
Abstract
In 2012, after the accident at the Fukushima Daiichi Nuclear Power Plant (FDNPP) that followed the Tohoku earthquake and tsunami in March 2011, no rock shell (Thais clavigera; currently recognized as Reishia clavigera; Gastropoda, Neogastropoda, Muricidae) specimens were found near the plant from Hirono to Futaba Beach (a distance of approximately 30 km). In July 2016, however, rock shells were again found to inhabit the area. From April 2017 to May 2019, we collected rock shell specimens monthly at two sites near the FDNPP (Okuma and Tomioka) and at a reference site ~ 120 km south of the FDNPP (Hiraiso). We examined the gonads of the specimens histologically to evaluate their reproductive cycle and sexual maturation. The gonads of the rock shells collected at Okuma, ~ 1 km south of the FDNPP, exhibited consecutive sexual maturation during the 2 years from April 2017 to May 2019, whereas sexual maturation of the gonads of specimens collected at Hiraiso was observed only in summer. The consecutive sexual maturation of the gonads of the specimens collected at Okuma might not represent a temporary phenomenon but rather a site-specific phenotype, possibly caused by specific environmental factors near the FDNPP.
Similar content being viewed by others


Introduction
The 2011 Tohoku earthquake (Mw 9.0) on 11 March 2011 generated a tsunami that caused meltdown of three nuclear reactors at the Fukushima Daiichi Nuclear Power Plant (FDNPP), owned by the Tokyo Electric Power Company. Hydrogen explosions in the reactor buildings caused the emission of hundreds of petabecquerels (PBq) of radionuclides to the environment1. Nevertheless, the estimated amount of radionuclide leakage from the FDNPP is only about one-tenth the total amount released by the 1986 Chernobyl Nuclear Power Plant disaster in Ukraine, which has been estimated as 5300 PBq, excluding noble gases1. Releases of FDNPP-derived radionuclides to the environment have occurred via four major pathways2: 1) Earliest and largest was the initial venting and explosive release of gases and volatile radionuclides to the atmosphere. 2) Subsequently, contaminated material, including radionuclides, was leaked directly from the reactors to the sea during emergency cooling efforts. Finally, releases of radionuclides to the sea via 3) groundwater discharge and 4) river runoff are ongoing. The amounts released via groundwater and river discharge are much smaller than the initial atmospheric fallout and the direct leakage to the sea. Although transport model simulations have indicated that more than 80% of the atmospheric fallout during the 2011 disaster was onto the ocean surface, and that deposition was highest onto coastal waters near the FDNPP, this evaluation was not confirmed by direct observations of atmospheric fallout over the ocean2. Several studies have estimated that total deposition from the atmosphere onto the ocean surface was 5–15 PBq, and direct leakage of 137Cs from the FDNPP into the sea was 3–6 PBq3,4,5,6,7,8,9,10,11,12. Less is known about the amounts of other chemical substances leaked from the FDNPP to the environment (especially the marine environment) following the nuclear disaster, although there is limited information about boric acid and hydrazine13.
Despite the large number of studies that have estimated the amount of radionuclide leakage from the FDNPP to the environment and that have examined the behavior and fate of leaked radionuclides in the environment, little is known about the ecological effects of radionuclides generally, and those derived from the nuclear disaster in particular, on wildlife13,14,15,16. However, some data on the activity concentrations of radionuclides (i.e., gamma emitters) in aquatic organisms (fishes and shellfishes) have been collected, mainly for regulatory purposes17,18,19,20.
Starting in December 2011, following the FDNPP accident in March 2011, we investigated the intertidal zones along the coastline of eastern Japan to evaluate possible ecological effects of harmful substances leaked from the FDNPP into the sea on marine organisms inhabiting areas close to the FDNPP. We found that in 2012 the number of intertidal species decreased significantly with decreasing distance from the power plant. In particular, we were able to collect no rock shell (Thais clavigera) specimens along the coast near the plant from Hirono to Futaba Beach, a distance of approximately 30 km, even though we were able to collect rock shell specimens from many other sites struck by the tsunami13. Furthermore, quantitative surveys in 2013 showed that species richness and population densities of sessile invertebrates, especially Arthropoda, in intertidal zones were much lower at sites near the FDNPP or within several kilometers southward than at other sites, and they were also lower in 2013 than in 1995. These findings strongly suggest that the intertidal biota around the power plant was affected by the nuclear accident13.
According to the results of quantitative quadrat surveys of sessile invertebrates conducted from 2014 to 2019 at seven intertidal sites in Ibaraki, Fukushima, and Miyagi prefectures, including sites near the FDNPP, species richness and population densities in intertidal zones near the FDNPP did not start to increase again until at least 4–5 years after the 2011 disaster14. In April, July, August, and September from 2012 to 2017, we also monitored the population density and spawning behavior of rock shells at sites near the FDNPP. Although in July 2016, rock shells were again found to be inhabiting the area where no rock shells were found in 2012, rock shell population densities and reproductive performance near the FDNPP remained lower in 2017 than before the accident. Thus, sessile invertebrate larval recruitment from remote areas to the intertidal zones near the FDNPP was not clearly observed until 2016 at the earliest. These findings suggest that environmental factors inhibited invertebrate reproduction, recruitment, or both in the intertidal zones near the FDNPP14.
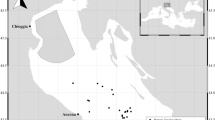
Besides the recovery or increase in rock shell populations at sites near the FDNPP, normal reproduction, in terms of sexual maturation, copulation, egg-laying, hatching, survival, and growth (including metamorphosis and settlement) in the early life history stages and recruitment to populations, is also important to maintain rock shell populations, including at sites near the FDNPP. Therefore, to investigate whether normal reproduction was occurring, we collected rock shell (T. clavigera and Thais bronni) specimens monthly from April 2017 to May 2019 at two sites in Fukushima prefecture near the FDNPP (Ottozawa, Okuma town, ca. 1 km south of the FDNPP, and Tomioka fishing port, Tomioka town, ca. 10 km south of the FDNPP) and at a reference site in Ibaraki prefecture (Hiraiso, Hitachi-naka city, ca. 120 km south of the FDNPP) (Fig. 1 and Supplementary Table S1). We then examined their gonads histologically to evaluate reproductive cycles and sexual maturation in these specimens. Our results revealed a strange phenomenon—consecutive sexual maturation—in a rock shell population in the vicinity of the FDNPP.

