
Abstract
Light is one of the most important limiting factors for photosynthesis and the production of plants, especially in the regions where natural environmental conditions do not provide sufficient sunlight, and there is a great dependence on artificial lighting to grow plants and produce food. The influence of light intensity, quality, and photoperiod on photosynthetic pigments content and some biochemical and growth traits of cucumber seedlings grown under controlled conditions was investigated. An orthogonal design based on a combination of different light irradiances, ratio of LEDs and photoperiods was used. Treaments consisted of three light irradiance regimes (80, 100, and 150 µmol m−2 s−1) provided by light-emitting diodes (LEDs) of different ratios of red and blue (R:B) (30:70, 50:50, and 70:30) and three different photoperiods (10/14, 12/12, and 14/10 h). The white light was used as a control/reference. Plant height, hypocotyl length, stem diameter, leaf area, and soluble sugar content were highest when exposed to LM9 (150 µmol m−2 s−1; R70:B30; 12/12 h) light mode, while the lowest values for the above parameters were obtained under LM1 (80 µmol m−2 s−1; R30:B70; 10/14 h). Higher pigments contents (chlorophyll a, chlorophyll b, and carotenoid) were obtained when light regime LM9 (150 µmol m−2 s−1; R70:B30; 12/12 h) was applied. In general, cucumber seedlings grown under the LM9 regime showed a significant increase in growth as well as photosynthetic capacity. It seems that the content of photosynthetic pigments is the key factor responsible for the performance of cucumber seedlings grown under different lighting modes, compared to other traits studied. We recommend monitoring the content of chlorophyll a, b, and their ratio value when studying the light requirement of cucumber plants.
Similar content being viewed by others

Introduction
The cucumber is an economically important vegetable grown in over 80 countries worldwide1, and its annual production is estimated to be about 80 million tons, including about 3 million tons grown in the European Union (EU 28)2. High temperatures, humidity, light irradiance, and nutrient availability are ideal conditions for this typical subtropical plant, which is highly sensitive to adverse environmental conditions3. Characterized by their tenderness, these plants thrive in a temperature range between 18.3 to 23.9 °C, with a minimum temperature of 15.6 °C and a maximum of 32.2 °C. Germination of the cucumber takes place in soil that has a temperature range from 15.6 to 35 °C. However, germination is substantially impeded below 15.6 °C4.
The energy given by light is a key concern in cucumber cultivation, as it is in other plant development, and temperature regulation must be addressed in conjunction with light irradiance5. The total plant leaf area, carbohydrate production, and, consequently, productivity are all affected by radiation6. During the winter months, low carbohydrates supply and reduced output may lead many plants to fail7. As a result, the quality of the vegetable is directly influenced by light. The crops cultivated in such low light levels have less dry matter, and the colour is green when harvested but quickly becomes yellow when stored. The sensitivity of these young vegetables to low light irradiance is greater than that of the older vegetables from the same plant3.
The use of artificial lighting suitable for indoor cultivation under controlled environments has increased plant productivity in densely populated areas and space missions8. The spectral characteristics of electric light sources must meet the physiological requirements of plants for photosynthesis and photomorphogenic development9. However, the distribution and variation of the spectrum of conventional light sources (fluorescent tubes, sodium vapor lamps, metal halides etc.) is fixed and may not be ideal for the light requirements of different plant species. Light-emitting diodes (LEDs) are on the rise and have great potential for horticultural lighting due to their energy efficiency, longevity, and flexibility of use10. In addition, LEDs are becoming more suitable for research and commercial agriculture under controlled conditions due to their low radiation, heat, and broad spectral match11.
Brown et al.12 and Tennessen et al.13 reported that plant light demand research can determine the various functions of light properties in terms of spectral irradiance when growing conditions have been supplied with LEDs and modified to emit light photons at specific wavelengths. It is worth noting that LED provides the ideal spectral distribution range that promotes plant growth with optimal longevity and light energy efficiency14. During the shift from conventional to LED light sources, many inventions have been made, such as the combination of fluorescent lamps and LED15, the replacement of fluorescent tubes with LEDs16, and the retrofitting of LEDs for fluorescent tubes without ballasts17.
Many works have attempted to explain the influence of light on growth, development, morphology, and photosynthesis in various plants18,19,20,21. The combination of LED light quality has been observed to strongly influence the physiological and developmental processes of plants22,23,24,25,26.
Many scholars have reported different accounts of plants grown under different illumination irradiances, providing insights into plant growth and development as well as their photoperiodic requirements25,27,28,29,30.
The aim of this study was to determine the best light irradiance, light quality, and photoperiod for the growth of cucumber (Cucumis sativus var. Building No. 4) seedlings and to find out which physiological characteristic is the best as an indicator of optimal lighting. This experiment was designed on the hypothesis that the response of the photosynthetic apparatus of cucumber seedlings and their growth parameters will be varied base on the characteristics of the applied light modes.
