
Abstract
Accurate morphological ant mimicry by Myrmarachne jumping spiders confers strong protective benefits against predators. However, it has been hypothesized that the slender and constricted ant-like appearance imposes costs on the hunting ability because their jumping power to capture prey is obtained from hydraulic pressure in their bodies. This hypothesis remains to be sufficiently investigated. We compared the jumping and prey-capture abilities of seven Myrmarachne species and non-myrmecomorphic salticids collected from tropical forests in Malaysian Borneo and northeastern Thailand. We found that the mimics had significantly reduced abilities compared with the non-mimics. The analysis using geometric morphometric techniques revealed that the reduced abilities were strongly associated with the morphological traits for ant mimicry and relatively lower abilities were found in Myrmarachne species with a more narrowed form. These results support the hypothesis that the jumping ability to capture prey is constrained by the morphological mimicry and provide a new insight into understanding the evolutionary costs of accurate mimicry.
Similar content being viewed by others


Introduction
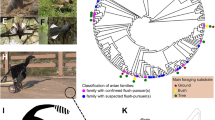
Morphological ant mimicry is widespread in spiders1. Ants possess species-specific combinations of strong defensive traits, including powerful mouthparts and stings, chemical defenses, general aggressiveness, and nest-mate-recruiting ability2. Thus, morphological ant mimicry has been considered to be a good example of the Batesian mimicry1. Some of the most striking examples of morphological ant mimicry are found in the Myrmarachne, the largest genus of jumping spiders (Salticidae) with more than 200 species3,4,5. Myrmarachne has two body regions, a cephalothorax and an abdomen, but a constriction in the cephalothorax makes this portion resemble the separate head and thorax of an ant. The lengthened pedicel, in combination with a slender abdomen with a constriction in the anterior part, simulates the petiole and postpetiole segments of an ant abdomen and also enhances the spiders’ resemblance to that of ants. Additionally, most Myrmarachne species bear a striking resemblance to a particular species or genus of an ant6 (Fig. 1). For example, Myrmarachne cornuta is an accurate mimic of ants belonging to the genus Tetraponera, which have an extremely slender and elongated body shapes7. In contrast, Myrmarachne maxillosa has a more oval and stocky-shaped body, matching the appearance of the ants belonging to the genus Polyrhachis8. Myrmarachne plataleoides is a well-known accurate morphological mimic of the weaver ant Oecophylla smaragdina and has a short and convex cephalothorax and abdomen similar to the body form of its ant model9. The accurate ant-like appearances of Myrmarachne spiders confer protection against predation by visually oriented predators10, such as mantids11,12, and other spiders13.

Studied species of Myrmarachne ant-mimetic jumping spiders, except M. malayana. (a) Myrmarachne acromegalis, (b) M. cornuta, (c) M. hashimotoi, (d) M. melanocephala, (e) M. maxillosa, and (f) M. plataleoides. (Photo credit: Hashimoto Y.).
Spiders are distantly related to insects and they have a completely different body plan. For example, unlike insects, spiders lack extensor muscles in major leg joints and instead use hydraulic pressure generated in their bodies to extend their legs for jumping14. To drive the hydraulic force for leg extension, spiders pump hemolymph fluid from the abdomen into the cephalothorax through the pedicel, and then increase the hemolymph pressure via a cephalothorax compression. In this hydraulic mechanism, sufficient body volume and hemolymph flow are required to produce a high fluid pressure15,16. Therefore, the slender and constricted ant-like appearances of Myrmarachne spiders are expected to constrain their jumping abilities. Indeed, earlier observations have suggested that, despite belonging to a jumping spider family, some species of Myrmarachne spiders were observed to be unable to jump and were only able to hunt prey in a close range17. However, such observations were based on a relatively few species and largely qualitative analyses. Furthermore, because each Myrmarachne species morphologically mimics the characteristics of a particular species or genus of an ant, the variations in the jumping abilities are expected to be associated with the variations in their ant-mimetic morphological traits. However, complex variation associated with body shape is a difficult type of variation to quantify18. Consequently, the relationship between the body shape and jumping ability in Myrmarachne spiders had not been explicitly tested before. In this study, we used geometric morphometric techniques (an approach that provides a mathematical description of biological forms) to quantify the body shapes of Myrmarachne spiders and test the hypothesis that a reduction in the jumping proficiency is associated with the shape adaptations related to ant mimicry. Since high species richness and morphological diversity of Myrmarachne spiders are found in tropical regions, particularly in Southeast Asia, where numerous ant species exist3,18, we collected Myrmarachne species with different body shapes and non-myrmecomorphic jumping spiders from three tropical forests in Southeast Asia. We then measured the jumping distances to capture prey and prey-capture success rates of seven Myrmarachne species and quantitatively compared the results with those of non-myrmecomorphic jumping spiders using geometric morphometrics. This study, for the first time, provides quantitative evidence to show that the accurate resemblance of Myrmarachne spiders to ants is likely to constrain their jumping ability for catching prey. In addition, our findings provide novel insights to further our understanding of the evolution and maintenance of accurate mimicry in ant-mimetic jumping spiders.
