
Abstract
The spike protein of SARS-CoV-2 engages the human angiotensin-converting enzyme 2 (ACE2) receptor to enter host cells, and neutralizing antibodies are effective at blocking this interaction to prevent infection. Widespread application of this important marker of protective immunity is limited by logistical and technical challenges associated with live virus methods and venous blood collection. To address this gap, we validated an immunoassay-based method for quantifying neutralization of the spike-ACE2 interaction in a single drop of capillary whole blood, collected on filter paper as a dried blood spot (DBS) sample. Samples are eluted overnight and incubated in the presence of spike antigen and ACE2 in a 96-well solid phase plate. Competitive immunoassay with electrochemiluminescent label is used to quantify neutralizing activity. The following measures of assay performance were evaluated: dilution series of confirmed positive and negative samples, agreement with results from matched DBS-serum samples, analysis of results from DBS samples with known COVID-19 status, and precision (intra-assay percent coefficient of variation; %CV) and reliability (inter-assay; %CV). Dilution series produced the expected pattern of dose–response. Agreement between results from serum and DBS samples was high, with concordance correlation = 0.991. Analysis of three control samples across the measurement range indicated acceptable levels of precision and reliability. Median % surrogate neutralization was 46.9 for PCR confirmed convalescent COVID-19 samples and 0.1 for negative samples. Large-scale testing is important for quantifying neutralizing antibodies that can provide protection against COVID-19 in order to estimate the level of immunity in the general population. DBS provides a minimally-invasive, low cost alternative to venous blood collection, and this scalable immunoassay-based method for quantifying inhibition of the spike-ACE2 interaction can be used as a surrogate for virus-based assays to expand testing across a wide range of settings and populations.
Similar content being viewed by others

Introduction
Coronavirus disease 2019 (COVID-19) is a potentially deadly infectious disease that is caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)1. Serological testing is an important part of the effort to slow the spread of SARS-CoV-2, and detects the presence of antibodies in blood samples from exposed individuals2. Antibodies are detectable in 3–10 days after infection, and anti-SARS-CoV IgG antibodies persist for at least 5 months after the resolution of symptoms3,4. Neutralizing antibodies (NAbs) are of particular importance because they can bind to viral proteins and inhibit viral entry into host cells, thereby preventing infection5. For SARS-CoV-2, the receptor binding domain (RBD) of the surface spike protein engages the human angiotensin-converting enzyme 2 (ACE2) receptor to gain entry into host cells, and anti-SARS-CoV-2 NAbs are effective at blocking this interaction and preventing viral entry6,7.
Serological testing has been widely used to identify individuals who have been exposed to SARS-CoV-2. Importantly, due to their functional role in blocking viral entry, the measurement of NAbs provides information on the magnitude of immune response to prior infection and the level of protection against re-infection8. In addition, NAbs can be used to assess conferred immunity through vaccination, as neutralizing antibody response is frequently used as a correlate of protection following vaccination, including vaccination against SARS-CoV-29,10.
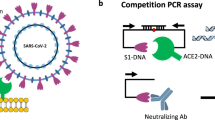
Conventional methods for measuring neutralizing activity require either a non-replicating virus with the SARS-CoV-2 spike protein expressed from a viral vector (“pseudotyped virus” or “pseudovirus”) or replication-competent SARS-CoV-2. These approaches limit widespread application, as the assays are time-consuming and require specialized laboratory containment facilities. SARS-CoV-2 neutralizing antibodies can also be quantified utilizing a surrogate virus neutralization test (sVNT), which detects NAbs without using live virus and can be implemented with immunoassay techniques6,11. The sVNT approach replicates the virus-host interaction by incubating a blood sample with purified versions of viral spike protein and human ACE2 receptor. Neutralizing antibodies prevent the spike protein from binding to ACE2, and the degree of inhibition is quantified in a competitive immunoassay to indicate the level of NAbs in the sample. There is a high level of agreement in results derived from sVNT and live virus methods, and the specificity and sensitivity of sVNT are comparable to conventional neutralization tests11,12.