Results
Growth parameters
The effect of light type LED on plant morphology and growth characteristics of cucumber seedlings is shown in Fig. 1. Different light conditions had significant effects on the morphological characteristics of cucumber seedlings. Plant height, stem diameter, total leaf area, hypocotyl length, shoot fresh weight, root fresh weight, shoot dry weight and root dry weight were significantly higher under LM9 treatment than the other treatments, while the lowest value for all these parameters was observed under LM1. Plant height, stem diameter, total leaf area, hypocotyl length, shoot fresh weight, root fresh weight, shoot dry weight and root dry weight were increased by 85.07, 52.73, 57.37, 172.81, 77.94, 98.88, 133.33 and 62.5% when LM9 was applied on cucumber seedlings (Fig. 1A–H) as compared to control (WL) while application of LM8 increased these attributes by 79.1, 47.27, 40.16, 149.19, 67.64, 90, 111 and 46.59% respectively as compared to control (WL). The water content of plants exposed to LM9 (92.4%) was significantly higher than under the other LED light modes, although the values were not significantly different between LM1–LM6, LM8, and WL (Fig. 1I).

Effect of modes LED light on plant morphology and growth characteristics of cucumber seedlings; where Plant height (A), Stem diameter (B), Total leaf area (C), Hypocotyl length (D), Shoot Fresh weight (E), Root Fresh weight (F), Shoot dry weight (G), Root Dry weight (H), Water content % (I). Means followed by the same letter within the same series are not significantly different according to Duncan’s multiple range test (P ≤ 0.05). See Table 4 for LM abbreviations.
On the other hand, according to the R-values, the order of influence of the three factors on growth characteristics of cucumber seedlings was observed in this study by using the orthogonal array design (Table 1). Table 1 shows that the order of impact of the three factors on plant height, stem diameter, leaf area, Hypocotyl length, shoot fresh weight, root fresh weight, shoot dry weight, root dry weight, and water content was (A > B > C), (A > B > C), (A > B > C), (A > B > C), (A > B > C), (A > B > C), (A > B > C), (A > C > B), and (C > B > A), respectively.
Based on the average of growth characteristics derived from three factors at each level, the best combination of different factors with the levels to get the highest results on growth performance was A3B3C2, which indicated that the maximum of these parameters presented at the irradiance of light (150 µmol m−2 s−1), the ratio of (R70:B30), and photoperiod (12/12 h).
ANOVA (Table 1) showed that these three factors were significant effects on growth performance parameters of cucumber seedlings (p ˂ 0.05), excepted factor C on plant height, hypocotyl length, shoot fresh weight, root fresh weight, and shoot dry weight had no significant effects, and also excepted factor B on root fresh weight and root dry weight had no significant effects.
Photosynthetic pigments content
The contents of chlorophyll and carotenoids in the leaves of cucumber seedlings under different LED light modes is shown in Fig. 2. Compared with WL treatment, the levels of Chl a, Chl b, total chlorophyll and carotenoid in the leaves of cucumber seedlings were higher with LM9 than with the other modes of LED light, with LM7 and LM8 showing no statistical difference, while LM1 mode showed the lowest levels of Chl a, Chl b and carotenoid (Fig. 2A–D). The ratio of carotenoid to total chlorophyll was higher with LM3 than the other modes of LED light, with LM4, LM5 and WL showing no statistical difference between the values, while LM1 mode showed the lowest ratio (Fig. 2E).

Effect of modes LED light on photosynthetic pigments content [chlorophyll a (A), chlorophyll b (B), total chlorophyll (C), carotenoid (D), and total chlorophyll/carotenoid contents (E)] in cucumber seedlings. Means followed by the same letter within the same series are not significantly different according to Duncan’s multiple range test (P ≤ 0.05). See Table 4 for LM abbreviations.
On the other hand, according to the R-values, the order of influence of the three factors on photosynthetic pigments content of cucumber seedlings was observed in this study (Table 2). Table 2 shows that the order of impact of the three factors on chlorophyll a, chlorophyll b, total chlorophyll, carotenoid, and total chlorophyll/carotenoid was (A > B > C), (A > B > C), (A > B > C), (A > B > C), and (C > A > B), respectively.
Based on the average of photosynthetic pigments content derived from three factors at each level, the A3B3C2 was the best combinations gave the highest chlorophyll a, chlorophyll b, total chlorophyll, and carotenoid, which indicated that the maximum of these parameters presented at (irradiance 150 µmol m−2 s−1 + ratio (R70:B30) + photoperiod 12/12 h). While the best combination of different factors with the levels for the highest total chlorophyll/carotenoid was A1B3C3, which indicated that the maximum of these parameters presented with (irradiance 80 µmol m−2 s−1 + ratio (R70:B30) + photoperiod 14/10 h).
ANOVA (Table 2) showed that these three factors were significant effects on photosynthetic pigments content of cucumber seedlings (p ˂ 0.05), excepted factor B on chlorophyll b, and factor C on chlorophyll b and Carotenoid had no significant effects.
