
Abstract
The extensive peat bogs of Southern Scandinavia have yielded rich Mesolithic archaeological assemblages, with one of the most iconic artefacts being the bone point. Although great in number they remain understudied. Here we present a combined investigation of the typology, protein-based species composition, and absolute chronology of Maglemosian bone points. The majority of the bone points are made from cervids and bovines. However, changes both in species composition and barb morphology can be directly linked to a paucity of finds lasting nearly 600 years in Southern Scandinavia around 10,300 cal BP. We hypothesize that this hiatus was climate-driven and forced hunter-gatherers to abandon the lakes. Furthermore, the marked change in bone points coincides with a change in lithic technology. We, therefore, propose that the Maglemose culture in Southern Scandinavia is fundamentally divided into an Early Complex and a Late Complex.
Similar content being viewed by others


Introduction
The biological and geological record of the transition from the Late Glacial to the Early Holocene is manifested by a dramatic change in vegetation due to climatic warming1. With the increase in temperature vast amounts of buried stagnant ice gradually melted forming water-filled depressions. The Early Holocene landscape was therefore characterized by numerous shallow lakes and ponds in a relatively open birch and pine-dominated woodland2. These formed crucial hunting and fishing grounds for the first Maglemosian people living in Southern Scandinavia.
Throughout the Early Mesolithic barbed bone points were frequently lost in the lakes, presumably in connection with spearfishing. These lakes gradually evolved into bogs where peat accumulated. Fuel shortages, especially during the First and Second World Wars, resulted in industrialized peat exploitation in these bogs, which in turn caused these artefacts to be uncovered and recognized for what they are3,4,5.
The barbed bone points soon became closely associated with the Maglemose culture (c. 11,000–8000 BP). However, despite a long history of research on the typology of these characteristic items6,7,8, their chronological placement has largely been indirectly deduced from stratigraphy and pollen dating4,9. Species identification of bone points has previously been based on such evidence as bone debitage from habitation sites, or the absence of specific skeletal elements in a given faunal assemblage3,10, p. 278,11, which makes the identification of each artefact circumstantial. Based on these types of analyses, the majority of the Maglemosian bone points were thought to have been made from both ribs and long bones of “large ungulates”, translating to aurochs, elk, red- and roe deer (Bos primigenius, Alces alces, Cervus elaphus, and Capreolus capreolus)12.
We analyzed 126 bone points from Southern Scandinavia (Fig. 1 and Supplementary Information, Section 1) using a combination of morphological typology, radiocarbon dating, and proteomic analyses. Using these techniques, we were able to assess the selection of raw materials and date the typological variation. This study is the first attempt to investigate whether a single artefact type can be used as a proxy for both human and animal populations. We demonstrate that such comprehensive analyses can provide information about species dynamics, resource exploitation, human–environment interactions, and perhaps population mobility. Our integrated analyses provide a robust new framework for Maglemose chronology, that extends to changes in lithic manufacture.

Overview of the approximate find locations for the 126 barbed bone points from Southern Scandinavia. All bone points found in paleolake systems, now grown into peat bogs. While the selection is finite the spread indicates the approximate Eastern → Western extent of sediments conducive for preservation (for further information on each artefact including species identifications see Supplementary Dataset 1 and Supplementary Figures 1–22 and 29–30) (Map: digital elevation model was produced using open source Copernicus data and information (from the European Union—EU-DEM layers), and then merged with spatial geographical data in QGIS v.3.19 (www.qgis.org), followed by a final correction in Adobe Illustrator v. 24.3 (www.adobe.com).
Results
Bone points
Local bone point typo-chronologies have previously been proposed for two sites in Sweden, i.e. Rönneholms mosse8 and Motala13. However, since the present study comprises material from a much wider geographical region, this fine-scale classification of the bone points might not be representative. Therefore, to reduce bias, we divided the material into two groups; fine-barbed bone points and large-barbed bone points.
Radiocarbon dating
Prior to this study, only five radiocarbon (14C) dates on Danish barbed bone points had been published14,15. With the addition of previously published dates from Scania in Sweden (n = 20)8, unpublished dates from Denmark (n = 7) as well as 21 new 14C measurements acquired specifically for this project, we were able to model the radiocarbon-date distribution of 50 bone points (excluding double dates on the same artefact) (Fig. 2; Table 1; Supplementary Information, section 2; Supplementary Dataset 1). After calibration into cal. years BP in OxCal v.4.316, the artefacts separated into two distinct phases (Fig. 2). The fine-barbed bone points are all confined to the mid-late Preboreal and the beginning of the Boreal (c. 11,200–10,100 cal BP). The larger-barbed bone points are restricted to the end of the Boreal and the beginning of the Atlantic chronozones (9658–8413 cal BP). Between the fine-barbed bone points and the larger-barbed bone points, there is a clear gap in the radiocarbon ages lasting nearly 600 years. In order to explore this hiatus, we summed the radiocarbon dates and performed a Kernel Density Estimation (KDE) simulation to explore periods of activity. This indicates temporal and spatial morphological patterns and confirms the age correlated distributions of the two types of bone points in both regions (Fig. 2).

Radiocarbon dates from bone points showing the separation of the two types. (a) A Bayesian model assuming two phases performed on 52 bone points. Double dates on the same artefact marked with *. Carbon distributions colored to denote finding the place or phase boundaries, (b) Summed radiocarbon dates of bone points (shown in light grey), KDE to visualize activity and hiatus (light blue), boundaries marked in dark grey, (c) Summed radiocarbon dates from habitation in Denmark (see Supplementary Figure 23) (shown in light grey), KDE to visualize activity and hiatus (light blue), boundaries marked in dark grey, (d) biozones (Preboreal onset after Jessen et al.1)(e) the two bone point complexes separated by hiatus (see Table 1 and Supplementary Dataset 1).
