
Abstract
The vertical migration and accumulation of Microcystis colonies is a critical process in algal bloom formation. This work explored the effect of wind and light intensity on the vertical migration of Microcystis colonies. The wind-driven currents, light-driven changes in mass density of colonies, and the effect of colony size was coupled to simulate the vertical motion of colonies via Ansys Fluent and MATLAB. Results showed that light causes Microcystis to exhibit a ‘day-sinking and night-floating’ (d-n) phenomenon, however, wind weakens the phenomenon by forming a turbulent drag force that inhibits the vertical movement of Microcystis. This study proposed a kinetic ratio-based method, that there is a specific equilibrium turbulent kinetic energy and when turbulent kinetic energy of the water body is greater than the equilibrium turbulent kinetic energy, the d-n phenomenon does not occur. For Lake Taihu, the wind-driven turbulent kinetic energy is usually greater than the equilibrium turbulent kinetic energy. Therefore, Microcystis colonies may not exhibit the d-n phenomenon. Our findings provide a new theoretical basis for current process-based models in simulating algal blooms in large shallow lakes.
Similar content being viewed by others

Introduction
Algal blooms caused by lake eutrophication are of a global environmental and human health concern, threatening our drinking water supplies, and the ecological and economic sustainability of our freshwater ecosystems1. Microcystis is generally the dominant algal species in eutrophic lakes. Its strong vertical migration ability allows Microcystis to compete better than other algae for light and nutrients by occupying an optimized position2. Lake Taihu is the third largest freshwater lake in China, with an area of 2360 km2, and algal blooms have a serious impact on the lake environment. In Lake Taihu, Microcystis comprises more than 85% of the summer phytoplankton biomass3. Therefore, studying the vertical movement of Microcystis is essential for understanding its growth and treatment for eliminating algal blooms. Three factors typically affect the floating/sinking of Microcystis in water: the mass density of the Microcystis population4, wind-driven currents5, and Microcystis colony size6.
Ibelings et al.7 noticed that changes in light intensity affect photosynthesis and respiration processes of Microcystis, resulting in variations in the mass density of colonies. The buoyancy force varies with mass density, causing the Microcystis to sink gradually during the day and float at night. Previous studies cultured Microcystis aeruginosa8 and cyanobacteria purified from the field9 in the laboratory. Under varying light intensities, the gas vesicle expansion pressure, mass density, intracellular carbohydrate mass, and protein content were tested. Evidently, the variations in the mass density of Microcystis were driven by light intensity. The main reason for the variations in mass density was the variation in intracellular carbohydrate mass in Microcystis cells as a result of photosynthesis and respiration processes. Visser et al.10 determined the relationship between the rates of density change in Microcystis and changes in photon irradiance in laboratory experiments, and provided critical information for simulations of the ‘day-sinking and night-floating’ (d-n) phenomenon of Microcystis cultures in still water.
Wind-driven currents changed the pattern of the d-n phenomenon of Microcystis in still water11. George et al.12 proposed a concept of critical wind speed, and only when the actual wind speed is below the critical wind speed, Microcystis can float to the surface of the water, and sink to the deeper layers vice versa13. Critical wind speed was observed in different parts of the world, such as Nakdong River14, Lake Dianchi15, Mickeysloe Bay16, Lake Kinneret17, Lake Taihu18. The critical wind speed was different in each in-situ investigation.
The size of colonies affects the ability of Microcystis to resist disturbance by wind-driven currents. Medrano et al.11 suggested that the larger the colony size of Microcystis, the more resistant it is to the disturbance of wind-driven currents, which indicates that large colonies can congregate at the water surface under increased wind-speed conditions. Wu et al.19 observed that Microcystis in Lake Taihu, with a colony size of 36–120 µm, were evenly distributed vertically under windy conditions and congregated at the surface when there was no wind. Laboratory test results from Xiao et al.20 and a numerical simulation test from Zhao et al.21 showed that the ability of Microcystis to resist the disturbance of wind-driven currents is positively correlated with colony size. Microcystis has a resistance limit for turbulent kinetic energy (TKE). When TKE generated by wind is greater than the resistance limit, Microcystis cannot easily float on the surface.