While sVNT provides a more streamlined alternative to conventional assays for neutralizing antibodies, the use of venous blood limits the screening of large numbers of people in non-clinical settings due to the costs and logistical constraints associated with collecting, processing, and transporting venous blood. There is an alternative in finger stick dried blood spot (DBS) sampling13. With DBS, a lancet is used to prick the finger, and up to five drops of blood are placed on filter paper, where it dries. In comparison with venipuncture, DBS sampling is low-cost, reduces biohazard risk during sample collection and transport, and it affords the possibility of self-sampling, with participants collecting their own blood and returning the sample through the mail, without the need for a cold chain or special handling14. Due to these advantages, DBS sampling is the foundation of neonatal screening programs in the US, and it is increasingly applied as a minimally-invasive alternative to venipuncture in community- and population-based health research, including several applications in infectious disease epidemiology15,16.
We recently validated a protocol for quantifying anti-RBD SARS-CoV-2 IgG antibodies in DBS samples that is being used in several community-based seroprevalence studies17,18,19. Here, we describe a method for quantifying neutralization of spike-ACE2 binding in DBS samples. In contrast to previously described methods, the protocol is relatively short (< 4 h), can be implemented with widely available laboratory infrastructure, does not require venous blood, and can be scaled up for high-throughput processing. We describe the assay protocol, as well as results demonstrating performance characteristics and the validity of measuring SARS-CoV-2 neutralizing antibodies in DBS samples.
Materials and methods
The DBS method is based on modifications to a commercially available protocol (Meso Scale Diagnostics V-PLEX SARS-CoV-2 Panel 2 Kit; K15386U-2), designed for the multiplex capture of SARS-CoV-2 antibodies in serum on the Meso Scale Diagnostics QuickPlex SQ 120 Imager. This method utilizes a 96-well solid phase plate with antigens printed as arrays in discrete spots within each well, an electrochemiluminescent detection system, and a competitive binding protocol. The assay principle is as follows: diluted sample is incubated in plate wells in the presence of ACE2 conjugated with an electrochemiluminescent label; neutralizing antibodies, if present, compete with conjugated ACE2 for viral antigen binding sites; after separation an electrical impulse generates light in proportion to bound ACE2 at each spot in the array; the signal is negatively proportional to the concentration of neutralizing antibodies in the sample, and % neutralization is calculated as follows: % neutralization = 100 × (1 − (sample signal/negative control signal)).
Serum assay protocol
Serum samples were quantified per manufacturer’s instructions. Briefly, the assay plate included two SARS-CoV-2 antigens coated to the bottom of each well: spike and RBD. The nucleocapsid antigen is also available in the plate but does not bind ACE2 and is not a target for neutralizing antibodies; it was therefore not used in this protocol. The assay plate was blocked for 30 min and washed. Serum samples were diluted (1:40) and 25 μL were transferred to each well. The plate was then incubated at room temperature for 60 min with shaking at 700 rpm, followed by the addition of SULFO-TAG conjugated ACE2, and continued incubation with shaking for 60 min. The plate was washed, 150 μL MSD GOLD Read Buffer B was added to each well, and the plate was read using the QuickPlex SQ 120 Imager. Mean fluorescence intensity (MFI) values were generated for each sample. Sample diluent was used as the negative control for the calculation of % neutralization.
DBS assay protocol
Three levels of DBS control material (low, mid, and high neutralization antibodies) were manufactured by diluting neutralizing monoclonal antibody to SARS CoV-2 spike (provided with kit) in assay diluent, mixing with an equal volume of washed red blood cells, then transferring to filter paper (Whatman 903 Protein Saver Card, #10534612) in 60 μL drops using a pipette (Rainin pipet lite LTS L100)13. A DBS negative control sample was made with the same procedure, using assay diluent.
