
Abstract
Stress can predispose to depressive episodes, yet the molecular mechanisms regulating the transition from the initial stress response to a persistent pathological depressive state remain poorly understood. We profiled the development of an enduring depressive-like state by assessing affective behavior and hippocampal function during the 2 months following social-defeat stress. We measured remodeling of hippocampal extracellular matrix (ECM) during this period, as we recently identified ECM changes to mediate cognitive impairment during the sustained depressive-like state. Affective disturbance and cognitive impairments develop disparately after social stress, with gradual appearance of affective deficits. In contrast, spatial memory was impaired both early after stress and during the late-emerging chronic depressive-like state, while intact in-between. Similarly, we observed a biphasic regulation of the hippocampal ECM coinciding with hippocampus-dependent memory deficits. Together our data (1) reveal a dichotomy between affective and cognitive impairments similar to that observed in patients, (2) indicate different molecular processes taking place during early stress and the chronic depressive-like state, and (3) support a role of the ECM in mediating long-lasting effects on memory. From a translational point of view, it is important to prioritize on temporal phenotypic aspects in animal models to elucidate the underlying mechanisms of depression.
Similar content being viewed by others
Introduction
Major depressive disorder (MDD) is a highly prevalent and debilitating neuropsychiatric disorder with a complex set of symptoms and high rates of relapse1. MDD is characterized by persistent disturbances in the affective domain, including depressed mood and anhedonia1. In addition to these well-characterized affective symptoms, cognitive dysfunction is prominent among depressed patients, affecting multiple domains, such as executive function, attention, memory and learning2,3. Despite the high occurrence and disabling effects on patients’ everyday life, cognitive dysfunction in depression has received little attention and therapeutic strategies against depression are mainly aimed at alleviating its mood-related symptoms. This poses serious limitations as residual cognitive symptoms often persist after improvement of mood symptoms and prevent functional recovery of patients4,5. Moreover, these residual symptoms are associated with an increased risk of relapse and recurrence of depressive episodes6. Taken together, increasing data suggest that rather than being an epiphenomenon of affective symptoms, cognitive dysfunction represents a core trait of the disease that is necessary to tackle in order to reach full recovery and prevent relapse.
Although the etiology of MDD remains elusive, stressful life experiences have been strongly linked to the development of depression and other neuropsychiatric disorders7. The hippocampus is highly susceptible to the effects of stress and given its critical involvement in cognitive function (e.g., spatial and temporal order processing), stress-induced hippocampal abnormalities appear to have a strong contribution to the cognitive deficits associated with MDD8,9. For example, attenuated hippocampal activity during hippocampus-dependent tasks has been demonstrated in MDD patients10,11, underscoring the potentially causal relation between deficits in recollection and declarative memory and hippocampal atrophy. Similarly, in chronically stressed animals, hippocampal memory deficits are present alongside structural changes and altered activity of hippocampal neurons12. Despite the vast data describing these stress-induced depression-related changes, the neural substrates and mechanisms that underlie the transition from initial stress exposure to a pathological depressive state remain elusive. In particular, the molecular mechanisms that propel these enduring effects of stress are poorly characterized as preclinical studies have largely focused on the short-lasting effects of acute and/or chronic stress.
Recently, we employed the social defeat-induced persistent stress (SDPS) model in rats to investigate the molecular mechanisms that underlie hippocampus-dependent cognitive dysfunction during a sustained depressive-like state13. In this preclinical model, brief social defeat stress is combined with a prolonged social isolation period (a subthreshold stressor14) that together result in enduring affective deficits and cognitive dysfunction, thereby recapitulating the core depression symptoms. Importantly, these deficits are prominent months after the last social defeat stress exposure, reminiscent of the human depressive state that can emerge and persist long after a stressful period15,16. As a novel pathological mechanism, we identified changes in hippocampal extracellular matrix (ECM), which mediate these cognitive impairments during the sustained depressive-like state13.
The brain ECM is a heterogeneous molecular network that forms a scaffold around neurons and synapses, supporting structural stability and regulating plasticity17,18. The pivotal role for ECM in mediating experience-dependent plasticity in the adult brain has become increasingly apparent. In particular, perineuronal nets (PNNs), lattice-like matrix structures predominantly coating parvalbumin-expressing interneurons19, have been shown to gate the closure of critical periods20, to sustain fear memories21 and to regulate addiction-related memories22,23. Furthermore, increasing evidence supports the involvement of abnormal ECM in diseases with disturbed cognitive processing, as diverse as schizophrenia24 and Alzheimer’s disease25.