Results
Body shape comparisons
Canonical variate analysis (CVA) of the body shape of the seven Myrmarachne species (M. acromegalis, M. cornuta, M. hashimotoi, M. malayana, M. maxillosa, M. melanocephala, and M. plataleoides) and non-myrmecomorphic spiders (not identified to the species level but belonging to the same family, i.e., Salticidae) showed that the CVA1 axis clearly distinguished the mimics from non-mimics (Fig. 2a). Transformation grids, used to interpret shape changes associated with CVA1, demonstrated that the non-myrmecomorphic spiders (Non-mimics) were characterized by a broader cephalothorax and abdomen with a shorter pedicel than those of Myrmarachne spiders (Fig. 2b). The CVA2 axis revealed that the seven Myrmarachne species could be divided into two distinct groups: M. maxillosa and M. malayana had a positive score on the CVA2 axis (Group: Broad mimics), whereas the remaining five species had a negative score on the CVA2 axis (Group: Slender mimics). The body shape of the Slender mimics species was observed to be more slender and more elongated than that of the Broad mimics species, which had broader body forms. The discriminant function analysis (DFA) of the body shapes for pairwise comparisons among the Non-mimics group and the two groups of Myrmarachne species showed significant differences in body shape between the groups (Non-mimics vs. Broad mimics: the Mahalanobis distance = 9.51, T-square = 2204.17; Non-mimics vs. Slender mimics: the Mahalanobis distance = 7.89, T-square = 1968.28; Broad mimics vs. Slender mimics: the Mahalanobis distance = 6.73, T-square = 1048.02; Permutation tests of the Mahalanobis distances P < 0.001 for all comparisons; Fig. 3).

Differences in the body shape among Myrmarachne and non-myrmecomorphic jumping spiders. (a) Canonical variate analysis of seven Myrmarachne species and non-myrmecomorphic jumping spiders based on 16 landmarks of the body shape. Scatter plot of the first two canonical variate axes with 90% confidence ellipses (Non-mimics: Non-myrmecomorphic jumping spiders, MAC: M. acromegalis, MCO: M. cornuta, MHA: M. hashimotoi, MME: M. melanocephala, MMX: M. maxillosa, MMY: M. malayana, MPL: M. plataleoides). (b) Transformation grids illustrate the shape changes associated with the first two canonical variates in positive and negative directions with scaling factors of -9 to 9 for CV1 and -6 to 6 for CV2.

Discriminant function analysis for pairwise competitions among non-myrmecomorphic spiders (Non-mimics) and the two groups of Myrmarachne species (Broad mimics and Slender mimics). The mean body-shape differences of each group have been demonstrated with wireframe outlines (left). Cross validation scores of body-shape variables of each group have been shown as histograms (right). Permutation test with the T-square statistic for the Mahalanobis distance P < 0.001 for all comparisons. (Non-mimics: Non-myrmecomorphic jumping spiders; Broad mimics: M. maxillosa and M. malayana; Slender mimics: M. acromegalis, M. cornuta, M. hashimotoi, M. melanocephala, and M. plataleoides).
Jumping distance comparisons
Both groups of Myrmarachne species discriminated by CVA and DFA showed significantly shorter jumping distances than the Non-mimics, and the distances differed between the two groups (GLMM_g, χ2 = 102.63, df = 2, P < 0.001, Non-mimics: 2.81, CI = 2.50–3.12, Broad mimics: 1.00, CI = 0.80–1.21, Slender mimics: 0.68, CI = 0.57–0.79, the Tukey HSD test P < 0.05 for all comparisons; Fig. 4, Supplementary Table S1). The Slender mimics group presented shorter jumps than those exhibited by the Broad mimics group. The two-block partial least square (PLS) analysis revealed a high correlation between the body shape and jumping distance among the seven Myrmarachne species, indicating that the species with a shorter jumping distance typically had a slender body (RV coefficient = 0.699, P = 0.005; Fig. 5a, b).