In summary, mass density of colonies, colony size, as well as wind-driven current play critical roles in the floating–sinking process of Microcystis, while the mass density is partially driven by light and the turbulence was partially driven by wind. The multiple effects of wind and light essentially affect the Microcystis floating–sinking process. Therefore, it is important to investigate this issue. Li et al.22 found that turbulent mixing may effectively reduce the colony size of Microcystis. Chien et al.23 considered the effects of light-driven mass density variations and Microcystis colony size on the floating and sinking of Microcystis under hydrostatic conditions and suggested that the d-n phenomenon only occurs in large Microcystis colonies. Medrano et al.24 integrated the effects of Microcystis colony buoyancy variations and turbulent disturbances and proposed that Microcystis would be evenly distributed vertically when the effects of turbulent disturbance dominated. Liu et al.25 studied the effects of wind-driven currents and Microcystis colony size on floating velocity. The vertical distribution of Microcystis in Milford Lake, USA was measured and simulated, and it was proposed that the mixing process caused by wind-driven currents had an important effect on the vertical distribution of Microcystis. Although these studies considered the combined effects of multiple factors on Microcystis floating/sinking, the motions were simulated based on diffusion–dispersion models, which did not incorporate the biological effects of wind disturbance and light on the mass density variations of colonies.
The mechanism of upward movement of Microcystis, considering both wind and light effects, needs to be clarified. Furthermore, the conditions that allow Microcystis to behave like a laboratory hydrostatic culture with the d-n phenomenon are unknown. Therefore, the movement patterns of Microcystis in lakes (i.e., Lake Taihu, China) remain unclear. This study investigated the influence of wind and light on the floating and sinking processes of Microcystis and studied the vertical motion of Microcystis colonies. To investigate the effects of wind and light on the floating and sinking movements of Microcystis, we assessed the influence of the intensity of wind-driven currents and changing mass density on floating and sinking movements.
Methods
Model overview
The intensity of wind-driven currents was simulated using Ansys Fluent, and compared with the in situ measurements. Disturbance of wind-driven currents, light-driven variations in mass density and the effect of colony size were coupled when simulating the vertical motion of Microcystis using MATLAB. We used SPSS 20 to analyze correlation of the data. Details about simulations can be seen in Appendix A (Supplementary).
In-situ investigation
Lake Taihu (30°55ʹ40ʺ–31°32ʹ58ʺ N; 119°52ʹ32ʺ–120°36ʹ10ʺ E) is located in the lower part of the Yangtze River Delta, China. It is a well-known large, shallow, and eutrophic lake. In 2017, we used an acoustic Doppler current profiler (ADCP) at two sampling sites in Meiliang Bay and Gonghu Bay in northern Lake Taihu to measure the stratified TKE in seven layers under different wind speeds throughout the year. Details about in-situ investigation and data analysis can be seen in Appendix A. Supplementary data.
Simulations
Simulation of the intensity of wind-driven currents
The wind-driven currents were simulated based on the multiphase flow Eulerian model in Ansys Fluent, which allows for the modeling of multiple separate, yet interacting phases. Momentum exchange between atmosphere and water is based on the value of the fluid–fluid exchange coefficient. The volume fraction follows the continuity equation. The turbulence intensity of wind-driven currents was simulated based on the ‘realizable k–ε model’26. To ensure the convergence and accuracy of the algorithm, the solution method in the SIMPLEC algorithm with pressure–velocity coupling and the discrete second-order upwind format was used.
Wind stress determines the efficiency of kinetic energy transfer between the atmosphere and water, and the morphology of surface waves. The strength of the wind stress strongly depends on the average wind speed at the water surface27. The wind stress in the wind-driven current model is simulated as follows28:
where \({F}_{x}\) and \({F}_{y}\) are wind stresses; \({\rho }_{air}\) is the air density; \({\rho }_{water}\) is the water density; \({C}_{d}\) is the drag coefficient of wind stress29; and \({u}_{wind}\) and \({v}_{wind}\) are the components of wind speed in the horizontal plane.
Details about simulations can be seen in Appendix A. Supplementary data.
Simulation of the influence of wind-driven currents on the Microcystis floating and sinking process
Movement of the Microcystis colony in turbulence can be considered analogous to the sediment transport process in turbulent currents21. The turbulent force, \({F}_{w}\), on the Microcystis population under wind-driven current disturbance is composed of turbulent drag, virtual mass, and pressure-gradient forces.
The upward direction is regarded as the forward direction; thus, the formula30 for the turbulent force, \({F}_{w}\), per unit mass of the Microcystis population is represented as follows:
where \(\rho\) and \({\rho }_{p}\) are the mass densities of water and Microcystis, respectively; \(v\) and \(u\) are the vertical velocities (Vy) of water and Microcystis colony, respectively; \({Re}_{d}\) is the Reynolds number of colonies, which is simulated as \({Re}_{d}=\frac{\rho {d}_{p}\left|v-u\right|}{\mu }\); \({C}_{D}\) is the drag coefficient of turbulent flow30, and \({d}_{p}\) is the Microcystis colony size.