In the present study, we studied the development of SDPS-induced depressive-like state by assessing affective behavior, as well as cognitive function during the days and weeks following social defeat stress. In addition to this temporal profiling of the core depressive-like symptoms, we characterized ECM remodeling over this same timeframe in order to understand how aberrant ECM that underlies the cognitive deficit at the late stage develops following initial social defeat stress.
Materials and methods
The full method section can be found in the Supplementary Information online.
Animals and the social defeat-induced persistent stress (SDPS) paradigm
All experiments were approved by the central ethics committee of the Netherlands and the Animal Users Committee of the Vrije Universiteit Amsterdam, in accordance with the relevant guidelines and regulations. The SDPS paradigm was carried out as previously described13, where male Wistar rats (≥ 9 weeks; Envigo, Netherlands) were used in a resident-intruder paradigm with male Long-Evans rats (> 4 months; Charles River, UK) as residents. SDPS rats underwent daily social defeat sessions of 15-min, including 5 min of physical contact. The defeat was repeated for five consecutive days and each day a new resident was used. From the first defeat session onwards, SDPS rats were single-housed until the end of the experiment. Control rats remained pair-housed throughout the experiment. During defeat sessions, they were transferred to an empty social defeat box, in which they remained for the session of defeat.
Time points taken for behavioral, and molecular analysis were named ‘2 weeks post-defeat’ (behavior: day 17, 18; molecular: day 20 post-defeat), ‘4 weeks post-defeat’ (behavior: day 31, 32; molecular: day 33 post-defeat), and ‘8 weeks post-defeat’ (behavior: day 56, 58; molecular: day 60 post-defeat).
Behavioral testing
All behavioral testing was performed during the dark phase of a 12 h light–dark cycle (lights on at 7 PM), under a dim red light as described previously13. Affective function was tested with the Social Approach Avoidance (SAA) test, in which exploration and approach to a caged unfamiliar Long-Evans rat was measured during the first minute of the test, and was calculated as the time spent near the social target vs. time spent near the empty box; interaction ratio: social target zone/(social target zone + empty box zone). Cognitive function in terms of spatial memory was assessed with the Object Place Recognition (OPR test), in which exploration and approach of a relocated object was analyzed during the first minute of the test, and was calculated as time spent exploring the relocated object compared to the stable object; discrimination index: relocated object/(relocated object + stable object).
Immunohistochemistry
Following transcardial perfusion with ice-cold 4% PFA in PBS and overnight post-fixation, brains were transferred to 30% sucrose in PBS at 4 °C until sectioned using a cryostat and processed for immunohistochemistry as described previously13. Free-floating sections (35 μm) were incubated in blocking solution (2.5% BSA, 0.2% Triton, 5% goat serum in PBS) for 2 h at RT and incubated with primary antibodies (mouse anti-chondroitin sulfate proteoglycan 1:1000, cat-301 MAB5284; rabbit anti-parvalbumin 1:1000, Swant #235) overnight at 4 °C. After washing in PBS, sections were incubated with fluorescent-conjugated secondary antibodies (anti-mouse Alexa-488 1:400, Invitrogen A11001; anti-rabbit Alexa-568 1:400, Invitrogen A11011) for 2 h at RT. Thereafter, sections were washed in PBS, mounted in 0.2% gelatin and cover-slipped using polyvinyl alcohol mounting medium with DABCO (Merck, 10981).
Images were acquired on a fluorescent microscope (Leica DM5000), and analyzed by Fiji software26, using automated threshold and particle analysis to detect the number of PNN+ and PV+ cells and their double immunoreactivity. False-positive cells were excluded manually during the analysis. During image acquisition and cell quantification, the researcher was blind to the experimental groups. On average, PNNs and PV+ cells were counted in 2–3 sections per animal, and subsequently averaged per animal.
Tissue preparation, immunoblotting, MMP extraction & zymography
Dorsal hippocampus27 samples were homogenized to either isolate synaptosomes for immunoblotting using a sucrose gradient and processed for immunoblotting as described previously13 or to isolate ECM-bound MMPs for zymography as described28.
Immunoblotting
Normalized immunoblotting data were subsequently log2-transformed and presented vs. those of control samples. In addition, data were compared to the log2-transformed quantification of perisynaptic ECM protein levels at 8 weeks post-defeat, as shown previously13.