Published and unpublished radiocarbon dates from the faunal remains of Eurasian elk (n = 73), red deer (n = 33), and bovines (aurochs + bison Bison bonasus, n = 73) reveal reduced frequencies in the faunal remains of these species around c. 10,000 cal. BP in Denmark (see Supplementary Figure 24; Supplementary Information, Section 2; Supplementary Dataset 2). However, no equivalent decline in the dates from Southern Sweden has been observed. If this decline in radiocarbon-dated remains from Eastern Denmark is representative of a decline in the animal populations this would have implications for the availability of raw material for the manufacture of bone points. The absence of bone points also corresponds to a partial absence of dated habitation sites in Eastern Denmark (not including the island of Bornholm) and Jutland (Fig. 2). However, it is during this same gap in the Danish record, that most of the dated habitation sites on the west coast of Sweden occur, as well as on the island of Bornholm (see Supplementary Figures 23 and 24; Supplementary Information, section 2; Supplementary Dataset 3).
Protein analysis
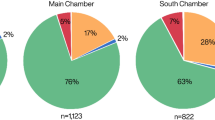
One-hundred and twenty barbed bone points were analyzed by ZooMS17 (see Supplementary Information, section 3, Supplementary Dataset 4). They turned out to derive from three groups of mammals: 74 from cervids, 43 from bovines and three from brown bear (Ursus arctos) (Fig. 3).

Species identifications of bone points. (a) Histogram of 120 summed protein mass spectrometry identifications separated by radiocarbon hiatus (see Supplementary Dataset 4). Each column refers to barbed bone points made of respectively long bone and rib. Colours refer to stylized animal portraits. Gradient colours (blue vs. orange) indicate either red deer or elk, (b) selected bone points of each class.
Similarly to aurochs and bison18, red deer and elk could not be distinguished using previously published markers, e.g., from Welker et al.17. However, through mining of published whole-genome data, we were able to construct more complete sequences for the species revealing five single amino acid polymorphisms (SAPs) between red deer and elk resulting in five potential tryptic peptides (biomarkers) (Supplementary Table 1; Supplementary Information, section 4). In three of these biomarkers, one or both sequences also matched (100%) with environmental bacterial sequences as identified using BLASTp and were discarded (Supplementary Information, section 4). LC–MS/MS analyses of four reference samples (two red deer and two elks) confirmed that the two remaining biomarkers can be used to discriminate between the two species (Supplementary Information, section 4). In the MALDI spectra, the red deer peptide m/z 2216 (GETGPAGRPGEVGPPGPPGPAGEK, peptide COL1A1T66/67, position 910-934) results from a missed cleavage because the A → P substitution reduces the efficiency of tryptic cleavage at the adjacent arginine19. As m/z 2216 is also present in aurochs we can only use this marker to discriminate red deer once aurochs has been discounted. Unfortunately, masses corresponding to the equivalent sequence in elk were not detected in any of the MALDI spectra, so we can not confirm that samples lacking the m/z 2216 are elk using ZooMS. Of the 74 bone points previously identified as red deer/elk, 22 were reassigned to red deer. Two Preboreal bone points analyzed by LC–MS/MS (VHM13821 and A37811) had diagnostic peptides for elk, while two from the Boreal period (A40894 and A42422) were identified as red deer, confirming the earlier MALDI-MS based identification (Supplementary Table 2).
Discussion
Radiocarbon dates from Denmark and Scania (southern Sweden) revealed a hiatus starting at c. 10,300 cal BP, lasting nearly 600 years that separates the Maglemose period into two complexes represented by bone points of a markedly different form. Protein analysis revealed that the fine-barbed points (i.e. Preboreal and Early Boreal) were also materially different from the larger-barbed bone points (i.e. Late Boreal).
Fine-barbed points of the Early Maglemose Complex (n = 45) were predominantly manufactured from cervid long bones (see Fig. 3) despite evidence for the presence of other large ungulates—aurochs, bison, reindeer (Rangifer tarandus), and wild horse (Equus ferus)—in Southern Scandinavia at this time20,21,22. This overrepresentation of cervids may be a reflection of the species abundance at the time but more likely represent a deliberate choice by Early Maglemosian hunter-gatherers. Seven of the 45 were identified as red deer based on our novel biomarker. One of the seven was also radiocarbon dated (Øresø Mølle, A52426, 9481 ± 47) and represents the earliest occurrence of red deer in Denmark.
The bone point morphology of the Late Maglemose Complex changes to a larger and more varied barb design. The number of bone points made of bovine ribs increases substantially (from 6 to 93%) in this large-barbed (n = 75) assemblage (Fig. 3) and those of brown bear appears for the first time. All confirmed bison remains from Denmark and Southern Sweden have been radiocarbon dated to no later than the Preboreal20,23. This seemingly leaves aurochs as the sole persisting bovine species and assumed source of bone points produced after the Preboreal. The bone points made from the aurochs ribs represent the very last remnants of this species before its disappearance on the Danish islands c. 7000 cal BP24,25. The selection of aurochs rather than cervid ribs probably reflects its preferable mechanical properties (Supplementary Information 5). It could, therefore, be argued that the selection of the raw materials by Maglemosian hunter-gatherers was based upon practical, rather than for example spiritual, considerations26.
The majority of the large-barbed bone points date to the Maglemose culture, while seven dates extend into the beginning of the Kongemose culture (c. 8500–7400 cal BP), after which simple bone points without barbs seem to have been preferred (at least in Central Scania)27,28. The transition from large-barbed bone points to simple bone points seems to be synchronous with the arrival of trapezoid lithic armaments29,30,31. This change in material culture may reflect a change of economy and seasonal rounds of the local population: coastal and inland groups merging within Southern Scandinavia as sea levels rose; seasonal spearfishing in the lakes losing its importance by communities that relocated their demographic centres to the coasts during the Kongemose32. However, the transition to simple bone points and Blak type trapezoid armaments might also represent the arrival of new migrants, whose new technologies ended the microlithic tradition completely.