available at http://www5b.biglobe.ne.jp/~t-kamada/CBuilder/mapmap.htm and have been modified by the authors.
Sampling sites of the rock shell (Thais clavigera and Thais bronni) between April 2017 and May 2019. Two sites were near the Fukushima Daiichi Nuclear Power Plant (FDNPP) in Fukushima prefecture: Ottozawa, Okuma town, and Tomioka fishing port, Tomioka town are approximately 1 km and 10 km south of the FDNPP, respectively. The reference site, Hiraiso, Hitachi-naka city, Ibaraki prefecture, is approximately 120 km south of the FDNPP. Water temperature was observed at Onahama, Iwaki city, and at Matsukawa-ura, Soma city, by the Fukushima Prefectural Research Institute of Fisheries Resources. *The maps used in this figure are based on a map of Japan
Results
Reproductive cycles and sexual maturation in female T. clavigera and T. bronni
Female reproductive cycles were recognized in temporal changes in the monthly average sexual maturation scores of ovaries from T. clavigera and T. bronni specimens collected at each site (Fig. 2). Ovaries of developmental grades III and IV (corresponding to maturation scores of 3 and 2.5, respectively) in T. clavigera and T. bronni were recognized as sexually mature (Fig. 2 and Table 1).

Time series of maturation scores (purple line) and developmental grades (colored vertical bars) of female Thais clavigera and Thais bronni specimens collected at (a) Hiraiso (T. clavigera), (b) Tomioka (T. clavigera), (c) Okuma (T. clavigera), and (d) Okuma (T. bronni).
At the reference site (Hiraiso), the female maturation score of T. clavigera specimens decreased sharply during August and September 2017 from a peak in June 2017 (Fig. 2a), and a gradual increase was observed from February to May 2018. The peak female maturation score in 2018 was observed in July, and then it sharply decreased from August to September 2018. A gradual recovery of the female maturation score was observed from February to May 2019. On the basis of the ovarian developmental grade composition and the female maturation scores of the T. clavigera specimens collected at Hiraiso, we judged the mature period of T. clavigera at Hiraiso to be June and July of each year. We judged October to February to be the non-reproductive season of T. clavigera at Hiraiso, because immature ovaries (grade I/recovery stage) were predominantly observed in the T. clavigera specimens collected in those months.
At Tomioka, the female maturation score of T. clavigera specimens peaked from May to July 2017 and then decreased sharply in August and September 2017 (Fig. 2b). Similar to the trend at Hiraiso, the score at Tomioka increased again from February to May 2018. The peak female maturation score in 2018 was observed in May and June, and a sharp decrease was observed in July. Although a brief increase was observed in August 2018, the score then decreased again until October. The female maturation score was zero from October 2018 to January 2019, but it rapidly recovered from February to March 2019. On the basis of the ovarian developmental grade composition and female maturation scores of the T. clavigera specimens collected at Tomioka, we judged their mature period to be May to June or July and their non-reproductive season to be from September to February, when grade V (spent stage) ovaries were predominantly observed.
At Okuma, the female maturation scores of T. clavigera specimens were at peak levels during April–September 2017. Thereafter, female maturation scores remained high (> 2.5) throughout most of the survey period. Moreover, the percentage occurrence of mature ovaries (i.e., grades III and IV/mature stage) was high at Okuma during most of the survey period. Thus, the reproductive cycles of female T. clavigera specimens differed markedly at Okuma compared to those at Hiraiso and Tomioka (Fig. 2c). In addition, in T. clavigera specimens collected at Okuma and Hiraiso in the reproductive and non-reproductive seasons (June 2018 and January 2019, respectively; Fig. 3), maturation was clearly observed in ovarian tissues of the specimens collected at Okuma in both June 2018 and January 2019, whereas it was observed in only one female specimen collected at Hiraiso in June 2018. Therefore, consecutive sexual maturation apparently occurred in female T. clavigera at Okuma, the site closest to the FDNPP.

Examples of ovarian tissues of Thais clavigera collected at Okuma and Hiraiso. (a) Female T. clavigera collected at Okuma on 16 June 2018 in the reproductive season, (b) female T. clavigera collected at Okuma on 24 January 2019 in the non-reproductive season, (c) female T. clavigera collected at Hiraiso on 18 June 2018 in the reproductive season, and (d) female T. clavigera collected at Hiraiso on 25 January 2019 in the non-reproductive season. ov, ovary; dg, digestive gland. Bars = 200 µm. (a)–(c) Ovarian tissues are thick, and many mature oocytes can be seen. Therefore, we judged these ovaries to be mature (developmental grade III; see Table 1). (d) The ovarian tissue is thin, and oogonia and early stage oocytes are present, together with connective tissues and hemocytes. Therefore, we judged this ovary to be in the recovery stage (developmental grade I; see Table 1).
At Okuma, we also collected specimens of T. bronni, a T. clavigera relative, and examined the female reproductive cycle, mature period, and other reproductive characteristics for comparison with the female T. clavigera specimens collected at the same site (Fig. 2d). Although the female maturation scores of the T. bronni specimens showed a reproductive cycle, high (> 2.5) female maturation scores and a high percentage occurrence of mature ovaries (i.e., grades III and IV/mature stage) were observed for much longer periods compared with female T. clavigera specimens at Hiraiso and Tomioka. Thus, though the tendency was not as marked as in female T. clavigera, consecutive sexual maturation was also observed in female T. bronni specimens collected at Okuma.
Reproductive cycles and sexual maturation in male T. clavigera and T. bronni
The temporal changes in the average monthly maturation scores of testes of T. clavigera and T. bronni specimens represent the male reproductive cycle (Fig. 4). Developmental grades III and IV (corresponding to maturation scores 2.5 and 3, respectively) were recognized as sexually maturity in testes of T. clavigera and T. bronni (Fig. 4 and Table 2).