To analyze the interaction between the Microcystis population and the continuously generated and dissipated eddies in the wind-driven current and to reflect the randomness of turbulent flow31, a random walk model was used to calculate the time step in this study.
Details` about simulations can be seen in Appendix A. Supplementary data.
Simulation of the influence of light on Microcystis floating and sinking
The effect of light on the floating and sinking of Microcystis colonies is mainly reflected in the light-driven variations in the mass density and thus in the population buoyancy. Visser et al.10 proposed a relationship between the variations in the mass density of Microcystis in still water and photon irradiance. In this study, the mass density under light-driven conditions was simulated based on the relationship between light intensity and mass density of Microcystis. The initial mass density32 of the Microcystis population was set to 985 kg m−3.
Simulation of the kinetic ratio for Microcystis floating and sinking
According to the force analysis, the effect of light can be expressed in terms of the mass density force, Fρ, and the effect of wind can be expressed in terms of the turbulent disturbance force, Fw. The ratio of these two forces is defined as the kinetic ratio, k, which determines the dominance of wind or light in the vertical movement of Microcystis. Equation (4) describes k as follows:
where \({f}_{w}\) is the turbulent flow disturbance force per unit mass of the Microcystis colony (Eq. 3); \({f}_{\rho }\) is the mass density force per unit mass of the Microcystis colony, \({f}_{\rho }=g\frac{\left(\rho -{\rho }_{p}\right)}{{\rho }_{p}}\); and \(\rho\) and \({\rho }_{p}\) are the mass densities of water and Microcystis colonies, respectively.
Details about simulations can be seen in Appendix A. Supplementary data.
Parameters
Parameter selection
-
1.
Wind speed
Diurnal wind speed in Lake Taihu from January 1, 1956 to September 30, 2019 was obtained from the China Meteorological Data Network (http://www.data.cma.cn). In the past 70 years, the average diurnal wind speed in the Lake Taihu area was 3.39 m s−1. From the diurnal wind speed frequency distribution, the typical wind speed was 0–5 m s−1, accounting for 93.3% of the measurements. Therefore, wind speeds of 1, 2, 3, 4, and 5 m s−1 and an extremely high value of 10 m s−1 were selected to cover the wide range of wind speeds for the simulation of wind-driven currents.
-
2.
Light intensity.
In still water, the light intensity varied at different depths. According to the Lambert–Beer law, the light intensity, I, at a water depth, y, is simulated as follows10:
$${{I}}_{{y}} ={{I}}_{{S}}/{{e}}^{{\eta y}}$$(5)$${{I}}_{{S}}={{I}}_{{MAX}}{{\sin}}{(}{\pi t}{/}{{D}}_{{L}}{)}$$(6)where η is the extinction coefficient, set at − 2 m−1; \({{I}}_{{MAX}}\) is the maximum light intensity at noon, set at 1000 μmol; and \({{D}}_{{L}}\) is the duration of light, set at 12 h.
-
3.
Microcystis colony size
The size and mass density of Microcystis colonies were determined from samples collected from Meiliang Bay and Gonghu Bay in the northern Lake Taihu. The average colony size of Microcystis in Lake Taihu was 342.7 µm33. In the study, values of 100, 300, 500, and 1000 µm were selected as four typical colony sizes of Microcystis in simulations, representing small to large Microcystis colonies. In each simulation, the size of Microcystis colony remains unchanged.
-
4.
Number of Microcystis colony
Considering the randomness of turbulence, the migration of 1000 colonies of the same size was simulated under the same wind speed. The colony whose final position was the median value of 1000 colonies was selected as the typical colony.
Computational domain
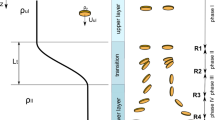
The two-dimensional vertical mathematical model in Ansys Fluent for the simulation of different strengths of wind-driven currents is shown in Fig. 1. A water depth of 2 m was used to emulate the actual water depth of Lake Taihu. The bottom height was set to 0 m. To guarantee the complete simulation of wind-driven currents, the fetch was set to 100 m. The strength of the wind field on the water surface was determined by the simulated wind speed that remained unchanged during the simulation. To weaken the influence of offshore flow on the intensity of the wind-driven current, we considered the average value recorded on five vertical lines as the final strength of the wind-driven currents. Vertical lines at 5-m intervals were established in the middle of the water body, and data were recorded (X = 40 m, 45 m, 50 m, 55 m, and 60 m). The average value was used to evaluate the effect of wind-driven currents on the floating–sinking of Microcystis.

Schematic diagram of the mathematical model for wind-driven current simulations.