GLMM_g predicted probabilities of jumping distances for non-myrmecomorphic spiders (Non-mimics) and the two groups of Myrmarachne species (Broad mimics and Slender mimics). The vertical bands represent the 95% confidence intervals of the expected jumping distances. The values in the graph represent the mean ± standard error of the expected values. Different letters assigned to the boxes denote significant differences (Tukey’s HSD post hoc test P < 0.05). (Non-mimics: Non-myrmecomorphic spiders; Broad mimics: M. maxillosa and M. malayana; Slender mimics: M. acromegalis, M. cornuta, M. hashimotoi, M. melanocephala, and M. plataleoides).

Relationship between the body shape and jumping distance of the seven Myrmarachne species. (a) Two-block partial least squares analysis (PLS) of the jumping distance against the mean body shape of the seven Myrmarachne species. The horizontal axis of the scatter plot represents the first PLS axis for body shape based on 16 landmarks and the vertical axis represents the first PLS axis for the jumping distance of the seven species. (MAC: M. acromegalis, MCO: M. cornuta, MHA: M. hashimotoi, MME: M. melanocephala, MMX: M. maxillosa, MMY: M. malayana, and MPL: M. plataleoides). (b) Transformation grids illustrate the shape changes associated with the PLS shape vector in positive and negative directions. (Box plots showing the variation in the jumping distances and wireframe outlines showing the mean body shapes of the seven Myrmarachne species have been provided in Supplementary Fig. S2).
Prey-capture success rate comparisons
Prey-capture success rate of Myrmarachne spiders were significantly lower than that of the Non-mimics (GLMM_b, χ2 = 17.77, df = 2, P < 0.001, Non-mimics: 0.74, CI = 0.63–0.83, Broad mimics: 0.67, CI = 0.46–0.82, Slender mimics: 0.38, CI = 0.27–0.50; Fig. 6, Supplementary Table S1). However, multiple comparisons showed no significant difference in the prey-capture rate between the Non-mimics and those in the Broad mimics group (Tukey’s HSD test P = 0.750). In contrast, the prey-capture success rate of the Slender mimics group was significantly lower than that of the Non-mimics group (Tukey’s HSD test P < 0.001).

GLMM_b predicted probabilities of prey-capture success rates for non-myrmecomorphic spiders (Non-mimics) and the two groups of Myrmarachne species (Broad mimics and Slender mimics). The vertical bands represent the 95% confidence intervals of the expected prey-capture success rates. The values shown in the graph represent the mean ± standard error of the expected values. Different letters assigned to the boxes denote significant differences (Tukey’s HSD post hoc test P < 0.05). (Non-mimics: Non-myrmecomorphic spiders; Broad mimics: M. maxillosa and M. malayana; Slender mimics: M. acromegalis, M. cornuta, M. hashimotoi, M. melanocephala, and M. plataleoides).
Discussion
This study provides, for the first time, quantitative evidence to support the hypothesis that the accurate morphological mimicry of ants by Myrmarachne jumping spiders may impose constraints on their jumping abilities and prey-capture success rates. Here, we employed geometric morphometric methods to quantify and discriminate the body shape variations in Myrmarachne and non-myrmecomorphic jumping spiders, and confirmed that Myrmarachne spiders had significantly slenderer and more elongated body shapes than non-mimics. The Myrmarachne spiders presented shorter jumping distances and lower prey-capture success rates than the non-myrmecomorphic jumping spiders, and the degree of these reductions was significantly correlated with the morphological changes resulting from the specific ant model selection. For example, among the Myrmarachne species investigated in the present study, M. cornuta, which mimics Tetraponera ants with an extremely slender body with elongated petiole, showed the shortest jumping distance to catch its prey. In contrast, M. maxillosa and M. malayana, which mimic Polyrhachis ants with more oval and stocky-shaped bodies, were able to jump to distances equal to their own body length to capture prey. Overall, these results suggested that reduction in jumping and prey-capture abilities of Myrmarachne spiders was associated with the morphological adaptations related to ant mimicry.
