
Abstract
An extensive network of storm water conveyance systems in urban areas, often referred to as the “underground storm drain system” (USDS), serves as significant production habitats for mosquitoes. Knowledge of whether USDS habitats are suitable for newly introduced dengue vectors Aedes aegypti and Ae. albopictus will help guide surveillance and control efforts. To determine whether the USDS functions as a suitable larval habitat for Culex, Ae. aegypti and Ae. albopictus in southern California, we examined mosquito habitat utilization and larval survivorship using laboratory microcosm studies. The data showed that USDS constituted 4.1% of sampled larval habitats for Ae. aegypti and Ae. albopictus, and 22.0% for Cx. quinquefasciatus. Furthermore, USDS water collected in the summer completely inhibited Aedes larval development, but yielded a 15.0% pupation rate for Cx. quinquefasciatus. Food supplementation in the microcosms suggests that nutrient deficiency, toxins and other factors in the USDS water led to low success or complete failure of larval development. These results suggest that USDS habitats are currently not major productive larval habitats for Aedes mosquitoes in southern California. Our findings prompt inclusion of assessments of pupal productivity in USDS habitats and adult mosquito resting sites in the mosquito surveillance program.
Similar content being viewed by others


Introduction
Rapid urbanization in the past several decades have altered microclimatic conditions and natural ecology, which may subsequently affect the ecology of disease vectors and risk of mosquito-borne diseases in heavily populated landscapes1,2,3. For example, more mosquito larval habitats and higher abundance of the yellow fever mosquito Aedes aegypti, the most important vector of dengue, chikungunya, and Zika viruses, were found in urban areas than in suburban and rural areas in Côte d’Ivoire, Africa4. Similarly, not only were more larval habitats found for the Asian tiger mosquito, Aedes albopictus, in urban areas of southern China, Aedes larvae developed faster and the adult emergence rate was higher than in suburban and rural areas, partly due to higher ambient temperatures associated with heat islands in urban areas5.
Aedes aegypti and Ae. albopictus are considered as the most invasive mosquitoes in the world6,7. In the state of California, USA, Ae. albopictus was introduced in 2001 and considered established in 2011, while Ae. aegypti was discovered in 20138. In southern California, common aquatic mosquito habitats include peridomestic sources (e.g., small artificial water-holding containers, unmaintained ornamental ponds and swimming pools)9, and an extensive network of storm water conveyance systems consisting of catch basins, manhole chambers, underground vaults, pipelines and tunnels, and is collectively referred to as the “underground storm drain system (USDS)”10.
The USDS is designed to rapidly direct water from heavy rainstorms to large channels to slow runoff and reduce erosion, improve water quality and avoid flooding of streets, homes, and businesses. However, non-stormwater runoff into the USDS from landscape irrigation of residential and commercial establishments, as well as highways and streets, in highly populated southern California is tremendous. The USDS has been documented as suitable, year-round larval habitats for Culex quinquefasciatus mosquitoes at times and places in the system where subsurface water drainage is slow and irregular, as often by design or damage10,11. In addition, the arid climate of southern California can desiccate small containers rapidly, so large numbers of stable aquatic habitats in the USDS may pose very significant challenges for the control of Aedes-transmitted viruses if these habitats are suitable to egg laying and larval development for invasive Aedes. However, because aquatic habitats in the USDS tend to have low dissolved oxygen, above normal electrical conductivity and salinity levels10, and perhaps residual pesticides from dry-weather runoff during summer months12, USDS water may inhibit larval development and oviposition choice by gravid female mosquitoes.
Larval habitats in the USDS are extremely difficult to access and apply pesticides for mosquito control. A better understanding of the larval ecology and pupal productivity of invasive Aedes mosquitoes in USDS aquatic habitats will facilitate the development of rational control strategies. Given the rapid spread of Ae. aegypti and Ae. albopictus in many regions of the world13,14,15, information on the larval ecology of invasive Aedes mosquitoes in USDS water will be useful in assessing the impact of environmental changes on the risk of arboviruses transmitted by these exotic Aedes mosquitoes in southern California. The objective of this study was to assess habitat utilization by Ae. aegypti and Ae. albopictus following their discovery in Orange County, California, in 2015 and to examine whether the USDS provides suitable habitats for Ae. aegypti and Ae. albopictus larvae through life table studies in controlled microcosms and extensive field surveys. We investigated: (1) habitat usage by invasive Aedes mosquitoes from 2016–2019, (2) suitability of the USDS habitat for invasive Aedes egg laying, hatching, and larval development, and (3) impact of Cx. quinquefasciatus larvae, the predominant mosquito species in urban southern California16,17, on Aedes oviposition preference and larval survival.
Methods
Ethics and vertebrate animals
The field surveys and collections were conducted on accessible public areas or private residential areas with property owners’ permission. The study did not involve human participants, or endangered or protected species. Laboratory mice were used as a blood source for mosquitoes. All experimental protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of the University of California, Irvine (UCI) (IACUC protocol number: AUP-19-165). All methods were carried out in accordance with relevant IACUC guidelines and regulations.
Study sites and mosquito larval habitat surveillance
The study was carried out in Orange County, California, USA. Orange County is a highly urbanized county with an estimated population density of approximately 1470 people/km2 according to U.S. Census Bureau, an average annual low/high temperature range of 13–25 °C, 65% relative humidity, and annual precipitation of about 350 mm according to U.S. Climate Data. Annual rainfall was 261 mm, 311 mm, 198 mm and 475 mm for 2016, 2017, 2018 and 2019, respectively. A major drought event occurred in December 2017 and February 2018 when the total rainfall in the 3-month period was 20.6% of the 30-year average. Both Ae. aegypti and Ae. albopictus were discovered in the county in 20158. Culex quinquefasciatus is the most abundant mosquito in the county and breeds readily in a variety of residential, commercial and USDS water sources, and is the primary vector of West Nile virus in southern California18.