In addition to wind speed, wind-driven currents are generally affected by fetch and duration27. To achieve practical results, the duration of the simulation was set to 48 h, and why? Due to the variance of wind-driven currents during the early stage, the simulated results of wind-driven currents in the 24–48 h period were used. The simulation time in following sections was counted from the 24 h. The effects of wind and light on the floating and sinking of Microcystis were considered in the model. To analyze the multiple effects of wind and light on the floating and sinking process of Microcystis, the vertical movement of different Microcystis colonies was simulated for 24 h under different intensities of wind-driven current disturbances and light intensity variations. The wind-driven current data for the 0–24 h period are represented as data from the 24 to 48 h period in the following sections. The light intensity was set to be the same as described in the section above, and the Microcystis colonies were initially placed on the water surface.
Results
Results of wind-driven current simulation
The distribution of velocity in the Y direction (Vy, Fig. 1) and of Turbulent kinetic energy (TKE) at 12 h from the start of the selected simulation and the comparison between the simulation and measured data at Lake Taihu (Fig. 2). The Vy of wind-driven currents oscillated within a certain frequency and had a conspicuous peak value (Fig. 2a). With an increase in wind speed, the oscillation ranges of Vy of wind-driven currents also increased. The magnitude of the wind-generated vertical flow velocity gradually decreases with an increase in water depth. Within 50 cm below the water surface, the oscillation range of the wind-generated vertical flow velocity was substantial, which changed rapidly. At depths between 50 and 200 cm below the water surface, the oscillation ranges of the wind-generated vertical flow velocity decreased. As the wind speed increased, the degree of turbulence in wind-driven currents increased simultaneously (Fig. 2b, Table 1). The wind-driven TKE gradually decreased with increasing water depth.

Simulated structure of wind-driven currents (a), relationship between depth and TKE (b), relationship between TKE and wind speed (c).
The average values measured from different layers in the water column were compared with the simulated mean TKEs of the entire water column of the wind-driven current at 12 h (Fig. 2c). The average TKE of the measured wind-driven currents in the entire water column was correlated with wind speed. The simulated results and measured data show a similar pattern.
Effect of wind on Microcystis floating and sinking
Disregarding the effect of light, the mass density of colony was set to a constant value of 985 kg m−332, and the effect of wind on the Microcystis floating and sinking process was simulated. Firstly, different sizes of colonies were placed on the water surface, and the vertical migration was simulated under the different intensities of wind-driven currents. Table 2 shows the median value and standard deviation of the final position of Microcystis colonies in the water column at the end of simulation. We found that with the increase of wind speed, the median value of the final position with the same size decreased after 1 h (0–1 h), and the final position with the smaller particle size was closer to the bottom. Besides, with the increase of wind speed, the standard deviation of the final position increases after 1 h, indicating a greater measure of dispersion. The migration trajectory of the typical colony was plotted in Fig. 3.

Vertical migration of Microcystis colonies at various constant wind speeds: (a) 1 m s−1, (b) 2 m s−1, (c) 3 m s−1, (d) 4 m s−1, (e) 5 m s−1, and (f) 10 m s−1.
The vertical variation patterns that were simulated by placing Microcystis colonies of different sizes at the water surface under different intensities of wind-driven current disturbance (Fig. 3). The wind speed remained unchanged under different simulation conditions. Two Microcystis movement patterns, Mode I and Mode II, were observed. Mode I resisted the disturbance of wind-driven currents allowing Microcystis to continue floating on the water surface, whereas Mode II could not resist the disturbance of wind currents; Microcystis sank and was turbulent in the water column. When the wind speed was extremely high (u = 10 m s−1), the colonies of all tested sizes were in Mode II. When the wind speed was the lowest (u = 1 m s−1), only the small colonies (d = 100 µm) sank (Mode II), and the remaining colonies were in Mode I. As the wind speed increased, the movement of smaller colonies changed from Mode I to Mode II, and they sank; larger colonies maintained their resistance to the wind-driven current disturbance.
The effect of variations in the intensity of wind field on the floating and sinking of Microcystis was not well understood. The behaviors of two groups of Microcystis were simulated using varying wind field strengths. In the first group, the Microcystis colonies were placed on the water surface, and the wind speed was increased from 1 to 10 m s−1 for simulating Microcystis sinking. In the second group, Microcystis colonies were placed at the water bottom, and the wind speed was decreased from 10 to 1 m s−1 to simulate Microcystis floating. Table 3 shows the median value and standard deviation of the final position of Microcystis colonies in the water column at the end of simulation. We found that with the increase of wind speed, the final position of colonies with the same particle size after 1 h was consistent with that under constant wind speed. The migration trajectory of the typical colony was plotted in Fig. 4.

