
Abstract
The ladybird Eriopis connexa is an important natural enemy of several pest arthropods in agroecosystems. High population of this predator is frequently observed in strawberry and soybean crops associated with spider mites. We used two-sex life table parameters to evaluate under laboratory conditions, the suitability of three species of spider mites (Tetranychus evansi, Tetranychus urticae, Tetranychus ogmophallos), and a species of aphid (Myzus persicae) as a prey for the predator E. connexa. Eriopis connexa completed immature development on all prey species except on T. evansi, in which all individuals of predator died before reaching the pupal stage. Among prey species that allowed the immature development of E. connexa, T. urticae and M. persicae provided a faster development time to the predator. Oviposition days, longevity and fecundity of E. connexa on T. urticae and M. persicae were substantially longer/higher than on T. ogmophallos. Net reproductive rate (R0), intrinsic rate of increase (r), and finite rate of increase (λ) of E. connexa feeding on T. urticae and M. persicae were also higher than those on T. ogmophallos. Based on the overall performance of the ladybird, the order of suitability of prey species was M. persicae > T. urticae > T. ogmophallos > T. evansi.
Similar content being viewed by others


Introduction
Spider mites and the green peach aphid Myzus persicae (Sulzer) (Hemiptera: Aphididae) are important pests in several crops, with the potential to cause significant yield losses1,2. The two‐spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) is the most important pest mite worldwide for its extensive damage to several crops and numerous cases of pesticide resistance3. Τhe tomato red spider mite Tetranychus evansi Baker and Pritchard (Acari: Tetranychidae), and the peanut red spider mite Tetranychus ogmophallos Ferreira and Flechtmann (Acari: Tetranychidae) are emerging pests in tomato and peanut, respectively, and have caused considerable economic damage to these crops4,5,6.
Chemical control is the main method used for controlling populations of mites and aphids6,7,8. However, the efficacy of this method is not always effective, since these pests become more prone to evolving pesticide resistance due to their high biotic potential and high genetic variability3,4,6,7. Intensification of chemical sprays has led also to environmental pollution, and poisoning risks to farmers and consumers8,9. For this reason, alternative control measures have been sought worldwide9,10,11. Biological control is a potential alternative control method, mainly when combined with other control measures, thus helping to implement integrated pest management programs11,12.
Ladybirds (Coleoptera: Coccinellidae) have been used in farm systems as natural enemies of phytophagous arthropods and maintained some pests below the level of economically significant damage12,13,14. Among these ladybirds, Eriopis connexa (Germar) (Coleoptera: Coccinellidae) has a high pest-control potential due to its high foraging capacity, voracity, and polyphagy14,15,16,17. This ladybird is a Neotropical predator of several pest arthropods, including eggs of Diuraphis noxia (Mordvilko), Rhopalosiphum maidis (Fitch), Rhopalosiphum padi (L.), Acyrthosiphon pisum (Harris), Schizaphis graminum (Rondani), Cinara atlantica (Wilson) (Hemiptera: Aphididae), Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), Diatraea saccharallis (Fabricius) (Lepidoptera: Pyralidae), and Macrosiphum euphorbiae (Thomas) (Hemiptera: Aphididae)12,15,17,18,19,20.
Eriopis connexa is a control agent for aphids in pine, cotton, wheat, and citrus plants13. In Brazil, a field study of key mortality factors of Myzus persicae (Sulzer) (Homoptera: Aphididae) in cabbage recorded E. connexa as a potential control for this pest21. This predator was efficacious in controlling Chaetosiphon fragaefolii Cockerell (Hemiptera: Aphididae) on strawberry in a greenhouse in Argentina22. Because of its potential as an aphid predator, this ladybird was introduced into the United States to control the Russian wheat aphid, D. noxia and pea aphid A. pisum20,23. Preliminary investigations indicated that E. connexa is often associated with spider mites in strawberry and soybean crops (Matos and Andrade, personal information). However, studies that evaluated the suitability of spider mites for E. connexa performance are scarce. Knowing the suitability of a predator on different prey species can be useful for practical purposes in biological control programs24,25.
Life table parameters are accurate and reliable tools to track how prey species can be suitable for natural enemies given that such tools generate detailed information about the development, survival, longevity, fecundity, and life expectancy of a population26. These tools have been used to evaluate the suitability of several pests to various natural enemies27,28. To date, studies assessing suitability of prey species to E. connexa are limited only to some parameters such as immature development phase, longevity, and fecundity13,16,18,29. Furthermore, few studies that used life table parameters to evaluate suitability of prey species to this predator have been based on a female age-specific life table method16,30,31. In this method, only female individuals are taken into consideration disregarding the male population and variation in developmental rates among individuals of a population. According to Chi and Liu32, this may result to errors in the estimation of life table parameters. Given these limitations, Chi and Liu26 and Chi32 developed a theoretical model of life table analysis namely “two-sex life table”. That method considers ages or stages and development rates of both sexes. Such design allows deeper knowledge on biology of predators and population growth parameters, which are fundamental for pest management efficiency. Therefore, this study aimed to use the two-sex life table to evaluate the suitability of three species of spider mites (T. evansi, T. urticae, T. ogmophallos) and a species of aphid M. persicae as prey for E. connexa.