Biochemical traits
Light modes of LEDs had significant effects on the accumulation of biochemical compounds, including the contents of nitrate, soluble protein, and soluble sugar (Fig. 3). LM3 provided the highest nitrate and soluble protein contents; this shows the importance of red light in improving nitrate and soluble protein content (Fig. 3A,B). Also, LM9 exhibited the highest accumulation of soluble sugar in the cucumber seedlings (6.40%FW) (Fig. 3C).

Effect of modes LED light on biochemical traits; Nitrate content (A), Soluble protein content (B), and Soluble sugar content (C) in cucumber seedlings. Means followed by the same letter within the same series are not significantly different according to Duncan’s multiple range test (P ≤ 0.05). See Table 4 for LM abbreviations.
On the other hand, according to the R-values, the order of influence of the three factors on biochemical traits of cucumber seedlings was observed in this study (Table 2). Table 2 shows that the order of impact of the three factors on nitrate content, soluble protein, and soluble sugar contents was (A > B > C) with all of them.
Based on the average of biochemical traits derived from three factors at each level, the best combination of different factors with the levels for the highest nitrate content and soluble protein was A1B3C3, which indicated that the maximum of these parameters presented at (irradiance 80 µmol m−2 s−1 + ratio (R70:B30) + photoperiod 14/10 h). While the best combination of different factors with the levels for the highest soluble sugar content was A3B3C2, which indicated that the maximum of soluble sugar content presented with (irradiance 150 µmol m−2 s−1 + ratio (R70:B30) + photoperiod12/12 h).
ANOVA (Table 2) showed that these three factors were significant effects on biochemical traits of cucumber seedlings (p ˂ 0.05), excepted factors C on soluble sugar content had no significant effects.
Correlation analysis
Pearson’s correlation31 was carried out among the morphological, photosynthetic pigments content, and biochemical traits observed in this study as shown in Table 3. There was a highly significant positive correlation between PH with [(SD R2 = 0.935), (TLA R2 = 0.965), (HL R2 = 0.983), (SFW R2 = 0.977), (RFW R2 = 0.970), (SDW R2 = 0.990), (RDW R2 = 0.949), (Chl a R2 = 0.977), (Chl b R2 = 0.969), (ToCh R2 = 0.916), (Car. R2 = 0.975), and (Sug. R2 = 0.972)]. While the correlation relation between PH with (WC%, ToCh/Car., Nit., and Pro.) were found significantly negative (R2 = − 0.866, − 0.458, − 0.753, and − 0.734, respectively). Additionally, a positive significant relationship was found between SD and [(TLA R2 = 0.983), (HL R2 = 0.937), (SFW R2 = 0.963), (RFW R2 = 0.956), (SDW R2 = 0.951), (RDW R2 = 0.984), (Chl a R2 = 0.966), (Chl b R2 = 0.986), (ToCh R2 = 0.943), (Car. R2 = 0.962), and (Sug. R2 = 0.981)]. Whereas the negative correlations were found between SD and WC %, Nit., and Pro. (R2 = − 0.953, − 0.814, and − 0.846, respectively). The relationship between Nit. and Pro. with all morphological traits was high significantly negative, while the relationship between Sug. and all morphological traits was high significantly positive as shown in Table 3.
Discussion
For proper growth and development, plants are grown under constantly changing light conditions. Some light wavelengths are critical for plant growth and development. Plants are observed to detect subtle changes in light quality by light receptors. These light receptors can initiate signal transduction through various pathways to alter plant appearance32,33,34. It has been observed that photosynthetically active radiation (PAR; 400–700 nm) plays a direct role in photosynthetic processes of plants. Red light in the range of 610–700 nm and blue at 425–490 nm is the optimal light spectrum for photosynthesis in plants35. Recently, there are many research results that focused on the effects of LED light on morphogenesis and photosynthesis. They observed that red and blue light improved plant production when light irradiance and quality were controlled36,37,38.
Growth parameters
The photosynthetic process in leaves requires the capture of light, which is influenced by the wavelength (light spectrum), intensity and angle of incidence39, and total leaf area. It was observed that cucumber seedlings responded strongly to LM9 in terms of plant height, stem diameter, total leaf area, hypocotyl length, shoot fresh weight, root fresh weight, shoot dry weight, and root dry weight (Fig. 1). This finding agreed with the results of Naznin et al.37 and Yang et al.40 in pepper seedlings and Yang et al.41 in tomato seedlings, they observed that a mixture of red and blue LED light was efficient in producing strong seedlings. The combination of red and blue light was shown to be the most beneficial in promoting plant growth and development in the majority of research. When cucumber seedlings were grown under a combination of red and blue light (R5: B5), yields were higher than when they were grown under red light only42. Under a combination of blue and red light, Kim and Hwang confirmed that high grade 'Mini Chal' tomato (Solanum lycopersicum L.) could be grown in plant factories43. Furthermore, the barrier tissue cells in the leaves were very well developed and the spongy tissue cells were organized in an ordered manner under red + blue light44.