Vertical migration of Microcystis colonies under varying wind speeds: (a) simulations in which colonies were place on the water surface and (b) simulations in which colonies were placed at the base of the water column.
Results indicate that the movement patterns of the Microcystis colonies under varying wind intensities were the same as those with constant wind (Fig. 4). Small colonies (d = 100 µm) maintained Mode II movements during the simulation. Even when the colonies were initially on the water surface (Fig. 5a), they exhibited sinking in the short term. Medium-sized colonies (d = 300 µm) could float on the water surface with Mode I movement, when the wind speed was low (u < 3 m s−1). When the wind speed was higher than 3 m s−1, the colonies could not resist the wind-driven current and sank into the water with Mode II movement. Colonies with large sizes (d = 500 and 1000 µm) were more resistant to wind-driven current disturbances and remained on the surface longer than in the other scenarios. When the wind speed was low, they came up to the water surface and exhibited Mode I movement. However, under extremely high wind speeds, the colonies sank into the water and exhibited Mode II movement.

Vertical migration of Microcystis colonies under varying light intensity.
Effect of light on Microcystis floating and sinking
The effect of variation in light intensity on the mass density of Microcystis colonies was included in the model. The light intensity was set according to the variation in light intensity within a day by setting 0–12 h as the time with light and 12–24 h as the time without light. Microcystis colonies were placed on the water surface at the beginning of the day, and the vertical movement of the Microcystis colonies with different colony sizes was simulated with the change of light intensity over 24 h.
The result shows that the variations in light intensity can affect the vertical movement of Microcystis (Fig. 5). In the first 12 h of light exposure, all Microcystis colonies started to sink after approximately 2 h of exposure, eventually reaching the bottom. In the first 12 h without light, all Microcystis colonies floated to the surface due to reduced mass density. All the Microcystis colonies showed the ‘day-sinking and night-floating’ (d-n) phenomenon when simulating with the variations in day and night light intensities. The floating and sinking speeds of large Microcystis colonies were faster than those of smaller colony sizes. When simulating the floating and sinking speeds of Microcystis colonies in still water using Stokes’ formula, the mass density driven by the variation of light intensity played a key role8.
Multiple effect of wind and light on the Microcystis floating and sinking process
Table 4 shows the median value and standard deviation of the final position of Microcystis colonies in the water column at the end of simulation. We found that with the increase of wind speed, the final position of colonies with the same particle size after 24 h was consistent with the law above. The migration trajectory of the typical colony was plotted in Fig. 6.

Vertical migration of Microcystis colonies under the effect of wind and light: (a) 1 m s−1, (b) 2 m s−1, (c) 3 m s−1, (d) 4 m s−1, (e) 5 m s−1, and (f) 10 m s−1.
The result shows that the smaller colonies with d = 100 μm did not show the ‘day-sinking and night-floating’ (d-n) phenomenon under the several given wind speed conditions, and the influence of wind was dominant (Fig. 6). When the wind speed was less than 3 m s−1, the medium-sized colonies (d = 300 μm) exhibited the d-n phenomenon, which is similar to the behavior under hydrostatic conditions. When the wind speed was higher than 3 m s−1, the phenomenon no longer occurred. The d-n phenomenon was still observed in colonies with large sizes (d = 500 μm and 1000 μm), when the wind speed was higher than 3 m s−1. However, when the wind speed was greater than 4 m s−1, no colonies showed the phenomenon. From the diurnal migration trajectory of Microcystis, as the wind speed increased, the position of Microcystis in the water column gradually approached the bottom.
Discussion
According to Eq. (4), when the kinetic ratio (k) > 1, the effect of wind dominates, and when k < 1, the effect of light becomes dominant. When the effect of wind dominated, the turbulent drag force dominated the vertical movement, and the more colonies were trapped in the water layer. When the effect of light dominated, the change in mass density drove the sinking and floating process of the colony, which led to the tendency of the ‘day-sinking and night-floating’ (d-n) phenomenon. Light had a greater effect on the floating and sinking of larger colonies while in comparison, smaller colonies were more affected by wind disturbance. Under the effects of wind and light, the average kinetic ratio of Microcystis with different colony sizes was shifted by wind-driven currents. By calculating the average kinetic ratio, we analyzed the dominant factors of Microcystis’ vertical movement, which was affected by intensities of light and wind. The variations of light intensity were observed to affect the mass density of Microcystis colonies. During the day, photosynthesis caused increasing mass density, and Microcystis colonies tend to sink. The average TKE at the corresponding location is shown in Fig. 7. Under the constant effect of wind-driven currents, the kinetic ratios of different colony sizes varied considerably. When the colony size was < 100 μm, the kinetic ratio was generally < 1, indicating that the wind played a dominant role. When the colony size reached 1000 μm or more, the effect of light could still play a dominant role, unless when the wind speed was greater than 5 m s−1. For each colony size, an equilibrium point was observed in which the effects of light and wind negated each other; a wind speed less than this point indicated light dominance, and vice versa, which is defined as ‘equilibrium TKE’.