MMP extraction & zymography
The isolated pellet (Supplementary Materials and Methods online) was solubilized in non-reducing sample buffer (2% SDS) and heated (37 °C, 15 min) before loading on an SDS-PAGE gel containing gelatin (0.1% gelatin, 8% SDS); samples (10 µg of protein) and recombinant mouse MMP-9 as positive control (5 ng, ab39309, Abcam). The gels were washed with 2.5% Triton X-100 (2 × 20 min) and then incubated for 7 days (50 mM Tris, pH 7.5, 10 mM CaCl2, 1 μM ZnCl2, 1% Triton X-100, and 0.02% NaN3; 37 °C, 80 rpm). Incubation was followed by Coomassie staining and destaining (5% HAc) until clear bands were visible. Gels were scanned and analyzed using Gel Doc EZ imager (Biorad, Herculus, CA, USA), and normalized based on Coomassie input.
Statistics
All data were analyzed using IBM SPSS Statistics 24. For group comparisons, two-tailed Student’s t-tests (with or without correction for unequal variation) were applied for normally distributed data, unless stated otherwise, and Mann–Whitney U-test was used when normality was violated. Data were checked for normality using the Saphiro-Wilk test. All group data are depicted as mean ± SEM, with overlaid individual data. Statistical significance level was set for P-values < 0.05, trend for 0.05 < P-value < 0.10. Correlations were made for SAA and OPR behavior per batch of animals, using Spearman rank (n < 15), or Pearson’s product moment (n ≥ 15). Details of all statistical testing can be found in Supplementary Table S1, and individual data points are given in Supplementary Table S2.
Results
Affective dysfunction and cognitive impairment develop disparately following social defeat stress
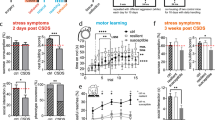
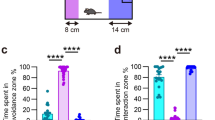
To characterize the development of depressive-like symptoms following a stressful experience, we performed a temporal analysis and assessed affective behavior and cognitive function at several time points (24 h/48 h, 2, 4 and 8 weeks) following the last social defeat encounter (Fig. 1a). The effect of SDPS on affective behavior was tested by the social approach avoidance (SAA) test, where interaction towards an unfamiliar Long-Evans rat is used to measure motivation for social interaction. Cognitive function was assessed by the object place recognition (OPR) test with a 15-min retention interval. This test measures short-term spatial memory and is heavily dependent on hippocampal function. In order to avoid carryover effects from repeated testing, independent groups of animals were used for each time point. In addition, these low-stress tests allowed analysis of ECM expression in absence of stress-by-test-induced effects. As others14 and we13 have previously shown, social isolation employed during and after social defeat is an intricate part of the SDPS paradigm and serves as a subthreshold stressor that contributes to the incubation of a chronic depressive-like state. In adult males, prolonged social isolation alone does not lead to alterations in behavior (Supplementary Fig. S1), hippocampal plasticity or corticosterone levels13.

Affective and cognitive impairments develop disparately after social defeat stress. (a) Adult rats were subjected to the SDPS (social defeat-induced persistent stress) paradigm; a 5-day social defeat paradigm followed by an 8-week period of individual housing. Control rats were not exposed to social defeat and remained pair-housed throughout the experiment. Behavior was assessed with social approach avoidance (SAA) and object place recognition (OPR) tests at four different time points: 24 h/48 h, 2 weeks, 4 weeks and 8 weeks after the last social defeat session. (b) Affective deficits emerged gradually following SDPS. At 24 h after the last social defeat session, no difference in approach towards a social target between SDPS and control groups was present. At week 2 post-defeat and thereafter decreased approach towards a social target was observed in the SDPS group (c) SDPS impaired spatial memory in a temporally dynamic manner. SDPS rats failed to discriminate between a stable and relocated object both shortly (48 h) after social defeat stress and at the chronic depressive state (week 8), whereas at week 2 and 4 post-defeat the SDPS rats showed intact spatial memory, similar to the control group. Behavior at each time point was assessed in an independent group of SDPS and control rats. Student’s t-test or Mann–Whitney U test were used to test statistical significance between SDPS and control group at each time point. #P < 0.05 compared to a fictive group with a mean of 0.5 and equal variation29; *P < 0.05; **P < 0.01. Data are expressed as mean ± SEM.
A day after (24 h) the last social defeat exposure, control and SDPS animals showed similar interaction time spent in the SAA test (control vs. SDPS, P = 0.935), indicating that recent stress does not affect approach behavior towards an unfamiliar social target in rats (Fig. 1b). At 2 weeks post-defeat, the SDPS group displayed decreased time spent with the social target (control vs. SDPS, P = 0.028), demonstrating the emergence of diminished motivation for social interaction and exploration. Similarly, at week 4 and 8 post-defeat, the SDPS rats showed decreased interaction ratios (control vs. SDPS, week 4 P = 0.023; week 8 P = 0.026). Taken together, these results demonstrate a gradually developing, yet persistent, effect of SDPS on affective behavior as observed previously13,30.