Combining red and blue light is more effective for plant development than monochromatic red light. Phytochrome-dependent elongation of hypocotyls and cotyledons was seen in plants cultivated under monochromatic red light45. Plants exposed to a mixture of red and blue light had greater photoreceptor excitation, like phytochromes, cryptochromes, and phototropins, and had higher photosynthetic activity than plants exposed to monochromatic red or blue light46. According to Yousef and coauthors showed that a combination of R and B LED light with a high R portion was effective in producing vigorous grafted tomato seedlings compared to blue and red light alone20,47,48. Also, Dong et al.49, as compared to blue light alone, red light alone, and sunshine, combined red and blue light (1/3 blue light at 450–460 nm + 2/3 red light at 620–630 nm, at 400 lx and 12 h photoperiod for 60 days) improved the DW and bio efficiency of Cordyceps militaris mushroom. Plant production was considerably improved for most species when combined light wavelengths with a considerable proportion of red light supplemented by blue light were used50.
Effects of mode (regime) on photosynthetic pigments content
Chlorophyll content directly affects photosynthetic ability and primary production51. Moreover, the chlorophyll content of plants is affected by the quality of light. Many studies have explained the beneficial effects of using blue lights21,38,41. Our results showed that the combination of red and blue LED light with high red light (LM9) was observed to be favorable for chlorophyll a, chlorophyll b, and carotenoids content (Fig. 2). This is in agreement with the findings of Yang et al.41 on tomato seedlings and on pepper seedlings37, they have observed that the content of these pigments was more when seedlings were exposed to a mixture of red and blue LED lights than white fluorescent lights exposure.
Chlorophyll content, photosynthetic enzyme activity, stomatal aperture and carbohydrate release in plants were all affected by red and/or blue light52. Because of the high amount of carbohydrates in the leaves, red light increased the total chlorophyll content in plants, which encouraged photosynthesis. However, it prevented the movement of carbohydrates from the leaves to enhance photosynthesis, suppressing photosynthesis52. By raising the ratio of chl a/b, enhancing the activities of ribulose-1,5-bisphosphate carboxylase (Rubisco) and phosphoenolpyruvate carboxylase, and encouraging stomatal opening, blue light improved photosynthesis per unit leaf area53. Plant yield was affected by red and/or blue light during morphogenesis. Cell division and expansion were aided by red light, resulting in increased leaf area and root elongation, whereas cell division and expansion were hindered by blue light, resulting in reduced leaf area and root elongation54. Plant photomorphogenesis was affected by blue light, which increased chlorophyll a/b ratios and facilitated stomatal opening55. Reduced photon uptake due to reduced leaf area could be the cause of reduced plant growth under LM1.
Effect of light mode (regime) on chosen biochemical traits
The study presented the significance of the use of red light in metabolite accumulation in cucumber seedlings. LM3 was more effective in increasing nitrate and soluble sugar content in cucumber seedlings while WL was observed to increase soluble protein content. The study by Bian et al.56 proved that soluble sugar in lettuce was higher when irradiated with red, green, and blue LED light (4:1:1) than under other types of LED light. Also, Xiaoying et al.18 observed that tomato seedlings grown under blue LED light had higher soluble sugar levels than under other types of LED light57, they also observed that soluble sugar levels were higher than other types of LED light in pepper, tomato and cucumber seedlings grown under red LED light and (red:blue). Their results proved that soluble sugars and proteins respond to different light qualities in vegetable crops grown under controlled conditions and that this also varies among different species and cultivars. In the present study, it was found that at a light irradiance of (80 ± 2 μmol m−2 s−1) and 14/10 h photoperiods, a combination of R70:B30 LED light was more effective in reducing nitrate concentration for cucumber seedlings than the white fluorescent light. Bian et al.56 reported that the effect of red and blue light had negative effects on nitrate assimilation by decreasing the activity and expression of nitrate assimilation-related genes NR and NiR in hydroponically grown lettuce, while the addition of green light with red and blue light had positive effects on nitrate assimilation by increasing activity.
The correlation analysis performed showed that most of the tested parameters were significantly and positively correlated. Only traits such as water content %, nitrate, and protein content were negatively associated. Total chlorophyll/carotenoid ratio and did not show significant relationship with the other traits studied (Fig. 4).

Radar plot of all studied traits under various light modes. Where PH Plant height, SD Stem diameter, TLA Total leave area, HL Hypocotyl length, SFW Shoot fresh weight, RFW Root fresh weight, SDW Shoot dry weight, RDW Root dry weight, WC % Water content, Chl a Chlorophyll a, Chl b Chlorophyll b, ToCh Total Chlorophyll contents, Car. Carotenoid, ToCh/Car. Total Chlorophyll contents/Carotenoid, Nit. Nitrate content, Pro. Protein content, and Sug. Sugar content.