Relationship between kinetic ratio and TKE: (a) 100 µm, (b) 300 µm, (c) 500 µm, (d) 1000 µm.
Microcystis colonies tend to float at night when the mass density decreases. The vertical velocity of wind-driven currents oscillates within a certain frequency and has a conspicuous peak value, the oscillation ranges of the vertical velocity of wind-driven currents also increases with the increase in wind speed. Hence, the water turbulence produces a drag force on the colonies, which acts in opposition to the direction of their movement. This means that wind-driven currents create an upward drag force when Microcystis sinks during the day, and a downward drag force is created when colonies rise at night. The magnitude of the drag force is related to the strength of the turbulence, and the ability of colonies to resist the drag force is highly dependent on the square of the colony size (Eq. 3), which also indicates that the smaller colonies are more dispersed in the water column under the same wind speed.
Realistically, a lake cannot be absolutely still. Microcystis populations generally begin to grow in spring and become smaller after autumn. The illumination time and light intensity above the water surface variations depend on the season and weather in Lake Taihu34. The TKE corresponding to normal colony sizes and wind speeds in Lake Taihu throughout the year are plotted in Fig. 8 and listed in Table 2. The relationship between Microcystis colony size and equilibrium TKE obtained from this study is shown as a solid line. Most Microcystis colonies can grow to 300–400 µm in April33. According to the actual wind speed statistics, the average wind speed in April was 3.6 m s−1, and the measured TKE strength was approximately 45 cm2 s-2, which was greater than the equilibrium TKE. Therefore, wind dominated the vertical movement of Microcystis. There was less time for the d-n phenomenon, and Microcystis was distributed in the water layer. From July to August, the colony size was approximately 500 µm35, the average wind speed was 3.4 m s−1, and the measured TKE was approximately 42 cm2 s−2. Although it was closer to equilibrium TKE, the wind-dominated period was longer. The daytime measured results36 showed that large colonies float easily on the water surface, whereas small colonies are mixed in the water layer. With wind speeds of 2–3 m s−1, this effect does not change owing to the light conditions. Although wind has a dominant effect most of the time, light can play a dominant role in the floating and sinking of Microcystis during relatively still periods. However, when the wind speed increases, the drag force of turbulent currents starts to dominate. Thus, it is difficult for light to alter the trajectory of Microcystis.

Relationship between colony sizes and equilibrium TKE.
The method proposed in this study, which combines the influence of wind-driven currents and the change in mass density of Microcystis on the migration of Microcystis, has broad applicability in the field of algal bloom prediction in shallow lakes in the future. For other shallow lakes, such as Lake Chaohu37 and Lake Dianchi15, the influence of wind-driven currents on the formation of water blooms has received more attention. This study provides an effective simulation method and provides theoretical guidance for this field. However, in deep lakes, such as Lake Erie38 and Lake Xiapu39, turbulence caused by temperature stratification cannot be ignored. Besides, the formation of algal blooms decreases the transparency of the water column and influences the vertical distribution of light intensity, which suggests a positive feedback regulation of Microcystis surface scum formation and stability by self-shading40. In our follow-up study, this mechanism will be further explored.
Conclusions
In conclusion, Microcystis tends to exhibit the ‘day-sinking and night-floating’ (d-n) phenomenon from the changing light intensities, however, wind currents generate turbulent drag forces that prevent the vertical movement of Microcystis and weaken this d-n phenomenon. Colonies with smaller sizes are less resistant to turbulence and are more dispersed in the water column. The existence of the d-n phenomenon can be determined comparing the kinetic ratio and the equilibrium TKE. When the TKE of the water body is greater than the equilibrium TKE, the d-n phenomenon does not occur for Microcystis. For Lake Taihu, the Microcystis colonies do not exhibit the d-n phenomenon, because the effect of wind dominates the vertical movement of Microcystis. In the absence of this phenomenon, the Microcystis colonies remain in the still water layer where the TKE is less than their equilibrium TKE. Our method highlights the use of a critical threshold of the kinetic ratio, which help simplify the numerical simulation and forecasting of Microcystis blooms.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request. Equipment and settings: All figures were created in Excel 2016.
References
Paerl, H. W. & Huisman, J. Climate. Blooms like it hot. Science 320, 57–58 (2008).