The next day (48 h after the last social defeat session), the SDPS group showed a deficit in the OPR test, assessing short-term spatial memory. A significant group effect was found (control vs. SDPS, P = 0.041), as the SDPS rats spent significantly less time exploring the relocated object when compared with the control group, thereby demonstrating an impairment in hippocampus-dependent spatial memory. Furthermore, whereas the control group showed a preference for the relocated object (control, P = 0.029 vs. a fictive control with a mean of 0.5 and equal variation29), the SDPS group failed to perform this task by showing an OPR discrimination index around chance levels (SDPS, P = 0.908 vs. a fictive control) (Fig. 1c). Surprisingly, at week 2 post-defeat, the SDPS group displayed normal short-term memory retention (SDPS, P = 0.003 vs. a fictive control) similar to the control group (control, P = 0.008 vs. a fictive control), while no between group differences were found (control vs. SDPS, P = 0.298). Also, at week 4 post-defeat both groups showed a preference for the relocated object (control, P = 0.018 vs. a fictive control; SDPS, P < 0.001 vs. a fictive control; and control vs. SDPS, P = 0.789). In accordance with our previous study13, 8 weeks after the last social defeat stress encounter, SDPS rats showed impaired memory function as indicated by decreased exploration of the relocated object compared with the control group (control vs. SDPS, P = 0.002) and a loss of preference for the displaced object (SDPS, P = 0.067 vs. a fictive control), whereas the control group retained information over the object’s location (control, P < 0.001 vs. a fictive control).
Notably, all animals were defeated by the same group of residents in a span of 4 weeks, ensuring no between-batch differences in the quantity or quality of received aggression that could explain these behavioral differences. In support, we did not observe differences in stress-induced arrest in weight gain among batches (Supplementary Fig. S2; P = 0.129).
Taken together, the current data show that hippocampal function is restored for a limited period of time after the short-term stress effects subside and before the long-term depression-related changes emanate. No significant correlation between SAA and OPR behavior was observed in any of the time points studied (Supplementary Fig. S3), indicating a degree of independence between these two behavioral domains, as shown previously30.
Cognitive impairment coincides with dysregulation of hippocampal extracellular matrix
Our previous study identified increased levels of hippocampal ECM during the depressive-like state evident months after the last social defeat exposure13. Specifically, SDPS increased the number of PNN-coated PV-expressing interneurons in the dorsal CA1 hippocampal subregion, but not in CA2/3 or DG13, together with an increase in the expression of perisynaptic ECM proteins in the dorsal hippocampus. Moreover, we showed that enzymatic disruption of CA1-ECM could restore the number of PNNs and, importantly, rescue the SDPS-induced hippocampal memory deficits, thereby demonstrating a causal relationship between hippocampal ECM dysregulation and cognitive dysfunction during the sustained depressive-like state13. However, at what moment after the defeat stress this increase in pericellular and perisynaptic ECM emerged was not clear.
To unravel the progression of SDPS-induced dysregulation of hippocampal ECM, in the present study we profiled temporal changes in the ECM at 72 h, 2, 4 and 8 weeks after social defeat stress, by quantifying the number of PNN+/PV+-cells in the CA1 pyramidal layer (Fig. 2a). Early after social defeat stress (72 h), a reduced number of PNN+/PV+-cells was found in the SDPS animals compared with the control group (control vs. SDPS, P = 0.042). This shows that social defeat stress induces an immediate ECM remodeling resulting in decreased number of PNN+/PV+-cells. At week 2 and 4 post-social defeat, no statistically significant difference in the number of PNN+/PV+-cells between the control and SDPS groups was present (week 2, control vs. SDPS, P = 0.144; week 4, control vs. SDPS, P = 0.126), revealing a recovery of the perisomatic ECM after the cessation of acute stress. Moreover, as shown previously13, an increased number of PNN+/PV+-cells was present 8 weeks following stress, characterizing the sustained depressive-like state (week 8, control vs. SDPS, P = 0.001) (Fig. 2b,c). Prolonged social isolation alone does not lead to alterations in the number of PNN+/PV+-cells (Supplementary Fig. S1). The density of PV+ interneurons was not affected in any of the time points studied (72 h, control vs. SDPS, P = 0.977; week 2 control vs. SDPS, P = 0.155; week 4 control vs. SDPS, P = 0.110; week 8 control vs. SDPS, P = 0.333; Fig. 2d), indicating a specific regulation of PNNs by SDPS both early after stress and at the phase when the depressive-like state appears (cf. Fig. 1).