Given that Myrmarachne evolved from ancestor of a non-myrmecomorphic jumping spider5, the reduction in the jumping distance of Myrmarachne spiders may be explained by the unique jumping mechanism in spiders. The mechanism used by jumping spiders involves the use of a hydraulic catapult method in which spiders pump hemolymph fluid from the abdomen into the cephalothorax through the pedicel, and then increase the hemolymph pressure by compressing the cephalothorax15,16. In this hydraulic system, proper function requires an adequate enclosed space and fluid flow16. Therefore, the ant-like appearance of Myrmarachne spiders, such as narrow and constricted body with an elongated pedicel, seems unsuitable for generating the high hemolymph pressure required for jumping. Our geometric morphometric analysis, using a two-block PLS analysis, of Myrmarachne spiders further showed that the lower abilities of jumping were found in Myrmarachne species with narrower and more elongated forms. However, at present, no detailed studies have assessed the hydraulic mechanisms of jumping in Myrmarachne spiders. Thus, our assumption that the morphological ant mimicry by Myrmarachne spiders limits their hydraulic forces to jump should be addressed in future studies, for example, by analyzing the biomechanics of the hydraulic system in the mimetic spiders.
The present study showed that not only the jumping ability but also the prey-capture success rate was considerably reduced in Myrmarachne species with a slender body shape. As the results were only obtained under laboratory conditions, it is necessary to verify the prey-capture success rate under natural conditions via field observations. However, our results suggest the possibility that Myrmarachne species changed their diets in nature or may have evolved specialized prey-capture behavior to compensate for the poor jumping ability. Although none of the field evidence indicated that Myrmarachne species have a specialized predation behavior, such as snatching prey from ants as observed in Menemerus jumping spiders20, our previous study using stable isotope analysis demonstrated that Myrmarachne species with slender body shapes, such as M. hashimotoi, had low δ15N values similar to those of nectar-feeding ants and arthropod herbivores, but not similar to those of predators, in the same study sites21. In contrast, the δ15N values of Myrmarachne species with broad body shapes, such as M. malayana and M. maxillosa, were similar to those of omnivorous ants and non-myrmecomorphic jumping spiders. These findings indicated that plant-based diets were the main source of nutrition, especially for the species with slender body shapes, although some species of Myrmarachne have been observed to feed on the eggs of other spiders22. Jackson et al.22 and Nyffeler et al.23 reviewed previous studies on plant feeding by spiders and found that more than 40 species of jumping spiders, representing a dozen different subfamilies, were observed feeding on plant-derived food, such as nectar and honeydew, under natural conditions. Therefore, it is not surprising that some Myrmarachne spiders engage in phytophagy. However, plant-based diets are known to have a low protein content and are thus poor alternatives for animal prey24. If Myrmarachne spiders with slender body shapes indeed depend on plant-based diets as important food sources, this raises a further interesting question of whether the predominant plant feeding of Myrmarachne spiders interferes with their growth and fecundity. Clarification on the nutritional ecology of Myrmarachne spiders, with particular attention to their morphological resemblance to ants, will be a fascinating future research direction that will enable a better understanding of the costs of the mimetic resemblance.
In conclusion, our study showed that Myrmarachne spiders were less efficient at capturing prey than non-myrmecomorphic jumping spiders, based on observations under laboratory conditions. Moreover, our geometric morphometric methods allowed us to quantitatively analyze the body shapes of Myrmarachne spiders, which revealed that the variations in their body shapes, resulting from specific model-mimic resemblances, were strongly associated with a reduction in their jumping and prey-capture abilities. In particular, a significant reductions in prey-capture proficiency was observed in Myrmarachne species with a slender and elongated body shape, which may influence these species to be phytophagous. Overall, the present study supported our hypothesis that the accurate morphological mimicry of ants by Myrmarachne spiders constrained the jumping and prey-capture abilities. Although our research focused on the constraints placed upon the jumping and prey-capture abilities due to mimicry, the accurate resemblance between Myrmarachne spiders and ants may potentially interfere with other activities because selection for mimicry can entail radical phenotypic changes in the mimics. Indeed, previous studies have suggested that the mimicry of Myrmarachne spiders imposes costs in the form of detriment effects on their life-history traits, such as mating25,26 and locomotion27. Furthermore, the resemblance between Myrmarachne spiders and ants can be achieved by not only body-form modifications but also combining a variety of morphological changes, all of which may potentially impose constraints on the mimics. For example, Myrmarachne spiders have thin legs like ants instead of the stout legs of typical salticids; however, thin legs were also found in non-myrmecomorphic salticids28. As salticid spiders usually use their legs to grasp and hold onto prey29, the thin legs of Myrmarachne spiders may also interfere with their prey-capture success rates. If the fitness costs of close mimetic resemblance greatly outweighs the benefits in a mimic species, then inaccurate mimicry may be selected for10,30. Therefore, to better understand how and why the accurate morphological mimicry evolved and is maintained in Myrmarachne spiders, we acknowledge that more studies should focus on the constraints imposed by the accurate morphological mimicry of ants. The methods used in the present study can be used in future investigations to provide novel insights into the evolutionary costs of accurate morphological mimicry in ant-mimetic jumping spiders.