Yamamoto, Y., Shiah, F. K. & Chen, Y. L. Importance of large colony formation in bloom-forming cyanobacteria to dominate in eutrophic ponds. Ann. Limnol. Int. J Limnol. 47, 167–173 (2011).
Chen, Y. W., Qin, B. Q., Teubner, K. & Dokulil, M. T. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J. Plankton Res. 25, 445–453 (2003).
Walsby, A. E. The nuisance algae: Curiosities in the biology of planktonic blue-green algae. Water Treat. Exam. 19, 359–373 (1970).
Reynolds, C. S. & Walsby, A. E. Water-blooms. Biol. Rev. 50, 437–481 (1975).
Yonggang, L., Wei, Z., Ming, L. I., Amp, D. X. & Man, X. Effect of colony size on Microcystis diurnal vertical migration. J. Lake Sci. 25(3), 386–391 (2013).
Ibelings, B. W., Mur, L. & Walsby, A. Diurnal variations in buoyancy and vertical distribution in populations of Microcystis in two shallow lakes. J. Plankton Res. 13, 419–436 (1991).
Kromkamp, J. C. & Mur, L. R. Buoyant density variations in the cyanobacterium Microcystis aeruginosa due to variations in the cellular carbohydrate content. FEMS Microbiol. Lett. 1, 105–109 (1984).
Kromkamp, J. & Walsby, A. E. A computer model of buoyancy and vertical migration in cyanobacteria. J. Plankton Res. 12, 161–183 (1990).
Visser, P. M., Passarge, J. & Mur, L. R. Modelling vertical migration of the cyanobacterium Microcystis. Hydrobiologia 349(1–3), 99–109 (1997).
Medrano, E. A., Uittenbogaard, R. E., Pires, L. M. D., van de Wiel, B. J. H. & Clercx, H. J. H. Coupling hydrodynamics and buoyancy regulation in Microcystis aeruginosa for its vertical distribution in lakes. Ecol. Model. 248, 41–56 (2013).
George, D. G. & Edwards, R. W. The effect of wind on the distribution of chlorophyll A and crustacean plankton in a shallow eutrophic reservoir. J. Appl. Ecol. 13, 667 (1976).
Hutchinson, P. A. & Webster, I. T. On the distribution of blue-green algae in lakes: Wind-tunnel tank experiments. Limnol. Oceanogr. 9, 374–382 (1994).
Ha, K., Kim, H. W., Jeong, K. S. & Joo, G. J. Vertical distribution of Microcystis population in the regulated Nakdong River, Korea. J. Limnol. 1, 225–230 (2000).
Ma, X., Wang, Y., Feng, S. & Wang, S. Vertical migration patterns of different phytoplankton species during a summer bloom in Dianchi Lake, China. Environ. Earth Sci. 74, 3805–3814 (2015).
Ndong, M. et al. A novel Eulerian approach for modelling cyanobacteria movement: Thin layer formation and recurrent risk to drinking water intakes. Water Res. 127, 191–203 (2017).
Hozumi, A., Ostrovsky, I. S., Sukenik, A. & Gildor, H. Turbulence regulation of Microcystis surface scum formation and dispersion during a cyanobacteria bloom event. Inland Waters. 10, 51–70 (2020).
Zhu, W., Chen, H., Xiao, M., Miquel, L. & Li, M. Wind induced turbulence caused colony disaggregation and morphological variations in the cyanobacterium Microcystis. J. Lake Sci. 33, 349 (2021).
Wu, X. & Kong, F. Effects of light and wind speed on the vertical distribution of Microcystis aeruginosa colonies of different sizes during a summer bloom. Int. Rev. Hydrobiol. 94, 258–266 (2009).
Xiao, M. et al. The influence of water oscillation on the vertical distribution of Microcystis colonies of different sizes. Fresenius Environ. Bull. 22, 3511–3518 (2013).
Zhao, H. et al. Numerical simulation of the vertical migration of Microcystis (cyanobacteria) colonies based on turbulence drag. J. Limnol. 76, 190–198 (2017).
Li, M., Xiao, M., Zhang, P. & Hamilton, D. P. Morphospecies-dependent disaggregation of colonies of the cyanobacterium Microcystis under high turbulent mixing. Water Res. 141, 340–348 (2018).
Chien, Y. C., Wu, S. C., Chen, W. C. & Chou, C. C. Model simulation of diurnal vertical migration patterns of different-sized colonies of Microcystis employing a particle trajectory approach. Environ. Eng. Sci. 30, 179–186 (2013).
Medrano, E. A., van de Wiel, B. J. H., Uittenbogaard, R. E., Pires, L. M. D. & Clercx, H. J. H. Simulations of the diurnal migration of Microcystis aeruginosa based on a scaling model for physical-biological interactions. Ecol. Model. 337, 200–210 (2016).