Larval mosquito surveillance in Orange County was conducted from 2016 to 2019 by the Orange County Mosquito and Vector Control District (OCMVCD) through its routine mosquito surveillance and treatment program, following the recommendations of the California Department of Public Health and the Mosquito and Vector Control Association of California19. Briefly, OCMVCD staff conducted routine inspection for aquatic habitats in randomly selected public areas, and performed door-to-door mosquito larval and adult sampling on residential or commercial premises upon the request of the residents or business owners while distributing public education materials for vector control and personal protection. Arial photography was used to examine the presence of abandoned swimming pools in residential areas. In addition to surface aquatic habitats, subsurface habitats (e.g., catch basins, underground drains, manhole chambers, and public utility vaults) were examined for larval abundance of all mosquito species. In 2019, OCMVCD completed 5,622 mosquito service requests, and conducted 11,813 inspection and treatments on routine sites using a variety of public health-approved adulticides and larvicides. A total of 38,099 underground drains and catch basins and 6925 km of flood channels were treated. In addition, a total of 17,783 km of gutters and 3562 neglected swimming pools were inspected and treated. The larval distribution data reported here were based on this extensive field sampling effort20.
Larval sampling used standard mosquito dippers or pipettes, and specialized modifications of these to sample hard to reach areas. Mosquito larvae from each source were collected, transferred into a uniquely-numbered vial with isopropyl alcohol (70%), and submitted to the laboratory for identification; if present, live pupae were collected and held in site-specific labelled rearing chambers (BioQuip Products, Inc., Rancho Dominguez, CA) until emergence. Third and fourth instar mosquito larvae (1–100, depending on sample size) and emerged adults were identified to species using a stereo microscope (40–50x) and morphological features described in taxonomic keys21,22. Results were uploaded to OCMVCD’s data management system, along with collection date, GPS location, and habitat type for each sample site. For this study, larval habitats were classified into six types: small container, underground system, ornamental water features, marsh, pools/spas, and creek (Table S1). The container classification included flowerpots/vases, saucers, tires, bowls, boxes, buckets, dishes, tree holes, etc. Underground storm drain system referred to larval habitats such as catch basins, manhole chambers, underground drains, and public utility vaults that were below the ground. Water feature included flood control channels, ponds, fountains, birdbaths, street gutters and small reservoirs, etc. Marsh included both fresh and salt water marshes.
Mosquito strains and water source for laboratory studies
We examined the effect of USDS water on oviposition substrate preference and larval development in microcosms in an insectary with climate control (27 ± 1 °C, 70 ± 10% relative humidity, and 12 h light/12 h dark photoperiod) at UCI. To minimize potential bias on behavior and ecology from mosquito colonization, this study did not use previously established laboratory mosquito colonies. Instead, we used Ae. aegypti and Ae. albopictus adults reared from field-collected eggs using ovicups in residential areas of Orange and Los Angeles Counties, California, respectively. Culex quinquefasciatus were also reared from eggs of field-collected, blood-engorged adult mosquitoes using gravid traps in Orange County23.
All experiments reported here used two types of habitat water: (1) USDS water collected from seven manhole chambers or catch basins (33°47′01.9"N, 117°53′19.0"W, Orange City, manhole; 33°52′25.0"N, 117°57′02.6"W, Fullerton City, manhole; 33°44′44.4"N, 118°06′24.2"W, Seal Beach City, manhole; 33°55′38.9"N, 117°56′51.4"W, La Habra City, manhole; 33°52′48.9"N, 117°55′21.4"W, Fullerton City, catch basin; 33°54′35.2"N, 117°56′02.5"W, Fullerton City, catch basin; 33°52′25.0"N, 117°57′02.6"W, Fullerton City, catch basin); and 2) flowerpot water from vases of three cemeteries in Orange County (33°50′29.0"N, 117°53′57.9"W; 33°46′21.5"N, 117°50′35.8"W; 33°46′12.3"N, 117°50′21.4"W). Water (including sediments) from each breeding source was collected with mosquito dippers and mixed together by habitat type into 18.9 L (five-gallon) Nalgene™ containers. The containers were transported to the laboratory in shaded ice containers, and stored overnight in a refrigerator at 4 °C. The experiments described below were conducted on the field-collected water for the two habitat types. We selected flowerpot water as the comparison substrate because flowerpot containers showed the highest larval positivity rate in the study area.
Oviposition preference test
To examine whether USDS water attracts or repels egg laying by Ae. aegypti and Ae. albopictus mosquitoes, a two-choice oviposition preference test was conducted. Briefly, this experiment used two ovicups placed within a mosquito cage (1 × 0.5 × 0.5 m3), one ovicup with 200 ml USDS water and another with 200 ml flowerpot water. Adult mosquitoes were bloodfed on mice; fully engorged females 3-days post-bloodfeeding were used for oviposition preference tests. Ten gravid Ae. aegypti females were released into a cage and allowed to lay eggs for three days, and the number of eggs in each ovicup were counted. Five replicates were used. The same experiment was conducted for Ae. albopictus.
To evaluate whether the presence of Cx. quinquefasciatus larvae has any impact on the egg laying behavior of invasive Aedes mosquitoes, the two-choice oviposition preference test described above was used. One ovicup contained 200 ml USDS water and ten first-instar Cx. quinquefasciatus larvae, while the second ovicup contained 200 ml USDS water only. Ten gravid Ae. aegypti or Ae. albopictus females were released into a cage and allowed to lay eggs for three days. Five replicates were used. We also conducted this experiment using flowerpot water with the same design and same number of replicates to determine whether the impact of Cx. quinquefasciatus larvae on Aedes mosquito egg laying behavior was similar across different water substrate types.