Materials and methods
Growth conditions and plant materials
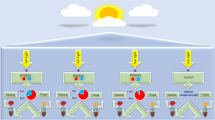
The experimental system included 10 chambers; each had a dimension of 60 × 60 × 60 cm. The details of growth conditions and LEDs light are shown in Table 4 and Fig. 5. The manufacturer of the tested LED lamps is Kedao Technology Corporation (Huizhou, China) with the type of UH-BLDT0510. The plant experiments were complied with local and national regulations and following Fujian Agriculture and Forestry University (Fujian, China) regulations. Cucumber seeds (Cucumis sativus var. Built No. 4) were provided by the Tianjin Kernel Cucumber Research Institute. Cucumber seeds were sown in 32-cell plug trays (W 4 cm × L 4 cm × H 6 cm/cell) that was filled with commercial growing substrate (N1: P1: K1 ≥ 3%, Organic matter ≥ 45% pH 5.5–6.5). Ten days after planting, the germinated seedlings were transferred to pots (W 10 cm × L 10 cm × H 8.5 cm) and were left there for 20 days. In total, 20 seedlings were sown in each growth chamber. Irrigation was provided for the seedlings daily or as required. Seedlings began receiving fertilization based on water-soluble nutrients two times per week through irrigation one week after sowing. The air conditioner and ventilation fans were relied on in the chambers to standardize temperatures as well as possible.

Spectrum distribution of the treatments LED light and in the experiment and environmental conditions.
Multiple-factor experiment design
The multiple-factor experimental regular fractional design L9 (33) was used in this experiment (Table 4) i.e. 3 levels were chosen for each of the 3 improvement criteria and 9 tests from all possible combinations. When considering the technical feasibility of the advanced LED lighting unit, the parameters for improving the lighting system in the factory were chosen at the following levels:
-
A.
The irradiances of LEDs light averaged over the whole time of plant growing period, PPFD (A1-A3): 80, 100, 150 μmol m−2 s−1.
-
B.
The ratio of PPDD values from Red and Blue LEDs (B1-B3): (R:B) = 30:70, 50:50, and 70:30.
-
C.
The light period during Light/Dark (C1–C3): 10/14 h, 12/12 h, and 14/10 h.
Additionally, a white fluorescent lamp (WL) was used as a control.
Measurements and calculations
Measurement of growth and biomass parameters
On the 30th day after planting, growth metrics were measured. Using a ruler, the height of the plant was measured from the base of the rhizome to the top of the plant (cm). Digital calipers (mm) were used to measure stem diameter, and an electronic balance was used to weigh the fresh and dry mass (0.0001 g). Pandey and Singh's58 method for calculating total leaf area (cm2) was used. To acquire the dry weight, fresh shoots and roots were placed in Petri plates without cover and placed in a drying oven at 75 °C for at least 48 h.
Measurement of photosynthetic pigments
After 30 days of transplanting chlorophyll and carotenoid contents were determined from fresh medium-aged leaves with excluded the edges and veins of leaves. Tissues of fresh leaves (0.2 g) were cut, ground well then used in 5 mL of 95% ethanol and filtered, the filtrate was made up to 25 mL by adding 95% ethanol. Absorbance of the filtered solution at 665 nm (OD665), 649 nm (OD649) and 470 nm (OD470) was measured using a spectrophotometer (UV-5100B, Unico. Shanghai, China) while the chlorophyll content was determined using the equations below59: Chl a (mg g−1FW) = (13.95OD665 − 6.88OD649) V/200 W; Chl b (mg g−1FW) = (24.96OD649 − 7.32OD663) V/200 W; Total Chlorophyll (mg g−1FW) = Chl a + Chl b; C (mg g−1FW) = (1000OD470 − 2.05Chl a − 114.80Chl b) V/(245 × 200 W). Where (Chl a) = chlorophyll a, (Chl b) = chlorophyll b, (C) = carotenoid; (V) = volume (25 mL) and (W) = sample weight (g).
Measurement of biochemical traits
To measure the biochemical traits, fresh leaves were chopped into small pieces and fresh samples weighed (0.5 g, 0.2 g, and 0.5 g) for nitrate, protein, and sugar, respectively; they were used to determine the content of soluble nitrate60. The soluble protein content was evaluated using coomassie brilliant blue G250 method61. Also, the content of soluble sugar was evaluated using the anthrone colorimetric method62. The absorbance of the solution extracted was estimated at 410 nm (OD410), 595 nm (OD595), and 630 nm (OD630) using a UV-5100B spectrophotometer (Unico, Shanghai, China). The biochemical traits were expressed using the following equations: Soluble nitrate content (mg kg−1FW) = (C × VT)/(W × VS); Soluble protein content (mg g−1FW) = (C × VT)/(VS × W × 1000); Soluble sugar content (%) = (C/VS × VT)/(W × 106) × 100. Where C = nitrite; sugar (%) ; protein value from the standard curve, VT = total volume of samples extracted (mL), VS = taken sample solution (mL), and W = leaf fresh weight (g).