Liu, H., Zheng, Z. C., Young, B. & Harris, T. D. Three-dimensional numerical modeling of the cyanobacterium Microcystis transport and its population dynamics in a large freshwater reservoir. Ecol. Model. 398, 20–34 (2019).
Shih, T. H., Liou, W. W., Shabbir, A., Yang, Z. & Zhu, J. A new k-ε eddy viscosity model for high Reynolds number turbulent flows. Comput. Fluids. 24, 227–238 (1995).
Geernaert, G. L., Larsen, S. E. & Hansen, F. Measurements of the wind stress, heat flux, and turbulence intensity during storm conditions over the North Sea. J. Geophys. Res. 92, 127–139 (1987).
Large, W. G. & Pond, S. Open ocean momentum flux measurements in moderate to strong winds. J. Phys. Oceanogr. 11, 324–336 (1981).
Sellers, H. Development and application of “U.S.E.D.”: A hydroclimate lake stratification model. Ecol. Model. 21, 233–246 (1984).
Morsi, S. A. & Alexander, A. J. An investigation of particle trajectories in two-phase flow systems. J. Fluid Mech. 55, 193–208 (1972).
Gosman, A. D. & Loannides, E. Aspects of computer simulation of liquid-fuelled combustor. AIAA J. 81, 482–490 (1981).
Li, M. et al. To increase size or decrease density? Different Microcystis species has different choice to form blooms. Sci. Rep. 6, 37056 (2016).
Li, M., Zhu, W. & Gao, L. Analysis of cell concentration, volume concentration, and colony size of Microcystis via laser particle analyzer. Environ. Manag. 53, 947–958 (2014).
Sun, D., Li, Y., Wang, Q. & Gao, J. Light scattering properties and their relation to the biogeochemical composition of turbid productive waters: A case study of Lake Taihu. Appl. Opt. 48(11), 1979–1989 (2009).
Li, M., Zhu, W., Gao, L., Huang, J. & Li, L. Seasonal variations of morphospecies composition and colony size of Microcystis in a shallow hypertrophic lake (Lake Taihu, China). Fresenius Environ. Bull. 22, 3474–3483 (2013).
Zhu, W. et al. Vertical distribution of Microcystis colony size in Lake Taihu: Its role in algal blooms. J. Great Lakes Res. 40, 949–955 (2014).
Chen, Y. Y. & Liu, Q. Q. On the horizontal distribution of algal-bloom in Chaohu Lake and its formation process. Acta Mech. Sinica-Prc. 30(005), 656–666 (2014).
Beletsky, D., Hawley, N., Rao, Y. R., Vanderploeg, H. A. & Ruberg, S. A. Summer thermal structure and anticyclonic circulation of Lake Erie. Geophys. Res. Lett. 39, 6605 (2012).
Ishikawa, T. & Qian, X. Numerical simulation of wind-induced current and water exchange at the mouth of Takahamairi Bay of the Lake Kasumigaura during the formation of diurnal thermocline. Tohoku Univ. 2, 419–428 (1998).
Wu, H., Wu, X. & Yang, T. Feedback regulation of surface scum formation and persistence by self-shading of Microcystis colonies: Numerical simulations and laboratory experiments. Water Res. 194(3), 116908 (2021).
Acknowledgements
We are grateful for the grants for this study from the Science and Technology Project of Jiangsu Province [BE2018737] and Research and Innovation Plan for Postgraduates in Jiangsu Province [B200203049]. We would like to thank Elsevier (https://cn.webshop.elsevier.com) for English language editing.
Funding
This work was funded by a project supported by the Science and Technology Project of Jiangsu Province [BE2018737] and Research and Innovation Plan for Postgraduates in Jiangsu Province [B200203049].
Author information
Authors and Affiliations
Contributions
Z.X. performed the simulations and significantly contributed to drafting the manuscript. W.Z. analyzed the influence of wind and light on the floating and sinking processes of Microcystis. Y.Z. drafted the manuscript. X.F. participated in the simulations. H.C. and G.F. analyzed the Lake Taihu field data. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xue, Z., Zhu, W., Zhu, Y. et al. Influence of wind and light on the floating and sinking process of Microcystis. Sci Rep 12, 5655 (2022). https://doi.org/10.1038/s41598-022-08977-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-08977-5
This article is cited by
-
Wind-driven hydrodynamic characteristics of Lake Taihu, a large shallow lake in China
Environmental Science and Pollution Research (2024)
-
The Vertical Migration Dynamic of Microcystis aeruginosa in a Water Column
Journal of Applied Phycology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