Material and methods
Eriopis connexa rearing
Rearing of E. connexa was initiated with specimens of adults collected from volunteer plants. Ladybirds were kept in cages made of a PVC tube (10-cm-height by 10-cm-diameter), lined internally with bond paper, and sealed with voile fabric. The insects were fed with eggs of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) and an artificial diet, which consisted of beer yeast and honey (1:1). Eggs laid by E. connexa were removed daily from the cages and kept in 12.0-cm diameter Petri dishes sealed with PVC film. The newly hatched larvae were transferred to new Petri dishes and fed with eggs of E. kuehniella. Water was supplied with a moistened polyethylene sponge33. The insects were maintained in a climate-controlled room at 25 ± 1 °C, 60 ± 10% relative humidity, and 12-h photoperiod.
Plant materials
The seedlings of different plants used for the rearing of aphid and spider mites species in this study were purchased from "Casa de sementes" (Jaboticabal, São Paulo, Brazil). The plant materials used were obtained with prior permission, and the present study is in compliance with relevant guidelines and legislation.
Myzus persicae rearing
Rearing of M. persicae started with insects collected from cabbage plants (Brassica oleracea L. var. Acephala) at São Paulo State University, Jaboticabal Campus. The rearing substrate was leaf discs (2.5 cm in diameter) from cabbage leaves with the abaxial side facing upward, which were placed in 5.0-cm-diameter Petri dishes filled with a 5.0-mm layer of 1% agar-water, for disc turgidity. Nymphs and adults of M. persicae were placed on the discs, and the Petri dishes were sealed with PVC film and kept in a climate-controlled chamber at 23 ± 1 °C, 70 ± 10% RH, and 12-h photoperiod. The agar-water layer and leaf discs were changed twice a week.
Rearing of spider mites
The original colonies of T. urticae, T. evansi, and T. ogmophallos used in this study was established from jack bean (Canavalia ensiformis L. cv. commun), tomato (Solanum lycopersicum L. var. cerasiforme), and peanut (Arachis hypogaea L., cv. Granoleico) plants, respectively, maintained in a screen house of the Acarology laboratory at São Paulo State University, Jaboticabal Campus, Brazil. After identification of the spider mite species, live specimens from each species were used to establish the laboratory colony on the same plant species as in the original colony. The host plants were 35–60 day-old and were grown into 5-L pots which were by 80% filled with a homogeneous mixture of soil, sand, and tanned bovine(1:1:1). All spider mite specie were kept climate-controlled rooms at 25 ± 1 °C, 60 ± 10% relative humidity, and 12-h photoperiod. Plants deteriorated by mites and/or senescent were periodically replaced with new ones.
Experimental procedure
Development and reproduction of E. connexa were evaluated in four groups of prey diet: T. urticae, T. evansi, T. ogmophallos, and M. persicae under the same conditions described above for the stock colony. Initially, 10 pairs of freshly emerged females and males of E. connexa were maintained in a 350-mL transparent plastic cup (7 cm in diameter and 10 cm in height) covered by fine nylon netting (40 mesh), to mate and lay eggs. Fifty freshly deposited eggs (< 1-day old) were transferred into 12.0-cm-diameter Petri dishes sealed with PVC film until hatching (Fig. 1). Newly hatched larvae (< 24 h old) were placed individually in 9-cm-diameter Petri dishes, sealed with PVC film. A cohort of 50 larvae of E. connexa was used per prey diet (number of replications). A surplus of 200 individuals with the same ratio of different stages of prey was provided to each predator stage daily to ensure an abundance of food. Upon reaching adulthood, ladybirds were sexed and transferred in couples to new plastic cups. In the case of treatments where the number of females emerged was greater than the number of males, some males from the rearing stock were used to form couples (Fig. 1). As a source of water, a small Petri dish with wetted cotton wool (3 cm in diameter) was used. The experimental units were examined every 24 h to determine the duration of each developmental stage and survivorship. The pre-oviposition period (APOP: period between adult emergence and its first oviposition), total pre-oviposition period (TPOP: period from egg to first oviposition), oviposition days (number of days in which oviposition occurred), longevity of each sex, sex ratio and fecundity were also determined. In units where males died before females, other males from the stock colony were used to replace them. Data on males that came from stock colonies to form the couples or those used to replace died males before females were not used in statistical analysis. Eggs laid were removed at each observation time. The experiment was considered complete after all predators had died.

