
Abstract
Bovine tuberculosis (bTB) is an important animal health and economic problem for the cattle industry and a potential zoonotic threat. Wild badgers (Meles meles) play a role on its epidemiology in some areas of high prevalence in cattle, particularly in the UK and Republic of Ireland and increasingly in parts of mainland Europe. However, little is known about the involvement of badgers in areas on the spatial edge of the cattle epidemic, where increasing prevalence in cattle is seen. Here we report the findings of a study of found-dead (mainly road-killed) badgers in six counties on the edge of the English epidemic of bTB in cattle. The overall prevalence of Mycobacterium tuberculosis complex (MTC) infection detected in the study area was 51/610 (8.3%, 95% CI 6.4–11%) with the county-level prevalence ranging from 15 to 4–5%. The MTC spoligotypes of recovered from badgers and cattle varied: in the northern part of the study area spoligotype SB0129 predominated in both cattle and badgers, but elsewhere there was a much wider range of spoligotypes found in badgers than in cattle, in which infection was mostly with the regional cattle spoligotype. The low prevalence of MTC in badgers in much of the study area, and, relative to in cattle, the lower density of sampling, make firm conclusions difficult to draw. However, with the exception of Cheshire (north-west of the study area), little evidence was found to link the expansion of the bTB epidemic in cattle in England to widespread badger infection.
Similar content being viewed by others


Introduction
Bovine tuberculosis (bTB), primarily caused by Mycobacterium bovis, is an important animal health and economic problem for the British cattle industry, and an important zoonotic infection globally, particularly where bovine milk, the main source of human infection, is not treated1. M. bovis, is a member of the M. tuberculosis complex (MTC) group of bacteria and predominantly infects cattle, but has a wide host range that includes badgers (Meles meles), other wildlife, camelids, goats, humans and some companion animals. Badgers are reported to act as one of the ‘reservoirs’ of M. bovis bacilli, at least in some areas2,3,4,5. M. tuberculosis, the overwhelmingly most frequent cause of human TB and M. microti, which mainly circulates in field voles (Microtus agrestis)6, are also found in Great Britain.
The incidence of TB in cattle in England and Wales has been rising since the mid-1980s, and the area in which it is considered endemic has gradually expanded to cover most of the south west of England and south Wales, the West Midlands and parts of mid-Wales2. Although usually described as a single spreading epidemic, the regional distributions of different spoligotypes of M. bovis might suggest a series of regional epidemics7. The government’s strategy for achieving ‘officially TB-free (OTF)’ status for England is based on geographic areas of risk, with different surveillance and control measures applied to different areas: the Low Risk Area (LRA), the Edge Area and the High Risk Area (HRA). The Edge Area is a boundary of intermediate TB incidence in cattle, situated between the HRA and LRA and, at the time of this study, comprised eleven counties (or part-counties), where surveillance suggested that the historically low incidence of TB in cattle was increasing.
The control of TB in cattle in many parts of the world is complicated by the involvement of wildlife species, such as deer8 and, in England, Wales, the island of Ireland and increasingly in some other European countries, badgers (Meles meles)2,3. It is unclear whether or not badgers are technically a reservoir host in the HRA (i.e. not only a source of infection to cattle—the ‘target’ host—but able to maintain infection without reintroduction from cattle)2,9, and even less is known about the role of badgers in cattle infection in the Edge Area. Thus the expansion of the cattle epidemic in the Edge Area could be the result of cattle-to-cattle, badger-to-badger, cross species transmission, or, indeed, other even less well-understood transmission routes such as through soil and farm waste10,11,12,13; and the relative contributions of each potential transmission route, or if they differ between regions in the Edge Area, are unknown.
Road traffic and ‘found dead’ surveys have previously been used to study several wildlife diseases, including TB in badgers in England and Wales. Following the discovery of TB in badgers in the 1970s and 1980s14,15,16,17, several surveys of road-killed badgers for TB were undertaken in different areas of England and Wales (reviewed in Krebs et al.18). More recently, surveys of ‘found dead’ badgers in Northern Ireland and Wales have been published19,20,21, and also a study of road-killed badgers in Cheshire, NW England, in 201422, a county that until 2018 straddled the Edge Area and the HRA. In each study, infection in badgers was found largely, although by no means totally, in areas in which TB in cattle was, at the time, most prevalent. Government agencies have also undertaken occasional small road-kill surveys around particular cattle outbreaks and found infected badgers, but we are not aware of these data having been published.