Methods
Study site and study animals
The field sites and laboratories at each site were located at the Danum Valley Field Centre (4°55′N, 117°40′E) and the Lambir Hills National Park (4°20′N, 113°50′E) in Malaysian Borneo, and at the Sakaerat Environmental Research Station (14°30′N, 101°56′E) in northeastern Thailand. Sampling was conducted during four sampling events at the Danum Valley Field Centre-September 2004 (7 days), February 2005 (6 days), December 2006 (6 days), and January 2008 (5 days); five sampling events at the Lambir Hills National Park–August 2007 (7 days), July 2008 (7 days), February 2009 (9 days), September 2009 (5 days), and December 2012 (5 days); and three sampling events at the Sakaerat Environmental Research Station–March 2013 (6 days), September 2013 (5 days), and September 2014 (5 days). The spiders were collected on the day before testing for their jumping ability and prey-capture success rate and were held individually in plastic tubes containing a piece of damp cotton wool without food in the laboratory. For the tests, we used adult females and subadults of the spiders because they naturally spend much of their time foraging and not seeking partners. The subadults, which are the last instar before reaching sexual maturity, have almost the same size as that of the adults31. Small flies (1.5–3.0 mm body length) were used as prey items, which were collected in the field by sweep-netting on the day of testing. Eighty-six individuals from seven Myrmarachne species (MAC: M. acromegalis, n = 9; MCO: M. cornuta, n = 6; MHA: M. hashimotoi, n = 29; MMY: M. malayana, n = 14; MMX: M. maxillosa, n = 10; MME: M. melanocephala, n = 10; MPL: M. plataleoides, n = 8) were used in this study. Recently, Prószyński segregated 12 genera from the genus Myrmarachne without considering synapomorphies, or quantitative or phylogenetic analyses32. In addition, the monophyly of “Myrmarachne” in Prószyński’s sense is not supported, and his idea has not been followed by subsequent researchers4,33. Therefore, in this study we followed the previous definition of the genus Myrmarachne. Seventy individual non-myrmecomorphic jumping spiders were used in this study, including 12 salticid genera and unknown genera of the subfamily Salticinae (Carrhotus, n = 1; Colyttus , n = 1; Cosmophasis, n = 1; Epeus, n = 7; Evarcha, n = 1; Parabathippus, n = 5; Phintella, n = 2; Ptocasius, n = 23; Rhene, n = 3; Simaetha, n = 3; Telamonia, n = 3; Thiania, n = 3; and Unknown, n = 17) (pictures of the studied genera of non-myrmecomorphic salticids are shown in Supplementary Fig. S1).
Geometric analysis
Geometric analyses were performed using photographs of the ventral view of each spider body, where it was easy to determine the anatomically homologous points in both the non-myrmecomorphic and Myrmarachne jumping spiders. All photographs of the 86 individuals of the seven Myrmarachne species and the 70 individuals of non-myrmecomorphic salticids were taken using an Olympus digital camera (E-1) on a trinocular mount of a Leica MZ16a stereo-microscope. In each photograph, we digitized 16 landmarks using the ImageJ software version 1.50i (National Institute of Health, Bethesda, MD)34 (Fig. 7). The raw landmark coordinates were superimposed using a generalized procrustes analysis (GPA) to scale all configurations to the centroid size, translate them to the same location, and rotate them to the same orientation. Following the GPA, to visualize the body shape differences and group separation among spider taxa, we used CVA. DFA was used to test the statistical accuracy of the group separation indicated by CVA. For this analysis, the Mahalanobis distances, T-square tests, cross-validation scores, and wireframe results have been reported for pairwise comparisons among the groups. Permutation tests (10,000 rounds) with the T-square statistic for the Mahalanobis distance were performed to test the null hypothesis of equal group means. Cross-validation scores were used to assess the classification accuracy. Wireframe graphs of the mean and displaced landmarks described the most significant shape differences. Finally, a two-block PLS analysis was performed to assess the degree of association between the body shape and jumping distance. The two-block PLS analysis is a method to find a linear combination between two sets of data that can be used to analyze the relationship between shape and function35. The analysis provides an RV coefficient, which is analogous to a squared correlation coefficient and is a measure of the strength of association between two sets of variables. A permutation test with 10,000 permutations was performed to assess the statistical significance of this coefficient against the null hypothesis of complete independence between the two blocks of variables. All analyses were performed using MORPHOJ software (version 1.06)36.