Statistical analysis
All above mentioned measurements were made with 9 replicates. The Orthogonal Experimental design method was used to determine the number of experiments to be conducted. All the data were subjected to an one-way analysis of variance (ANOVA). Duncan's multiple range tests63 was used to test the significant difference between the means at 0.05 significance level using SPSS software (Version 16 SPSS Inc. Chicago, Illinois). The importance of the three factors for the measured parameters was assessed according to the effectiveness of each factor64 by the range value (R) using Excel 365 (v16.0). The most important impact factor has the greatest R-value. Correlation analysis was done, and Pearson correlation coefficients are shown65.
Conclusions
Based on our experimental work and the data obtained, we propose the following mechanism or "cascade of effects": the light mode acts first on photosynthetic pigments. This, in turn, increases the photosynthetic output of plants and then the soluble sugar content, which could indicate a higher production of proteasomal. The latter were directly used by plants to build up the shoot and root biomass and increase the photosynthetic area of plants.
In summary, we believe that light irradiance is more important (has a greater effect) than light ratio and photoperiod in the case of cucumber seedlings (Fig. 4). This is evident in the case of LEDs of 150 μmol m−2 s−1 (LM 7, 8 and 9), where in general the highest values of the most studied traits/properties were observed, especially the content of photosynthetic pigments. The spectral light ratio (red:blue) proved to be the second factor affecting the studied properties. A higher red:blue ratio (70:30) was the best. Finally, it looks that the longest light period (10–14 h per day) did not play a significant role in establishing better photosynthetic performance and growth of cucumber seedlings. Therefore, it is necessary to perform molecular analyses and link them to morphological and biochemical traits to learn more about the mechanisms of the effect of LED light on seedling growth.
Data availability
All data available within the article.
References
Adhikari, B. N. et al. Expression profiling of Cucumis sativus in response to infection by Pseudoperonospora cubensis. PLoS One 7, e34954 (2012).
FAOSTAT—Crops, F. http://www.fao.org/faostat/en/#data (2020).
Haifa. Crop Guide: Growing Cucumbers. https://www.haifa-group.com/cucumber-fertilizer/crop-guide-growing-cucumbers (2018).
Schrader, W., Aguiar, J. & Mayberry, K. Cucumber Production in California (UCANR Publications, 2002).
An, S. et al. Evaluation of air temperature, photoperiod and light intensity conditions to produce cucumber scions and rootstocks in a plant factory with artificial lighting. Horticulturae 7, 102 (2021).
Baudoin, W. et al. Good Agricultural Practices for greenhouse vegetable production in the South East European countries-Principles for sustainable intensification of smallholder farms. (2017).
FAO, F. The future of food and agriculture—Trends and challenges. (2017).
Yeh, N. & Chung, J.-P. High-brightness LEDs—Energy efficient lighting sources and their potential in indoor plant cultivation. Renew. Sustain. Energy Rev. 13, 2175–2180 (2009).
Bula, R. J. et al. Light-emitting diodes as a radiation source for plants. HortScience 26, 203–205 (1991).
Bourget, C. M. An introduction to light-emitting diodes. HortScience 43, 1944–1946 (2008).
Morrow, R. C. LED lighting in horticulture. HortScience 43, 1947–1950 (2008).
Brown, C. S., Schuerger, A. C. & Sager, J. C. Growth and photomorphogenesis of pepper plants under red light-emitting diodes with supplemental blue or far-red lighting. J. Am. Soc. Hortic. Sci. 120, 808–813. https://doi.org/10.21273/JASHS.120.5.808 (1995).
Tennessen, D. J., Singsaas, E. L. & Sharkey, T. D. Light-emitting diodes as a light source for photosynthesis research. Photosynth. Res. 39, 85–92 (1994).
Schubert, E. F. & Kim, J. K. Solid-state light sources getting smart. J. Sci. 308, 1274–1278 (2005).
Robertson, J. (Google Patents, 2007).
Robertson, J. J. & Currie, R. M. (Google Patents, 2005).
Kit, J. (Google Patents, 2009).
Xiaoying, L., Shirong, G., Taotao, C., Zhigang, X. & Tezuka, T. Regulation of the growth and photosynthesis of cherry tomato seedlings by different light irradiations of light emitting diodes (LED). Afr. J. Biotechnol. 11, 6169–6177 (2012).
Yousef, A. F. et al. The influence of light spectral composition on the growth, pigments, biochemical and chlorophyll fluorescence characteristics of tomato seedlings (Solanum lycopersicum L.). Fresenius Environ. Bull. 29, 3575–3588 (2021).
Elmardy, N. A. et al. Photosynthetic performance of rocket (Eruca sativa. Mill.) grown under different regimes of light intensity, quality, and photoperiod. PLoS ONE 16, e0257745 (2021).
Liang, D. et al. Increasing the performance of Passion fruit (Passiflora edulis) seedlings by LED light regimes. Sci. Rep. 11, 1–13 (2021).
Borowski, E., Michałek, S., Rubinowska, K., Hawrylak-Nowak, B. & Grudzinski, W. The effects of light quality on photosynthetic parameters and yield of lettuce plants. J. Acta Scientiarum Polonorum 14, 177–188 (2015).