Egg hatching
To investigate the effects of different habitat water sources on egg hatching, 50 Ae. aegypti or Ae. albopictus eggs on separate filter papers were introduced into ovicups with 200 ml USDS water or flowerpot water. Deoxygenized distilled water that we routinely use in laboratory mosquito colony maintenance was used as a positive control. The experiment was conducted in an insectary with climate control (27 ± 1 °C). The number of larvae hatched were counted daily for six days continuously. Five replicates were used.
Larval survivorship
A life table study was conducted on Ae. aegypti and Ae. albopictus larvae to determine the effect of USDS water and flowerpot water on larval development and survivorship. Twenty-five newly hatched Ae. aegypti or Ae. albopictus larvae were introduced into a microcosm that contained 200 ml USDS or field flowerpot water. The number of dead and surviving larvae was recorded daily until they pupated. Pupae were counted, and removed to different paper cups for emergence to adults. Four replicates were used for each type of habitat water per species. We included Cx. quinquefasciatus in the larval life table study for method validation purposes because the larvae of this species were known to successfully develop into pupae and adults in USDS water in southern California10.
Larval survivorship experiments were conducted in two different seasons. The first was in the summer (August–September) 2019 when the density of invasive Aedes species peaked19, and also insecticide runoff from mosquito and residential/agricultural pest control applications were at the highest levels in southern California24. The second was in the winter (December) 2019 when there was little insecticide treatment for mosquito and pest control. This design enabled us to examine seasonality in larval survivorship and the impact of environmental insecticide runoff in USDS water. To determine whether USDS water’s nutritional deficiency plays a major role in limiting Aedes larval development, we repeated the larval survival experiment by adding 0.1 g Tetramin Tropical Flakes, the standard larval mosquito diet in insectaries, to the microcosms every 2 days. The number of dead and surviving larvae, pupae, and emergent adults was recorded daily.
Data analysis
All aquatic habitats that were positive or negative for the larvae of Ae. aegypti, Ae. albopictus and Cx. quinquefasciatus (the predominant species), were mapped using ArcGIS 10.7.1. The proportion of aquatic habitats positive for Ae. aegypti and Cx. quinquefasciatus was calculated for each habitat type from 2016 to 2019. To examine variation in Aedes and Culex larval positivity rate among different groups of larval habitats within the USDS, larval positivity rates for Ae. aegypti and Cx. quinquefasciatus were calculated for underground water retention vaults, underground catch basins/manholes, and underground pipelines/tunnels. The Chi-square test was used to examine the statistical significance. Culex quinquefasciatus was analyzed because it was the most common species, whereas Ae. albopictus was not included in the analysis due to insufficient number of Ae. albopictus positive habitats. To determine whether USDS water attracted or repelled oviposition of invasive Aedes mosquitoes, a pairwise t test was used to compare egg number in USDS water ovicups to flowerpot water ovicups for each Aedes species. Similarly, a pairwise t-test was used to test the effect of Cx. quinquefasciatus larvae on Aedes mosquito oviposition choice.
To examine the effect of water sources on egg hatching, the t-test was used to analyze the egg hatching rate. The analysis of larval life table study data focused on pupation rates and larval-to-pupal development times. The pupation rate was calculated as the proportion of first-instar larvae that molted into pupae. The effect of water sources and larval food supplementation on pupation rate was analyzed using non-parametric Wilcoxon test. The t-test was used to analyze the duration of larval-to-pupal development. Kaplan–Meier survival analysis was used to determine the effects of food supplementation and water source on larval development for each species, and the log-rank test was conducted to determine their statistical significance. All statistical analyses were performed using JMP software (JMP 14.2, SAS Institute Inc.).
Results
Ecological characterization of mosquito larval habitats
A total of 6,072 records with GPS coordinates, presence or absence of mosquito larvae and habitat type classification, were analyzed from 2016 to 2019 (Table S1). Within each sampling year, a majority (88.2% on average) of the larval habitats were collected once, 7.8% twice, and 4.1% more than 2 times. A total of 21 mosquito species across four genera were identified, including nine Aedes species, two Anopheles species, eight Culex species, and two Culiseta species (Table S2). Culex quinquefasciatus, Ae. aegypti, Cx. tarsalis, and Culiseta incidens were the dominant larval species, with occurrence constituting 65.5%, 18.3%, 17.1% and 12.8%, respectively, of the total 6072 larval collections. Of the six aquatic habitat types, containers, water features and USDS were the most common larval habitats for mosquitoes, comprising 31.4%, 24.8% and 17.4% respectively, of total aquatic habitats positive for any mosquito species.
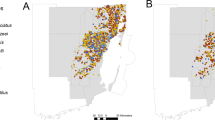
Aedes aegypti and Ae. albopictus, the two focal species of the present study, exhibited differences in abundance and spatial distribution (Fig. 1). Over the four-year study period, Ae. aegypti was found in 18.3% (1,111/6,072) and Ae. albopictus was detected in 0.4% (25/6,072) of the collections (χ2 = 1,145.3; d.f. = 1, P < 0.0001), clearly demonstrating Ae. aegypti was more abundant than Ae. albopictus in the study area. The distribution map of these two invasive Aedes species showed that Ae. aegypti was spreading rapidly over time (Fig. 1). The biggest expansion period occurred between 2017 and 2018. The total number of habitats positive for Ae. aegypti each year was 48 (9.2%; total number of habitats n = 520), 110 (5.2%; n = 2099), 570 (33.1%; n = 1718) and 383 (22.1%; n = 1735) between 2016 and 2019; and 13 (2.5%), 6 (0.3%), 5 (0.3%), and 1 (0.1%) for Ae. albopictus. The overall habitat positivity rate of Ae. aegypti among all aquatic habitats exhibited an increasing trend from 9.2% in 2016 to 22.1% in 2019.