Ongoing long-term studies of badger TB in Gloucestershire (SW England)23 in the HRA, show long term persistence of TB in badger setts, and recent badger surveys around an isolated hotspot of TB in cattle in Cumbria (NW England), suggested that infection of imported origin in cattle was circulating in both badgers and cattle24. While these studies provide information about M. bovis in badgers and cattle in specific locations, there remains a lack of research into the relationship between M. bovis in cattle and badgers in other risk areas, particularly where the epidemic in cattle is spreading. Our study contributes to this knowledge gap.
Here we describe a study of road-killed badgers in northern Edge Area counties of England (Fig. 1; Cheshire, Derbyshire, Nottinghamshire, Leicestershire, Warwickshire and Northamptonshire), carried out in 2016–2017, and compare those findings with reported outbreaks of TB in cattle over the same area and time, collected by routine, government-regulated surveillance.

Map of the counties where RTA badgers were collected for testing.
Results
Badger carcasses
Overall, 610 badger carcasses suitable for necropsy and sampling, were collected from the study area (Fig. 2). Of these, 246 (40%) were collected by farmers and farming groups, 219 (36%) by badger or conservation groups and 145 (24%) by project staff, veterinarians, local authorities or other groups. These proportions differed by county (SI 1). The age and sex of the badger carcasses collected, when recorded (> 98% of carcasses), was overall 53% male and 46% female, and the age ratio 76% adult : 24% juvenile, with 13 (3%) of the adults classified as ‘old’ on the basis of pelage and tooth wear.

(A) the geographic distribution of badger carcasses sampled with those culture positive for confirmed Mycobacterium tuberculosis complex (MTC) in red and those negative in tan. (Original map generated in R-Googlemaps).
The availability of badger carcasses varied over time, with a peak in carcass collection in February 2017 (SI 1), and the number collected was capped at 100 badgers per county.
Mycobacterium tuberculosis complex (MTC) in badgers
Overall, 51/610 (8.3%, 95% CI 6.4–11%) badgers were positive for MTC by culture and IS6110 PCR. The prevalence of infection in males was approximately twice that in females (11% c.f. 5%, chi-square 6.019 p = 0.0142, odds ratio 2.16, relative risk 2.13). The prevalence did not vary significantly with the age of the badger, the time of year or the type of collector (‘farmer’, ‘wildlife’ group or ‘other’).
At post mortem examination, only one badger, which was MTC culture positive, had obvious, widespread lesions of TB, and another had atypical gross lesions from which MTC was isolated. The prevalence of MTC in the badgers varied between counties, such that Cheshire, Leicestershire and Warwickshire had higher prevalence than Derbyshire, Nottinghamshire and Northamptonshire (χ2 = 12.7, p = 0.026).
Spoligotyping of badger MTC revealed a variety of spoligotypes, including those such as SB0129, SB0263, SB0272 and SB0140, frequently found associated with cattle infection in England25 but also a range of novel and rare spoligotypes (Figs. 2 and 3, and Supplementary Information). The most common spoligotype of M. bovis found in badgers (SB0129), was found particularly in the north of the study area, but also sporadically elsewhere. Other major spoligotypes of M. bovis isolated from cattle were found more sporadically in badgers.

Overall distribution of spoligotypes of MTC isolates from northern EDGE Area cattle (circles) and badgers (squares) key: grey SB0129, red SB0263, blue SB0140, olive green SB0272, gold SB1016, orange SB1016, chocolate SB1414, green SB1512, yellow rare/untypable, purple M. microti. Cattle breakdowns with no known spoligotype are shown as white circles; these are usually breakdowns defined by skin tests but without MTC being cultured for typing. (Original map generated in R version 4.1.0 and package RgoogleMaps40).
Mycobacterium tuberculosis complex bacteria in cattle
The results of farm-level surveillance testing of cattle for TB over the same period and area as the badger study are shown in Fig. 3. In total, there were 730,654 individual tests undertaken at 5,765 cattle holdings, and reported herd sizes ranged from 1 to 2,900 animals (median 62). There were 5227 positive animals, giving rise to 660 herd ‘breakdowns’ or outbreaks (11% of holdings) over the study period.
In the vast majority (91%) of herd breakdowns, the index case was detected by skin test during routine surveillance testing. When M. bovis was isolated from cattle, almost all (98.3%) of the spoligotypes from cattle were those previously identified in cattle in England. Each had a distinctive geographical range (Fig. 3) but some were also found sporadically in other regions. For example, the predominant spoligotype found in cattle during 2016–2017 was SB0129, particularly in the northern part of the study area, but the same spoligotype was also more sporadically found in other parts of the study area. Spoligotype SB0272 was the predominant spoligotype found in the southern part of the study areas, SB0263 was found sporadically throughout the study area, but mainly in western Cheshire.
