berglund, B. E.) 455–484 (Chichester, Wiley, 1986).
Traverse, A. Paleopalynology. Second Edition, 813 p. (Dordrecht, Springer, 2007).
Punt W. (ed.) The Northwest European pollen flora (NEPF), vol. 2 (1980), vol. 3 (1981), vol. 4 (1984) vol. 5 (1988), vol. 6 (1991), vol. 7 (1996), vol. 8 (2003) (Elsevier, Wim Punt, Amsterdam, 1980–2003)
Fægri, K. & Iversen, J. Textbook of pollen analysis (John Wiley and Sons, 1989).
Moore, P. D., Webb, J. A. & Collinson, M. E. Pollen analysis 2nd edn. (Blackwell, 1991).
Beug, H.-J. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete (Pfeil, 2004).
Reille, M. Pollen et spores d’Europe et d’Afrique du Nord. (Marseille, Laboratoire de Botanique Historique et Palynologie, 1992).
Katz, O. et al. Rapid phytolith extraction for analysis of phytolith concentrations and assemblages during an excavation: An application at Tell es-Safi/Gath Israel. J. Archaeol. Sci. 37(7), 1557–1563. https://doi.org/10.1016/j.jas.2010.01.016 (2010).
Brown, D. A. Prospects and limits of a phytolith key for grasses in the central United States. J. Archaeol. Sci. 11, 345–368. https://doi.org/10.1016/0305-4403(84)90016-5 (1984).
Rosen, A. M. Preliminary identification of silica skeletons from Near Eastern archaeological sites: an anatomical approach. in Phytolith Systematics: Emerging Issues, Advances in Archaeological and Museum Science (eds. Rapp, G. Jr. & Mulholland, S. C.) 129–148 (New York, Plenum Press, 1992)
Mulholland, S. C., Rapp Jr. G. A morphological classification of grass silica-bodies. in Phytolith Systematics: Emerging Issues, Advances in Archaeological and Museum Science (eds. Rapp, G. Jr. & Mulholland, S. C.) 65–89 (New York, Plenum Press, 1992)
Piperno, D. R. Phytoliths: A comprehensive Guide for Archaeologists and Paleoecologists (Altamira Press, 2006).
Albert, R. M., & Weiner, S. Study of phytoliths in prehistoric ash layers from Kebara and Tabun caves using a quantitative approach. in Phytoliths: applications in earth sciences and human history, (eds. Meunier, J.D. & Colin, F.) 251–266 (Tokyo, Balkema Publisher, 2001)
Albert, R. M. et al. Phytolith-rich layers from the Late Bronze and Iron Ages at Tel Dor (Israel): Mode of formation and archaeological significance. J. Archaeol. Sci. 35(1), 57–75. https://doi.org/10.1016/j.jas.2007.02.015 (2008).
Albert, R. M., Ruíz, J. A. & Sans, A. PhytCore ODB: A new tool to improve efficiency in the management and exchange of information on phytoliths. J. Archaeol. Sci. 68, 98–105 (2016).
Portillo, M., Kadowaki, S., Nishiaki, Y. & Albert, R. M. Early Neolithic household behavior at Tell Seker al-Aheimar (Upper Khabur, Syria): A comparison to ethnoarchaeological study of phytoliths and dung spherulites. J. Archaeol. Sci. 42, 107–118 (2014).
Tsartsidou, G. et al. The phytolith archaeological record: strengths and weaknesses evaluated based on a quantitative modern reference collection from Greece. J. Archaeol. Sci. 34, 1262–1275. https://doi.org/10.1016/j.jas.2006.10.017 (2007).
Neumann, K., Strömberg , A. E. C., Ball, T., Albert, R. M., Vrydaghs, L. Scott-Cummings, L. (International Committee for Phytolith Taxonomy ICPT). International Code for Phytolith Nomenclature (ICPN) 2.0. Annals of Botany, 124(2): 189–199 (2019).