Maps of mosquito larval sampling sites in Orange County, California, from 2016 to 2019, showing the distribution of Aedes aegypti and Ae. albopictus positive larval habitats. The maps were generated using ArcGIS 10.7.1 (ESRI, USA).
In terms of habitat utilization, containers were the most common habitats for Ae. aegypti (Fig. 2). Among all Ae. aegypti positive habitats (n = 1111), 80.1% were classified as containers, 13.8% water features, and only 4.1% USDS over the four-year study (Fig. 3). Although USDS sources constituted a small proportion of larval habitats for Ae. aegypti, habitat positivity rate in the USDS exhibited an increasing trend (Fig. 3). Other habitat types showing an increasing trend of Ae. aegypti larval positivity included containers, water features, and pools/spas, probably due to increasing abundance of Ae. aegypti in the study area (Fig. 3). Among Ae. albopictus positive habitats (n = 25), 88.0% were classified as containers, 8.0% water features and 4.0% marsh. In comparison, containers and USDS sources were consistently the main habitats of larval Cx. quinquefasciatus, constituting 28.3% and 22.0% of mosquito-positive habitats during the four-year study (Fig. 2). The habitat positivity rate of Cx. quinquefasciatus exhibited a decreasing trend in all six habitat types (Fig. 3).

Dynamics of habitat utilization of Aedes aegypti and Culex quinquefasciatus mosquitoes from 2016 to 2019 in Orange County, California.

Positivity rates of Aedes aegypti and Culex quinquefasciatus larvae in six types of aquatic habitats from 2016 to 2019 in Orange County, California. (A) Container; (B) Underground storm drain system; (C) Water feature; (D) Marsh; (E) Pool/Spa; and (F) Creek. Positivity rate was calculated as the proportion of larval habitats positive for Ae. aegypti or Cx. quinquefasciatus larvae among all habitats examined.
Because USDS habitats in our study represented several groups of underground aquatic habitats (underground water retention vaults, underground catch basins/manholes, and underground pipelines/tunnels), we examined whether these groups of aquatic habitats differed in mosquito larval positivity rate. A particularly interesting question is whether some USDS structures (e.g., catch basins) that are prone to collect debris and concentrate organic materials, which can serve as potential larval food sources, had a higher positivity rate for Aedes mosquito larvae. We found significant variations in Ae. aegypti positivity rate among the USDS habitat groups (χ2 = 15,6, d.f. = 2, P < 0.001), with the lowest positivity rate in the underground catch basins and manholes (0%), and the highest in the underground pipelines/tunnels (5.8%) (Table S3). In contrast, underground catch basins and manholes showed the highest positivity rate for Cx. quinquefasciatus (96.8%), whereas underground water retention vaults exhibited the lowest positivity rate (52.6%) (Table S3; χ2 = 63.7, d.f. = 2, P < 0.0001). Overall, these results demonstrated a low Ae. aegypti larval positivity rate but a high Cx. quinquefasciatus positivity rate in USDS aquatic habitats.
Laboratory oviposition preference
The total numbers of eggs laid by Ae. aegypti and Ae. albopictus were similar (42.3 vs. 38.5 per female, t = 0.42, d.f. = 1, P > 0.05; Fig. 4). Aedes albopictus mosquitoes preferred USDS water for egg laying over the flowerpot water (t = 2.88, d.f. = 1, P < 0.05), but Ae. aegypti exhibited no preference for any aquatic substrate (Fig. 4). Water from USDS did not attract or repel Ae. aegypti in their egg laying choice.

Oviposition substrate preference of Aedes aegypti and Ae. albopictus mosquitoes in a two-choice oviposition preference test with waters from underground storm drain systems and flowerpots.
In southern California, Culex larvae were present in the majority of containers and USDS habitats, and reached up to 80% habitat positivity, as shown in Fig. 3. It is thus interesting to examine whether the presence of Culex larvae attracted or repelled egg laying by the invasive Aedes species. We found that the presence of Cx. quinquefasciatus larvae did not attract or repel egg laying by Ae. aegypti in USDS or flowerpot water, and did not have a significant impact on the number of eggs laid in either type of oviposition substrate (Fig. S1A). A similar result was found for Ae. albopictus (Fig. S1B).
Egg hatching
The egg hatching rate of Ae. aegypti in USDS water was 37.8%, similar to the 42.5% hatching rate observed in the flowerpot water (Fig. 5). In contrast, the Ae. albopictus egg hatching rate was 1.0% in flowerpot water, significantly lower than the 18.0% hatching rate observed in USDS water. The low hatching rate of Ae. albopictus eggs in USDS water cannot be attributed to poor egg quality or other unsuitable environmental conditions because the same batch of Ae. albopictus eggs exhibited a 35.3% hatching rate in the deionized water under the same environmental conditions (Fig. 5).

Egg hatching rate of Aedes aegypti and Ae. albopictus in waters collected in summer 2019, from underground storm drain systems and flowerpots in Orange County, California. The error bar indicates standard error.
Larval survivorship
To determine whether USDS water was suitable for larval development of the invasive Aedes mosquitoes, larval-to-pupal survivorship was examined in microcosms under controlled insectary conditions. The first experiment was conducted in summer 2019 using field-collected USDS water and flowerpot water. Both Ae. aegypti and Ae. albopictus larvae exhibited high pupation rate (> 94%) and fast larva-to-pupa development (7–8 days) in the control (flowerpot water, Table 1), suggesting the flowerpot water fully met the nutritional needs of Aedes larvae and supported their development. On the other hand, Ae. aegypti and Ae. albopictus larvae exhibited a zero pupation rate in the USDS water (Table 1). Aedes aegypti larvae died within two days after they were placed in the microcosms with USDS water (Fig. 6A). Aedes albopictus larvae developed very slowly in the USDS water, and none successfully developed into pupae (Fig. 6B). The pupation rate pattern was very similar between Ae. aegypti and Ae. albopictus (Table 1), suggesting that summer USDS water was not suitable for larval development for the two invasive Aedes species.