Interestingly, the hiatus of dated barbed bone points is also evident in the radiocarbon dates from the classic Maglemosian habitation sites in Eastern Denmark, in effect dividing them into two periods of occupation (see Fig. 2 and Supplementary Figure 23, Supplementary Information, section 2; Supplementary Dataset 2). Further, a similar gap in radiocarbon dates of faunal remains further indicates a decline in population size in Eastern Denmark, but interestingly not in Scania (apart from Rönneholms mosse) (see Supplementary Figure 24, Supplementary Information, section 2; and Supplementary Dataset 3). The δ13C values obtained from the bone points indicate vegetational change over time from an open environment to a more closed setting (see Supplementary Information, sections 6 and 8). This is unlikely to represent a behavioural change in the animals, but instead probably reflects a landscape characterized by denser forests.
The gap in radiocarbon dates during the Early Holocene might be interpreted as evidence for a decline in human habitation in Eastern Denmark. There is only weak evidence for human presence during the ‘bone point hiatus’. The 14C dates from the two settlement sites of Draved and Klosterlund in Jutland fall before and partly during the hiatus33,34. However, the radiocarbon dates from these sites are conventional and from the very early days of radiocarbon dating when samples were not processed to remove secondary humic acids; a factor that can often result in misleadingly young dates35. Five AMS dates from uniserial bone harpoon points date to the hiatus in Southern Scandinavia. These are from Tunebjerg Øst (9050 ± 40 14C BP) and Trunderup Mose (8845 ± 60 14C BP), both sites on Funen36, from Rönneholms mosse in Scania (8610 ± 90 14C BP)37 and from Vallensgård Mose on the present-day island of Bornholm in the Baltic Sea (9250 ± 60 14C BP, 8875 ± 65 14C BP)38, which would have been connected to Continental Europe at this time.
When large-barbed bone points appear following the hiatus (at c. 9650 cal BP), they show markedly different morphological traits, from their predecessors. They appear once the former major lakes fill and deepen again. Most of the smaller ponds would probably at this time, have grown into fens. Until recently, it was believed that the new bone point morphology appeared before the first indications of the pressure blade lithic industry in Denmark at c. 9000 cal BP39. This industry is characterized by small regular blades, created by applying pressure rather than direct percussion. Blades of this type were utilized as cutting-edge inserts in slotted bone points. However, based on a previously unpublished radiocarbon date obtained from a slotted bone implement (Clarks Type B1 or B2) from Fugle Å, near Ulkestrup Lyng in Store Åmose (AAR-11949, 8360 ± 55 14C BP) (see Fig. 2 and Supplementary Figure 29); pressure flaking is now contemporary with the emergence of large-barbed bone points.
The reason for this gap in the radiocarbon record which seemingly separates two cultural traditions is currently unknown. Several factors could have caused a small and confined human population to almost disappear from the archaeological record; i.e. epidemics, warfare, changes in subsistence strategy, climate, and migration. However, most of these factors are difficult to tie to the disappearance of bone points in the Southern Scandinavian lakes. Several local studies have revealed climatic fluctuations in the Northern Hemisphere during the Early Holocene including lower temperatures and increased precipitation40,41,42,43,44,45,46,47. To our knowledge, no evidence of a similar cool interval or increase in precipitation between c. 10,300 to 10,000 cal BP has been reported for Southern Scandinavia. This may be due to a lack of high-resolution studies of sediments from this period coupled with a sampling bias, further complicated by contemporaneous erosion events48,49,50.
High-resolution studies coupled with direct radiocarbon dating of sediment cores conducted in Southern Sweden, however, do not show an increase in precipitation in lake levels51, 52 at that time. Rather, they indicate dry climate and the lowest water levels of the entire Holocene during the early Boreal51,52,53. The northern expanse of hazel (Corylus avellana) at this time1 is also believed to have been facilitated by the markedly lowered water levels in the lakes and fens20. Pollen analyses of the Åmose basin on Zealand and Rönneholms mosse in Scania also show several marked water level changes that occurred during the Early Holocene48,49, although these are not correlated by direct radiocarbon dating of the sediments. During the transition from the Preboreal to the Boreal, a brief, but significant lowering of the water levels took place, which was shortly followed by renewed transgression54. In Åmosen, this regression led to erosion of the littoral zone and the redeposition of sediments, basically removing most Preboreal riparian sites and sediments associated with this period50. As in Scania, the lowering of the water level was also followed by a transgression48. The reason for these water level fluctuations is difficult to determine. One factor during the Early Preboreal could be the melting of stagnant ice, whereby glacier ice melted in situ. Temperature oscillations may also account for some of these water-level changes, with dryer climatic conditions resulting in increased evaporation55. “The general notion of a particularly dry climate during this period also seems to be reflected in sediment cores from Eastern Denmark, Southern Sweden and Western Germany, which all contain increased levels of charcoal, argued to have been caused by wildfires48, 56,57. Similarly, Jørgensen observed pollen from the shrub Ephedra sp. in Åmosen at the interphase between the Preboreal and the Boreal periods. Ephedra is known to thrive in arid environments, indicating that Åmosen might have been arid at this time period, although he also argues that it could have been transported by the wind48. If the climate did indeed force hunter-gatherers to relocate and abandon their old activity areas, these actions might be reflected in radiocarbon dates elsewhere in the form of potential bust and boom cycles and a rapid diffusion of technologies.
Interruption in 14C dates or drastic changes in the archaeology elsewhere in Europe is also centred around 10,000 cal. BP. In Eastern Fennoscandia, a c. 200-year long gap seems to occur shortly after 10,300 cal. BP, which has been interpreted as a decline in the local population as a result of an abrupt climate event58. Radiocarbon dates of human skeletal remains in Central Germany also reveal a similar gap, indicating either an absence of humans in the period or potential preservation or sample bias59. Interestingly, in North-western Europe this period marks a radical change in microlithic technology contemporaneous with increased droughts and severe wildfires in the region60. In Southern Norway, a similar diffusion event occurred, where pressure blade technology suddenly appeared61. These two studies testifiy to the notion that the environment must have affected the Early Mesolithic populations of Northern Europe.
The falling water levels62 would almost certainly have impacted the fishing and hunting opportunities and may have caused humans to move away in search of viable fishing grounds. This may account for the observed decrease of butchered animal remains discarded in environments conducive for preservation. However, ecological stress caused by increasing temperatures, coupled with potential wildfires in drying mixed coniferous forest, may also have triggered the movement of humans and animals alike.