Anderson, P. C. Insight into plant harvesting and other activities at Hatoula, as revealed by microscopic functional analysis of selected chipped stone tools. Le site de Hatoula en Judée occidental. (eds. Lechevallier, M. & Ronen, A.) 277–293 (Paris, Association Paléorient, 1994)
Fugazzola Delpino, M.A. La vita quotidiana del Neolitico. Il sito della Marmotta sul Lago di Bracciano. in Settemila anni fa il primo pane. Ambienti e culture delle società neolitiche (eds. Pessina, A. & Muscio G.) 185–192 (Udine, Museo Friulano di Storia Naturale, 1998–1999)
Mineo, M. Monossili d'Europa: costruite anche per le rotte marine? in Ubi minor: le isole minori del Mediterraneo centrale: dal Neolitico ai primi contatti coloniali (eds. Guidi, A., Cazzella, A. & Nomi, F.). Scienze dell'Antichità 22, 453–475 (2016)
Helwig, K., Monahan, V. & Poulin, J. The identification of hafting adhesive on a slotted antler point from a southwest Yukon ice patch. Am. Antiq. 73, 279–288. https://doi.org/10.1017/S000273160004227X (2008).
Steigenberger, G. & Herm, C. Natural resins and balsams from an eighteenth-century pharmaceutical collection analysed by gas chromatography/mass spectrometry. Anal. Bioanal. Chem. 401, 1771–1784. https://doi.org/10.1007/s00216-011-5169-y (2011).
van den Berg, K. J., Boon, J. J., Pastorova, I. & Spetter, L. F. M. Mass spectrometric methodology for the analysis of highly oxidized diterpenoid acids in Old Master paintings. J. Mass Spectrom. 35, 512–533. https://doi.org/10.1002/(SICI)1096-9888(200004)35:4%3c512::AID-JMS963%3e3.0.CO;2-3 (2000).
Behre K. E. Anthropogenic Indicators in Pollen Diagrams, A.A. (Rotterdam, Balkema, 1986).
Mercuri, A. M. et al. Anthropogenic Pollen Indicators (API) from archaeological sites as local evidence of human-induced environments in the Italian peninsula. Ann. Bot. 3, 143–153 (2013).
Andersen, S.-T., Identification of wild grass and cereal pollen. in Danmarks Geologiske Undersøgelse (ed. Aaby, B.) 69–92 (Geological Survey of Denmark, 1978).
Bottema, S. Cereal-type pollen in the Near East as indicators of wild or domestic crops. in Préhistoire de l’agriculture: nouvelles approches expérimentales et ethnographiques (ed. Anderson P. C.) 95–106 (Paris, CRA, 1992). https://doi.org/10.1007/BF00217499.
Lagerås, P. Long-term history of land-use and vegetation at Femtingagölen—a small lake in the Småland Uplands, southern Sweden. Veg. Hist. Archaeobot. 5, 215–228 (1996).
Joly, C., Barillé, L., Barreau, M., Mancheron, A. & Visset, L. Grain and annulus diameter as criteria for distinguishing pollen grains of cereals from wild grasses. Rev. Palaeobot. Palynol. 146, 221–233. https://doi.org/10.1016/j.revpalbo.2007.04.003 (2007).
Punt, W. Umbelliferae. Rev. Palaeobot. Palynol. 42, 155–364 (1984).
Ellis, M. B. & Ellis, J. P. Microfungi of Land Plants. An Identification Handbook (London, Croom Helm, 1985) (Figure 1270).
Ellis, M. B. & Ellis, J. P. Microfungi of Land Plants. An Identification Handbook (London, Croom Helm, 1985) (Figures 174; 176).
Rottoli, M., Pessina, A. Neolithic agriculture in Italy: an update of archaeobotanical data with particular emphasis on northern settlements. in The Origins and Spread of Domestic Plants in Southwest Asia and Europe. (eds. Colledge, S. & Conolly, J.) 141–154 (Routledge, New York, 2016)
Gurova, M. Prehistoric sickles in the collection of the National Museum of Archaeology in Sofia. in Southeast Europe and Anatolia in Prehistory: Essays in Honor of Vassil Nikolov on his 65th Anniversary (eds. Bacvarov, K. & Gleser, E.) 159–165 (Bonn, Verlag Dr. Rudolf Habelt GmbH, 2016)
Sidéra, I. Nouveaux éléments d'origine proche-orientale dans le Néolithique ancien balkanique. in Analyse de l'industrie osseuse. in Préhistoire d'Anatolie. Genèse de deux mondes (ed. Otte, M.), 215–239 (Liège, ERAUL, 1997)
Mellaart, J. Excavations at Hacılar: Fourth preliminary report, 1960. Anat. Stud. Anat. Stud. 11, 39–75 (1961).
Nag, P. K., Goswami, A., Ashtekar, S. P. & Pradhan, C. K. Ergonomics in sickle operation. Appl. Ergon. 19(3), 233–239 (1988).