Experimental set up of Eriopis connexa reared on Tetranychus evansi. The same experimental set up was followed for the experiments of Eriopis connexa on Tetranychus urticae, Tetranychus ogmophallos and M. persicae. Number of replications = 50.
Complementary test
As E. connexa was unable to complete its immature development phase feeding only on T. evansi, a complementary test was performed to evaluate its development when fed with T. evansi combined with a complementary food source. For this purpose, ladybird larvae were daily fed only with T. evansi until the fourth instar, and after that, T. urticae were added to the diet. Larval mortality and adult emergence were recorded.
Statistical analyses
The software TWOSEX-MSChart by Chi34 available at http://140.120.197.173/Ecology/prod02.htm was used to estimate development and reproduction raw data and to calculate population parameters, using the procedure “two-sex life table”26,32. The following parameters were estimated: age-stage–specific survival rate (sxj), age-specific survival rate (lx), age-specific fecundity (mx), net reproduction rate (R0), intrinsic rate of increase (r), stage-specific fecundity (fxj), finite rate of increase (λ), average generation time (T), age-stage life expectancy (exj), and age-stage reproductive value (vxj).
The variance and standard errors of development, fecundity, reproduction period, population parameters and survival curves were estimated using the bootstrap35,36,37,38. During this procedure, data of each of these biological parameters were re-sampled 100,000 times. Differences among treatments were compared by the paired bootstrap test, based on the 95% confidence interval of differences implemented in the TWOSEX-MSChart39,40.
Results
Life stage duration, fecundity, sex ratio, and life table parameters
The duration of the larval stages of the E. connexa was substantiality shorter on M. persicae and T. urticae than on T. ogmophallos, and T. evansi, except for the first instar of the last prey species (Table 1). Similar pattern was observed for the predator pupal stage, also shorter on the former two prey species (Table 1). Thus, the immature development time for both sexes was shorter on M. persicae (18.4 ± 0.28 and 18.3 ± 0.31 days, respectively for male and female) and T. urticae (18.7 ± 0.27 for both sexes) than on T. ogmophallos (22.7 ± 0.24 and 22.8 ± 0.24 days, respectively for male and female). The immature survival of E. connexa fed with M. persicae and T. urticae was higher than 95% (Table 1). Sex ratio was significantly higher on the latter previously mentioned prey species than T. ogmophallos. Eriopis connexa fed with T. evansi could not complete its developmental cycle, with no larva reaching the pupal stage (Table 1). However, in the complementary test, E. connexa larvae fed with T. evansi completed their biological cycle after T. urticae was added to the diet, at the beginning of the fourth instar (Table 2).
Pre-oviposition and total pre-oviposition periods were significantly shorter for E. connexa fed with M. persicae and T. urticae compared to those fed T. ogmophallos (Table 3). Likewise, longevity, fecundity, and oviposition days increased for the predator when with fed T. urticae and M. persicae (Table 3). All parameters for the predator population growth were significantly influenced by the prey species (Table 3). Net reproduction rate (R0), intrinsic rate of increase (r), and finite rate of increase (λ) were significantly higher for ladybirds fed with M. persicae and T. urticae compared to those fed with T. ogmophallos.
Age- and stage-specific survival and fecundity rate
Survival-rate curves (sxj) show the probability of a freshly oviposited egg to survive to age x and develop to stage j (Fig. 2). Due to changes in the development and survival, the stages of the predator fed with different prey, the sxj curves showed significant overlap. The probability of a freshly laid egg surviving to the adult stage was higher for ladybird adults fed with T. urticae and M. persicae (0.5 for females and 0.46 for males) than for those fed with T. ogmophallos (0.34 for females and 0.40 for males). The probability of newly hatched larvae of the predator reaching the adult stage of when feeding on T. evansi was zero (Fig. 2).

Age-stage-specific survival rate (sxj) of Eriopis connexa fed on Tetranychus urticae, Tetranychus evansi, Tetranuchus ogmophallos, or Myzus persicae.
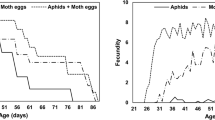
Age-specific survival rate (lx) expresses the survival probability that an individual of E. connexa would survive until age x (Fig. 3). The 84% of E. connexa individuals fed with M. persicae and T. urticae remained alive for 96.0 ± 5.14 days and 98.0 ± 6.03 days, respectively, showing a significantly higher lx than that of T. ogmophallos (6.0 ± 3.3 days) (Fig. 3). Similar patterns were observed for 50% and 16% of ladybirds’ survival on M. persicae (110.0 ± 1.79 and 122.0 ± 2.06 days, respectively) and T. urticae (109 ± 1.06 and 122.0 ± 2.85 days, respectively) which were higher than on T. ogmophallos (45.0 ± 2.37 and 57.0 ± 3.48 days, respectively). Age-stage-specific fecundity fx4 (daily number of eggs produced per female of age x) showed higher egg-laying peaks for females of E. connexa fed with T. urticae (11.04 eggs) and M. persicae (10.03 eggs) on the 58th and 63rd day of their age, respectively. For female adults fed with T. ogmophallos, the egg-laying peak (4.4 eggs) occurred at 38 days of age (Fig. 3).