SDPS induces a temporally dynamic regulation of CA1 perineuronal nets. (a,b) The number of chondroitin sulfate proteoglycan (CSPG+)-rich perineuronal nets (PNNs; green) and parvalbumin-expressing (PV+, red) interneurons were quantified with immunohistochemical (IHC) analysis at four time points: 72 h, 2 weeks, 4 weeks and 8 weeks after the last social defeat session (n = 3–5 animals per group; based on 2–3 sections per animal). Examples of staining for each time point is shown. Scale bar (75 µm) is indicated. (c) Decreased number of CSPG+PV+ -cells was present at 72 h after stress in the SDPS group. At week 2 and 4 no difference in the number CSPG+PV+-cells was observed. At the chronic depressive state (week 8) an increased number of CSPG+PV+-cells was found in the SDPS group, revealing a biphasic regulation of PNNs in response to SDPS. (d) SDPS did not alter the total number of PV-expressing cells in any of the time points studied. Student’s t-test was used to test statistical significance between SDPS and control group at each time point. *P < 0.05. Data are expressed as mean ± SEM.
Apart from the perisomatic PNNs surrounding PV-interneurons, perisynaptic extracellular space is enriched with ECM proteins that are vital for synaptic function17,31. Therefore, we also investigated the expression of individual ECM proteins in the synaptic fraction of the dorsal hippocampus at 72 h and 2 weeks following social defeat stress (Fig. 3a). We found the expression of the perisynaptic ECM to follow an identical pattern with that of PNNs. Namely, early after stress we found decreased expression of ECM proteins, followed by a transient recovery period during which perisynaptic ECM regulation was absent, before increasing at the long-term (Fig. 3b,d). Specifically, at the early time point (72 h), synaptic expression of several chondroitin sulfate proteoglycans (CSPG) from the lectican family, including brevican (Bcan), neurocan (Ncan) and phosphacan (Phcan) was reduced in the SDPS group (vs. control: Bcan, P = 0.039; Ncan, P = 0.029; Phcan, P = 0.022; Acan, P = 0.450). Similarly, the expression of the link-proteins Tenascin-R (TnR) and Hyaluronan and proteoglycan link protein 1 (Hapln1) was decreased shortly after social stress in the SDPS group (vs. control: TnR160, P = 0.007; TnR180, P = 0.047; Hapln1, P = 0.021) (Fig. 3c,d). At week 2 post-defeat, no difference in the expression of synaptic ECM proteins between the control and SDPS groups was present (vs. control: Bcan, P = 0.993; Ncan P = 0.760; Phcan, P = 0.671; Acan, P = 0.300; TnR160, P = 0.327; TnR180, P = 0.629; Hapln1, P = 0.491), illustrating a transient recovery of perisynaptic ECM protein levels, similar to what was observed for the number of PNNs.

SDPS induces a biphasic regulation of perisynaptic extracellular matrix. (a) Immunoblot (IB) analysis was performed at two time points: 72 h and 2 weeks (n = 6–8 animals per group) after the last social defeat session, and was compared to the log2-transformed quantification of perisynaptic ECM protein levels 8 weeks post-defeat13, presented vs. their respective control. (b,c) Decreased expression of chondroitin sulfate proteoglycans Brevican (Bcan), Neurocan (Ncan) and Phosphacan (Phcan), together with glycoproteins Tenascin (TnR) and Hyaluronan and proteoglycan link protein 1 (Hapln1) were present shortly after stress (72 h) in the SDPS group. At week 2 no difference in protein expression level was observed. (d) Example blots of each protein at the 72 h and week 2 post-defeat time point for Control (C) and SDPS (S). Full blots, as well as corresponding gels for total protein (sample normalization) are available in Supplementary Figs. S4 and S5 online. Student’s t-test was used to test statistical significance between SDPS and control group at each time point. *P < 0.05; **P < 0.01. Data are expressed as mean ± SEM.
Taken together, this temporal profile of hippocampal ECM organization shows that social defeat stress induces a persistent, yet dynamic ECM remodeling both at the perisynaptic, as well as at the pericellular level.