Time series of maturation scores and development grades of male Thais clavigera and Thais bronni collected at (a) Hiraiso (T. clavigera), (b) Tomioka (T. clavigera), (c) Okuma (T. clavigera), and (d) Okuma (T. bronni).
At Hiraiso, the male maturation score of T. clavigera specimens decreased sharply in July 2017 from a peak in June (Fig. 4a). Subsequently, it gradually increased to another peak in July 2018, which was followed by a sharp decrease in August. Thus, the trend in 2018 was similar to that in 2017. The male maturation score gradually recovered from January to May 2019. On the basis of the testicular developmental grades of the testicular tissue and the male maturation scores, we judged the mature period of male T. clavigera at Hiraiso to be June and July. In the non-reproductive season, from August to November or December, the testes of most specimens were immature (grade I/recovery stage).
At Tomioka, the male maturation score of T. clavigera specimens peaked in May–June 2017 and then decreased sharply in July and August (Fig. 4b). It increased thereafter from November 2017 to April 2018, in a similar pattern to that at Hiraiso. In 2018, the peak male maturation score was observed in May and June, and a sharp decrease was observed in July. A gradual recovery was observed from November 2018 to May 2019. On the basis of the testicular developmental grades and the male maturation scores, we judged their mature period to be April–June (but not, perhaps, in 2019). In the non-reproductive season, from August to October, immature testes (grade I/recovery stage) were predominantly observed.
At Okuma, the male maturation score of T. clavigera specimens was at peak levels during April–July 2017, and it decreased sharply in September 2017 (Fig. 4c). Thereafter, male maturation scores remained high (≥ 2.5), and a high percentage occurrence of mature testes (i.e., mature stage/grades III and IV), were observed throughout most of the survey period (except in August and September 2018). Thus, the male reproductive cycle differed markedly between specimens collected at Okuma and those collected at Hiraiso and Tomioka. Therefore, we also observed apparently consecutive sexual maturation in male T. clavigera specimens collected at Okuma, although the pattern appeared somewhat weaker than that in female T. clavigera collected there.
We also compared the reproductive cycle, mature period, and other male reproductive characteristics in male T. bronni specimens collected at Okuma with those of the male T. clavigera specimens collected at the same site (Fig. 4d). Although the male maturation score of T. bronni specimens collected at Okuma showed a reproductive cycle (characterized by an abrupt decline of the male maturation score in September 2017 and August 2018, similar to the declines observed in male T. clavigera specimens collected at Okuma), high (≥ 2.5) male maturation scores and a high percentage occurrence of mature testes (i.e., grades III and IV/mature stage) were observed over a much longer period at Okuma than at Hiraiso and Tomioka, Therefore, we also observed apparently consecutive sexual maturation in male T. bronni at Okuma, although, similar to female T. bronni compared with female T. clavigera, the tendency was less marked in male T. bronni than in male T. clavigera.
Discussion
We observed differences in shell size (i.e., shell height and weight) of T. clavigera specimens among the three sites; in general, specimens collected at Okuma were bigger than those collected at Hiraiso and Tomioka. Because population densities (i.e., abundances) of mussels (Mytilus galloprovincialis and Septifer virgatus) and barnacles (Chthamalus challengeri), which are prey for T. clavigera, were higher at Okuma than at Tomioka and Hiraiso14, the growth rate of the T. clavigera population might have been faster at Okuma than at Tomioka and Hiraiso. However, because the shell height of all of the studied T. clavigera specimens collected at Okuma, Tomioka, and Hiraiso exceeded 12 mm, the minimum shell height of a mature specimen21 (Supplementary Table S1), the reproductive cycle and sexual maturation in the female and male T. clavigera specimens could be compared among the sites.
We inferred that the breeding season of the T. clavigera population at Hiraiso during the summer, on the basis of the observed female and male reproductive cycles (Figs. 2a and 4a). Kon et al.22 reported that the breeding season of the T. clavigera population at Yoriihama, Niigata, Japan, extended from late spring to summer. In general, T. clavigera populations in Japan mature from late spring to summer, and the females deposit their egg capsules beneath ledges or on other substrates (e.g., tetrapods and similar concrete structures set along the coast for wave protection)23. At Okuma, however, we observed sexual maturation in both female and male T. clavigera specimens throughout most of each year of the survey period (April 2017 to May 2019) (Figs. 2c and 4c). Moreover, we observed consecutive sexual maturation in both female and male T. bronni specimens collected at Okuma, though the tendency was less marked than it was in T. clavigera (Figs. 2d and 4d). Although we observe a rapid regression of the testis in male T. clavigera specimens collected at Okuma in September of both 2017 and 2018, they recovered rapidly to maturation in October of both years (Fig. 4c). Similar to T. clavigera, we also observed a rapid regression of the testis in male T. bronni specimens collected at Okuma in September 2017 and August 2018, followed by recovery to maturation 2 months later, in November 2017 and October 2018, respectively (Fig. 4d). Therefore, both T. bronni and T. clavigera populations apparently breed throughout most of the year at Okuma. In fact, we observed egg laying (i.e., egg capsules deposited on substrates) by T. clavigera and T. bronni at Okuma as late as 21 September in 2017 and 10 September in 2018. Unfortunately, it is impossible to directly observe egg laying by T. clavigera and T. bronni on substrates at Okuma and other sites from October to next March because the tide level is high, even at low tide, and wave action is strong. To the best of our knowledge, consecutive sexual maturation and egg laying in mid-September have not been reported previously in T. clavigera and T. bronni.
It is possible that, with respect to their reproductive cycle, the T. clavigera population at Tomioka might be intermediate between Okuma and Hiraiso, because we observed spent ovaries (grade V/spent stage) in female T. clavigera specimens predominantly from September to February at Tomioka, whereas at Hiraiso, we observed immature ovaries (grade I/recovery stage) predominantly from October to February (Fig. 2b). Thus, the non-reproductive period differed slightly between Tomioka and Hiraiso.