Comparison of MTC prevalence, spoligotypes and distributions in cattle and badgers
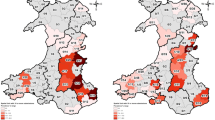
Spatially co-incident prevalence of MTC in badgers and on cattle farms was examined using a Bayesian bivariate logistic common-components geostatistical model26. Formal comparison of spoligotype distributions was only possible for SB0129 as the other spoligotypes found in badgers were too rare and sparsely distributed to be compared with those found in cattle. The spatial prevalence of both MTC and SB0129 was modelled in the badger and cattle (farm) populations using a combination of spatially-continuous risk surfaces (‘common’, ‘farm’, and ‘badger’), as well as a covariate adjusting for observation type (badger or farm). This enabled smoothed MTC and SB0129 risk surfaces to be fitted, describing how prevalence in farms and badgers varied in common over space, and also any additional departures from this common surface which might indicate spatial segregation of infection by observation type. The results, in terms of the posterior mean estimate of the spatial odds ratio for being MTC or SB0129 positive are shown in Fig. 4 (MTC regardless of spoligotype) and Fig. 5 (for SB0129). Only those regions where the posterior probability of the odds ratio is either greater or lower than 1 is at least 0.95 are plotted. For MTC (regardless of spoligotype), common high prevalence areas were seen in east Cheshire, in south west Derbyshire, on the border with Staffordshire, and in south Warwickshire, with areas of lower combined prevalence in east Cheshire, northern Derbyshire and central Northamptonshire. Similar areas of high prevalence in cattle were observed in east Cheshire, west Derbyshire and south Warwickshire, but no evidence of badger-specific foci was seen. Spatial comparison of SB0129 demonstrated obvious spatial foci of increased odds in east Cheshire and Derbyshire’ border with Staffordshire, but no pronounced evidence of additional cattle or badger-specific spatial foci was found for SB0129 (Fig. 5).

Mean posterior spatial odds ratio of MTC in the Edge region, highlighting regions of high posterior confidence that the spatial odds is above or below 1 (Pr(OR > 1) > 0.5 or Pr(OR < 1) > 0.5). For example, the common map indicates high prevalence foci in badgers and cattle in east, but not west Cheshire, in Derbyshire on the border with Staffordshire, but not in north-west Derbyshire, and in south Warwickshire. Areas of high prevalence for farms are seen in East Cheshire on the border with Staffordshire and in south Warwickshire. No areas of additional high prevalence are seen for badgers. Increase in red represents increased risk, and blue indicates decreased risk. Map tiles by Stamen Design, under CC BY 3.0. Data by OpenStreetMap, under ODbL.

Mean posterior spatial odds ratio of SB0129 in the Edge region, highlighting regions of high posterior confidence that the spatial odds is above or below 1 (Pr(OR > 1) > 0.5 or Pr(OR < 1) > 0.5). The common map indicates high prevalence foci in Cheshire and Derbyshire and east Leicestershire. A small area of additional high prevalence for farms is seen in south west Derbyshire, bordering Staffordshire. No areas of additional high prevalence are seen for badgers. Increase in red represents increased risk. Map tiles by Stamen Design, under CC BY 3.0. Data by OpenStreetMap, under ODbL.
Discussion
Stakeholders collected the badger carcasses and recorded their locations in this study, as described previously22. Although there was variation between counties, the type of stakeholder (farming group, wildlife group or other) was not overall associated with differences in the sex and age of badger carcasses collected or the prevalence of infection with MTC. Furthermore, the sex ratio, and proportions of animals by age were similar to those described in previous studies of live badgers27,28. Thus, while using road-killed badgers might involve biases in terms of space and time, there was no evidence that different stakeholder groups influenced the outcome of this study in terms of badger demographics or infection prevalence. As the density of badgers is not known in the Edge Area, and 100 carcasses were collected from each county (with counties of different areas), it is impossible to determine if heterogeneities in sampling density reflected heterogeneities in badger population density, stakeholder engagement or roads and road usage.
Overall, 51/610 (8.3%, 95% CI 6.4–11%) carcasses were culture positive and confirmed by PCR as MTC, a figure similar to that found for Wales in 2014–201621. Previous studies found that the prevalence of infection in badgers was higher in adults than cubs, and in males than females, although in some cases these differences were not statistically significant16,20,21,29. This study found no significant difference in prevalence by age, although the numbers of, and prevalence in, younger badgers was small; but that the prevalence of infection in males was approximately twice that in females. It has been suggested this difference might reflect the larger home ranges of, and more territorial conflicts between, males compared to females, both of which could increase transmission20,21.