To begin the protocol, DBS samples and DBS controls were eluted overnight in a non-binding 96-well plate (Corning 96 Well TC-Treated Microplate; Sigma #CLS3599). One 5.0 mm disc was removed using a pneumatic hole punch (Analytical Sales and Services #327500, Flanders, NJ), 100 μL assay diluent were added to each well, and the plate was covered and incubated overnight at 4 °C.
The following day, the elution plate was removed from the refrigerator and rotated for 1 h at 300 rpm. The MULTI-SPOT 96-well assay plate was blocked for 30 min at room temperature, washed, and 25 μL of each sample were transferred from the elution plate to the assay plate in duplicate. The assay plate was then incubated at room temperature for 60 min while shaking at 700 rpm, followed by the addition of 25 μL of SULFO-TAG conjugated ACE2, and continued incubation with shaking for 60 min. The plate was washed, 150 μL MSD GOLD Read Buffer B was added to each well, and the plate was read using a MESO QuickPlex SQ 120MM Imager. Mean MFI values were generated for the duplicates of each sample, and the DBS negative control was used to calculate % neutralization for each sample.
Evaluation of assay performance
The following aspects of DBS assay performance were evaluated: dilution series of confirmed positive and negative samples, agreement with results from serum samples, analysis of results from DBS samples with known COVID-19 status, and precision and reliability. Assay precision, or intra-assay variability, was calculated as the percent coefficient of variation (CV; SD/mean × 100) for 10 replicates each of DBS control samples with high, medium, and low levels of neutralizing antibodies. Reliability, or inter-assay variability, was assessed by running the three DBS control samples across ten separate assay plates.
Agreement between results from DBS and serum samples was evaluated in a set of matched samples collected from n = 20 convalescent individuals with PCR positive confirmed cases of COVID-19, and n = 10 negative samples collected as DBS in early 2019, before the pandemic. Immediately following collection of venous blood, five drops of whole blood (60 μL each) were transferred by pipet to filter paper (Whatman #903, GE Healthcare, Pascataway, NJ), allowed to dry overnight, and then stacked and stored at – 30 °C. DBS samples with known SARS-CoV-2 seropositivity status were drawn from an ongoing community-based seroprevalence study and collected between June 24 and November 11, 2020. Serostatus was determined based on the presence of IgG antibodies against SARS-CoV-2 RBD, using an immunoassay protocol previously validated for use with DBS17,18. These samples were categorized as seronegative (n = 38), seropositive with PCR confirmed COVID-19 (n = 51), and seropositive with no PCR test (n = 32). Individuals with PCR confirmed cases of COVID-19 reported an average of 5.5 symptoms. Cases for seropositive and untested individuals were mild, with an average of 1.7 symptoms, and 59.5% reporting no symptoms of infection. An additional set of negative DBS samples, collected and stored as dried blood spots in early 2019, were also assayed (n = 27). All samples were de-identified and all research activities were implemented under protocols approved by the institutional review board at Northwestern University (#STU00212371, #STU00212457 and #STU00212472). All participants were 18 years of age or older and provided informed consent. All research was performed in accordance with the relevant guidelines and regulations.
All samples were analyzed in duplicate using the DBS and serum protocols, as described above. Passing-Bablok regression models and difference plots were used to evaluate patterns of agreement in results20,21,22,23. Statistical analyses were implemented with Stata/SE (version 15.1 for Windows, StataCorp, College Station, TX). Results are presented for neutralization of the spike-ACE2 interaction only. Results for RBD neutralization of RBD-ACE2 interaction were highly correlated with spike (Spearman R = 0.90) and are included in supplementary information.
Results
Analysis of DBS samples from convalescent, PCR confirmed cases of COVID-19 demonstrated that the SARS-CoV-2 Spike-ACE2 interaction can be neutralized in a dose-dependent manner (Fig. 1). There was no evidence of neutralization in pre-pandemic negative DBS samples. Results from a set of matched DBS and serum samples, including confirmed positive and negative cases, indicated a high level of agreement in % neutralization across the two sample types (concordance correlation = 0.991) (Fig. 2). A Bland–Altman difference plot indicated that the pattern of association was linear and consistent across the measurement range, with relatively tight 95% limits of agreement (bias = 0.63% inhibition; lower limit = − 8.4; upper limit = 9.6) (Figure S1).