Survival curve of Aedes aegypti and Ae. albopictus larvae in microcosms with waters collected in summer, 2019, from underground storm drain systems and flowerpots in Orange County, California. (A) Ae. aegypti with waters from underground storm drain systems and flowerpots without food supplementation; (B) Ae. albopictus with waters from underground storm drain systems and flowerpots without food supplementation; (C) Ae. aegypti with waters from underground storm drain systems and flowerpots with food supplementation; (D) Ae. albopictus with waters from underground storm drain systems and flowerpots with food supplementation. Chi-squared and P-value of Kaplan–Meier survival analysis log-rank test is shown. The error bar indicates standard error.
We conducted additional experiments to determine whether complete inhibition of larval development in invasive Aedes mosquitoes in the summer USDS water was due to the water’s nutritional deficiency or presence of toxins in water runoff from residential and commercial applications of insecticides during the summer season. We added Tetramin Tropical Flakes, the standard larval mosquito diet in insectaries, to the microcosms. Food supplementation decreased pupation rate of Ae. aegypti and Ae. albopictus in the control (flowerpot water, Table 1), as a consequence of habitat over-nutrition by food addition. In contrast, food supplementation led to a marginally significant increase in the larval pupation rate of Ae. aegypti from 0% without food to 12.0% after adding food (Z = 1.82, P = 0.069, Fig. 6C and Table 1), but did not change the pupation rate of Ae. albopictus—the pupation rate remained at 0% (Fig. 6D and Table 1). These results suggest that the inability of Ae. albopictus larvae to develop into pupae in the summer USDS water was not entirely due to nutritional deficiency, but could have been caused by pesticide residues to which Ae. albopictus larvae were particularly sensitive.
To test this possibility, we conducted the third microcosm experiment by using USDS water in winter when there was no larval habitat treatment. We found food supplementation increased the pupation rate of Ae. aegypti from 2 to 61% (Z = 2.22, P < 0.05; Table 2; Fig. S2). For Ae. albopictus, the addition of food increased the pupation rate from 2 to 87% (Z = 1.96, P < 0.05; Fig. S2). The significant contrast observed in Ae. albopictus pupation rates between summer and winter in the USDS water with food supplementation (0% vs. 87%; Z = 2.02, P < 0.05) suggests that the presence of toxins in the USDS water in summer was also one of the driving forces for complete inhibition of Ae. albopictus larval development.
Although summer USDS water completely inhibited the development of invasive Aedes larvae, Cx. quinquefasciatus larvae demonstrated a 15.0% pupation rate, significantly lower than the 81% pupation rate observed in the flowerpot water (Z = 2.19, P < 0.05; Table 1). Food supplementation significantly increased its pupation rate from 15.0% to 63.0% in the summer USDS water (Z = 1.96, P < 0.05; Table 1), suggesting that USDS summer water was not highly toxic to Cx. quinquefasciatus larvae. Rather, summer USDS water conferred nutrimental constraint to Cx. quinquefasciatus. In winter, nutrimental constraint of the USDS water to Cx. quinquefasciatus was more severe due to its lower pupation rate in the USDS water without the addition of food and larger difference in pupation rate induced by food supplementation (82.0% vs. 2.0%; Z = 2.23, P < 0.05; Table 2).
Discussion
This study was motivated by the question of the contribution of man-made USDS aquatic habitats to the productivity of invasive Ae. aegypti and Ae. albopictus pupae or adults in the urban environment of southern California. The USDS serves to drain run‐off water from homes, businesses, and streets in the urban environment, and also creates hard-to-treat stagnant water bodies in urban areas from thousands of kilometers of street gutters and underground pipes, numerous catch basins, and manhole access chambers10,25. Because stagnant water in the USDS in warm southern California has previously been shown to provide large numbers of suitable larval habitats predominated by Cx. quinquefasciatus10, we were interested in whether USDS offers productive larval habitats for newly invasive Ae. aegypti and Ae. albopictus to the region.
This study showed the rapid spread of invasive Ae. aegypti mosquitoes in Orange County, California, as evidenced by increasingly wider ranges of larval distribution and habitat positivity rates over a four-year period from 2016 to 2019. The largest expansion period occurred from 2017 to 2018. Although mosquito surveillance efforts were substantially enhanced since 2017, the observed Ae. aegypti spatial range expansion cannot be attributed to yearly differential sampling efforts: sampling efforts were comparable among 2017, 2018 and 2019. The distribution of Ae. albopictus was sporadic and limited in its abundance and spatial range: among the 6,072 aquatic habitats examined, Ae. albopictus larvae occurred in only 0.4% of aquatic habitats, whereas Ae. aegypti was found in 18.3% of the habitats. Invasive Aedes larvae were found mostly in containers, but 4.7% of the USDS habitats were positive for Ae. aegypti and 0% for Ae. albopictus. The USDS water did not attract or repel Ae. aegypti from laying eggs, but it attracted egg laying by Ae. albopictus, and was conducive for their egg hatching. Interestingly, USDS water inhibited larval development of Ae. aegypti and Ae. albopictus, particularly during the summer season when no larvae were able to develop into pupae.