Most infected badgers had no macroscopic lesions suggestive of tuberculosis on post-mortem examination. The proportion of infected badgers with no obvious lesions (96%) was similar to that found previously in Cheshire22, but higher than that found in most studies of badgers in areas where infection is endemic in both cattle and badgers. The majority of positive cultures were from the ‘head and neck’, ‘thorax’ and ‘carcass’ pools of lymph nodes (27%, 27% and 21%, respectively). This is also similar to the findings of22, but contrasts with many previous studies, which found that isolation of M. bovis was most frequently from lung and thoracic lymph nodes30,31. These differences in infection sites and lesions may reflect differences sources of infection, in transmission routes and dose, or in the pathogenicity of the different genotypes of MTC found in the badgers across this study. It might also reflect differences in how carcasses were collected between these and other studies. Although it is often assumed that transmission of M. bovis among badgers is by the respiratory route, and requires lesions, transmission via bites is also thought to be important. It is conceivable that transmission via saliva and respiratory secretions through bites, grooming and other close contact, or, indeed, contaminated soil and bedding, associated with pharyngeal lymphoid infection is more likely, without the need for lesions. Indeed, such secretions might also enable shedding in faeces, with or without enteric infection.
There was variation in the frequency of infected badger carcasses between counties, with Cheshire, Leicestershire and Warwickshire having headline infection rates (10–15%) higher than Derbyshire, Nottinghamshire and Northamptonshire (4–5%). In Cheshire, 15/104 (13.5%, 95% CI 8–21%) badgers were confirmed MTC-infected, a prevalence not significantly different from that found in 201422. There are no recent studies of MTC prevalence in badgers across the rest of the study area with which the results can be compared. Spoligotyping was performed rether than whole genome sequencing to enable comparison with historical cattle data. The most common spoligotype in MTC isolated from badgers was SB0129, particularly in the northern part of the study area where it was also the most frequent spoligotype found in cattle. Elsewhere, a wide range of spoligotypes were found in badgers, whereas in cattle the predominant spoligotypes were those expected if those infections resulted from expansion of regional cattle spoligotypes from neighbouring High Risk Area counties. The majority of cattle outbreaks in Leicestershire during the study were not confirmed by culture and so the spoligotype(s) responsible could not be identified. Subsequent surveillance reports (APHA, 2019), suggest that cattle outbreaks when spoligotyped remain largely of SB0129 with some suggestion of SB0272 expanding across the border from Warwickshire.
More formal analysis of the prevalence of MTC and spoligotypes in both cattle and badgers across the study area found no pronounced evidence of either MTC or SB0129 -specific spatial foci beyond those in eastern Cheshire and the Derbyshire border with Staffordshire, where SB0129 was and remains the predominant spoligotype found in cattle32. The approach used is robust to differing sampling densities of either farms or badgers across space, necessary with the inevitable variation in population density and sampling density. However, the diversity of spoligotypes found in badgers combined with the relatively low numbers of MTC-positive badgers in much of the study area, made further quantitative analysis difficult.
Thus overall, it appears that different transmission patterns may apply to different areas of the expanding edge of the TB epidemic in cattle in England. In the northwest of the study area, in Cheshire, there was evidence of persistence of infection in both badgers and cattle with the same spoligotype (SB0129), which together with an earlier study22 might suggest that infection in the two species is part of the same epidemic. Further east however, in Derbyshire and Nottinghamshire, there appeared to be an expanding epidemic of SB0129 in cattle, with little evidence of widespread infection in badgers, although few badgers were collected in the southwest corner of Derbyshire where TB has previously been described in badgers33. Where MTC-infected badgers were found, they were usually also infected with spoligotype SB0129. Further south in the study area, the prevalence of infection with MTC varied in both badgers and cattle, but whereas in cattle spoligotyping suggested an expanding epidemic from neighbouring High Risk Area counties, along with occasional long distance movements, a diversity of spoligotypes, including M. microti and occasional spoligotypes usually found in distant regions, was found in badgers. With the exception of in Cheshire, therefore, this study found little evidence to directly link the expanding epidemic in cattle in England to widespread badger infection. However, longer term and longitudinal studies will be required to understand better whether the expanding epidemics of regional spoligotypes in cattle are followed or led by similar dynamical changes in the prevalence of infection in badgers.
Materials and methods
Badger and sample collection
All badgers used in this study were found dead. The majority of badgers collected in this study were collected from road-sides and had lesions compatible with vehicle collisions. A small number were found dead in fields or farm buildings, some of which also had lesions compatible with vehicle collisions. The approach used was that developed for a previous study in Cheshire in 201422: stakeholder groups collected carcasses, recorded where they were found, most often using smart phones, and the carcasses were then brought to the veterinary pathology facilities at either the University of Nottingham or University of Liverpool for necropsy and sampling. Stakeholders included farmers, veterinary practices, wildlife groups, local authorities and businesses as well as university staff. All experimental protocols were approved by the Defra and University of Nottingham.