Was the consecutive sexual maturation observed in the T. clavigera and T. bronni populations at Okuma related to radionuclides leaked from the FDNPP? First, the T. clavigera and T. bronni specimens collected at Okuma in the present study did not seem to be the direct offspring of parent shells that had probably been exposed to radionuclides leaked during the March 2011 nuclear disaster because no rock shell (T. clavigera) specimens were collected near the plant, from Hirono to Futaba Beach (a distance of approximately 30 km) in April 201213. The absence of rock shells at sites close to the FDNPP in April 2012 suggests the likely mortality of almost all individuals living there after March 2011 and, therefore, that reproduction and recruitment either did not occur there or were less successful in summer and autumn (the reproductive season and thereafter) of 2011. However, why adult rock shells living near the FDNPP disappeared or had little or no reproductive success in 2011 is still unknown13. Second, the T. clavigera and T. bronni specimens collected at Okuma in the present study seemed to be offspring of parent shells that had inhabited various sites to the north and south of the FDNPP14. The early life stage of T. clavigera and T. bronni, which lasts more than 2 months21, is a free-swimming planktonic veliger. Therefore, the T. clavigera and T. bronni specimens collected at Okuma might have been spawned in remote areas and have subsequently traveled as veligers to the intertidal zone at Okuma through dispersion and migration processes. In addition, the activity concentrations of radionuclides, including radiocesium (134Cs and 137Cs), in seawater have been mostly less than 0.1 Bq/L, even at sites close to the FDNPP, since April 2017 (https://radioactivity.nsr.go.jp/ja/contents/9000/8141/24/engan.pdf; in Japanese). Therefore, the dose rates of the T. clavigera and T. bronni specimens collected at Okuma in the present study could be much lower than 400 μGy/h (= 9.6 mGy/d), which is the threshold value estimated by the United Nations Scientific Committee on the Effects of Atomic Radiation for a detrimental effect on populations of aquatic organisms24. Thus, it appears to be unlikely that the consecutive sexual maturation observed in both T. clavigera and T. bronni populations at Okuma in the vicinity of the FDNPP is associated with radionuclides such as 137Cs leaked from the power plant.
In general, the reproductive cycle (including spawning behavior) of gastropods, including Thais spp., is controlled by both external (i.e., water temperature, illumination/photoperiod, daily and monthly tidal cycles, and physical factors such as rough seas and onshore winds) and internal factors (i.e., genetics, nutrition, and neurohormones/neuropeptides)25,26,27. In addition, at low latitudes, an extended reproductive season is common, whereas at high latitudes the season tends to be restricted by the need for the phytotrophic veliger stage to coincide with phytoplankton blooms. Variation in the length of the reproductive season within their geographic range is evident in two species, Hinia incrassata and Lacuna vincta, both with planktonic veligers, that occur at Plymouth, U.K. The former, whose range extends into warmer waters, breeds during spring and summer at Plymouth but throughout the year in Bermuda28, whereas the latter, whose range extends into colder waters, breeds during January–May at Plymouth and in June–July or August in the White Sea, an inlet of the Barents Sea29. Yamaguchi30 reported that the green snail (Turbo marmoratus), which has separate sexes, is able to reproduce at the age of about 4 years when its shell width reaches about 130 to 150 mm. At higher latitudes, however, this snail appears to breed only in the summer months, when the water temperature is higher, whereas at lower latitudes, mature individuals breed repeatedly throughout the year30. In addition, giant clam (Tridacna gigas) populations reproduce throughout the year at the low-latitude site of Palau31.
Different from the high water temperatures observed year-round in tropical and subtropical areas, water temperatures at Onahama, Iwaki city (on the southern Fukushima coast) fluctuated from 8.4 to 25.2 °C in 2017, from 7.0 to 24.2 °C in 2018, and 9.7 to 24.9 °C in 2019 (https://www.pref.fukushima.lg.jp/uploaded/attachment/363615.pdf; in Japanese), whereas those at Matsukawa-ura, Soma city (on the northern Fukushima coast), fluctuated from 6.1 to 26.1 °C in 2017, from 5.0 to 28.5 °C in 2018, and from 7.0 to 27.2 °C in 2019 (https://www.pref.fukushima.lg.jp/uploaded/attachment/363616.pdf; in Japanese) (Fig. 1). Thus, it is unlikely that the consecutive sexual maturation observed in both female and male T. bronni and T. clavigera specimens collected at Okuma in the present study was dependent on high water temperatures. Although we observed egg laying (i.e., egg capsules laid on substrates) by both T. clavigera and T. bronni at Okuma in September of both 2017 and 2018, to the best of our knowledge, consecutive sexual maturation and egg laying in mid-September by T. clavigera and T. bronni has not been reported previously. Thus, both these phenomena are unusual and differ markedly from our observations at Hiraiso and the findings reported by Kon et al.22 for the T. clavigera population in Niigata, Japan.
We observed consecutive sexual maturation in both sexes of T. clavigera and T. bronni specimens collected at Okuma over about 2 years (April 2017 to May 2019; Figs. 2c, 2d, 4c, and 4d). Thus, rather than a temporary phenomenon, it is probably a site-specific phenotypic response, possibly caused by specific environmental factors near the FDNPP.
We speculate that consecutive sexual maturation may be caused by either a maturation switch being turned on all the time, or that it is an irreversible response such that once maturation starts, a return to the original physiological phase is not possible. The endocrinology and reproductive physiology, such as controls for sexual maturation, of gastropods, especially prosobranchs, are not yet fully understood. It has been suggested that gastropods might have two co-existing regulatory systems for reproduction: neurohormones/neuropeptides and vertebrate-type steroid hormones32. Recently, however, doubts have been raised as to whether gastropods inherently have vertebrate-type steroids that function as sex hormones33,34,35,36,37. Thus, in the future, a comparative endocrinology study should compare expression levels of neuropeptides and their seasonal/temporal variations between the T. clavigera population at Okuma and that at Hiraiso to elucidate the mechanism of consecutive sexual maturation in the Okuma population.