Age-specific survival rate (lx), age-specific fecundity (mx), and age-stage-specific fecundity (fxj) of Eriopis connexa fed on Tetranychus urticae, Tetranychus ogmophallos, or Myzus persicae.
Age-specific fecundity mx (average daily fecundity per individuals at age x) showed the highest value when E. connexa fed with T. urticae (5.83 eggs) and M. persicae (5.13 eggs) at 57 and 49 days of age, respectively (Fig. 3). The life expectancy of freshly laid eggs (e01) was higher (about 107 days) for E. connexa fed with T. urticae and M. persicae than for those fed with T. ogmophallos (38.7 days) (Fig. 4). Therefore, adult females at 60 days of age can still live about 52 days feeding on these prey species. In contrast, the life expectancy was 18 and 39 days in adult ladybirds fed with T. evansi and T. ogmophallos, respectively.

Age-stage life expectancy (exj) of Eriopis connexa fed on Tetranychus urticae, Tetranychus evansi, Tetranychus ogmophallos, or Myzus persicae.
Age-stage reproductive value (vxj) of E. connexa adult females contributed more to population growth than other development stages. The peaks (vxj) of adult females fed with M. persicae, T. urticae, or T. ogmophallos were 58.4, 55.8, and 13.5, respectively (Fig. 5).

Age-stage reproductive value (vxj) of Eriopis connexa fed on Tetranychus urticae, Tetranychus ogmophallos, or Myzus persicae.
Discussion
Prey quality is a key factor affecting the growth, development and reproduction of predatory insects41. The suitability of a prey species can be evaluated by measuring its effect on biological attributes of the predator42. Prey are categorized as essential (supports development and reproduction), alternative (supports only survival) or rejected on the basis of quantitative data on the developmental rate, survival and reproductive capacity43,44,45. Due to the importance of predatory ladybirds in biological control, this study investigated effects of three spider mite species (T. evansi, T. urticae, and T. ogmophallos) and an aphid species (M. persicae) on the development, survival and reproductive performance of the predator E. connexa. Our findings indicated that E. connexa was able to complete its life cycle and reproduce on all prey studied, except for T. evansi, a diet that resulted in 100% mortality of the predator before reaching the pupal stage. However, the prey species that allowed the development of E. connexa had a strong influence on demographic parameters of the predator. For instance, E. connexa fed with T. urticae and M. persicae showed a faster immature developmental period and high survival rate when compared to the T. ogmophallos diet. Furthermore, the oviposition time, longevity for both sexes and fecundity on T. urticae and M. persicae diets were longer/higher when compared to T. ogmophallos. These results suggest that T. urticae and M. persicae are more suitable prey species for E. connexa, probably due to the quality difference in nutrient contents of the prey species46,47,48. Several studies reported that the variation of the chemical profiles, morphological or allelochemical features of host plants have also a direct effect on the nutritional value of herbivorous arthropods, in terms of their suitability for predators41,49,50. In our study, the prey species have been reared on different host plant families (B. oleracea, C.ensiformis, A.hypogaea cv. Granoleico and S. lycopersicum) that will presumably tend to have higher differences. Therefore, although not being evaluated in the present study, this fact could also have contributed as an additional cause of difference in the quality of prey species that affected development, survival and reproductive performance of E. connexa.
Most of the results obtained in this study are comparable to those of previously studies, which report that the type of prey had a significant influence on the developmental period, survival rate, and reproduction of several other ladybirds47,51,52. For instance, the duration of E. connexa immature phase (larva-adult) reported by Silva et al.16 on five prey species [immature of E. kuehniella, S. frugiperda, D. saccharalis, R. maidis, S. graminum] at 25 or 26 °C, lasted 12.6–17.4 days, values similar to those found in our study. Values of E. connexa development time (larva-adult) found by Nascimento et al.16 on Plutella xylostella (L.) larvae, the brassica aphid, Lipaphis pseudobrassicae (Davis) when provided separate or mixed are also comparable (17.1–24.7 days) to those of this study. On the other hand, Duarte et al.30 reported higher developmental duration (22.9–47.48 days) on three aphid species [Brevicoryne brassicae L., Macrosiphum euphorbiae Thomas, and Pterocallis sp. (Hemiptera: Aphididae)], but similar survival rates (37.5–97.5%) with our findings.