Contrasting abundance and spatial distribution patterns between Ae. aegypti and Ae. albopictus in our study are remarkable given that both species were introduced/discovered in Orange County at the same time, 2015. The reason for this is not clear, but interspecific competition between Ae. albopictus and Ae. aegypti larvae is not likely a driving factor. Literature from other field studies in the eastern US, Asia, and South America suggest that Ae. albopictus larvae generally outcompete Ae. aegypti larvae, and Ae. albopictus has contributed to the declines in Ae. aegypti in southern North America26. A more plausible hypothesis is that the pattern of high Ae. aegypti abundance and occurrence in our study area may be determined by environmental conditions in the local area that help the survival of adult mosquitoes or eggs (e.g., desiccation) of Ae. aegypti but are less favorable to Ae. albopictus, rather than the result of competition among aquatic larvae. Further research is needed to combine population genetic analysis and field ecological studies to examine the founder effects and determine differential impacts of environmental conditions on Ae. aegypti and Ae. albopictus.
To circumvent the potential impact of mosquito colonization on their behavior and life history traits, the present study used Ae. aegypti and Ae. albopictus reared from field-collected eggs in the study area. To minimize the confounding effect of environmental factors such as temperature and predators, all studies were conducted in an insectary with climate regulation. Thus, although the experiments were not conducted in the field, the experimental setup enabled us to address the question of our interests. We found 4.1% of USDS habitats were positive for Ae. aegypti, and 0% for Ae. albopictus larvae, and USDS water was receptive to egg laying and permissive to egg hatching. However, USDS water was not permissive to invasive Aedes larval development and survival during the summer months when gravid females are available to lay eggs. Although winter USDS water allows larval development at a low population rate (~ 1.3–2%), the number of adult mosquitoes that can lay eggs is low at this time of the year. Therefore, the overall contribution of USDS aquatic habitats to the productivity of invasive Aedes mosquitoes is low. Other studies have previously reported the presence of invasive Aedes larvae in urban USDS habitats, e.g., Ae. albopictus larvae in urban stormwater catch basins and manhole chambers in China27, Ae. aegypti larvae in underground habitats in Brazil28, Puerto Rico29,30, Mexico31,32 and the United States33,34. One key unanswered question is what proportion of larvae can develop into pupae and emerge into adults. Mosquito surveillance programs that examine both larval and pupal abundance in the USDS water should address this question. Further, it is critical to determine whether the low success or complete failure of Aedes larval development into pupae in the USDS water as observed in the present study is unique to southern California or is a general phenomenon.
The mechanisms for the impact of USDS water on invasive Aedes larval development and survival are not clear. The zero pupation rate observed in Ae. aegypti and Ae. albopictus with summer USDS water can be attributed to nutrient deficiency, as well as poor water quality from low dissolved oxygen and nutrients, and abnormal electrical conductivity and salinity levels10 and perhaps residual insecticidal activity from pyrethroids (e.g., bifenthrin, a common household insecticide) found in Orange County’s urban runoff35. We tried to discern these possibilities by adding larval food to the microcosms and repeating the larval life table studies using USDS water from winter when there were few habitat treatments. A 12% pupation rate in Ae. aegypti and zero pupation in Ae. albopictus in microcosms with USDS water and food supplementation suggest that the USDS water was toxic to both species, with Ae. albopictus larvae suffering higher mortality. Water chemistry analyses by other investigators have demonstrated significant runoff in storm drain system from urban insecticide use in several counties in southern California36,37, suggesting pesticide runoff in USDS may be a widespread event. Considering the evidence of low pupation rates (~ 2%) for these two invasive Aedes species in the microcosm studies with winter USDS water when there was no active habitat treatment and thus low concertation of toxins, and drastically increased pupation rate after food supplementation, we conclude that nutrimental deficiency, toxins, and poor water quality in USDS summer water act together, leading to complete inhibition of larval development of Ae. aegypti and Ae. albopictus in the summer.
There were several limitations with the present study. First, we did not perform water chemistry analysis and thus could not pinpoint the precise reasons for the inhibition of invasive Aedes larval development in the USDS water. In addition to the reported low dissolved oxygen and nutrient content, abnormal electrical conductivity and salinity levels10, insecticides from many urban applications such as structural pest control, landscape maintenance, residential home and garden use, and mosquito control drain into the USDS12,35,38,39 and may fluctuate temporally and spatially as a consequence of dilutions by natural rainfall and irrigation, depending upon landscapes. Second, the microcosm experiments on the suitability of the USDS habitat for invasive Aedes egg laying, hatching and larval development used water mixtures from seven collection sites. The possibility of point-source effects on the observed results cannot be ruled out, in that a single sample that contained pesticide residue could be mixed into other samples, thereby affecting the results of the entire batch. However, the potential adverse effects from mixing water samples did not prevent development of Cx. quinquefasciatus. High levels of kdr-mediated pyrethroid resistance in Orange County populations of Cx. quinquefasciatus may have contributed to successful emergence of this mosquito in the microcosm experiments40. Future research will need to conduct water chemistry analysis to address this question. Third, we did not examine density dependence of larval survivorship in the microcosm studies. Nonetheless, our experiments using field mosquitoes under well-controlled conditions support the notion that USDS habitats are not major productive habitats for Ae. aegypti and Ae. albopictus in Orange County, California. The low quality of USDS water and slight attractiveness to ovipositing invasive Aedes may allow USDS habitats to act as ecological traps for invasive Aedes in southern California41.
In sum, this study examined the spread and aquatic habitat usage of two invasive Aedes species in the arid urban environment of southern California over a period of four years immediately after their introduction, and tested the suitability of USDS water for their egg laying, hatching, and larval development. Our results show rapid spread of Ae. aegypti in small containers in residential and commercial areas. Water from USDS habitats was receptive to egg laying and hatching, but completely inhibited successful development of invasive Aedes larvae to pupae. The notion that USDS habitats are currently not productive habitats for these invasive Aedes mosquitoes in southern California prompts including pupal productivity in the larval mosquito surveillance program. Finally, this work demonstrated that assessment of the impact of urbanization on the risk of vector-borne diseases requires careful examination of its impact on the environment and the ecology of disease vectors.