The possibility that nuclear receptors, such as the retinoid X receptor (RXR), are involved in the development of consecutive sexual maturation in T. clavigera and T. bronni at Okuma should also be investigated. RXR is known to be involved in the induction of imposex, a type of endocrine disruption, in gastropods38,39,40,41,42,43,44,45. Imposex is an irreversible syndrome in which male sexual characteristics and organs such as the penis and vas deferens are superimposed in female prosobranch gastropods. Very low concentrations (ppt or ng/L) of organotin compounds such as tributyltin and triphenyltin, which are used in antifouling paints for ships and fishing nets, are known to induce imposex in prosobranch gastropods46,47,48,49,50,51,52,53. In severe cases, imposex may result in a sex change (i.e., spermatogenesis in the ovary, or a change of ovarian tissue to testicular tissue)54,55,56,57. Thus, RXR may also have a physiological effect of inducing gonad abnormalities, including the consecutive sexual maturation observed in the T. clavigera and T. bronni specimens collected at Okuma.
Rock shells (T. clavigera, and T. bronni) have a free-swimming planktonic veliger early life stage that lasts more than 2 months21. Therefore, the T. clavigera and T. bronni specimens collected at Okuma could have originated in remote areas and have traveled to the intertidal zone at Okuma as veligers through dispersion and migration processes. Therefore, it is unlikely that consecutive sexual maturation is congenital in the T. clavigera and T. bronni specimens collected at Okuma. Instead, exposure of T. clavigera and T. bronni larvae in the (late) planktonic stage or in the juvenile stage after settlement in the intertidal zone at Okuma to certain environmental factors around the FDNPP may have triggered the induction of consecutive sexual maturation in the Okuma populations.
Changes in life history traits and population dynamics of marine organisms in response to environmental change are known to occur in nature, although the causes underlying the responses remain unidentified58. Kodama et al.59 reported changes in the growth and reproductive traits of the dragonet, Callionymus valenciennei, in Tokyo Bay, Japan, that coincided in a decrease in stock size. The minimum standard length at which the dragonet attains gonadal maturation was smaller in the 2000s (4.8 cm) than in the 1990s (6.0 cm). In addition, the timing of the onset of spawning was earlier in the 2000s (spring) than in the 1990s (summer). Kodama et al.59 also found significant changes in the growth in both sexes from the 1990s to the 2000s; growth after sexual maturity was attained was significantly decreased in the 2000s compared with the 1990s. Changes in life history traits may reflect a trade-off in the allocation of available energy resources between reproduction and somatic growth; under limited prey abundance, more energy resources may be allocated to reproduction to enhance stock recovery. These findings suggest that consecutive sexual maturation in T. clavigera and T. bronni populations at Okuma, where no individuals of these species were observed from December 2011 to July 2016, following the FDNPP accident13,14, may reflect such a trade-off to enhance the recovery of the populations, although we have no data indicating that such an energy resource allocation trade-off has occurred. It is also unclear whether this life history change (i.e., consecutive sexual maturation) might, for example, cause the rock shell populations to be more resilient to the negative effects of low water temperature on larval and juvenile survival in winter. Thus, the impacts of consecutive sexual maturation on T. clavigera and T. bronni populations are unknown. Future studies in ecotoxicology as well as in the developmental biology and ecology of the early life stages of these species are needed to identify the causal factors of consecutive sexual maturation and to evaluate its effects on the population dynamics of T. clavigera and T. bronni at Okuma.
Further field studies are also necessary to constrain the geographic area in which consecutive sexual maturation in T. clavigera specimens occurs, and whether other species in the intertidal zone at Okuma also exhibit consecutive sexual maturation.
Methods
Collection of rock shell specimens
From April 2017 to May 2019, we collected specimens of rock shells monthly at two sites in Fukushima prefecture near the FDNPP. At Ottozawa (Okuma town, ca. 1 km south of the FDNPP), we collected both T. clavigera and T. bronni (currently, recognized as Reishia clavigera (Küster, 1860) and Reishia bronni (Dunker, 1860), respectively; Gastropoda, Neogastropoda, Muricidae) specimens, and at Tomioka fishing port (Tomioka town, ca. 10 km south of the FDNPP), we collected T. clavigera specimens. We also collected T. clavigera specimens at a reference site in Ibaraki prefecture (Hiraiso, Hitachi-naka city, ca. 120 km south of the FDNPP). On each collection date, we collected 7–14 individuals, on average 10.3 T. clavigera and 10.0 T. bronni individuals (Fig. 1, Supplementary Table S1). After collection, the specimens were dissected and their sex was determined by examining their accessory sex organs60. Then, the specimens were fixed in Bouin’s fluid (see below) for 48 h for later histological examination of the gonads.
Histopathological examination of T. clavigera and T. bronni gonads
The gonads of the rock shell specimens collected monthly at each site were fixed in Bouin’s fluid (saturated picric acid : formaldehyde solution : acetic acid = 15 : 5 : 1, all reagents were guaranteed to be extra pure by Nacalai Tesque, Inc.) for 48 h, embedded in paraffin (Sigma-Aldrich, Paraplast Plus), cut into 5–6 μm sections with a rotary microtome (Leica, RM2125RT), and stained with hematoxylin (Tissue-Tek Hematoxylin 3G) and eosin (Tissue-Tek Eosin) by an autostaining machine (Leica, Autostainer XL). Histopathological examination of the gonads was conducted under a light microscope (Olympus, BX40) equipped with a digital camera (Olympus, DP22), and the images were analyzed by using image analysis software (Olympus, cellSens Standard V2.2 (Build 17,989) (https://www.olympus-lifescience.com/en/software/cellsens/#!cms[focus]=cmsContent6017)) installed on a personal computer (Hewlett-Packard, DP2-PC-S-1–21).