The presence and continued use of fine-barbed bone points in Northern Germany during the hiatus12,63 suggest that fishing practices were not as disrupted further to the south. While dates from the Swedish west coast; Huseby Klev and Balltorp64,65,66 and a further 10 AMS dates from charred remains from the habitation site of Ålyst on Bornholm (Supplementary Figure 24; Supplementary Dataset 2), also suggest the arrival of new groups there. Subsistence at the Swedish sites focused primarily on marine mammals, fish, and birds67. Unfortunately, the coastlines of the Early Holocene in Denmark became submerged during the Atlantic period due to rising sea levels (see Supplementary Information 7), meaning that Early Mesolithic coastal sites are rarely located and excavated in this region68,69. Consequently, it is not possible to study and compare coastal settlements which could corroborate a shift in economic strategy and subsistence on a regional scale during the Early Mesolithic in this area. However, human exploitation of marine resources along coastal environments in Southern Scandinavia can be inferred from stable isotope data obtained from human remains. Individuals from Køge Sønakke off the coast of Eastern Zealand in Denmark70, Österöd71 and Huseby Klev65,72 from the South-western coast of Sweden are contemporaneous with the identified hiatus. These individuals yielded collagen δ13C and δ15N values consistent with partially or fully marine diet (Supplementary Dataset 2). Indeed the presence of harpoons during the Mesolithic has strongly been tied to the hunting of marine mammals, although the fact that they ended up in the potentially dried up lakes may indicate a more varied use36.
In summary, radiocarbon dates show a hiatus, spanning c. 600 years, separating the two classified types of bone points at c. 10,300 cal BP. The hiatus of dated barbed bone points are also evident in the radiocarbon dates from the habitation sites in Eastern Denmark, in effect dividing the Maglemose into two complexes, displaying distinctly different technologies; fine-barbed bone points and percussion knapping in the Early Maglemose Complex, and large-barbed bone points and pressure flaking in the Late Maglemose Complex. These two complexes represent two radically different material cultures and technological traditions which challenges the notion of a period in relative stasis. We are confident that this surprising gap in the South Scandinavian material will find more parallels elsewhere in Europe.
The cause of this radiocarbon gap is currently unknown, but we hypothesize that climate change caused water-levels to drop and thereby forced the Mesolithic people to relocate and adapt their subsistence strategies, potentially along the now submerged coasts. Another possibility coupled to low water levels could be the increase of wildfires due to an exceedingly dry climate, which may also have forced hunter-gatherers to relocate.
The introduction of pressure blade technology is now inferred to be synchronous with the emergence of large-barbed bone points after the hiatus and may indicate the transmission of influences from Sweden. Species identification by ZooMS and LC–MS/MS indicates that a conscious selection was carried out in regards to the specific species (as well as the preferred skeletal elements, i.e. rib or long bone) for manufacture.
The pressure blade industry which characterises the Late Maglemose Complex is thought to have originated at c. 20,000 cal BP in the area of present-day Siberia/Northern China from where it spread westward to Western Russia73, and subsequently into Northern Fennoscandia39,74,75. Results from archaeological studies on the diffusion of pressure blade technology correspond well with genetic studies from Norway and Sweden. These have shown that the individuals involved in the spread of pressure blade technology, were genetically admixed between Western and Eastern Hunter-Gatherers76, and thus indicate migration routes westwards from Russia into the Scandinavian peninsula while the route into Denmark remains inconclusive75 as it could also have spread through Poland and Germany. Most recently, re-analyses of lithic remains attributed to the oldest sequence at Huseby Klev (deep pit) revealed the presence of pressure blade technologies, which are securely dated to the hiatus and the subsequent decades (10,040–9610 cal BP). Moreover, the same study also reported on aDNA extracted from chewed birch pitch revealing that the individuals were of genetically admixed ancestry77. This novel lithic technology coupled with a changed bone point morphology and the pattern of radiocarbon dates from Western Sweden and Central Scandinavia78 favours the spread of this technology through Scania and into present-day Denmark.
It is hoped that future high-resolution sedimentary studies are conducted to elucidate if and in what ways the local climate affected the people inhabiting this area. Similarly, direct radiocarbon dates of material from classic Maglemosian sites, as well as genetic studies, will hopefully help to illuminate whether the Early and Late Maglemose Complex are also genetically as well as technologically distinct.
Methods
Materials
The bone points analyzed (n = 127) all derive from Southern Scandinavia (Denmark and Southern Sweden) (see Supplementary Information 1). The bone points were, based on barb morphology and skeletal element used in their manufacture (i.e. long bone versus rib), and to some extent species, divided into two groups.
A subset of the bone point samples was further submitted for AMS radiocarbon (14C) dating (n = 23), protein analysis to determine the species of each artefact (n = 120), and carbon (δ13C) and nitrogen (δ15N) isotopic analyses (n = 19). This has resulted in two distinctive groups extending from the Preboreal into the Atlantic Period, corresponding to the Maglemose and slightly beyond, into the subsequent Kongemose culture.
Radiocarbon dating
We submitted bone powder or bone fragments (mean weight 100 mg) of 23 bone points from Denmark for AMS dating at the Oxford Radiocarbon Accelerator Unit based on their typological grouping. Collagen was extracted from bone powder using ORAU pretreatment codes AF (samples A44111, 436, 487 and 534) and AG (all other samples79. The extracted collagen was combusted, graphitised and dated according to Dee and Ramsey80,81 and Ramsey et al.80,81. Of the 23, 21 were successfully dated and were subsequently merged with 24 published AMS dates8,14,15 as well as seven unpublished dates from Zealand (see Supplementary Dataset 1). Two artefacts (FP1469: 9375 ± 45 14C BP, 9208 ± 55 14C BP, and Brokøb B: 7940 ± 65 14C BP, 7890 ± 65 14C BP) were dated twice. Isotopic analysis of the dated collagen samples was conducted offline using combustion IRMS at the Oxford Radiocarbon Accelerator Unit, as were a further five samples dated at The Tandem Laboratory at Uppsala University for which sufficient collagen remained (Supplementary Dataset 1).