Ventral body views of non-myrmecomorphic (left) and Myrmarachne (right) jumping spiders showing the 16 landmarks used in this study.
Testing procedures to measure the jumping distance and prey-capture success
All tests were conducted in a plastic cup (diameter: bottom, 9 cm and top, 10 cm, and height: 3.6 cm) (see Supplementary Fig. S3 and Supplementary Movie S1). The cup had a clear plastic top and a hole (diameter 1 cm) on the side. For each trial, a single spider and a single prey were introduced into the arena through the hole in the side of the cup. During the testing, the hole was covered with a cotton plug. The jumping distance and prey-capture success of the 86 individuals of the seven Myrmarachne species and the 70 individuals of non-myrmecomorphic salticids were recorded for 3 min with a video camera (Panasonic NV-GS202, VDR-D300, and Sony HDR-CX590V) that was positioned above the arena. Jumping distance was measured from screen captures using Adobe Photoshop CC (Adobe Systems Inc) and the distance at which the spider launched its attack was measured between the head of the spider and the prey. The longest jump by each spider was selected and used in the data analysis. If spiders only performed one jump, this distance was used in the analysis. The relative jumping distance was used to compare the jumping distance of spiders, whereby the jumping distance (JD) was divided by the body length (BL). Each spider was used just once per test. The spiders that did not respond to the prey in their trials were not included in the analysis.
Statistical analysis
To compare the jumping distances and prey-capture successes between non-myrmecomorphic jumping spiders and the Myrmarachne species, we used generalized linear mixed models (GLMMs). Owing to the small sample size of each Myrmarachne species, we aggregated the seven species into groups that were discriminated by CVA and DFA, and included “species of Myrmarachne and genus of non-myrmecomorphic salticids” as a random intercept effect in the GLMMs to control for potential differences among species within each group. We used a GLMM with gamma distribution (GLMM-g) to compare the jumping distances, and GLMM with binomial distribution and logistic regression link (GLMM-b) to compare the prey-capture success. Analysis of the GLMMs was performed using the R package lme4 (version 1.1–21)37. A post hoc pairwise comparison of our GLMM estimated means was conducted using Tukey’s HSD test and the R package multcomp (version 1.4–10)38. All statistical analyses were performed using R (version 3.60)39.
References
Nelson, X. J. & Jackson, R. R. How spiders practice aggressive and Batesian mimicry. Curr. Zool. 58, 620–629. https://doi.org/10.1093/czoolo/58.4.620 (2012).
Hölldobler, B. & Wilson, E. O. The Ants (Harvard University Press, Cambridge, 1990).
Yamasaki, T. & Ahmad, A. H. Taxonomic study of the genus Myrmarachne of Borneo (Araneae: Salticidae). Zootaxa 3710, 501–556. https://doi.org/10.11646/zootaxa.3710.6.1 (2013).
Yamasaki, T. et al. New species of the ant-mimicking genus Myrmarachne MacLeay, 1839 (Araneae: Salticidae) from Sarawak, Borneo. Zootaxa 4521, 335–356. https://doi.org/10.11646/zootaxa.4521.3.24 (2018).
Maddison, W. P. A phylogenetic classification of jumping spiders (Araneae: Salticidae). J. Arachnol. 43, 231–292. https://doi.org/10.1636/arac-43-03-231-292 (2015).
Hashimoto, Y. et al. Pattern of co-occurrence between ant-mimicking jumping spiders and sympatric ants in a Bornean tropical rainforest. Raffles B. Zool. 64, 70–75 (2016).