MMP-2 activity is decreased during the sustained depressive-like state
Matrix metalloproteinase-mediated breakdown of ECM assemblies is an essential mechanism by which ECM remodeling is maintained32,33. To assess whether altered MMP activity potentially drives the SDPS-associated ECM changes, we measured gelatinolytic activity of MMP-2 and MMP-9 using in-gel zymography at the two time points showing major ECM dysregulation (Fig. 4a), namely at 72 h and week 8 post-defeat. Early after social stress, we did not find a significant difference in the activity of either MMPs (vs. control: MMP-2, 118%, P = 0.619; MMP-9, 139%, P = 0.301; Fig. 4b,d). Notably, at 8 weeks after social defeat, we found decreased MMP-2 activity in the SDPS animals compared with controls (vs. control: 76%, P = 0.043), whereas no difference in MMP-9 activity was present (vs. control: 84%, P = 0.239; (Fig. 4c,d). Together, our data suggest that increased build-up of the perisynaptic and pericellular ECM, as observed during the sustained depressive-like state, could result from decreased MMP-2-mediated breakdown of the ECM in the hippocampus, whereas the early ECM effects after social defeat are possibly regulated by an MMP-2/MMP-9 independent mechanism.

MMP-2 activity is reduced during the sustained depressive-like state. (a) In-gel zymography assays (Zy) were performed at 72 h and 8 weeks after social defeat. (b) No difference in MMP-2 or MMP-9 activity was present shortly after social defeat; n = 6 animals per group. (c) At week 8 post-defeat, decreased MMP-2 activity was found in the SDPS group, without MMP-9 regulation; n = 6 animals (control); n = 4 (SDPS). *P < 0.05. Data are expressed as mean ± SEM. (d) Examples with equal loading showing the mature form of MMP-2 and MMP-9 (white bands), and the position of the 75 kDa marker band for each time point. Full gels are available in Supplementary Fig. 6 online.
Discussion
Despite a strong link between stressful life events and depression, understanding the mechanisms that underlie the transition from initial stress to a pathological depressive state remains elusive. To date, the majority of preclinical studies have focused on the short-lasting effects of stress, although it is well-recognized that stress can elicit long-lasting and late-appearing effects15,16,34. In the present study, we characterized the development of affective and cognitive impairments during the days and weeks following social defeat stress to unravel how these core depressive-like symptoms develop. Moreover, as our previous study demonstrated that changes in hippocampal ECM underlie hippocampus-dependent cognitive dysfunction during chronic depression13, we here characterized ECM remodeling over time to depict the progression of SDPS-induced hippocampal pathology.
The core depressive symptoms include persistent low mood and anhedonia that reflect abnormal emotion regulation and reward processing35. As an example, aberrant social interaction and avoidance of social contact are common in MDD patients36. Furthermore, the lack of social interaction can worsen the illness by augmenting stress responses, thereby perpetuating a vicious disease cycle37. In preclinical models, social interaction is often measured as the approach towards a social target, which is a naturally rewarding activity38. Here, we found that social deficits develop progressively during the weeks following social defeat stress. Social avoidance early after social defeat stress is generally employed to identify stress-susceptible individuals in mice39,40. In discordance with this, we here show that rats display intact social behavior 24 h after social defeat, which is followed by gradually deteriorating social interaction. Short-term isolation increases the likelihood of social exploration in male rats41 and hence this species-specific behavior might have masked the initial deficits in social interaction. On the other hand, the delayed emergence of affective deficits could suggest a gradual progression of the (mal)adaptation underlying emotional regulation in depression. As such, stress-induced early changes might not be accurate in predicting the emergence of enduring affective vulnerability. In support of this, we recently showed that acute effects of defeat stress on social behavior are poor predictors of depression vulnerability30. In fact, and in accordance with the present study, our previous data suggest that diminished social interaction displayed 5 weeks post-social defeat most faithfully predicts SDPS susceptibility and depression proneness. These findings indicate that defeat-induced affective deficits develop gradually following social stress and are likely facilitated by prolonged social deprivation42 in rats.
Although MDD is thought of as a primarily affective disorder, it is increasingly acknowledged that cognitive dysfunction is common among MDD patients and contributes profoundly to the disease burden43,44. While affective dysfunction and cognitive disturbance co-occur during depressive episodes, these symptoms can also be experienced independently of one another. Cognitive symptoms often persist even in remitted patients and can hinder functional recovery of patients45. Moreover, these residual symptoms can serve as a risk factor for relapse or potentially even increase susceptibility for a first depressive episode46. Additionally, recent studies suggest that cognitive performance of patients during premedication can predict antidepressant treatment response47. Thus, in addition to alleviating mood-affecting symptoms, it is critical to understand and relieve the cognitive deficits in depression, which can propagate disability in patients.