The total of 52 AMS dates was then calibrated to cal. years BP in OxCal v.4.3 using IntCal3 calibration curve16,82. We also compiled 68 published and unpublished radiocarbon dates performed on charcoal and bone associated with habitation sites, including human remains not directly associated with habitation (Supplementary Information 2), and 118 published and unpublished radiocarbon dates from faunal remains (elk n = 33, red deer n = 12, auroch and bison n = 73) spanning the Maglemose to infer presence or absence (Supplementary Information 2). We applied Bayesian phase modelling on the dates from habitations, assuming the coeval age of habitation events using OxCal v.4.382 (see Supplementary Dataset 3 and Supplementary Figure 25).
Proteomics
We performed ZooMS on 120 bone points from Denmark and Scania in Southern Sweden using protocols from83,84 (see Supplementary Information 3). Mass spectrometry was conducted on a Bruker MALDI-TOF–MS/MS instrument in reflector mode to acquire spectra from 800 to 3500 m/z. Taxonomic identification was completed using published markers17. As red deer (Cervus elaphus) and European Elk (Alces alces) cannot be distinguished with published markers85, we compared their collagen (COL1ɑ1 and COL1ɑ2) sequences and identified five single amino acid polymorphisms (SAPs). We then analyzed these SAPs to see if they provided unique tryptic markers for ZooMS analysis (see Supplementary Information 4).
To confirm our candidate ZooMS marker capable of discriminating between red deer and elk, four reference samples (two from each of these species) were digested with Trypsin, Elastase and Chymostrypsin and sequenced using LC–MS/MS (see Supplementary Information 3). In addition, we sequenced the tryptic peptides from four bone points, two from the Preboreal and two from the Boreal (see Supplementary Information 3).
Data availability
The mass spectrometry data for both ZooMS and LC-MS/MS have been deposited to the ProteomeXchange Consortium via the PRIDE86 partner repository with the dataset identifier PXD018050.
References
Jessen, C. A. et al. Early Maglemosian culture in the Preboreal landscape: archaeology and vegetation from the earliest Mesolithic site in Denmark at Lundby Mose Sjælland. Quat. Int. 378, 73–87 (2015).
Mortensen, M. F., Henriksen, P. S., Christensen, C., Petersen, P. V. & Olsen, J. Vegetation development in south-east Denmark during the Weichselian Late Glacial: palaeoenvironmental studies close to the Palaeolithic site of Hasselø. Danish J. Archaeol. 3, 33–51 (2014).
Sarauw, G. F. L. En Stenalders Boplads i Maglemose ved Mullerup Sammenholdt med Beslægtede Fund (H.H Thieles Bogtrykkeri, København, 1903).
Broholm, H. C. Nye fund fra den Ældste Stenalder, Holmegaard- og Sværdborgfundene. Aarbøger for Nordisk Oldkyndighed og Historie 1–144 (1924).
Mathiassen, T., Troels-Smith, J. & Degerbøl, M. Stenalderbopladser i Aamosen. (1943).
Clark, J. G. D. The Mesolithic Settlement of Northern Europe: A Study of the Food-Gathering Peoples of Northern Europe During the Early Post-Glacial Period (Greenwood Press, New York, 1936).
Verhart, L. B. M. Stone Age Bone and Antler As Indicators for ‘Social Territories’ in the European Mesolithic. In Contributions to the Mesolithic in Europe (eds Vermeersch, P. M. & Van Peer, P.) 139–151 (Leuven University Press, Leuven, 1990).
Larsson, L., Sjöström, A. & Nilsson, B. Lost at the bottom of the lake. Early and Middle Mesolithic leister points found in the bog Rönneholms Mosse, southern Sweden. In Working at the Sharp End: From Bone and Antler to Early Mesolithic Life in Northern Europe (eds Groß, D. et al.) 1–8 (Wacholtz, Kiel, 2019).
Andersen, K. Stenalder bebyggelsen i den Vestsjællandske Åmose (Fredningsstyrelsen, Copenhagen, 1983).
David, E. L’industrie en matières dures animale du Mésolithique ancien et moyen d’ Europe du nord, contribution de l’ analyse technologique à la définition du Maglemosien. (Université Paris X-Nanterre, 1999).
Leduc, C. Ungulates exploitation for subsistence and raw material, during the Maglemose culture in Denmark: the example of Mullerup site (Sarauw’s Island) in Sjælland. Danish J. Archaeol. 1, 62–81 (2012).
David, É The osseous technology of Hohen Viecheln: a Maglemosian idiosyncrasy? In From Bone and Antler to Early Mesolithic Life in Northern Europe (eds Groß, D. et al.) 1–36 (Wachholtz Verlag, Neumünster, 2019).
Gummesson, S. & Molin, F. Points of bone and antler from the Late Mesolithic settlement in Motala, eastern central Sweden. In Working at the Sharp End: From Bone and Antler to Early Mesolithic Life in Northern Europe (eds Groß, D. et al.) 1–25 (Wacholtz, Kiel, 2019).
Fischer, A. At the border of human habitat. The late Palaeolithic and early Mesolithic in Scandinavia. In The Earliest Settlement of Scandinavia and Its Relationship with Neighbouring Areas (ed. Larsson, L.) 157–176 (Almquist & Wiksell, Stockholm, 1996).
Fischer, A. Tissø og Amoserne som trafikforbindelse og kultsted i stenalderen. Historisk Samfund for Holbæk Amt 27–44 (2003).
Ramsey, C. B. Methods for summarizing radiocarbon datasets. Radiocarbon 59, 1809–1833 (2017).
Welker, F. et al. Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte du Renne. Proc. Natl. Acad. Sci. USA 113, 11162–11167 (2016).
Buckley, M. & Collins, M. J. Collagen survival and its use for species identification in Holocene-lower Pleistocene bone fragments from British archaeological and paleontological sites. Antiqua 1, 1–7 (2011).