Astruc, L., Tkaya, M. B. & Torchy, L. D. l’efficacité des faucilles néolithiques au Proche-Orient: approche expérimentale. Bulletin de la Société préhistorique française 109(4), 671–687 (2012).
Sigaut, F. Les techniques de récolte des grains : identification, localisation, problèmes d'interprétation. in Rites et rythmes agraires (ed. Cauvin, M.-C.) 31–43 (Lyon, Maison de l'Orient et de la Méditerranée Jean Pouilloux, 1991)
Magri, D. Late Quaternary vegetation history at Lagaccione near Lago di Bolsena (central Italy). Rev. Palaeobot. Palynol. 106(3–4), 171–208 (1999).
Gale, R., & Cutler, D. F. Plants in archaeology: identification manual of vegetative plant materials used in Europe and the Southern Mediterranean to c. 1500 (Westbury and Royal Botanic Gardens, Kew, 2000).
Chabal, L. & Feugère, M. L. Le mobilier organique des puits antiques et autres contextes humides de Lattara. Lattara 18, 137–188 (2005).
Chabal, L. (ed.) Quatre puits de l’agglomération routière gallo-romaine d’Ambrussum (Villetelle, Hérault). Supplément. Revue Archéologique de Narbonnaise, 42: 65–71 (2013).
Caruso Fermé, L. & Piqué Huerta, R. Landscape and forest exploitation at the ancient Neolithic site of La Draga (Banyoles, Spain). The Holocene, 24(3): 266 (2014).
Boschian, G. Il Riparo “Ermanno de Pompeis” presso l’Eremo di San Bartolomeo di Legio. Scavi 1990–1999. in Atti della XXXVI Riunione Scientifica IIPP, Preistoria e Protostoria dell’Abruzzo, Chieti-Celano, 27–30 settembre 2001, 105–116 (IIPP; Firenze, 2003).
Radi, G. & Danese, E. L’abitato di Colle Santo Stefano di Ortucchio (L’Aquila). in Atti della XXXVI Riunione Scientifica IIPP, Preistoria e Protostoria dell’Abruzzo, Chieti-Celano, 27–30 settembre 2001, 145–161 (IIPP; Firenze, 2003).
De Francesco, A. M., Bocci, M., Crisci, G. M., & Francaviglia, V. Obsidian provenance at several Italian and Corsican archaeological sites using the non-destructive X-ray fluorescence method. in Obsidian and ancient manufactured glass (eds. Liritzis, I., & Stevenson, C. M.) 115–129 (Albuquerque, UNM Press, 2012).
Degano, I. et al. Hafting of Middle Paleolithic tools in Latium (central Italy): New data from Fossellone and Sant’Agostino caves. PLoS ONE 14, e0213473 (2019).
Nardella, F. et al. Chemical investigations of bitumen from Neolithic archaeological excavations in Italy by GC/MS combined with principal component analysis. Anal. Methods 11, 1449–1459. https://doi.org/10.1039/c8ay02429d (2019).
Rageot, M. et al. Management systems of adhesive materials throughout the Neolithic in the North-West Mediterranean. J. Archaeol. Sci. 126, 105309 (2021).
Binder, D., Bourgeois, G., Benoist, F. & Vitry, C. Identification de brai de bouleau (betula) dans le néolithique de Giribaldi (Nice, France) par la spectrométrie de masse. Revue d’Archéométrie 14, 37–42 (1990).
Vuorela, I. Relative pollen rain around cultivated fields. Acta Bot. Fenn. 102, 1–27 (1973).
Robinson, M. & Hubbard, R. N. L. B. The transport of pollen in the bracts of hulled cereals. J. Archaeol. Sci. 4(2), 197–199. https://doi.org/10.1016/0305-4403(77)90067-X (1977).
Hall, V.A., The role of harvesting techniques in the dispersal of pollen grains of Cerealia. Pollen et Spores, XXX, 2, pp. 265–270.
Portillo, M., Llergo, Y., Ferrer, A. & Albert, R. M. Tracing microfossil residues of cereal processing in the archaeobotanical record: an experimental approach. Veg. Hist. Archaeobot. 26(1), 59–74. https://doi.org/10.1007/s00334-016-0571-1 (2017).
Negri, G. Nuovo erbario figurato (Hoepli ed., Milano, 1981).
Paris R. R. & Moyse H. Matière Médicale. Vol 2°, (Masson, Paris. 1976).
Bulgarelli, G. & Flamigni, S. Le piante tossiche e velenose (Hoepli ed., Milano, 2010).