Sarmento et al.53 have reported an exponential (Type I) functional response for adults of E. connexa fed on T. evansi, but the authors did not assess the subsequent effects on different developmental stages and reproduction resulting from that predator feeding with T. evansi diet, an aspect fundamental in conservation and growth of the predator in agro-system. In this study, we found that T. evansi is unsuitable for the development and reproduction of E. connexa. Similar results have been reported by Oliveira et al.54 for Cycloneda sanguinea (Linnaeus) (Coleoptrea: Coccinellidae) which did not complete successfully its biological cycle feeding on T. evansi. Indeed, the mite is known as prey that sequesters toxic secondary metabolites from plants and accumulates them in its body55, which possibly makes it unsuitable as a prey for E. connexa. This is also supported by the increased mortality during the last instar, since ladybird larvae need more nutrients to reach the pupal and adult stage56, i.e., food consumption and intake of toxic compounds increased during L457. Furthermore, when the diet changed to T. urticae at the beginning of the fourth instar, the predator successfully completed the immature development phase. This result also indicates the toxic effect of T. evansi on E. connexa development can be reversible in the presence of other diet. Likewise, Munyaneza and Obrycki58 demonstrated that Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae) eggs are an adequate diet for Coleomegilla maculata DeGeer (Coleoptera: Coccinellidae) only when the larva consumes aphids in the early stages of development. Nascimento et al.16 also found that the performance (developmental time and survival) of E. connexa improved when larvae fed on diets of mixed prey compared to simple P. xylostella larvae prey diet. However, the effects of mixed diets that include T. evansi need to be assessed in further studies to investigate this relationship.
Zazycki,et al.31 reported longevity and fecundity of E. connexa on pollen + E. kuehniella eggs at 25 °C (60 –130 days; 584 ± 96.50 eggs/female respectively), which were comparable to those found on T. urticae and M. persicae in this study. The reduced longevity and very low fecundity of E. connexa on T. ogmophallos could be possibly due to lack of some nutrients essential for full longevity and good reproductive performance. According to Adams59 and Lima et al.60, adult females need suitable sources of nutrients to develop mature ovaries and produce eggs. Given that the period of the first oviposition (APOP and TPOP) was substantially longer for E. connexa on T. ogmophallos, it may be that the insufficient source of nutrients in this prey prolonged maturation of the predator’s ovaries before the first oviposition. The results are in compliance with those by Omkar and James47 and Tian et al.25 who report that prey suitability affects the oviposition of Coccinella transversalis Fabricius, Coccinella undecimpunctata (Linnaeus), and Serangium japonicum Chapin (Coleoptera: Coccinellidae). Furthermore, females that reached reproductive maturity by feeding on low-quality diet and acquired low energy during their development may be able to control the sex ratio of their offspring61. This would justify the male biased sex ratio (0.46) of E. connexa on T. ogmophallos, while the ratio is 1:1 (0.52) on T. urticae and M. persicae. Thus rearing of E. connexa on T. ogmophallos could lead to male individuals in the population, in response to the poor quality of the prey species and as a strategy to reproduce. However, further investigations should be carried out to demonstrate the likely impacts of these lower quality prey species on the predator.
The intrinsic rate of increase (r) of E. connexa, reflecting the effects of biological parameters such as development, survival, fecundity, and sex ratio50 on the predator’s population, was higher on T. urticae and M. persicae (0.118–0.126 day−1) compared to that on T. ogmophallos (0.047 day−1). Zazycki et al.31 under similar conditions reported a r comparable value (0.126 day−1) on pollen and E. kuehniella eggs for the aforementioned prey species.
Ocerall, these findings suggest that T. urticae and M. persicae are more suitable prey species for E. connexa than T. ogmophallos, while T. evansi is not suitable for this predator. Although enlightening, these results are certainly preliminary for the actual potential of E. connexa as biological control agent of spider mites and M. persicae in the field. Given that under natural conditions E. connexa is a generalist predator and can consume alternative as well as essential prey items, the performance of E. connexa could improve even with low-quality prey. The next steps should include predation capacity evaluation, weight and size of different life stages as well as field studies in order to determine the performance of this predator for the control of spider mites and M. persicae in the agroecosystems.
References
Sun, M. et al. Reduced phloem uptake of Myzus persicae on an aphid resistant pepper accession. BMC Plant Biol. 18, 1–14 (2018).
Migeon, A. & Dorkeld, F. Spider Mites Web: A comprehensive database for the Tetranychidae. Available at http://www.montpellier.inra.fr/CBGP/spmweb (2021)
Sato, M. E. et al. Spiromesifen resistance in Tetranychus urticae (Acari: Tetranychidae): Selection, stability, and monitoring. Crop Prot. 89, 278–283 (2016).