Rodriguez, J., Gupta, N., Smith, R. D. & Pevzner, P. A. Does trypsin cut before proline?. J. Proteome Res. 7, 300–305 (2008).
Ekström, J. The Late Quaternary history of the urus (Bos primigenius Bojanus 1827) in Sweden. vol. 29 (Lund Univ., Dep. of Quaternary Geology, 1993).
Aaris-Sørensen, K., Mühldorff, R. & Petersen, E. B. The Scandinavian reindeer (Rangifer tarandus L.) after the last glacial maximum: time, seasonality and human exploitation. J. Archaeol. Sci.34, 914–923 (2007/6).
Aaris-Sørensen, K. Diversity and dynamics of the mammalian fauna in Denmark throughout the last glacial-interglacial cycle, 115–0 kyr bp. Fossils Strata 57, 1–59 (2010).
Aaris-Sørensen, K. Diversity and Dynamics of the Mammalian Fauna in Denmark Throughout the Last Glacial-Interglacial Cycle, 115–0 kyr BP (Wiley, New York, 2010).
Aaris-Sørensen, K. Depauperation of the Mammalian Fauna of the Island of Zealand during the Atlantic Period. Vidensk. Meddr Dansk Naturh. Foren. 142, 131–138 (1980).
Noe-Nygaard, N., Price, T. D. & Hede, S. Diet of aurochs and early cattle in southern Scandinavia: evidence from N and C stable isotopes. J. Archaeol. Sci. 32, 855–871 (2005).
McGrath, K. et al. Identifying archaeological bone via non-destructive ZooMS and the materiality of symbolic expression: examples from iroquoian bone points. Sci. Rep. 9, 11027 (2019).
Sjöström, A. Mesolitiska lämningar i Rönneholms mosse. Arkeologisk förundersökning 2010: Hassle 32:18, Stehag socken, Eslövs kommun 1–79 (Skåne. Lund University, Lund, 2011).
Sjöström, A. Mesolitiska lämningar i Rönneholms mosse. Arkeologisk förundersökning. Hassle 32:18, Stehag socken, Eslövs kommun Skåne 1–84 (. Lund University, Lund, 2014).
Fischer, A. Dating the early trapeze horizon. Radiocarbon dates from submerged settlements in Musholm Bay and Kalø Vig, Denmark. Mesolithc Misc. 15, 1–7 (1994).
Sørensen, S. A. Kongemosekulturen i Sydskandinavien (Egnsmuseet Færgegården, Jægerspris, 1996).
Sjöström, A. Ringsjöholm. A boreal-early atlantic settlement in Central Scania, Sweden. Lund Archaeol. Rev. 3, 5–20 (1997).
Fischer, A. People and the sea—settlement and fishing along the mesolithic coasts. In The Danish Storebælt Since the Ice Age—Man, Sea and Forest (eds Pedersen, L. et al.) 63–77 (A/S Storebælt Fixed Link, Copenhagen, 1997).
Tauber, H. Copenhagen radiocarbon dates VII. Radiocarbon 8, 213–234 (1966).
Tauber, H. Copenhagen radiocarbon dates X. Radiocarbon 15, 86–112 (1973).
Fischer, A. Food for Feasting? An evaluation of explanations of the neolithisation of Denmark and southern Sweden. In The Neolithisation of Denmark—150 Years of Debate (eds Fischer, A. & Krisiansen, K.) 343–393 (J. R Collis, Sheffield, 2002).
Andersen, S. H. & Petersen, P. V. Maglemosekulturens stortandede harpuner. Aarbøger Nordisk Oldkynd. Hist. 2004, 7–41 (2009).
Larsson, L. The colonization of South Sweden during the deglaciation. In The Earliest Settlement of Scandinavia and Its Relationship with Neighbouring Areas 24 (ed. Larsson, L.) 141–155 (Acta Archaeologica Ludensia, Stockholm, 1996).
Sørensen, L. & Casati, C. Hunter-gatherers living in a flooded world: the change of climate, landscapes and settlement patterns during the Late Palaeolithic and Mesolithic on Bornholm, Denmark. In Climate and Ancient Societies (eds Kerner, S. et al.) 41–69 (Museum Tusculanum, Copenhagen, 2015).
Sørensen, M. Early mesolithic regional mobility and social organization: evidence from lithic blade technology and microlithic production in southern Scandinavia. In Technology of Early Settlement in Northern Europe—Transmission of Knowledge and Culture (eds Knutsson, K. et al.) 173–201 (Equinox Publishing, London, 2018).
Bond, G. et al. A pervasive millennial-scale cycle in North Atlantic Holocene and Glacial Climates. Science 278, 1257–1266 (1997).
Björck, S. et al. High-resolution analyses of an early Holocene climate event may imply decreased solar forcing as an important climate trigger. Geology 29, 1107–1110 (2001).
Dahl, S. O., Nesje, A., Lie, Ø, Fjordheim, K. & Matthews, J. A. Timing, equilibrium-line altitudes and climatic implications of two early-Holocene glacier readvances during the Erdalen Event at Jostedalsbreen, western Norway. Holocene 12, 17–25 (2002).
Nesje, A., Dahl, S. O. & Bakke, J. Were abrupt Lateglacial and early-Holocene climatic changes in northwest Europe linked to freshwater outbursts to the North Atlantic and Arctic Oceans?. Holocene 14, 299–310 (2004).
Bakke, J., Dahl, S. O. & Nesje, A. Lateglacial and early Holocene palaeoclimatic reconstruction based on glacier fluctuations and equilibrium-line altitudes at northern Folgefonna, Hardanger, Western Norway. J. Quat. Sci. 2, 179–198 (2005).
Nesje, A. Latest Pleistocene and Holocene alpine glacier fluctuations in Scandinavia. Quat. Sci. Rev. 28, 2119–2136 (2009).
Berner, K. S., Koç, N. & Godtliebsen, F. High frequency climate variability of the Norwegian Atlantic Current during the early Holocene period and a possible connection to the Gleissberg cycle. Holocene 20, 245–255 (2010).