To quantitatively evaluate gonadal maturation in the rock shell specimens, female and male reproductive cells were scored according to a developmental grading system similar to that described by Takamaru and Fuji61. Female reproductive cells were categorized according to their degree of maturity as an (1) oogonium or an (2) early (eoc), (3) middle (moc), (4) late (loc), or (5) mature (mo) oocyte (Fig. 5). An oogonium has a nucleus containing chromatin chains and a small nucleolus, as well as small, hematoxylin-stained granules. The cytoplasm is thin and its stainability is weak (Fig. 5a). An early stage oocyte is surrounded by a single layer of follicular cells. The large nucleus contains dispersed or scattered chromatin and a large basophilic nucleolus. The early stage oocyte is larger than an oogonium, and it has an irregular shape (Fig. 5b). No yolk globule accumulation is observed in either the oogonium or the early stage oocyte. In the middle stage, the oocyte starts to accumulate eosinophilic yolk granules in the cytoplasm. Its nucleolus is large and basophilic, and chromatin is dispersed in the nucleus. The middle stage oocyte is larger than an early stage oocyte and contains a few yolk globules stained by eosin (Fig. 5c). The late stage oocyte accumulates more eosinophilic yolk globules in the cytoplasm. Its outline is becoming unclear, and its nucleolus is more weakly stained. The late stage oocyte is larger than the middle stage oocyte and contains several yolk globules stained by eosin (Fig. 5d). A mature oocyte has an unclear outline, and its cytoplasm is filled with yolk globules. Its nucleus has an indeterminate form, and the nuclear membrane has disappeared. Small yolk granules cluster around the nucleus (Fig. 5e).

Reproductive cells in ovaries of Thais clavigera: (a) oogonium (og), (b) early stage oocyte (eoc), (c) middle stage oocyte (moc), (d) late stage oocyte (loc), and (e) mature oocyte (mo). Bars = 200 µm.
The developmental grade of an ovary could range from I to V and was determined on the basis of the characteristics of the ovarian tissue and the primary cells observed in the ovary (Table 1). Then, a maturation score based on the developmental grade of the ovary was assigned to each female T. clavigera and T. bronni specimen collected monthly at each site (Table 1).
Male reproductive cells were also categorized according their degree of maturity as a (1) spermatogonium, (2) spermatocyte, (3) spermatid, or (4) spermatozoon (Fig. 6). The spermatogonium, which is about half the size of a spermatocyte, is an almost round cell having a nucleus that contains small granules stained by hematoxylin. The stainability of the cytoplasm is weak. A spermatocyte is a round cell with a large nucleus and scattered punctiform chromatin. The stainability of a spermatocyte is weak. The spermatid is generally an oval or round cell that is intensively stained by hematoxylin, but some spermatids are crescent- or rod-shaped, with a reduced volume of cytoplasm. The spermatozoon is a thread-shaped cell that is intensively stained by hematoxylin. Some cells almost the same size as spermatocytes but with many granules intensively stained by hematoxylin were also observed in the testes of T. clavigera and T. bronni.

Reproductive cells in a testis of Thais clavigera: spermatogonium (sg), spermatocyte (sc), spermatid (st), and spermatozoon (sz). Unidentified cells (uc) that were almost the same size as spermatocytes and contained many granules intensively stained by hematoxylin were observed in the testes of both Thais bronni and T. clavigera. Bar = 200 µm.
The developmental grade of a testis, ranging from I to V, was determined from the characteristics of the testicular tissue and the primary cells observed in the testis (Table 2). Then, a maturation score based on the developmental grade of the testis was assigned to each male T. clavigera and T. bronni specimen collected monthly at each site (Table 2).
Average maturation scores of ovaries and testes of T. clavigera and T. bronni specimens collected each month at each site were calculated. The temporal changes of these scores were considered to represent the reproductive cycle in each species61.
Data availability
All data generated or analyzed in this study are included in this published article or in the Supplementary Information.
References
Steinhauser, G., Brandl, A. & Johnson, T. E. Comparison of the Chernobyl and Fukushima nuclear accidents: a review of the environmental impacts. Sci. Total Environ. 470–471, 800–817 (2014).
Buesseler, K. et al. Fukushima Daiichi-derived radionuclides in the ocean: transport, fate, and impacts. Annu. Rev. Mar. Sci. 9, 173–203 (2017).
Aoyama, M. et al. 134Cs and 137Cs in the North Pacific Ocean derived from the March 2011 TEPCO Fukushima Dai-ichi Nuclear Power Plant accident, Japan. Part two: estimation of 134Cs and 137Cs inventories in the North Pacific Ocean. J. Oceanography 72, 67–76 (2016).
Charette, M. A. et al. Radium-based estimates of cesium isotope transport and total direct ocean discharges from the Fukushima nuclear power plant accident. Biogeosciences 10, 2159–2167 (2013).
Estournel, C. et al. Assessment of the amount of cesium-137 released into the Pacific Ocean after the Fukushima accident and analysis of its dispersion in Japanese coastal waters. J. Geophys. Res. 117, C11014 (2012).
Kawamura, H. et al. Preliminary numerical experiments on oceanic dispersion of 131I and 137Cs discharged into the ocean because of the Fukushima Daiichi nuclear power plant disaster. J. Nucl. Sci. Technol. 48, 1349–1356 (2011).
Kobayashi, T., Nagai, H., Chino, M. & Kawamura, H. Source term estimation of atmospheric release due to the Fukushima Dai-Ichi nuclear power plant accident by atmospheric and oceanic dispersion simulations. J. Nucl. Sci. Technol. 50, 255–264 (2013).
Miyazawa, Y. et al. Inverse estimation of source parameters of oceanic radioactivity dispersion models associated with the Fukushima accident. Biogeosciences 10, 2349–2363 (2013).
Rypina, I. I. et al. Short-term dispersal of Fukushima-derived radionuclides off Japan: modeling efforts and model-data intercomparison. Biogeosciences 10, 4973–4990 (2013).
Tsumune, D., Tsubono, T., Aoyama, M. & Hirose, K. Distribution of oceanic 137Cs from the Fukushima Dai-ichi nuclear power plant simulated numerically by a regional ocean model. J. Environ. Radioact. 111, 100–108 (2012).
Tsumune, D. et al. One-year, regional-scale simulation of 137Cs radioactivity in the ocean following the Fukushima Daiichi nuclear power plant accident. Biogeosciences 10, 5601–5617 (2013).
Smith, G. Sources, Effects and risks of ionizing radiation UNSCEAR 2013 Report. Volume I: Report to the General Assembly, Annex A: Levels and effects of radiation exposure due to the nuclear accident after the 2011 great east-Japan earthquake and tsunami. Journal of radiological protection: official journal of the Society for Radiological Protection, 34, 725 (2014).