Loconsole, D., Cocetta, G., Santoro, P. & Ferrante, A. Optimization of LED lighting and quality evaluation of romaine lettuce grown in an innovative indoor cultivation system. J. Sustain. 11, 841. https://doi.org/10.3390/su11030841 (2019).
Wang, J., Lu, W., Tong, Y. & Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 7, 250 (2016).
Yan, Z., He, D., Niu, G. & Zhai, H. Evaluation of growth and quality of hydroponic lettuce at harvest as affected by the light intensity, photoperiod and light quality at seedling stage. Sci. Hortic. 248, 138–144 (2019).
Yousef, A. F. et al. Photosynthetic apparatus performance of tomato seedlings grown under various combinations of LED illumination. PLoS ONE 16, e0249373 (2021).
Muneer, S., Kim, E. J., Park, J. S. & Lee, J. H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 15, 4657–4670 (2014).
Fu, Y. et al. Interaction effects of light intensity and nitrogen concentration on growth, photosynthetic characteristics and quality of lettuce (Lactuca sativa L. Var. youmaicai). J. Sci. Hortic. 214, 51–57. https://doi.org/10.1016/j.scienta.2016.11.020 (2017).
Nguyen, T., Tran, T. & Nguyen, Q. Effects of light intensity on the growth, photosynthesis and leaf microstructure of hydroponic cultivated spinach (Spinacia oleracea L.) under a combination of red and blue LEDs in house. J. Int. J. Agric. Technol. 15, 75–90 (2019).
Kang, J. H., KrishnaKumar, S., Atulba, S. L. S., Jeong, B. R. & Hwang, S. J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. J. Hortic. Environ. Biotechnol. 54, 501–509. https://doi.org/10.1007/s13580-013-0109-8 (2013).
Pearson, K. V. I. I. Note on regression and inheritance in the case of two parents. J. Proc. R. Soc. Lond. 58, 240–242 (1895).
Kim, H.-H., Goins, G. D., Wheeler, R. M. & Sager, J. C. Stomatal conductance of lettuce grown under or exposed to different light qualities. Ann. Bot. 94, 691–697 (2004).
Franklin, K. A., Larner, V. S. & Whitelam, G. C. The signal transducing photoreceptors of plants. Int. J. Dev. Biol. 49, 653–664. https://doi.org/10.1387/ijdb.051989kf (2004).
Ward, J. M., Cufr, C. A., Denzel, M. A. & Neff, M. M. The Dof transcription factor OBP3 modulates phytochrome and cryptochrome signaling in Arabidopsis. Plant Cell 17, 475–485. https://doi.org/10.1105/tpc.104.027722 (2005).
Dou, H., Niu, G., Gu, M. & Masabni, J. G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled environments. Horticulturae 3, 36 (2017).
Jishi, T., Kimura, K., Matsuda, R. & Fujiwara, K. Effects of temporally shifted irradiation of blue and red LED light on cos lettuce growth and morphology. Sci. Hortic. 198, 227–232. https://doi.org/10.1016/j.scienta.2015.12.005 (2016).
Naznin, M. T., Lefsrud, M., Gravel, V. & Azad, M. O. K. Blue light added with red LEDs enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants 8, 93. https://doi.org/10.3390/plants8040093 (2019).
Zheng, L. & Van Labeke, M.-C. Long-term effects of red-and blue-light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants. Front. Plant Sci. 8, 917. https://doi.org/10.3389/fpls.2017.00917 (2017).
Brodersen, C. R. & Vogelmann, T. C. Do changes in light direction affect absorption profiles in leaves?. Funct. Plant Biol. 37, 403–412. https://doi.org/10.1071/FP09262 (2010).
Yang, Z. et al. Plant growth and development of pepper seedlings under different photoperiods and photon flux ratios of red and blue LEDs. Trans. Chin. Soc. Agric. Eng. 33, 173–180. https://doi.org/10.11975/j.issn.1002-6819.2017.17.023 (2017).
Yang, X. et al. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Environ. Exp. Bot. 150, 161–171. https://doi.org/10.1016/j.envexpbot.2018.03.013 (2018).
Jeong, H. W. et al. Using light quality for growth control of cucumber seedlings in closed-type plant production system. Plants 9, 639 (2020).
Kim, H. M. & Hwang, S. J. The growth and development of ‘mini chal’tomato plug seedlings grown under various wavelengths using light emitting diodes. Agronomy 9, 157 (2019).
XiaoYing, L., ShiRong, G., ZhiGang, X., XueLei, J. & Tezuka, T. Regulation of chloroplast ultrastructure, cross-section anatomy of leaves, and morphology of stomata of cherry tomato by different light irradiations of light-emitting diodes. HortScience 46, 217–221 (2011).
Darko, E., Heydarizadeh, P., Schoefs, B. & Sabzalian, M. R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B. Biol. Sci. 369, 20130243 (2014).
Sabzalian, M. R. et al. High performance of vegetables, flowers, and medicinal plants in a red-blue LED incubator for indoor plant production. Agron. Sustain. Dev. 34, 879–886 (2014).