Data availability
The authors confirm that the data supporting the findings of this study are available within the paper and its supplementary information files.
References
Wilke, A. B. B. et al. Urbanization creates diverse aquatic habitats for immature mosquitoes in urban areas. Sci. Rep. 9, 15335. https://doi.org/10.1038/s41598-019-51787-5 (2019).
Neiderud, C.-J. How urbanization affects the epidemiology of emerging infectious diseases. Infect. Ecol. Epidemiol. 5, 27060 (2015).
Murdock, C. C., Evans, M. V., McClanahan, T. D., Miazgowicz, K. L. & Tesla, B. Fine-scale variation in microclimate across an urban landscape shapes variation in mosquito population dynamics and the potential of Aedes albopictus to transmit arboviral disease. PLoS Negl. Trop. Dis. 11, e0005640. https://doi.org/10.1371/journal.pntd.0005640 (2017).
Zahouli, J. B. et al. Urbanization is a main driver for the larval ecology of Aedes mosquitoes in arbovirus-endemic settings in south-eastern Côte d'Ivoire. PLoS Neglect. Trop. Diseases 11, e0005751 (2017).
Li, Y. et al. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl. Trop. Dis. 8, e3301. https://doi.org/10.1371/journal.pntd.0003301 (2014).
Benedict, M. Q., Levine, R. S., Hawley, W. A. & Lounibos, L. P. Spread of the tiger: global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis. 7, 76–85. https://doi.org/10.1089/vbz.2006.0562 (2007).
Powell, J. R. & Tabachnick, W. J. History of domestication and spread of Aedes aegypti–a review. Mem. Inst. Oswaldo Cruz 108(Suppl 1), 11–17. https://doi.org/10.1590/0074-0276130395 (2013).
Metzger, M. E., HardstoneYoshimizu, M., Padgett, K. A., Hu, R. & Kramer, V. L. Detection and establishment of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) Mosquitoes in California, 2011–2015. J. Med. Entomol. 54, 533–543. https://doi.org/10.1093/jme/tjw237 (2017).
Reisen, W. K., Meyer, R. P., Tempelis, C. H. & Spoehel, J. J. Mosquito abundance and bionomics in residential communities in Orange and Los Angeles Counties, California. J. Med. Entomol. 27, 356–367. https://doi.org/10.1093/jmedent/27.3.356 (1990).
Su, T., Webb, J. P., Meyer, R. P. & Mulla, M. S. Spatial and temporal distribution of mosquitoes in underground storm drain systems in Orange County, California. J. Vector Ecol. 28, 79–89 (2003).
Cummings RF, Semrow A, Nguyen K & T, M. Using a Remotely-Operated Vehicle (‘“Rover”’) for mosquito surveillance and control in underground storm drains. Proceedings and Papers of the Annual Conference California Mosquito and Vector Control Association 87, 187–189 (2019).
Harbison, J. E., Metzger, M. E., Walton, W. E. & Hu, R. Evaluation of factors for rapid development of Culex quinquefasciatus in belowground stormwater treatment devices. J. Vector Ecol. 34, 182–190. https://doi.org/10.1111/j.1948-7134.2009.00025.x (2009).
Kraemer, M. U. et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 4, e08347. https://doi.org/10.7554/eLife.08347 (2015).
Leta, S. et al. Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes albopictus. Int. J. Infect. Dis. 67, 25–35. https://doi.org/10.1016/j.ijid.2017.11.026 (2018).
Lounibos, L. P. & Kramer, L. D. Invasiveness of Aedes aegypti and Aedes albopictus and vectorial capacity for Chikungunya Virus. J. Infect. Dis. 214, S453–S458. https://doi.org/10.1093/infdis/jiw285 (2016).
Su, T. et al. Deployment and Fact Analysis of the In2Care(R) Mosquito Trap, A Novel Tool for Controlling Invasive Aedes Species. J. Am. Mosq. Control Assoc. 36, 167–174. https://doi.org/10.2987/20-6929.1 (2020).
Mian, L. S. et al. Residual activity of pyriproxyfen against mosquitoes in catch basins in Northwestern Riverside County, Southern California. J. Am. Mosq. Control Assoc. 36, 175–180. https://doi.org/10.2987/20-6963.1 (2020).
Kwan, J. L., Kluh, S., Madon, M. B. & Reisen, W. K. West Nile virus emergence and persistence in Los Angeles, California, 2003–2008. Am. J. Trop. Med. Hyg. 83, 400–412. https://doi.org/10.4269/ajtmh.2010.10-0076 (2010).
California Department of Public Health. Guidance for surveillance of and response to invasive Aedes mosquitoes and dengue, chikungunya, and Zika in California. , https://www.cdph.ca.gov/Programs/CID/DCDC/CDPH%20Document%20Library/InvasiveAedesSurveillanceandResponseinCA.pdf (2020).
Orange County Mosquito and Vector Control District. Annual Budget Fiscal Year 2019–2020. https://www.ocvector.org/files/fb678f362/AnnualBudget_1920.pdf. (2019).
Darsie, R. F. & Ward, R. A. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico 2nd edn. (University Press of Florida, 2005).
Meyer, R. P. & Durso, S. L. Identification of the mosquitoes of California. 1998. Mosquito and Vector Control Association of California Sacramento, CA.
Cummings, R. F. Design and use of a modified Reiter gravid mosquito trap for mosquito-borne encephalitis surveillance in Los Angeles County, California. Proc. Pap. Annu. Conf. California Mosq. Vector Control Assoc. 60, 170–176 (1992).
Cummings, R. et al. Fighting an Unpredictable Foe: An Overview of Two Consecutive Years of Record West Nile Virus Outbreaks, 2014–2015, in Orange County, California. Proc. Mosquito and Vector Control Association of California 84, 8 (2016).