Neutralization of SARS-CoV-2 Spike-ACE2 interaction in DBS samples from convalescent COVID-19 cases and pre-pandemic negative samples. Reduction in neutralization with dilution of 12 DBS samples (1:4, 1:16, 1:64, 1:256) after initial elution of one 5 mm punch in 100 μL assay buffer. Negative samples are indicated with dashed lines.

Agreement in neutralization results for matched serum and DBS samples. Scatterplot and Passing-Bablok regression line (95% CI) demonstrating agreement in % neutralization of SARS-CoV-2 Spike-ACE2 interaction in DBS and serum samples. PCR positive samples are indicated with open circles; negative samples are indicated with black circles. The concordance correlation of absolute agreement = 0.991.
Repeat analysis of three NAb-positive DBS control samples indicated a high level of precision and reliability for the assay, particularly at higher levels of neutralization of the spike-ACE2 interaction. Levels of intra-assay variability for high, mid, and low NAb positive DBS control samples were 0.2, 3.2, and 14.9%CV, respectively. Inter-assay variability was 0.7, 5.7, and 14.1%CV, respectively.
Analysis of DBS samples with known COVID-19 serostatus produced the expected pattern of results, with the highest levels of neutralization in samples from convalescent, PCR confirmed cases (Fig. 3). Median neutralization in this group was 46.9%, but values spanned the entire measurement range. All negative samples (seronegative and pre-pandemic) showed low levels of neutralization, with median % neutralization of 0.1%.

Neutralization of SARS-CoV-2 Spike-ACE2 interaction in DBS samples. Negative samples were collected in 2019 or were seronegative for anti-RBD IgG; PCR positive samples are from convalescent COVID-19 cases. Mild/asymptomatic samples are from individuals who were seropositive for anti-RBD IgG but did not get a PCR test for acute infection. The black line represents median neutralization for each group. Wilcoxon rank-sum tests were used to evaluate differences across the groups. *p < 0.05; ***p < 0.001.
We used the pre-pandemic, confirmed negative samples to establish a cut-off value for differentiating a positive surrogate neutralization response from background responses in SARS-CoV-2 negative samples. For these samples, mean % surrogate neutralization was 4.2% (SD = 4.5). Using 2SD above the mean as a criterion, the cut-off value is 13.2%. Neutralizing activity above this threshold was detected in 74.5% of samples from confirmed PCR positive cases. For individuals who tested seropositive for SARS-CoV-2 anti-RBD IgG, but who had mild/asymptomatic infections, 18.8% of samples tested above background.
Discussion
Serological testing is critical for tracking the spread of SARS-CoV-2, and for identifying the factors that influence transmission, immunity, and vulnerability to severe infection. High rates of asymptomatic and mild infection underscore the need for approaches that are amenable to large-scale application. Testing for neutralizing antibodies may be particularly important at this stage in the pandemic, as it can inform our understanding of immunity among vaccinated individuals and those previously exposed to SARS-CoV-2. We have validated a method for quantifying neutralization of the SARS-CoV-2 spike-ACE2 interaction in a single drop of blood, collected on filter paper following a simple finger stick. The protocol is based on a commercially available immunoassay platform, and can be scaled up for large-scale testing of samples collected in centralized or remote locations.
Our results demonstrate the feasibility of using DBS samples for serological testing of SARS-CoV-2 neutralizing antibodies with a competitive immunoassay that quantifies inhibition of the spike-ACE2 interaction. Neutralization is high in samples from PCR confirmed convalescent cases of COVID-19, and low in samples that are known to be negative for SARS-CoV-2. We document dose-dependent reduction in neutralization in dilutions of samples with high initial neutralization, as well as close correspondence in results from DBS in comparison with matched serum samples. The method also produces results that are precise and reliable, which is particularly important for studies with large numbers of samples assayed across multiple plates. Furthermore, the method can be implemented in ubiquitous BSL2 laboratories, without the specialized facilities and biohazard risks associated with live virus methods.