Yousef, A. F. et al. Light quality and quantity affect graft union formation of tomato plants. Sci. Rep. 11, 1–11 (2021).
Yousef, A. F. et al. Effects of light spectrum on morpho-physiological traits of grafted tomato seedlings. PLoS One 16, e0250210 (2021).
Dong, J. Z. et al. Light wavelengths regulate growth and active components of c ordyceps militaris fruit bodies. J. Food Biochem. 37, 578–584 (2013).
Ichimura, M., Watanabe, H., Amaki, W. & Yamazaki, N. In VI International Symposium on Light in Horticulture 907. 91–94.
Gitelson, A. A., Gritz, Y. & Merzlyak, M. N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 160, 271–282. https://doi.org/10.1078/0176-1617-00887 (2003).
Bondada, B. R. & Syvertsen, J. P. Leaf chlorophyll, net gas exchange and chloroplast ultrastructure in citrus leaves of different nitrogen status. Tree Physiol. 23, 553–559 (2003).
Li, Q. Effects of Light Quality on Growth and Phytochemical Accumulation of Lettuce and Salvia miltiorrhiza Bunge (Northwest A F University, 2010).
Bugbee, B. In VIII International Symposium on Light in Horticulture 1134. 1–12.
Baroli, I., Price, G. D., Badger, M. R. & von Caemmerer, S. The contribution of photosynthesis to the red light response of stomatal conductance. Plant Physiol. 146, 737–747 (2008).
Bian, Z., Cheng, R., Wang, Y., Yang, Q. & Lu, C. Effect of green light on nitrate reduction and edible quality of hydroponically grown lettuce (Lactuca sativa L.) under short-term continuous light from red and blue light-emitting diodes. Environ. Exp. Bot. 153, 63–71. https://doi.org/10.1016/j.envexpbot.2018.05.010 (2018).
Cui, J. et al. Effects of supplemental lighting with different light qualities on growth and physiological characteristics of cucumber, pepper and tomato seedlings. Acta Hortic. Sin. 36, 663–670 (2009).
Pandey, S. & Singh, H. A simple, cost-effective method for leaf area estimation. J. Bot. https://doi.org/10.1155/2011/658240 (2011).
Knight, S. & Mitchell, C. Enhancement of lettuce yield by manipulation of light and nitrogen nutrition. HortSci. Publ. Am. Soc. Hortic. Sci. 108, 750–754 (1983).
Cataldo, D., Maroon, M., Schrader, L. & Youngs, V. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 6, 71–80. https://doi.org/10.1080/00103627509366547 (1975).
Li, L. & Jiao, X. Determining of protein using coomassia brilliant blue G-250 method. In Manual of Modern Plant Physiology Experiment (in Chinese), 392e394 (Science Press, 1999).
Yemm, E. & Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 57, 508. https://doi.org/10.1042/bj0570508 (1954).
Duncan, D. B. Multiple range and multiple F tests. J. Biometr. 11, 1–42. https://doi.org/10.2307/3001478 (1955).
Roy, R. A Primer on Taguchi Method (Van Noshtrand Reinhold Int. Co., Ltd., 1990).
Pearson, E. S. The test of significance for the correlation coefficient. J. Am. Stat. Assoc. 26, 128–134 (1931).
Acknowledgements
The authors thank the valuable contributions for data collection provided by Naif Ali Elmardy and Helio Guiamba. Helpful suggestions were provided by Kui Lin and Xiwen Zheng for data analysis.
Author information
Authors and Affiliations
Contributions
Conceptualization, A.M.H. and A.F.Y.; methodology, A.M.H. and A.F.Y.; software, A.F.Y., M.O.K and M.A.A.; validation, A.M.H., A.F.Y., and M.A.A.; formal analysis, A.F.Y. and M.A.A.; investigation, A.F.Y.; resources, A.F.Y.; data curation, A.M.H. and A.F.Y.; writing—original draft preparation, A.F.Y., W.M.A., J.G.W. and H.M.K.; writing—review and editing, W.M.A., M.M.A., J.G.W., and H.M.K.visualization, A.F.Y.; supervision, A.F.Y.; project administration, M.O.K and A.F.Y.; funding acquisition, A.F.Y., All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hamedalla, A.M., Ali, M.M., Ali, W.M. et al. Increasing the performance of cucumber (Cucumis sativus L.) seedlings by LED illumination. Sci Rep 12, 852 (2022). https://doi.org/10.1038/s41598-022-04859-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-04859-y
This article is cited by
-
CsSHMT3 gene enhances the growth and development in cucumber seedlings under salt stress
Plant Molecular Biology (2024)
-
Optimizing supplemental light spectrum improves growth and yield of cut roses
Scientific Reports (2023)
-
Different combinations of red and blue LED light affect the growth, physiology metabolism and photosynthesis of in vitro-cultured Dendrobium nobile ‘Zixia’
Horticulture, Environment, and Biotechnology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