Balascio, N. L. & Bradley, R. S. Evaluating Holocene climate change in northern Norway using sediment records from two contrasting lake systems. J. Paleolimnol. 48, 259–273 (2012).
Jørgensen, S. Early Postglacial in Aamosen: Geological and Pollen-analytical Investigations of Maglemosian Settlements in the West-Zealand Bog Aamosen (Reitzel, Aigle, 1963).
Noe-Nygaard, N. Sedimentary, geochemical and ecological evolution of a Lateglacial-Postglacial lacustrine basin: lakelevel and climatic influence on flora, fauna and human population (Aamosen, Denmark). Foss. Strata 37, 1–436 (1995).
Noe-Nygaard, N., Abildtrup, C. H., Albrechtsen, T., Gotfredsen, A. B. & Richter, J. Palæobiologiske, sedimentologiske og geokemiske undersøgelser af Sen Weichel og Holocæne aflejringer i Store Åmose Danmark. Geol. tidsskr. 2, 1–65 (1998).
Gedda, B. Environmental and climatic aspects of the early to mid Holocene calcareous tufa and land mollusc fauna in southern Sweden (Lund University, Lund, 2001).
Digerfeldt, G., Björck, S., Hammarlund, D. & Persson, T. Reconstruction of Holocene lake-level changes in Lake Igelsjön, southern Sweden. GFF 135, 162–170 (2013).
Gaillard, M.-J. Postglacial paleoclimatic changes in Scandinavia and Central Europe. A tentative correlation based on studies of lake-level fluctuations. Ecol. Mediterr. 11, 159–175 (1985).
Nilsson, T. Die pollenanalytische Zonengliederung der spät- und postglazialen Bildungen Schonens. Geol. Föreningen Stockh. Förhandlingar 57, 385–562 (1935).
Digerfeldt, G. Reconstruction and regional correlation of Holocene lake-level fluctuations in Lake Bysjon South Sweden. Boreas 17, 165–182 (1988).
Dreibrodt, S. et al. Are mid-latitude slopes sensitive to climatic oscillations? Implications from an Early Holocene sequence of slope deposits and buried soils from eastern Germany. Geomorphology 122, 351–369 (2010).
Olsson, F., Gaillard, M. J., Lemdahl, G. & Greisman, A. A continuous record of fire covering the last 10,500 calendar years from southern Sweden—the role of climate and human activities. Palaeogeogr. Palaeoclimatol. Palaeoecol. 291, 128–141 (2010).
Manninen, M. A., Tallavaara, M. & Seppä, H. Human responses to early Holocene climate variability in eastern Fennoscandia. Quat. Int. 465, 287–297 (2018).
Grünberg, J. The Mesolithic burials of the Middle Elbe-Saale region. In: Mesolithic burials—Rites, symbols and socialorganisation of early postglacial communities (eds. Judith M. Grünberg, B. G., Larsson, L., Orscheidt, J. & Meller, H.) vol. 13,1 257–290 (Halle (Saale) Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt, Landesmuseum für Vorgeschichte 2016, 2016).
Crombé, P. Mesolithic projectile variability along the southern North Sea basin (NW Europe): hunter-gatherer responses to repeated climate change at the beginning of the Holocene. PLoS ONE 14, e0219094 (2019).
Solheim, S., Damlien, H. & Fossum, G. Technological transitions and human-environment interactions in Mesolithic southeastern Norway, 11 500–6000 cal. BP. Quat. Sci. Rev. 246, 106501 (2020).
Hammarlund, D., Björck, S., Buchardt, B., Israelson, C. & Thomsen, C. T. Rapid hydrological changes during the Holocene revealed by stable isotope records of lacustrine carbonates from Lake Igelsjön, southern Sweden. Quat. Sci. Rev. 22, 353–370 (2003).
Cziesla, E. & Pettitt, P. B. AMS-14C-Datieirungen von spätpaläolithischen und mesolithischen Funden aus dem Bützsee (Brandenburg). Archäol. Korresp. 33, 21–38 (2003).
Nordqvist, B. The Mesolithic settlements of the west coast of Sweden-with special emphasis on chronology and topography of coastal settlements. In Man and the Sea in the Mesolithic: Coastal Settlements Above and Below Present Sea Level; 1993; Kalundborg; Denmark (ed. Fischer, A.) 185–196 (Oxbow Books, Oxford, 1995).
Nordqvist, B. Coastal Adaptations in the Mesolitic [Mesolithic]: A Study of Coastal Sites with Organic Remains from the Boreal and Atlantic Periods in Western Sweden (Department of Archaeology Göteborg University, Gothenburg, 2000).
Johansson, G. En 10 000 år gammal boplats med organiskt material i Mölndal. Ytterligare en överlagrad Sandarnaboplats vid Balltorp. Västra Götalands län, Västergötland, Mölndal stad, Balltorp Ytterligare en överlagrad Sandarnaboplats vid Balltorp Västra Götalands län, Västergötland, Mölndal stad, Balltorp 1:124, Mölndal 182 Dnr 3.1.1-04306-2008(2014).
Boethius, A. Fishing for Ways to Thrive: Integrating Zooarchaeology to Understand Subsistence Strategies and Their Implications Among EARLY and Middle Mesolithic Southern Scandinavian Foragers (Lunds University, Lund, 2018).
Astrup, P. M. Sea-Level Change in Mesolithic Southern Scandinavia. Long- and Short-Term Effects on Society and the Environment 106 (Jutland Archaeological Society Publications, Højbjerg, 2018).
Fischer, A. & Petersen, P. V. Denmark—a sea of archaeological plenty. In Oceans of Archaeology (eds Fischer, A. & Pedersen, L.) 68–83 (Jutland Archaeological Society, Højbjerg, 2018).
Fischer, A. et al. Coast–inland mobility and diet in the Danish Mesolithic and Neolithic: evidence from stable isotope values of humans and dogs. J. Archaeol. Sci. 34, 2125–2150 (2007).
Ahlström, T. & Sjögren, K.-G. Kvinnan från Österöd—ett tidigmesolitiskt skelett från Bohuslän. In Situ Archaeologica 7, 47–69 (2007).