Finger stick DBS sampling is relatively low-cost and minimally-invasive, and samples can be self-collected in the home14,15. In addition, antibodies remain stable in DBS for several weeks at room temperature, and can be sent in the mail as nonregulated, exempt materials24. The possibility of remote serological testing, with large numbers of people, may be particularly advantageous when clinical resources are stretched thin and people are being encouraged to stay home. A disadvantage of DBS includes the small volume of blood in comparison with venipuncture. Other potential issues, depending on the application, are variable quality of self-collected samples, and the possibility that samples may be delayed or lost during shipment by mail.
Recent studies have documented neutralization antibody responses to vaccination and natural infection with SARS-CoV-28,25,26, and prior research with other viruses suggests that neutralizing antibodies are excellent indicators of protective immunity9. More widespread, community-based testing is important for determining the level of neutralizing antibodies that provide protection against COVID-19, and for estimating the level and duration of immunity in the general population—and subpopulations of particular interest—following multiple waves of community transmission27. Testing for neutralization antibodies can also play a role in evaluating the effectiveness of vaccines, and heterogeneity in vaccine responses across individuals8. Our quantitative DBS-based assay for neutralizing antibodies can help advance research and surveillance in these important areas.
References
Wu, F. et al. A new coronavirus associated with human respiratory disease in China. Nature 579, 265–269 (2020).
Krammer, F. & Simon, V. Serology assays to manage COVID-19. Science 368, 1060–1061 (2020).
Wajnberg, A. et al. SARS-CoV-2 infection induces robust, neutralizing antibody responses that are stable for at least three months. medRxiv. https://doi.org/10.1101/2020.07.14.20151126 (2020).
Marklund, E. et al. Serum-IgG responses to SARS-CoV-2 after mild and severe COVID-19 infection and analysis of IgG non-responders. PLoS ONE 15, e0241104 (2020).
McMahan, K. et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 590, 630–634 (2020).
Abe, K. T. et al. A simple protein-based SARS-CoV-2 surrogate neutralization assay. JCI Insight 5, 19 (2020).
Hoffmann, M. et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271–280 (2020).
Wu, F. et al. Neutralizing antibody responses to SARS-CoV-2 in a COVID-19 recovered patient cohort and their implications. medRxiv. https://doi.org/10.1101/2020.03.30.20047365 (2020).
Zinkernagel, R. M. On natural and artificial vaccinations. Annu. Rev. Immunol. 21, 515–546 (2003).
Walsh, E. E. et al. Safety and immunogenicity of two RNA-based Covid-19 vaccine candidates. N. Engl. J. Med. 383, 2439–2450 (2020).
Tan, C. W. et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2–spike protein–protein interaction. Nat. Biotechnol. 38, 1073–1078 (2020).
Johnson, M. et al. Evaluation of a novel multiplexed assay for determining IgG levels and functional activity to SARS-CoV-2. J. Clin. Virol. 130, 104572 (2020).
McDade, T. W. Development and validation of assay protocols for use with dried blood spot samples. Am. J. Hum. Biol. 26, 1–9 (2014).
Roberts, S. et al. Self-collected dried blood spots as a tool for measuring ovarian reserve in young female cancer survivors. Hum. Reprod. 31, 1570–1578 (2016).
McDade, T. W., Williams, S. & Snodgrass, J. J. What a drop can do: Dried blood spots as a minimally invasive method for integrating biomarkers into population-based research. Demography 44, 899–925 (2007).
Lehmann, S., Delaby, C., Vialaret, J., Ducos, J. & Hirtz, C. Current and future use of “dried blood spot” analyses in clinical chemistry. Clin. Chem. Lab. Med. 51, 1897–1909 (2013).