Melville, C. C., Andrade, S. C., Oliveira, N. T. & Andrade, D. J. Impact of Tetranychus ogmophallos (Acari: Tetranychidae) on different phenological stages of peanuts. Bragantia 77, 116–123 (2018).
Savi, P. J., de Moraes, G. J., Melville, C. C. & Andrade, D. J. Population performance of Tetranychus evansi (Acari: Tetranychidae) on African tomato varieties and wild tomato genotypes. Exp. Appl. Acarol. 77, 555–570 (2019).
Bass, C. et al. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 51, 41–51 (2014).
Tabet, V. G., Vieira, M. R., Martins, G. L. M. & Sousa, C. G. N. M. Plant extracts with potential to control of two-spotted spider mite. Arq. Inst. Biol. 85, 1–8 (2018).
Özkara, A., Akyil, D. & Konuk, M. Pesticides, environmental pollution, and health. In Environmental Health Risk—Hazardous Factors to Living Species (eds Larramendy, M. & Soloneski, S.) (InTech, 2016).
Faraone, N., Evans, R., LeBlanc, J. & Hillier, N. K. Soil and foliar application of rock dust as natural control agent for two-spotted spider mites on tomato plants. Sci. Rep. 10, 12108 (2020).
Taghizadeh, M., Iraninejad, K. H., Iranipour, S. & Vahed, M. M. Comparative study on the efficiency and consumption rate of Stethorus gilvifrons (Coleoptera, Coccinellidae) and Orius albidipennis (Hemiptera, Anthocoridae), the predators of Tetranychus urticae Koch (Acari, Tetranychidae). North-West. J. Zool. 16, 125–133 (2020).
Orr, D. & Lahiri, S. Biological control of insect pests in crops. In Integrated Pest Management: Current Concepts and Ecological Perspective (ed. Abrol, D. P.) 531–548 (Academic Press, 2014).
Oliveira, N. C., Wilcken, C. F. & Matos, C. A. O. Biological cycle and predation of three coccinellid species (Coleoptera, Coccinellidae) on giant conifer aphid Cinara atlantica (Wilson) (Hemiptera, Aphididae). Rev. Bras. Entomol. 48, 529–533 (2004).
Moghaddam, M. G. et al. Demographic traits of Hippodamia variegata (Goeze) (Coleoptera: Coccinellidae) fed on Sitobion avenae Fabricius (Hemiptera: Aphididae). J. Crop Prot. 5, 431–445 (2016).
Gómez, W. D. & Polanía, I. Z. Life table of the predatory beetle Eriopis connexa (Germar) (Coleoptera: Coccinellidae). Rev. UDCA Act. Divulg. Cient. 12, 147–155 (2009).
Silva, R. B. et al. Biological aspects of Eriopis connexa (Germar) (Coleoptera: Coccinellidae) fed on different insect pests of maize (Zea mays L.) and sorghum [Sorghum bicolor L. (Moench)]. Braz. J. Biol. 73, 419–424 (2013).
Nascimento, D. V., Lira, R., Ferreira, E. K. S. & Torres, J. B. Performance of the aphidophagous coccinellid Eriopis connexa fed on single species and mixed-species prey. Biocontrol. Sci. Technol. 31, 951–963 (2021).
Miller, J. C. A comparison of techniques for laboratory propagation of a South American ladybeetle, Eriopis connexa (Coleoptera: Coccinellidae). Biol. Control 5, 462–465 (1995).
Miller, J. C. & Paustian, J. W. Temperature-dependent development of Eriopis connexa (Coleoptera: Coccinellidae). Environ. Entomol. 21, 1139–1142 (1992).
Sarmento, R. A. et al. Fat body morphology of Eriopis connexa (Coleoptera, Coccinellidae) in function of two alimentary sources. Braz. Arch. Biol. Technol. 47, 407–411 (2004).
Van Driesche, R. et al. Catalog of Species Introduced into Canada, Mexico, the USA, or the USA Overseas Territories for Classical Biological Control of Arthropods 1985–2018 (USDA Forest Service, 2018).
Fidelis, E. G. et al. Coccinellidae, Syrphidae and Aphidoletes are key mortality factors for Myzus persicae in tropical regions: A case study on cabbage crops. Crop Prot. 112, 288–294 (2018).
Francesena, N. et al. Potential of predatory Neotropical ladybirds and minute pirate bug on strawberry aphid. Nat. Acad. Bras. Ciênc. 91, e20181001 (2019).
Reed, D. K. & Pike, K. S. Summary of an exploration trip to South America. IOBC Nearctic Reg. Newsl. 36, 16–17 (1991).