Badgers were collected from six English counties in the northern edge of cattle TB epidemic (Cheshire, Derbyshire, Nottinghamshire, Leicestershire, Warwickshire and Northamptonshire) over a 16 month period from 2016 to 2017.
Where possible badger carcasses were stored in dry, cool areas or refrigerated. With two exceptions, frozen badgers were not included in the study. Carcass collection and necropsy occurred within 72 h of collection, and obviously autolytic or heavily damaged carcasses were not processed due to the likelihood of contamination. Each badger’s sex, weight and age (adult or juvenile, based on teeth, size and pelage) were recorded.
Lymph nodes and visible ‘TB-like’ lesions were sampled for culture. In addition to lung samples, four pools of lymph nodes were sampled: a ‘thoracic pool’ of bronchial and mediastinal lymph nodes, an ‘abdominal pool’ comprising hepatic and mesenteric lymph nodes, a ‘head and neck’ pool of parotid, mandibular, retropharyngeal and cervical lymph nodes, and a ‘carcass pool’ of prescapular, axillary, and superficial inguinal lymph nodes. If possible lesions were observed, each sample of lesion material was processed separately. Pooled tissues were stored at 4 °C for up to 48 h before processing. Individual tissue samples were stored at − 80 °C in case needed for further study.
Tissue processing
Tissue samples for microbiological culture were processed in a Containment Level 3 (CL3) facility at the University of Nottingham, Sutton Bonington Campus. Samples were processed according to22. Briefly tissue pools or lesions were gently ground with sterile sand and 2 ml phosphate buffer saline (PBS; Dulbecco A, Oxoid). Samples were mixed with an equal amount of 5% oxalic acid (Fisher Scientific) and incubated at room temperature for 10 min to reduce non-mycobacterial contamination. The decontaminated pools (200 µl) were inoculated onto Stonebrink Selective agar supplemented with PACT (BD Diagnostics) and Middlebrook 7H11 agar slopes supplemented with PANTA (BD Diagnostics), and incubated at 37 °C for a minimum of 12 weeks. Cultures were examined approximately weekly for the appearance of colonies suggestive of mycobacteria.
Characterisation of mycobacteria
Characteristic and possible colonies were picked from slopes after 12 weeks of incubation. Cells were heat killed (80 °C for 30 min) and DNA was extracted by crude heat lysis, in which heat-killed colonies were frozen and heated to 95 °C for 5 min in sterile distilled water (SDW), centrifuged (13,000 × g; 3 min) and the supernatant used as template DNA for subsequent PCR.
Isolates were screened for the MTB signature genetic element, IS6110 by PCR34. For some colonies, the partial 16 s rRNA gene was amplified, sequenced (Source Bioscience, UK) and analysed to determine whether the colonies were part of the MTB complex or not35. DNA from confirmed members of the MTB complex were further interrogated by microarray-based spoligotyping (Alere Technology) according to36.
Cattle data
Cattle testing data were obtained from APHA’s surveillance programme. At the time of the study, different cattle testing regimens were used in different counties. In HRA and Edge counties, cattle were routinely tested at herd level six-monthly or annually, with additional testing if infection was detected. Furthermore, some holdings had groups of cattle tested due to movements or because of epidemiological links to outbreaks detected elsewhere. Some TB-infected cattle were also detected during routine post-mortem inspection in the abattoir. As described in22, the original data sets from APHA included cattle holding location, the results of each test (including spoligotype if known), the official ‘breakdown identifier’ and start date, and the number of cattle on the holding and tested on each occasion. For the purposes of this study, all test results in the study period were combined for each site, and TB-positive herds were defined as those with at least one animal having tested positive using the ‘single intradermal comparative cervical tuberculin’ (SICCT) test, at least one animal with two consecutive inconclusive skin test results, positive interferon-gamma blood tests, or if lesions which were culture positive were found at the abattoir.
Statistical analyses
The prevalence was estimated among the sampled badgers both overall and at a county level, as in previous studies19,20 on the assumption that the carcasses collected were representative of the overall population.