Edmunds, M. & Prószyński, J. On a collection of Myrmarachne spiders (Araneae: Salticidae) from peninsular Malaya. Bull. Br. Arachnol. Soc. 12, 297–323 (2003).
Edmunds, M. Do Malaysian Myrmarachne associate with particular species of ant?. Biol. J. Linn. Soc. 88, 645–653 (2006).
Mathew, A. P. Observations on the habits of two spider mimics of the red ant, Oecophylla smaragdina (Fabr.). Bombay Nat. Hist. Soc. 52, 249–263 (1954).
Cushing, P. E. Spider-ant associations: an updated review of myrmecomorphy, myrmecophily, and myrmecophagy in spiders. Psyche (Camb. Mass.) 2012, 1–23. https://doi.org/10.1155/2012/151989 (2012).
Nelson, X. J. & Jackson, R. R. Vision-based innate aversion to ants and ant mimics. Behav. Ecol. 17, 676–681. https://doi.org/10.1093/beheco/ark017 (2006).
Ramesh, A., Vijayan, S., Sreedharan, S., Somanathan, H. & Uma, D. Similar yet different: differential response of a praying mantis to ant-mimicking spiders. Biol. J. Linn. Soc. 119, 158–165. https://doi.org/10.1111/bij.12793 (2016).
Huang, J. N., Cheng, R. C., Li, D. & Tso, I. M. Salticid predation as one potential driving force of ant mimicry in jumping spiders. Proc. R. Soc. B 278, 1356–1364. https://doi.org/10.1098/rspb.2010.1896 (2011).
Parry, D. A. & Brown, R. H. J. The hydraulic mechanism of the spider leg. J. Exp. Biol. 36, 423–433 (1959).
Kropf, C. Hydraulic system of locomotion. In Spider Ecophysiology (ed. Nentwig, W.) 43–56 (Springer, 2013). https://doi.org/https://doi.org/10.1007/978-3-642-33989-9_4
Liu, C., Wang, Y., Ren, L. & Ren, L. A review of biological fluid power systems and their potential bionic applications. J. Bionic. Eng. 16, 367–399. https://doi.org/10.1007/s42235-019-0031-6 (2019).
Jackson, R. R. The biology of ant-like jumping spiders (Araneae, Salticidae): prey and predatory behaviour of Myrmarachne with particular attention to M. lupata from Queensland. Zool. J. Linn. Soc. 88, 179–190 (1986).
Adams, D. C., Rohlf, F. J. & Slice, D. E. Geometric morphometrics: ten years of progress following the ‘revolution’. Ital. J. Zool. 71, 5–16. https://doi.org/10.1080/11250000409356545 (2004).
Metzner, H. Jumping spiders (Arachnida: Araneae: Salticidae) of the world. https://www.jumping-spiders.com (2020).
Jackson, R. R., Salm, K. & Pollard, S. D. Snatching prey from the mandibles of ants, a feeding tactic adopted by East African jumping spiders. J. Arachnol. 36, 609–611. https://doi.org/10.1636/ST07-55.1 (2008).
Hyodo, F. et al. Stable isotope analysis reveals the importance of plant-based diets for tropical ant-mimicking spiders. Entomol. Sci. 21, 461–468. https://doi.org/10.1111/ens.12331 (2018).
Jackson, R. R., Pollard, S. D., Nelson, X. J., Edwards, G. B. & Barrion, A. T. Jumping spiders (Araneae: Salticidae) that feed on nectar. J. Zool. 255, 25–29. https://doi.org/10.1017/S095283690100108X (2001).
Nyffeler, M., Olson, E. J. & Symondson, W. O. Plant-eating by spiders. J. Arachnol. 44, 15–28. https://doi.org/10.1636/P15-45.1 (2016).
Lundgren, J. G. Nutritional aspects of non-prey foods in the life histories of predaceous Coccinellidae. Biol. Control. 51, 294–305. https://doi.org/10.1016/j.biocontrol.2009.05.016 (2009).
Nelson, X. J., Li, D. & Jackson, R. R. Out of the frying pan and into the fire: a novel trade-off for Batesian mimics. Ethology 112, 270–277. https://doi.org/10.1111/j.1439-0310.2006.01155.x (2006).
Nelson, X. J. & Jackson, R. R. Compound mimicry and trading predators by the males of sexually dimorphic Batesian mimics. Proc. R. Soc. B 273, 367–372. https://doi.org/10.1098/rspb.2005.3340 (2006).