Li, Y., Zhou, X., Duan, W. & Pang, B. Food consumption and utilization of Hippodamia variegata (Coleoptera: Coccinellidae) is related to host plant species of its prey, Aphis gossypii (Hemiptera: Aphididae). Acta Entomol. Sin. 58, 1091–1097 (2015).
Tian, M., Wei, Y., Zhang, S. & Liu, T. Suitability of Bemisia tabaci (Hemiptera: Aleyrodidae) biotype-B and Myzus persicae (Hemiptera: Aphididae) as prey for the ladybird beetle, Serangium japonicum (Coleoptera: Coccinellidae). Eur. J. Entomol. 114, 603–608 (2017).
Chi, H. Life table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 17, 26–34 (1988).
Midthassel, A., Leather, S. R. & Baxter, I. H. Life table parameters and capture success ratio studies of Typhlodromips swirskii (Acari: Phytoseiidae) to the factitious prey Suidasia medanensis (Acari: Suidasidae). Exp. Appl. Acarol. 61, 69–78 (2013).
Hosseini, A. et al. Life history responses of Hippodamia variegata (Coleoptera: Coccinellidae) to changes in the nutritional content of its prey, Aphis gossypii (Hemiptera: Aphididae), mediated by nitrogen fertilization. Biol. Control 130, 27–33 (2019).
Almeida, D. P., Berber, G. C. M., Aguiar-Menezes, E. L. & Resende, A. L. S. Evaluation of biological parameters of Eriopis connexa (Germar, 1824) and Coleomegilla maculata (DeGeer, 1775) (Coleoptera: Coccinellidae) fed with alternative prey developed at the integrated center for pest management - UFRRJ. Sci. Electron. Arch. 14, 8–16 (2021).
Duarte, W., Arévalo, H. & Polanía, I. Z. Influence of three aphid species used as prey on some biological aspects of the predator Eriopis connexa. J. Anim. Sci. 3, 193–199 (2013).
Zazycki, L. C. F. et al. Biology and fertility life table of Eriopis connexa, Harmonia axyridis and Olla v-nigrum (Coleoptera: Coccinellidae). Braz. J. Biol. 75, 969–973 (2015).
Chi, H. & Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 24, 225–240 (1985).
Santos, N. R. et al. Biological aspects of Harmonia axyridis fed on two prey species and intraguild predation with Eriopis connexa. Pesq. Agropec. Bras. 44, 554–560 (2009).
Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. National Chung Hsing University, Taichung, Taiwan. http://140.120.197.173/Ecology/prod02.htm (2021)
Efron, B. & Tibshirani, R. J. An Introduction to the Bootstrap (Springer, 1993). https://doi.org/10.1007/978-1-4899-4541-9.
Hesterberg, T. It’s time to retire the ‘n > = 30’ rule. In Proceedings of the American Statistical Association, Statistical Computing Section (CD-ROM). http://home.comcast.net/~timhesterberg/articles/JSM08-n30.pdf (2008)
Huang, Y. B. & Chi, H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 19, 263–273 (2012).
Akkopru, E. P., Atlıhan, R., Okut, H. & Chi, H. Demographic assessment of plant cultivar resistance to insect pests: A case study of the dusky-veined walnut aphid (Hemiptera: Callaphididae) on five walnut cultivars. J. Econ. Entomol. 108, 378–387 (2015).
Smucker, M. D., Allan, J. & Carterette, B. A comparison of statistical significance tests for information retrieval evaluation. In Proceedings of the Sixteenth ACM Conference on Conference on Information and Knowledge Management—CIKM ’07 623 (ACM Press, 2007).
Wei, M. et al. Demography of Cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of Pyrus bretschneideri (Rosales: Rosaceae) and P. communis pears with estimations of confidence intervals of specific life table statistics. J. Econ. Entomol. 113, 2343–2353 (2020).
Wu, X., Zhou, X. & Pang, B. Influence of five host plants of Aphis gossypii Glover on some population parameters of Hippodamia variegata (Goeze). J Pest Sci 83, 77–83 (2010).
Kalushkov, P. & Hodek, I. New essential aphid prey for Anatis ocellata and Calvia quatuordecimguttata (Coleoptera: Coccinellidae). Biocontrol Sci. Technol. 11, 35–39 (2001).
Hodek, I. & Honěk, A. Ecology of Coccinellidae (Springer, 1996).
Hodek, I. & Evans, E. Food relationships. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae) (eds Hodek, I. et al.) (Wiley, 2012).
Pervez, A. & Kumar, R. Preference of the aphidophagous ladybird Propylea dissecta for two species of aphids reared on toxic host plants. Eur. J. Environ. Sci 7, 130–134 (2017).