To detect evidence for co-occurrence of MTC positive badgers and SB0129 herd breakdowns, a bivariate Bayesian logistic common-components geostatistical model was used37. Observations consisted of a set of geographically located farms and badgers, with infection (breakdown) status recorded as a binary outcome variable (positive or negative), and observation type (farm or badger) as an explanatory variable. The outcome was modelled for each observation, yi, as a Bernoulli random variable with probability pi such that
where xi is the geographical location of observation i, and \(\alpha\) and \(\beta\) represent respectively the intercept and the log odds ratio for disease in badgers versus farms. The model makes use of three spatial Gaussian Processes, \({S}_{c}\left({x}_{i}\right)\), \({S}_{b}\left({x}_{i}\right)\), and \({S}_{f}\left({x}_{i}\right)\), characterised by a covariance function \({\Sigma }_{ijz}\) between the locations of observations i and j for “common”, “badger”, and “farm” spatial correlation components respectively. In each case, a Matérn correlation structure is assumed with sill and scale parameters \({\sigma }_{z}\) and \({\phi }_{z}\).48 Here, \({S}_{c}\left({x}_{i}\right)\) represents a disease risk surface common to both farms and badgers that allows disease prevalence to vary with space. Then, \({S}_{b}\left({x}_{i}\right)\) and \({S}_{f}\left({x}_{i}\right)\) represent respectively the additional spatial risk for badgers and farms respectively.
In a Bayesian context, prior distributions were assigned to the parameters as shown in Table 1.
In contrast with previous work in Cheshire22, the volume of data analysed here (5522 farm and 565 badger locations) precluded a naïve implementation of the three Gaussian processes. Instead, a low-rank approximation was used in which a number of “knot” points were chosen to represent the Gaussian process in 2-dimensional space, and the resulting representation projected onto the observation points. Here, 600 and 200 points were chosen for farms and badgers respectively by 2-dimensional k-means clustering38.
The model was implemented in Python 3.6. using the PyMC3 3.7 package39, with the No-U-Turn Sampler (NUTS) used to draw samples from the joint posterior distribution. The sampler was run for 20,000 samples, discarding the initial 1500 as tuning and burn-in. The convergence of 4 MCMC chains was inspected visually and found to be satisfactory. Traceplots for these runs are shown in Supplementary Fig. 1 together with Gelman-Rubin statistics demonstrating convergence (\(\widehat{R}\) in all cases).
To detect co-regionalisation of infection in both badgers and farms, the Bayesian predictive distribution of the Gaussian Process was evaluated on a fine grid over the study area38. To summarise each predictive distribution, both the mean posterior value of the Gaussian process, and the probability that the posterior distribution at each grid point was greater or less than zero, were calculated. Since the Gaussian process represented the spatial log odds ratio, this is equivalent to calculating the posterior probability of the spatial odds ratio being greater or less than 1, i.e. Pr(OR > 1) and Pr(OR < 1) respectively, noting that Pr(OR > 1) = 1 – Pr(OR < 1). To present the results, we plot maps of the mean posterior spatial odds ratio for pixels where Pr(OR > 1) > 0.95 or Pr(OR < 1) > 0.95. Thus our maps highlight regions where there was good evidence that the odds ratio is significantly different to 1 (i.e. no spatial effect).
References
Doran, P., Carson, J., Costello, E. & More, S. J. An outbreak of tuberculosis affecting cattle and people on an Irish dairy farm, following the consumption of raw milk. Irish Vet. J. 62, 390–397 (2009).
Allen, A. R., Skuce, R. A. & Byrne, A. W. Bovine tuberculosis in Britain and Ireland—a perfect storm? The confluence of potential ecological and epidemiological impediments to controlling a chronic infectious disease. Front. Vet. Sci. 5, 109. https://doi.org/10.3389/fvets.2018.00109 (2018).
Bouchez-Zacria, M., Courcoul, A. & Durand, B. The distribution of bovine tuberculosis in cattle farms is linked to cattle trade and badger-mediated contact networks in south-western France. Front. Vet. Sci. 2007–2015, 5. https://doi.org/10.3389/fvets.2018.00173 (2018).
Bailey, S. S., Crawshaw, T. R., Smith, N. H. & Palgrave, C. J. Mycobacterium bovis infection in domestic pigs in Great Britain. Vet. J. 198, 391–397. https://doi.org/10.1016/j.tvjl.2013.08.035 (2013).
Corner, L. A., Murphy, D. & Gormley, E. Mycobacterium bovis infection in the Eurasian badger (Meles meles): The disease, pathogenesis, epidemiology and control. J. Comp. Pathol. 144, 1–24. https://doi.org/10.1016/j.jcpa.2010.10.003 (2011).
Kipar, A. et al. Mycobacterium microti tuberculosis in its maintenance host, the field vole (Microtus agrestis): Characterization of the disease and possible routes of transmission. Vet. Pathol. 51, 903–914. https://doi.org/10.1177/0300985813513040 (2014).
Davidson, J. A. et al. Epidemiology of mycobacterium bovis disease in humans in England, Wales, and Northern Ireland, 2002–2014. Emerg. Infect. Dis. 23, 377–386. https://doi.org/10.3201/eid2303.161408 (2017).