Shamble, P. S., Hoy, R. R., Cohen, I. & Beatus, T. Walking like an ant: a quantitative and experimental approach to understanding locomotor mimicry in the jumping spider Myrmarachne formicaria. Proc. R. Soc. B 284, 20170308. https://doi.org/10.1098/rspb.2017.0308 (2017).
Pekár, S. Is inaccurate mimicry ancestral to accurate in myrmecomorphic spiders (Araneae)?. Biol. J. Linn. Soc. 113, 97–111. https://doi.org/10.1111/bij.12287 (2014).
Forster, L. M. A qualitative analysis of hunting behaviour in jumping spiders (Araneae: Salticidae). N. Z. J. Zool. 4, 51–62. https://doi.org/10.1080/03014223.1977.9517936 (1977).
Holen, Ø. H. & Johnstone, R. A. The evolution of mimicry under constraints. Am. Nat. 164, 598–613. https://doi.org/10.1086/424972 (2004).
Bartos, M., Szczepko, K. & Stanska, M. Predatory response to changes in camouflage in a sexually dimorphic jumping spider. J. Arachnol. 41, 381–386. https://doi.org/10.1636/Hi13-02.1 (2013).
Prószyński, J. Delimitation and description of 19 new genera, a subgenus and a species of Salticidae (Araneae) of the world. Ecol. Mont. 7, 4–32 (2016).
Maddison, W. P. & Szüts, T. Myrmarachnine jumping spiders of the new subtribe Levieina from Papua New Guinea (Araneae, Salticidae, Myrmarachnini). ZooKeys 842, 85–112. https://doi.org/10.3897/zookeys.842.32970 (2019).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. https://doi.org/10.1038/nmeth.2089 (2012).
Rohlf, F. J. & Corti, M. Use of two-block partial least-squares to study covariation in shape. Syst. Biol. 49, 740–753. https://doi.org/10.1080/106351500750049806 (2000).
Klingenberg, C. P. MorphoJ: an integrated software package for geometric morphometrics. Mol. Ecol. Resour. 11, 353–357. https://doi.org/10.1111/j.1755-0998.2010.02924.x (2011).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Soft. 67, 1–48. https://doi.org/10.18637/jss.v067.i01 (2015).
Hothorn, T., Bretz, F. & Westfall, P. Simultaneous inference in general parametric models. Biom. J. 50, 346–363. https://doi.org/10.1002/bimj.200810425 (2008).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/ (2019).
Acknowledgements
We thank the management and staff of the Institute of Tropical Biology and Conservation, University Malaysia Sabah for the provision of field and laboratory facilities, and the management and staff of the Danum Valley Management Committee and the Danum Valley Field Centre for their guidance and assistance, both in the field and at the Research Centre. Our study was conducted in accordance with the Memorandums of Understanding signed between University Malaysia Sabah and the Museum of Nature and Human Activities, Hyogo. We are grateful to the staff of the Forest Department of Sarawak (FDS) and the Sarawak Forestry Corporation for their permission to conduct research in Sarawak, to the National Research Council of Thailand (NRCT) for their permission to perform our research in the Sakaerat Environmental Research Station, and to Taksin Artchawakom (The Thailand Institute for Scientific and Technological Research) for the logistic arrangements in the station. We would also like to thank Editage for English language editing. Finally, we are grateful to Dr. Robert R Jackson and the referees for useful comments. This study was supported by the JSPS KAKENHI (Grant numbers: 16657028, 19570094, 24570109, and 17H01912) and by JST/JICA, SATREPS (JPMJSA1902).
Author information
Authors and Affiliations
Contributions
Y.H. conceived and designed the experiments. Y.H., T.E., T.Y., F.H., and T.I. performed the sampling and the experiments. T.Y. identified the Myrmarachne spiders to the species level and the non-myrmecomorphic salticids to the genus level. Y.H. analyzed the data and wrote the paper. All authors reviewed and accepted the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hashimoto, Y., Endo, T., Yamasaki, T. et al. Constraints on the jumping and prey-capture abilities of ant-mimicking spiders (Salticidae, Salticinae, Myrmarachne). Sci Rep 10, 18279 (2020). https://doi.org/10.1038/s41598-020-75010-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-75010-y
This article is cited by
-
Relationship between model noxiousness and mimetic accuracy in myrmecomorphic spiders
Evolutionary Ecology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