Ahlström, T. Mesolithic human skeletal remains from Tågerup, Scania, Sweden. In: Mesolithic on the Move. Papers Presented at the Sixth International Conference on the Mesolithic in Europe, Stockholm 2000 (eds. Larsson, L., Kindgren, H., Knutsson, K., Loeffler, D. & Åkerlund, A.) 478–484 (Oxbow Books, Oxford, 2003).
Desrosiers, P. M. The Emergence of Pressure Blade Making: From Origin to Modern Experimentation (Springer, Berlin, 2012).
Sørensen, M. The arrival and development of pressure blade technology in Southern Scandinavia. In The Emergence of Pressure Blade Making: From Origin to Modern Experimentation (ed. Desrosiers, P. M.) 237–259 (Springer, Cham, 2012).
Sørensen, M. et al. The first eastern migrations of people and knowledge into Scandinavia: evidence from studies of Mesolithic Technology, 9th-8th Millennium BC. Nor. Archaeol. Rev. 46, 19–56 (2013).
Günther, T. et al. Population genomics of Mesolithic Scandinavia: investigating early postglacial migration routes and high-latitude adaptation. PLoS Biol. 16, 1–22 (2018).
Kashuba, N. et al. Ancient DNA from mastics solidifies connection between material culture and genetics of mesolithic hunter–gatherers in Scandinavia. Nat. Commun. Biol. 2, 1–10 (2019).
Damlien, H., Kjällquist, M. & Knutsson, K. The pioneer settlement of Scandinavia and its aftermath: new evidence from Western and Central Scandinavia. In The Technology of Early Settlement in Northern Europe—Transmission of Knowledge and Culture 2 (eds Knutsson, K. et al.) 99–137 (Equinox Publishing, Sheffield, 2018).
Brock, F., Higham, T., Ditchfield, P. & Ramsey, C. B. Current pretreatment methods for AMS radiocarbon dating at the Oxford radiocarbon accelerator unit (Orau). Radiocarbon 52, 103–112 (2010).
Dee, M. & Bronk Ramsey, C. Refinement of graphite target production at ORAU. Nucl. Instrum. Methods Phys. Res. B 172, 449–453 (2000).
Ramsey, C. B., Higham, T. & Leach, P. Towards high-precision AMS: progress and limitations. Radiocarbon 46, 17–24 (2004).
Ramsey, C. B. C. B. Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Buckley, M., Collins, M., Thomas-Oates, J. & Wilson, J. C. Species identification by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 23, 3843–3854 (2009).
van Doorn, N. L., Hollund, H. & Collins, M. J. A novel and non-destructive approach for ZooMS analysis: ammonium bicarbonate buffer extraction. Archaeol. Anthropol. Sci. 3, 281 (2011).
Kirby, D. P., Buckley, M., Promise, E., Trauger, S. A. & Holdcraft, T. R. Identification of collagen-based materials in cultural heritage. Analyst 138, 4849–4858 (2013).
Perez-Riverol, Y. et al. The PRIDE database and related tools and resources in 2019: improving support for quantification data. Nucl. Acids Res. 47, D442–D450 (2019).
Acknowledgements
First and foremost, we would like to thank Axel Degn Johansson (Lundby, Denmark), and Uffe Seneca (Kalundborg, Denmark) for giving us access to their collection of barbed bone points. Additionally, we thank Peter Vang Petersen (The National Museum of Denmark) and Jeppe Boel Jepsen (Vendsyssels Historiske Museum) for access to their collections of barbed bone points. The Centre for Excellence in Mass Spectrometry, University of York, is thanked for access to MALDI-TOF-MS and so is Prof. Jesper Velgaard Olsen at the Novo Nordisk Center for Protein Research for providing access and resources. Karl Håkonsson is thanked for performing isotopic analyses at the Tandem Laboratory, Uppsala University. Likewise, Lisbeth Pedersen (Kalundborg Museum) is acknowledged for access to faunal remains from Mullerup and a barbed point from Ulkestrup Lyng. TZTJ is supported by the European Union’s EU Framework Programme for Research and Innovation Horizon 2020 under Grant Agreement no. 676154 (ArchSci2020). This study was also supported by NERC (NF/2018/1/5 - A chronology for (Danish) Maglemosian bone points). MM, AJT, LTL, and MJC are funded by the Danish National Research Foundation award PROTEIOS (DNRF128). HS was supported by the Villum Foundation (Grant No. 22917). Work at the Novo Nordisk Foundation Center for Protein Research is funded in part by a donation from the Novo Nordisk Foundation (Grant No. NNF14CC0001). Kristian Murphy Gregersen (Natural History Museum of Denmark) is thanked for providing reference material of elk and for access to unpublished radiocarbon dates as well as assistance with sampling. Søren Anker Sørensen (Museum Lolland-Falster) is thanked for providing reference material from red deer. Jan Dixon is also thanked for allowing us to use an unpublished AMS date from Sejerø. We also thank the following for valuable discussion: Harry Robson and Terry O’Connor (both from the University of York).
Author information
Authors and Affiliations
Contributions
T.Z.T.J. and M.J.C. conceived the study. T.Z.T.J. collected and prepared samples. D.C. performed radiocarbon dating by A.M.S. T.Z.T.J. and J.O. interpreted the radiocarbon data. T.Z.T.J., K.K.R., M.M. and A.J.T. prepared samples for MALDI-TOF-MS and LC-MS/MS and interpreted data. K.G. interpreted the isotope data. A.F., M.F.M. and L.S. provided unpublished radiocarbon dates. T.Z.T.J. wrote the manuscript with input from M.J.C., A.S., A.F., E.R., L.T.L., O.B., K.G., M.S., L.S., K.H.I., M.F.M., and N.M.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jensen, T.Z.T., Sjöström, A., Fischer, A. et al. An integrated analysis of Maglemose bone points reframes the Early Mesolithic of Southern Scandinavia. Sci Rep 10, 17244 (2020). https://doi.org/10.1038/s41598-020-74258-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-74258-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