In the current study, spatial memory deficits were present early after the last social defeat encounter. In accordance, several studies have described stress-induced deterioration of hippocampal function shortly after stress9,48. This early phase, i.e. days after stress, is accompanied with substantial structural remodeling of neurons and aberrant hippocampal plasticity, effects that are largely mediated via glucocorticoid signaling49,50,51. Similar to several other stress paradigms, we have shown that 24 h after social defeat glucocorticoid levels are increased, but these levels become equal in the long-term13. It is then plausible that the early ECM changes we observed here act additionally, or synergistically, to glucocorticoid-mediated effects that are associated with hippocampal dysfunction and memory deficits52,53.
Strikingly, after these early stress effects subsided, we here showed a period of apparent intact hippocampal function that lasted several weeks before subsequent deterioration. The reversibility of the memory deficit that coincides with recent stress suggests that the late appearing effects evolve gradually and might be mediated by independent neurobiological substrates. In support, early stress effects, like increased glucocorticoid response, as well as those on heart rate, body temperature, activity54, and water- and food intake, dampen over time and are no longer present during the sustained depressive-like state13. Together, these findings highlight a dichotomy in the development of affective and cognitive depression-related symptoms, and, importantly, dissociate more acute stress responses from the late-appearing pervasive deficits that characterize the enduring depressive-like state (Fig. 5). This divergent developmental course of affective and cognitive deficits in response to stress likely reflects dichotomous pathophysiological mechanisms underlying these endophenotypes of depression. Accordingly, these two behavioral domains affected in depression may require distinct therapeutic approaches in order to reach full recovery.

Schematic of the main conclusion: ECM imbalance coincides with cognitive deficits. Overview of the social-stress-induced behavioral impairments and changes in hippocampus ECM during the development from initial stress towards establishing a depressive-like state. Affective (grey) and cognitive (blue) deficits are not temporally associated. Yet, dysregulation of perisomatic (PNN) and perisynaptic ECM (green) coincides with hippocampal dysfunction in terms of spatial memory in the rat SDPS model of depression.
We recently identified an increase in hippocampal ECM to underlie hippocampus-dependent memory deficits at the sustained depressive-like state13. In the present study, we profiled ECM changes in parallel to the behavioral characterization, in order to understand how aberrant ECM evolves in response to stress. Moreover, many stress-related studies showed down- instead of up-regulation of ECM55,56,57,58, sparking our interest to study the temporal aspect of ECM regulation. Here, we found social defeat stress to induce an immediate decrease in the expression of synaptic ECM proteins and in the density of perineuronal nets (PNN+/PV+-cells), similar as observed immediately after several other stressors applied57,59, without affecting the number of PV+-cells. Chondroitin sulfate proteoglycans (CSPGs) are among the most abundant ECM components in the brain, forming mesh-like structures around synapses, and they can condense into PNNs enwrapping predominantly PV+-interneurons60,61. Furthermore, the glycoprotein TnR links CSPGs together and Hapln1 attaches CSPGs to the hyaluronan backbone, thereby providing stability to the molecular matrix62. Damage to the ECM can destabilize connectivity between neurons and glia, disrupt neural activity and affect neural plasticity63; all mechanisms that are critical for learning and memory. For example, genetic deletion of TnR results in changes in perisomatic inhibition and excitability of pyramidal cells that are linked to altered long-term potentiation, the cellular correlate of learning and memory64. In addition, TnR ablation impairs spatial and motor learning65. Similarly, loss of brevican disrupts long-term potentiation66, and results in memory deficits67. Together, the downregulation in the expression of several synaptic ECM proteins shortly after social defeat may result in ECM composition-dependent aberrant synaptic transmission that could contribute to the observed memory deficits.
Perineuronal nets are found to mainly enwrap PV+-interneurons in the hippocampal CA1 subregion19. Although the precise mechanisms by which PNNs regulate the activity of PV+-interneurons remains elusive, PNNs have been shown to modulate the excitability and high-frequency firing of PV+-interneurons68,69. Recently, it was shown that mainly Brevican, expressed by PV+-interneurons, controls PV-excitability via the modulation of excitatory inputs these neurons receive, and controls molecular components of synaptic and intrinsic plasticity67. Conversely, hippocampus-dependent learning, and intrinsic plasticity affect activity of PV+-interneurons and Brevican levels67,70. These reciprocal relations illustrate that an imbalance in PNNs can alter excitatory/inhibitory signaling as well as network plasticity crucial to cognitive processing71,72,73. Thus, similar to the increased number of PNNs months after social defeat that coincides with reduced inhibitory network activity and cognitive deficits13, the decreased number of PNNs found shortly after defeat stress may alter inhibitory activity of PV+-interneurons and thereby impair hippocampus-dependent memory.