Horiguchi, T. et al. Decline in intertidal biota after the 2011 Great East Japan Earthquake and Tsunami and the Fukushima nuclear disaster: field observations. Sci. Rep. 6, 20416. https://doi.org/10.1038/srep20416 (2016).
Horiguchi, T., Kodama, K., Kume, G. & Kang, I. J. Delayed recovery from declines in the population densities and species richness of intertidal invertebrates near Fukushima Daiichi Nuclear Power Plant. In Low-dose radiation effects on animals and ecosystems Long-Term Study on the Fukushima Nuclear Accident (ed. Fukumoto, M.) 65–88 (Springer, Berlin, 2019).
Kodama, K., Aramaki, T. & Horiguchi, T. Current status of the megabenthic community in coastal Fukushima Prefecture, Japan, in the wake of the Great East Japan Earthquake. Mar. Environ. Res. 140, 358–374 (2018).
Nohara, C. et al. The biological impacts of ingested radioactive materials on the pale grass blue butterfly. Sci. Rep. 4, 4946. https://doi.org/10.1038/srep04946 (2014).
Fisheries Agency of Japan. Results of the monitoring on radioactivity level in fisheries products. https://www.jfa.maff.go.jp/e/inspection/index.html (2020)
Horiguchi, T. et al. Radiocesium in seawater, sediments, and marine megabenthic species in coastal waters off Fukushima in 2012–2016, after the 2011 nuclear disaster. Mar. Environ. Res. 137, 206–224 (2018).
Wada, T. et al. Effects of the nuclear disaster on marine products in Fukushima. J. Environ. Radioact. 124, 246–254 (2013).
Wada, T. et al. Effects of the nuclear disaster on marine products in Fukushima: An update after five years. J. Environ. Radioact. 164, 312–324 (2016).
Nakano, D., Nagoshi, M. Growth and age of Thais clavigera (Küster), Prosobranch, in tidal zone around Shima Peninsula, Japan. In 25th Anniversary Memorial Journal for Toba Aquarium, 87–92 (1980) (in Japanese)
Kon, T., Honma, Y. & Murakawa, S. Studies on the maturity of gonad in some marine invertebrates – I. Seasonal changes in the gonads of prosobranch mollusk, Purpura (Mancinella) clavigera. Bull. Japan. Soc. Sci. Fish. 32, 484–491 (1966) (in Japanese).
Okutani, T. Marine mollusks in Japan, (ed. Okutani, T.) (Tokai University Press, Tokai, 2000) (in Japanese)
United Nations Scientific Committee on the Effects of Atomic Radiation. Sources and effects of ionizing radiation. Volume II: Effects, Scientific Annexes C, D and E. United Nations Scientific Committee on the Effects of Atomic Radiation sales publication E.11.IX.3 (United Nations, 2011)
Fretter, V. Prosobranchs in The mollusca Volume 7 Reproduction (eds. Tompa, A.S., Verdonk, N.H. & van den Biggelaar, J.A.M.) 1–45 (Academic Press, Inc., 1984).
Morishita, F. Neuropeptides and their physiological functions in mollusks in Biological effects by organotins (ed. Horiguchi, T.) 167–197 (Springer, 2017).
Sumikawa, S. Sex and Reproduction in Handbook of malacology Volume 1 (eds. Habe, T., Okutani, T. & Nishiwaki, S.) 159–176 (Scientist, Inc., 1994) (in Japanese)
Lebour, M. V. The eggs and larvae of some prosobranchs from Bermuda. Proc. Zool. Soc. London. 114, 462–489 (1945).
Kuznetsov, V.V. Seasonal and temperature conditions for the breeding of marine invertebrates in Data for a comprehensive study of the White Sea (ed. Palenichko, Z.G.) 32–52 (Akad. Nauk. SSSR, Moscow, 1963) (in Russian)
Yamaguchi, M. Green Snail. FFA Report 92/61 at https://spccfpstore1.blob.core.windows.net/digitallibrary-docs/files/af/af471a899f87b7444000345d543c5256.pdf?sv=2015-12-11&sr=b&sig=RjAGQi0dESjYNRnAXOhcZ3ouFX9SpQGkLr8www4CnG4%3D&se=2021-04-01T06%3A14%3A13Z&sp=r&rscc=public%2C%20max-age%3D864000%2C%20max-stale%3D86400&rsct=application%2Fpdf&rscd=inline%3B%20filename%3D%22FFA_1992_061.pdf%22 (1993).
Heslinga, G. A., Perron, F. E. & Orak, O. Mass culture of giant clams (F Tridacnidae) in Palau. Aquaculture 39, 197–215 (1984).
LeBlanc, G. A. et al. The endocrinology of invertebrates. In Endocrine disruption in invertebrates: endocrinology, testing, and assessment (eds de Fur, P. L. et al.) 23–106 (SETAC Press, Cambridge, 1999).
Horiguchi, T. Mechanism of imposex induced by organotins in gastropods. In Ecotoxicology of antifouling biocides (eds Arai, T. et al.) 111–124 (Springer, Berlin, 2009).
Horiguchi, T. Mode of action of organotins to induce the development of imposex in gastropods, focusing on steroid and the retinoid X receptor activation hypotheses. In Biological effects by organotins (ed. Horiguchi, T.) 199–219 (Springer, Berlin, 2017).
Horiguchi, T. & Ohta, Y. A critical review of sex steroid hormones and the induction mechanisms of imposex in gastropod mollusks. In Advances in invertebrate (neuro)endocrinology: a collection of reviews in the post-genomic era (2-volume set). Volume 1 Phyla other than Arthropoda (eds Saleuddin, S. et al.) 255–283 (Apple Academic Press Inc., London, 2020).
Scott, A. P. Do mollusks use vertebrate sex steroids as reproductive hormones? Part I: Critical appraisal of the evidence for the presence, biosynthesis and uptake of steroids. Steroids 77, 1450–1468 (2012).
Scott, A. P. Do mollusks use vertebrate sex steroids as reproductive hormones? II Critical review of the evidence that steroids have biological effects. Steroids 78, 268–281 (2013).