McDade, T. W. et al. Enzyme immunoassay for SARS-CoV-2 antibodies in dried blood spot samples: A minimally-invasive approach to facilitate community-and population-based screening. medRxiv. https://doi.org/10.1101/2020.04.28.20081844 (2020).
McDade, T. W. et al. High seroprevalence for SARS-CoV-2 among household members of essential workers detected using a dried blood spot assay. PLoS ONE 15, e0237833 (2020).
Demonbreun, A. R. et al. Patterns and persistence of SARS-CoV-2 IgG antibodies in a US metropolitan site. medRxiv https://doi.org/10.1101/2020.11.17.20233452 (2020).
Stockl, D., Dewitte, K. & Thienpont, L. M. Validity of linear regression in method comparison studies: Is it limited by the statistical model or the quality of the analytical input data?. Clin. Chem. 44, 2340–2346 (1998).
Passing, H. & Bablok, W. A new biometrical procedure for testing the equality of measurements from two different analytical methods. Application of linear regression procedures for method comparison studies in clinical chemistry, Part I. Clin. Chem. Lab. Med. 21, 709–720 (1983).
Bland, J. M. & Altman, D. G. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet 327, 307–310 (1986).
Bland, J. M. & Altman, D. G. Measuring agreement in method comparison studies. Stat. Methods Med. Res. 8, 135–160 (1999).
Centers for Disease Control & Prevention. Shipping guidelines for dried-blood spot specimens. Laboratory Quality Assurance and Standardization Programs. https://www.cdc.gov/labstandards/pdf/nsqap/Bloodspot_Transportation_Guidelines.pdf. Accessed 15 Jan 2021.
Seow, J. et al. Longitudinal observation and decline of neutralizing antibody responses in the three months following SARS-CoV-2 infection in humans. Nat. Microbiol. 5, 1598–1607 (2020).
Widge, A. T. et al. Durability of responses after SARS-CoV-2 mRNA-1273 vaccination. N. Engl. J. Med. 384, 80–82 (2021).
Crawford, K. H. et al. Protocol and reagents for pseudotyping lentiviral particles with SARS-CoV-2 Spike protein for neutralization assays. Viruses 12, 513 (2020).
Funding
Funding was provided by National Science Foundation (Grant No. 2035114), National Institutes of Health (Grant No. UL1TR001422).
Author information
Authors and Affiliations
Contributions
A.S. and T.M. designed and implemented the analyses, and drafted the manuscript. E.M., R.D., and A.D. provided input on study design and interpretation of results, and edited the manuscript. M.V., M.I., and A.D. provided samples and data. All authors reviewed and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
Dr. McDade reports that he has a financial interest in EnMed Microanalytics, a company that conducts lab tests using DBS samples. Dr. D'Aquila reports personal fees from Abbvie, outside the submitted work. Dr. McNally reports personal fees from Amgen, personal fees from AstraZeneca, personal fees from Cytokinetics, personal fees from Pfizer, personal fees from Tenaya Therapeutics, personal fees from 4D Molecular Therapeutics, outside the submitted work. The remaining authors declare no additional competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sancilio, A.E., D’Aquila, R.T., McNally, E.M. et al. A surrogate virus neutralization test to quantify antibody-mediated inhibition of SARS-CoV-2 in finger stick dried blood spot samples. Sci Rep 11, 15321 (2021). https://doi.org/10.1038/s41598-021-94653-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-94653-z
This article is cited by
-
Quantifying neutralising antibody responses against SARS-CoV-2 in dried blood spots (DBS) and paired sera
Scientific Reports (2023)
-
Detection of neutralizing antibodies against multiple SARS-CoV-2 strains in dried blood spots using cell-free PCR
Nature Communications (2022)
-
Durability of antibody response to vaccination and surrogate neutralization of emerging variants based on SARS-CoV-2 exposure history
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