Omkar, & Bind, R. B. Prey quality dependent growth, development and reproduction of a biocontrol agent, Cheilomenes sexmaculata (Fabricius) (Coleoptera: Coccinellidae). Biocontrol Sci. Technol. 14, 665–673 (2004).
Omkar, & James, B. E. Influence of prey species on immature survival, development, predation and reproduction of Coccinella transversalis Fabricius (Col., Coccinellidae). J. Appl. Entomol. 128, 150–157 (2004).
Farooq, M., Shakeel, M., Iftikhar, A., Shahid, M. R. & Zhu, X. Age-stage, two-sex life tables of the lady beetle (Coleoptera: Coccinellidae) feeding on different aphid species. J. Econ. Entomol. 111, 575–585 (2018).
Giles, K. L. et al. Host plants affect predator fitness via the nutritional value of herbivore prey: Investigation of a plant-aphid-ladybeetle system. Biocontrol 47, 1–21 (2002).
Savi, P. J., de Moraes, G. J. & Andrade, D. J. Effect of tomato genotypes with varying levels of susceptibility to Tetranychus evansi on performance and predation capacity of Phytoseiulus longipes. Biocontrol 66, 687–700 (2021).
Hodek, I. & Honěk, A. Scale insects, mealybugs, whiteflies and psyllids (Hemiptera, Sternorrhyncha) as prey of ladybirds. Biol. Control 51, 232–243 (2009).
Qureshi, J. A. & Stansly, P. A. Three Homopteran pests of citrus as prey for the convergent lady beetle: Suitability and preference. Environ. Entomol. 40, 1503–1510 (2011).
Sarmento, R. A. et al. Functional response of the predator Eriopis connexa (Coleoptera: Coccinellidae) to different prey types. Braz. Arch. Biol. Technol. 50, 121–126 (2007).
Oliveira, E. E. et al. Biological aspects of the predator Cycloneda sanguinea (Linnaeus, 1763) (Coleoptera: Coccinellidae) fed with Tetranychus evansi (Baker and Pritchard, 1960) (Acari: Tetranychidae) and Macrosiphum euphorbiae (Thomas, 1878) (Homoptera: Aphididae). Biosci. J. 21, 33–39 (2005).
de Moraes, G. J., McMurtry, J. A. & Baker, E. W. Redescription and distribution of the spider mites Tetranychus evansi and T. marianae. Acarologia 28, 333–343 (1987).
Iperti, G. Biodiversity of predaceous coccinellidae in relation to bioindication and economic importance. Agric. Ecosyst. Environ. 74, 323–342 (1999).
Cruz-Rivera, E. & Hay, M. E. Prey nutritional quality interacts with chemical defenses to affect consumer feeding and fitness. Ecol. Monogr. 73, 483–506 (2003).
Munyaneza, J. & Obrycki, J. J. Development of three populations of Coleomegilla maculata (Coleoptera: Coccinellidae) feeding on eggs of colorado potato beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 27, 117–122 (1998).
Adams, T. S. Effect of diet and mating status on ovarian development in a predaceous stink bug Perillus bioculatus (Hemiptera: Pentatomidae). Ann. Entomol. Soc. Am. 93, 529–535 (2000).
Lima, M. S., Pontes, W. J. T. & Nóbrega, R. L. Pollen did not provide suitable nutrients for ovary development in a ladybird Brumoides foudrasii (Coleoptera: Coccinellidae). Diversitas J. 5, 1486–1494 (2020).
Melville, C. C. et al. Peanut cultivars display susceptibility by triggering outbreaks of Tetranychus ogmophallos (Acari: Tetranychidae). Exp. Appl. Acarol. 78, 295–314 (2019).
Acknowledgements
We thank the Coordination for the Improvement of Higher Education Personnel (CAPES), Brazil—Finance Code 001, for granting scholarships to the authors S.T.S. Matos, P.J. Savi and C.C. Melville, and the National Council for Scientific and Technological Development (CNPq) for granting the research productivity scholarship to D.J. Andrade (Process No. 301492/2018-2). We also thank Professor Hsin Chi (National Chung Hsing University, Taichung, Taiwan, Republic of China) for assisting in the data analysis.
Author information
Authors and Affiliations
Contributions
D.J.A. and S.T.S.M. conceived the ideas and designed methodology; S.T.S.M. collected the data; P.J.S. participated in the data analysis; S.T.S.M., P.J.S., C.C.M., T.M.S.C., F.J.C. and D.J.A. led writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
de Matos, S.T.S., Savi, P.J., Melville, C.C. et al. Suitability of spider mites and green peach aphids as prey for Eriopis connexa (Germar) (Coleoptera: Coccinellidae). Sci Rep 12, 8029 (2022). https://doi.org/10.1038/s41598-022-12078-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-12078-8
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