Castro, L. F. C. et al. Imposex induction is mediated through the Retinoid X Receptor signalling pathway in the neogastropod Nucella lapillus. Aquat. Toxicol. 85, 57–66 (2007).
Horiguchi, T. et al. Retinoid X receptor gene expression and protein content in tissues of the rock shell Thais clavigera. Aquat. Toxicol. 84, 379–388 (2007).
Horiguchi, T. et al. Exposure to 9-cis retinoic acid induces penis and vas deferens development in the female rock shell Thais clavigera. Cell Biol. Toxicol. 24, 553–562 (2008).
Horiguchi, T. et al. Time course of expression of the retinoid X receptor gene and induction of imposex in the rock shell, Thais clavigera, exposed to triphenyltin chloride. Anal. Bioanal. Chem. 396, 597–607 (2010).
Horiguchi, T. et al. Establishment of a polyclonal antibody against the retinoid X receptor of the rock shell Thais clavigera and its application to rock shell tissues for imposex research. Ecotoxicology 19, 571–576 (2010).
Nishikawa, J. et al. Involvement of the retinoid X receptor in the development of imposex caused by organotins in gastropods. Environ. Sci. Technol. 38, 6271–6276 (2004).
Sternberg, R. M., Hotchkiss, A. K. & Leblanc, G. A. Synchronized expression of retinoid X receptor mRNA with reproductive tract recrudescence in an imposex-susceptible mollusc. Environ. Sci. Technol. 42, 1345–1351 (2008).
Urushitani, H. et al. Cloning and characterization of retinoid X receptor (RXR) isoforms in the rock shell Thais clavigera. Aquat. Toxicol. 103, 101–111 (2011).
Bryan, G. W. et al. The decline of the gastropod Nucella lapillus around south-west England: evidence for the effect of tributyltin from antifouling paints. J. Mar. Biol. Assoc. UK 66, 611–640 (1986).
Bryan, G. W. et al. The effects of tributyltin (TBT) accumulation on adult dog-whelks, Nucella lapillus: long-term field and laboratory experiments. J Mar. Biol. Assoc. UK 67, 525–544 (1987).
Bryan, G. W., Gibbs, P. E. & Burt, G. R. A comparison of the effectiveness of tri-n-butyltin chloride and five other organotin compounds in promoting the development of imposex in the dog-whelk, Nucella lapillus. J. Mar. Biol. Assoc. UK 68, 733–744 (1988).
Gibbs, P. E. & Bryan, G. W. Reproductive failure in populations of the dog-whelk, Nucella lapillus, caused by imposex induced by tributyltin from antifouling paints. J. Mar. Biol. Assoc. UK 66, 767–777 (1986).
Gibbs, P. E. et al. The use of the dog-whelk, Nucella lapillus, as an indicator of tributyltin (TBT) contamination. J. Mar. Biol. Assoc. UK 67, 507–523 (1987).
Horiguchi, T. et al. Imposex in Japanese gastropods (Neogastropoda and Mesogastropoda): effects of tributyltin and triphenyltin from antifouling paints. Mar. Pollut. Bull. 31, 402–405 (1995).
Horiguchi, T. et al. Effects of triphenyltin chloride and five other organotin compounds on the development of imposex in the rock shell Thais clavigera. Environ. Pollut. 95, 85–91 (1997).
Smith, B. S. Sexuality in the American mud snail, Nassarius obsoletus Say. Proc. Malacol. Soc. Lond. 39, 377–378 (1971).
Gibbs, P. E., Pascoe, P. L. & Burt, G. R. Sex change in the female dog-whelk, Nucella lapillus, induced by tributyltin from antifouling paints. J. Mar. Biol. Assoc. UK 68, 715–731 (1988).
Horiguchi, T. & Shimizu, M. Effects on aquatic organisms, mainly on molluscs. In Organotin pollution and its effects on aquatic organisms (eds Satomi, Y. & Shimizu, M.) 99–135 (Koseisha-Koseikaku, Tokyo, 1992).
Horiguchi, T. et al. Impact of tributyltin and triphenyltin on ivory shell (Babylonia japonica) populations. Environ. Health Perspect. 114(Supplement 1), 13–19 (2006).
Oehlmann, J. et al. Tributyltin (TBT) effects on Ocinebrina aciculata (Gastropoda: Muricidae): imposex development, sterilization, sex change and population decline. Sci. Total Environ. 188, 205–223 (1996).
Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 37, 637–669 (2006).
Kodama, K., Lee, J. H., Shiraishi, H. & Horiguchi, T. Changes in growth and reproductive traits of dragonet Callionymus valenciennei in Tokyo Bay, concurrent with decrease in stock size. Mar. Environ. Res. 93, 23–30 (2014).
Horiguchi, T. et al. Imposex and organotin compounds in Thais clavigera and T. bonni in Japan. J. Mar. Biol. Assoc. UK 74, 651–669 (1994).
Takamaru, T. & Fuji, A. Reproductive cycles in populations of Neptunea arthritica from the southern coastal waters of Hokkaido Japan. Suisan Zoshoku 29, 78–87 (1981) (in Japanese).
Acknowledgements
We are grateful to the Fukushima Prefectural Government for permission to enter “difficult-to-return zone” in the vicinity of the Fukushima Daiichi Nuclear Power Plant. This work was supported in part a Grant-in-Aid for Scientific Research (B) from the Japan Society for the Promotion of Science (JSPS), Japan, to T.H. (#15H04537) and by a Grant-in-Aid for Scientific Research (A) from JSPS to T.H. (#18H03962).
Author information
Authors and Affiliations
Contributions
T.H. planned and designed the study and collected the specimens. K.K. made histological preparations. T.H., K.K., and Y.O. observed the histological preparations under a light microscope. T.H. analyzed all of the data and wrote the manuscript and prepared the tables, figures, and supplementary information. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Horiguchi, T., Kawamura, K. & Ohta, Y. Consecutive sexual maturation observed in a rock shell population in the vicinity of the Fukushima Daiichi Nuclear Power Plant, Japan. Sci Rep 11, 560 (2021). https://doi.org/10.1038/s41598-020-80686-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-80686-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