Müller, G. C. et al. Control of Culex quinquefasciatus in a storm drain system in Florida using attractive toxic sugar baits. Med. Vet. Entomol. 24, 346–351 (2010).
Juliano, S. A. Species introduction and replacement among mosquitoes: interspecific resource competition or apparent competition?. Ecology 79, 255–268 (1998).
Gao, Q. et al. Aedes albopictus production in urban stormwater catch basins and manhole chambers of downtown Shanghai, China. PLoS ONE 13, e0201607. https://doi.org/10.1371/journal.pone.0201607 (2018).
Paploski, I. A. D. et al. Storm drains as larval development and adult resting sites for Aedes aegypti and Aedes albopictus in Salvador, Brazil. Parasites Vectors 9, 1–8 (2016).
Mackay, A. J., Amador, M., Diaz, A., Smith, J. & Barrera, R. Dynamics of Aedes aegypti and Culex quinquefasciatus in Septic Tanks. J. Am. Mosq. Control Assoc. 25, 409–416. https://doi.org/10.2987/09-5888.1 (2009).
Burke, R., Barrera, R., Lewis, M., Kluchinsky, T. & Claborn, D. Septic tanks as larval habitats for the mosquitoes Aedes aegypti and Culex quinquefasciatus in Playa-Playita, Puerto Rico. Med. Vet. Entomol. 24, 117–123. https://doi.org/10.1111/j.1365-2915.2010.00864.x (2010).
Manrique-Saide, P. et al. An assessment of the importance of subsurface catch basins for Aedes aegypti adult production during the dry season in a neighborhood of Merida, Mexico. J. Am. Mosq. Control Assoc. 29, 164–167. https://doi.org/10.2987/12-6320R.1 (2013).
Manrique-Saide, P. et al. Storm sewers as larval habitats for Aedes aegypti and Culex spp. in a neighborhood of Merida, Mexico. J. Am. Mosq. Control Assoc. 28, 255–257 (2012).
Rey, J. R., O’Meara, G. F., O’Connell, S. M. & Cutwa-Francis, M. M. Factors affecting mosquito production from stormwater drains and catch basins in two Florida cities. J. Vector Ecol. 31, 334–343. https://doi.org/10.3376/1081-1710(2006)31[334:fampfs]2.0.co;2 (2006).
Hribar, L. J. et al. Mosquito larvae (Culicidae) and other Diptera associated with containers, storm drains, and sewage treatment plants in the Florida Keys, Monroe County, Florida. Florida Entomol. 87, 199–203 (2004).
California Center for Urban Horticulture. Urban Run-Off Research. , https://ccuh.ucdavis.edu/urban-runoff-research (2020).
Budd, R., Wang, D., Ensminger, M. & Phillips, B. An evaluation of temporal and spatial trends of pyrethroid concentrations in California surface waters. Sci. Total Environ. 718, 137402. https://doi.org/10.1016/j.scitotenv.2020.137402 (2020).
Wolfand, J. M. et al. Occurrence of urban-use pesticides and management with enhanced stormwater control measures at the watershed scale. Environ. Sci. Technol. 53, 3634–3644 (2019).
Harbison, J. E. & Metzger, M. E. H2O: The Fundamental Link Between Stormwater Management and Mosquito Control Agencies, https://www.stormh2o.com/bmps/article/13009683/h2o-the-fundamental-link-between-stormwater-management-and-mosquito-control-agencies (2014).
Amweg, E. L., Weston, D. P., You, J. & Lydy, M. J. Pyrethroid insecticides and sediment toxicity in urban creeks from California and Tennessee. Environ. Sci. Technol. 40, 1700–1706. https://doi.org/10.1021/es051407c (2006).
Yoshimizu, M. H., Padgett, K. & Kramer, V. Surveillance of a kdr Resistance Mutation in Culex pipiens (Diptera: Culicidae) and Culex quinquefasciatus in California. J. Med. Entomol. 57, 645–648. https://doi.org/10.1093/jme/tjz208 (2020).
Gardner, A. M., Muturi, E. J. & Allan, B. F. Discovery and exploitation of a natural ecological trap for a mosquito disease vector. Proc. R. Soc. B 285, 20181962 (2018).
Acknowledgements
We thank San Gabriel Valley Mosquito and Vector Control District for their assistance in collecting Aedes albopictus eggs in the field, and Orange County Mosquito and Vector Control District for assisting with collecting habitat water and mosquitoes. We thank the three reviewers for their constructive comments. This work was supported by the Pacific Southwest Center for Excellence in Vector-Borne Diseases from funds provided by the U.S. Centers for Disease Control and Prevention (Cooperative Agreement U01CK000516), and the UCI Chancellor’s Fellow Fund. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
X.W., T.M., R.C., M.D., and G.Y. conceived the study. X.W., Y. L., S. O. and A. S. carried out the experiments. K.N. managed the larval database. X.W., G. Z., D. Z., Y. L., J. B. and G. Y. conducted the data analysis and data interpretation. X.W., R.C., and G.Y. drafted the manuscript. All authors contributed to the critical revision of the manuscript for important intellectual content.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, X., Zhou, G., Zhong, D. et al. Impact of underground storm drain systems on larval ecology of Culex and Aedes species in urban environments of Southern California. Sci Rep 11, 12667 (2021). https://doi.org/10.1038/s41598-021-92190-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-92190-3
This article is cited by
-
Defining the roles of local precipitation and anthropogenic water sources in driving the abundance of Aedes aegypti, an emerging disease vector in urban, arid landscapes
Scientific Reports (2024)
-
Climate, landscape, and life history jointly predict multidecadal community mosquito phenology
Scientific Reports (2023)
-
Field investigation combined with modeling uncovers the ecological heterogeneity of Aedes albopictus habitats for strategically improving systematic management during urbanization
Parasites & Vectors (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