Glaser, L. et al. Descriptive epidemiology and whole genome sequencing analysis for an outbreak of bovine tuberculosis in beef cattle and white-tailed deer in northwestern minnesota. PLoS ONE 11, e0145735. https://doi.org/10.1371/journal.pone.0145735 (2016).
Haydon, D. T., Cleaveland, S., Taylor, L. H. & Laurenson, M. K. Identifying reservoirs of infection: A conceptual and practical challenge. Emerg. Infect. Dis. 8, 1468–1473. https://doi.org/10.3201/eid0812.010317 (2002).
Milne, G. et al. Bovine tuberculosis breakdown duration in cattle herds: An investigation of herd, host, pathogen and wildlife risk factors. PeerJ 8, 2 (2020).
Barbier, E., Rochelet, M., Gal, L., Boschiroli, M. L. & Hartmann, A. Impact of temperature and soil type on Mycobacterium bovis survival in the environment. PLoS ONE 12, 2 (2017).
Barasona, J. A. et al. Environmental presence of mycobacterium tuberculosis complex in aggregation points at the wildlife/livestock interface. Transbound Emerg. Dis. 64, 1148–1158 (2017).
Sanchez-Hidalgo, A., Obregon-Henao, A., Wheat, W. H., Jackson, M. & Gonzalez-Juarrero, M. Mycobacterium bovis hosted by free-living-amoebae permits their long-term persistence survival outside of host mammalian cells and remain capable of transmitting disease to mice. Environ. Microbiol. 19, 4010–4021 (2017).
Muirhead, R. H. & Gallagher, J. Tuberculosis in wild badgers in gloucestershire—epidemiology. Vet. Rec. 95, 552–555 (1974).
Gallagher, J., Muirhead, R. H. & Burn, K. J. Tuberculosis in wild badgers. Vet. Rec. 98, 179–179 (1976).
Cheeseman, C. L., Jones, G. W., Gallagher, J. & Mallinson, P. J. The population-structure, density and prevalence of tuberculosis (Mycobacterium-Bovis) in badgers (Meles-Meles) from 4 areas in southwest England. J. Appl. Ecol. 18, 795–804. https://doi.org/10.2307/2402370 (1981).
Anderson, R. M. & Trewhella, W. Population dynamics of the badger (Meles meles) and the epidemiology of bovine tuberculosis (Mycobacterium bovis). Philos. Trans. R. Soc. Lond. B Biol. Sci. 310, 327–381. https://doi.org/10.1098/rstb.1985.0123 (1985).
Krebs, J., Anderson, R, Clutton-Brock, T, Morrison, I, Young, D & Donnelley, C. Bovine Tuberculosis in Cattle and Badgers, <http://www.bovinetb.info/docs/krebs.pdf> (1997).
Abernethy, D. A., Walton, E., Menzies, F., Courcier, E. & Robinson, P. Mycobacterium bovis surveillance in European badgers (Meles meles) killed by vehicles in Northern Ireland: An epidemiological evaluation. Epidemiol. Sante Anim. No 59–60(59–60), 216–218 (2011).
Goodchild, A. V., Watkins, G. H., Sayers, A. R., Jones, J. R. & Clifton-Hadley, R. S. Geographical association between the genotype of bovine tuberculosis in found dead badgers and in cattle herds. Vet. Rec. 170, 259. https://doi.org/10.1136/vr.100193 (2012).
Schroeder, P. et al. Temporal and spatial Mycobacterium bovis prevalence patterns as evidenced in the all wales badgers found dead (AWBFD) survey of infection 2014–2016. Sci. Rep. 10, 2 (2020).
Barron, E. S. et al. A study of tuberculosis in road traffic-killed badgers on the edge of the British bovine TB epidemic area. Sci. Rep. Uk 8, 17206. https://doi.org/10.1038/s41598-018-35652-5 (2018).
McDonald, J. L., Robertson, A. & Silk, M. J. Wildlife disease ecology from the individual to the population: Insights from a long-term study of a naturally infected European badger population. J. Anim. Ecol. 87, 101–112. https://doi.org/10.1111/1365-2656.12743 (2018).
DEFRA. TB surveillance in wildlife – Low Risk Area hotspots, <https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/914655/tb-surveillance-in-wildlife-sept2020.pdf> (2020).
Orton, R. J. et al. Identifying genotype specific elevated-risk areas and associated herd risk factors for bovine tuberculosis spread in British cattle. Epidemics-Neth 24, 34–42 (2018).
Fanshawe, T. R. & Diggle, P. J. Bivariate geostatistical modelling: A review and an application to spatial variation in radon concentrations. Environ. Ecol. Stat. 19, 139–160 (2012).