Les, D. H. Aquatic Dicotyledons of North America: Ecology, Life History, and Systematics (CRC Press, 2017).
Curti, L. Herbarium, un'inedita collezione di piante del XVIII secolo conservata presso l'orto Botanico dell'Università di Padova (Offset Invicta S.p.A., Padova, 1992).
Rottoli, M. Zafferanone selvatico (Carthamus lanatus) e cardo della Madonna (Silybum marianum), piante raccolte o coltivate nel Neolitico antico a “La Marmotta”? Bollettino di Paletnologia Italiana, 91–92, 47–61 (2000–2001).
Rottoli, M. “La Marmotta”, Anguillara Sabazia (RM), scavi 1989. Analisi paletnobotaniche: prime risultanze. Bullettino di Paletnologia Italiana 84, 305–315 (1993).
Van Geel, B. Non-pollen palynomorphs. in Tracking Environmental Change Using Lake Sediments: Terrestrial, vol. 3. (ed. Smol, J. P., Birks, H. J. B., Last W. M.) 99–119 (Algal and Siliceous Indicators, New York, 2001)
Hawksworth, David L., van Geel, Bas, Wiltshire, Patricia E. J. The enigma of the Diporotheca palynomorph. Rev. Palaeobot. Palynol. 235, 94–98 (2016). https://doi.org/10.1016/j.revpalbo.2016.09.010.
Krug, J. C., Benny, G. L., Keller, H. W. Coprophilous fungi. In Biodiversity of Fungi. Inventory and Monitoring Methods (ed. Foster M., Bill, G.) 467–499 (Elsevier Science, Amsterdam, 2004).
Acknowledgements
This research is dedicated to the memory of Filippo Maria Gambari, director of the Museo delle Civiltà between 2017 and 2020. This work has been possible only because of his support. We are also thankful to all the colleagues at the Museo delle Civiltà-Museo Preistorico Etnografico Pigorini and to everyone who made possible the excavation of La Marmotta from its very beginning. Funds have been obtained from the combination of several research projects: the ‘AGER’ project (PGR18BQHM7) funded by the Italian Ministry for Education, Universities and Research within the ‘Rita Levi Montalcini’ program; the project “I mutamenti nella concezione, nell'uso e nella percezione degli spazi”, Dipartimento d'Eccellenza 2019-2022 (429999), University of Pisa; the Marie Sklodowska Curie Individual Fellowship (UE) ‘QUANT’ (792544); the projects ‘La Difusión del Neolítico en el Mediterráneo Centro-Occidental (HAR2016-75201-P)’ and ‘Tools, Techniques and Specialists: the keys to understand the Mesolithic–Neolithic transition in Mediterranean Europe (Tool & Tech)’ (PID2020-112513RB-I00) both funded by the Spanish Ministry of Science, Innovations and Universities and Agencia estatal de investigación; the projects PID2019-105767GB-I00 and PDC2021-121613-I00 funded by MCIN/AEI/10.13039/501100011033 and the European Union “NextGeneration EU”/PRTR. This research is part of a Museo delle Civiltà-CSIC collaboration agreement.
Author information
Authors and Affiliations
Contributions
N.M., J.F.G., M.M.I.: research design and funding; N.M.: use-wear data acquisition and residue sampling; J.F.G., B.G., D.G., N.M.: lithic and use-wear analysis; N.M., J.J.I., F.P.: experimental practices; N.M., J.J.I., F.P.: quantitative analysis through confocal microscopy; M.M.O.: 3D scanning models; G.R., M.M.I.: spatial analysis and contextual information; L.C.: wood taxonomy, sampling and analysis; D.A., R.C.: pollen analysis; L.M.: NPPs analysis; M.P.: phytolith analysis; M.R.A.: organic residue extraction, GC and GC–MS analyses; M.R.O.: archaeobotanical remains. All authors drafted and revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mazzucco, N., Mineo, M., Arobba, D. et al. Multiproxy study of 7500-year-old wooden sickles from the Lakeshore Village of La Marmotta, Italy. Sci Rep 12, 14976 (2022). https://doi.org/10.1038/s41598-022-18597-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-18597-8
This article is cited by
-
Absolute dating of the European Neolithic using the 5259 BC rapid 14C excursion
Nature Communications (2024)
-
Reconstructing dietary practices at Tell Kamid el-Loz (Lebanon) during the Bronze and Iron Age III / Persian to Hellenistic periods using plant micro-remains from dental calculus and stable isotope analysis of bone collagen
Archaeological and Anthropological Sciences (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