Dugdale, H. L., Macdonald, D. W. & Newman, C. Offspring sex ratio variation in the European badger Meles meles. Ecology 84, 40–45 (2003).
Rogers, L. M., Cheeseman, C. L., Mallinson, P. J. & CliftonHadley, R. The demography of a high-density badger (Meles meles) population in the west of England. J. Zool. 242, 705–728 (1997).
Woodroffe, R. et al. Spatial association of Mycobacterium bovis infection in cattle and badgers Meles meles. J. Appl. Ecol. 42, 852–862. https://doi.org/10.1111/j.1365-2664.2005.01081.x (2005).
Murphy, D., Gormley, E., Costello, E., O’Meara, D. & Corner, L. A. L. The prevalence and distribution of Mycobacterium bovis infection in European badgers (Meles meles) as determined by enhanced post mortem examination and bacteriological culture. Res. Vet. Sci. 88, 1–5. https://doi.org/10.1016/j.rvsc.2009.05.020 (2010).
Corner, L. A. L., O’Meara, D., Costello, E., Lesellier, S. & Gormley, E. The distribution of Mycobacterium bovis infection in naturally infected badgers. Vet. J. 194, 166–172. https://doi.org/10.1016/j.tvjl.2012.03.013 (2012).
APHA. Bovine tuberculosis in England in 2019 Epidemiological analysis of the 2019 data and historical trends <https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/923195/tb-epidemiology-england-2019.pdf> (2020).
Atkins, P. J. & Robinson, P. A. Bovine tuberculosis and badgers in Britain: Relevance of the past. Epidemiol. Infect. 141, 1437–1444 (2013).
Eisenach, K. D., Cave, M. D., Bates, J. H. & Crawford, J. T. Polymerase chain-reaction amplification of a repetitive DNA-sequence specific for mycobacterium-tuberculosis. J. Infect. Dis. 161, 977–981 (1990).
Woo, P. C. Y., Leung, P. K. L., Leung, K. W. & Yuen, K. Y. Identification by 16S ribosomal RNA gene sequencing of an Enterobacteriaceae species from a bone marrow transplant recipient. J. Clin. Pathol. Mol. Pathol. 53, 211–215. https://doi.org/10.1136/mp.53.4.211 (2000).
Ruettger, A. et al. Rapid spoligotyping of mycobacterium tuberculosis complex bacteria by use of a microarray system with automatic data processing and assignment. J. Clin. Microbiol. 50, 2492–2495. https://doi.org/10.1128/Jcm.00442-12 (2012).
Fanshawe, T. R. & Diggle, P. J. Spatial prediction in the presence of positional error Rejoinder. Environmetrics 22, 129–131 (2011).
Banerjee, S. High-dimensional Bayesian geostatistics. Bayesian Anal 12, 583–614 (2017).
Salvatier, J. & W. T., Fonnesbeck C.,. Probabilistic programming in Python using PyMC3. PeerJ Comput. Sci. 2, e55. https://doi.org/10.7717/peerj-cs.55 (2016).
Loecher, M. & Ropkins, K. RgoogleMaps and loa: Unleashing R graphics power on map tiles. J. Stat. Softw. 63, 1–18 (2014).
Acknowledgements
This project was largely funded by DEFRA (SE3054 / OJEU 28406). The authors are grateful to the many individuals and organisations who collected carcasses and information for the study, and to the pathology and microbiology teams who supported the laboratory work. BMCS is a BloomsburySET Research Fellow, as part of the Connecting Capabilities Fund, Research England (CCF-17-7779).
Author information
Authors and Affiliations
Contributions
B.M.C.S., E.S.B., R.C., D.C., L.G., C.O., C.R., M.S., R.V., M.B. carried out sample processing, badger collection and took part in data collating, analysis, and wrote the main manuscript. C.J. carried out spatial analysis and mansucrip preparation. A.M. and A.P. identified stakeholders and took part in logistics for collection and project management. J.P. took part in stakeholder engagement and badger collection and carried out qualitative analysis of our sampling plan with stakeholders. All authors reveiwed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Swift, B.M.C., Barron, E.S., Christley, R. et al. Tuberculosis in badgers where the bovine tuberculosis epidemic is expanding in cattle in England. Sci Rep 11, 20995 (2021). https://doi.org/10.1038/s41598-021-00473-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-00473-6
This article is cited by
-
Performance of fur clips and livestock markers for identifying vaccinated badgers
European Journal of Wildlife Research (2024)
-
Tuberculosis in wild animals in India
Veterinary Research Communications (2024)
-
Farm management practices and host species richness associated with higher likelihood of tuberculosis positive farms in Western Spain
European Journal of Wildlife Research (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