The observed co-occurrence of ECM regulation in the synaptic fraction and at the level of PNNs suggests that stress induces a widespread ECM remodeling in the hippocampus that is presumably regulated by the enzymes that breakdown ECM, such as matrix metalloproteinases (MMPs)74. Although MMP-9 is mostly studied due to the much wider availability of tools, several MMP-s and their regulators (e.g. tissue inhibitor of metalloproteinases, TIMPs) have been shown to be involved in memory deficits75,76,77. Here, we did not observe major changes in MMP-2 or MMP-9 activity during the early phase after social defeat stress, suggesting that a reduction in the pericellular and -synaptic ECM levels could possibly result from a reduced production or release of ECM proteins, rather than increased breakdown of the ECM by MMP-2 or MMP-9. Although measuring enzyme activity is more informative than measuring MMP-protein levels, the assay might not report endogenous enzyme activity because of possible auto-activation within the gel and loss of MMP-bound TIMPs during preparation78. Alternatively, other ECM-cleaving enzymes, such as ADAMTS4/579,80, could be regulated in response to stress and contribute to the ECM breakdown. It is plausible that proteolysis of ECM during and shortly after stress exposure may promote susceptibility to adverse environmental effects by making the extracellular milieu more permissive to synaptic reorganization. Eventually, this could result in the emergence of a pathological circuitry, which in turn could mediate the long-term changes manifested in late-appearing cognitive deficits.
Interestingly, in the present study we found ECM levels, both at the perisynaptic and perisomatic level, to recover after the initial stress-induced downregulation. Thus, ECM remodeling displayed a similar profile as hippocampus-dependent memory performance, namely, both showing a full recovery after the acute stress phase before deteriorating at a later phase. Previous studies have demonstrated reversibility of stress-induced hippocampal structural changes, such as an increase in the complexity of dendritic arborization of CA3 neurons after a recovery period following stress that is concurrent with improvement of cognitive deficits81. This reversal of the early stress effects has been suggested to represent a dynamic process rather than merely a return to the pre-stress state82. Indeed, we describe a transient rescue of stress-induced structural and functional changes at the level of hippocampal ECM and memory performance, respectively. We hypothesize that the initial stress-induced ECM down-regulation possibly triggers a cascade of compensatory mechanisms, involving enzymes responsible for ECM breakdown, together with astrocytic and neuronal ECM production. Presumably these compensatory processes aim to counteract ECM-driven changes in PV activity, which is critical to experience-dependent plasticity and learning70. Albeit that individual housing in control rats is not able to carry long-term changes in affective or cognitive behavior13 (cf. Supplementary Fig. S1), neuronal plasticity13, stress hormone levels13 or PNN number (cf. Supplementary Fig. S1), it acts as a subthreshold stressor that promotes deficits in affective behavior14. Because SDPS animals are deprived of environmental and social enrichment, it is possible that in these conditions compensatory systems fail, in turn contributing to the later emerging pathological state, observed months after social stress.
Taken together, our findings suggest that ECM remodeling serves as a substrate in mediating both the short- and long-term effects of social stress, and that normal ECM levels are essential for optimal memory processing.
Limitations
To shed light on the causal relationship of the early- and late-appearing ECM changes in the SDPS model, it would be critical to investigate whether ECM alterations and cognitive decline observed during the chronic phase of the depression-like state could be attenuated by preventing ECM remodeling during initial stress exposure or shortly thereafter. Although the interdependence of the early- and late-appearing ECM regulation remains under investigation, our findings suggest a critical involvement of ECM remodeling in the progression of stress-induced hippocampal dysfunction as both phases coincided with a corresponding memory deficit (Fig. 5). In support, those time points for which ECM regulation was absent coincided with intact hippocampal function. In this light, it is of interest that PNN numbers seemed to start their long-term upregulation most likely already 4–5 weeks after stress cessation, although not reaching statistical significance (cf. Fig. 2c). At this time point, we had a relatively low n-number and variation seemed larger than normal. This could indicate that PNN changes precede the behavioral deficits that become overt only 2 months after social stress, underscoring the causal nature of ECM changes as shown before13. Furthermore, as we found MMP-2 activity downregulated during the sustained depressive-like state, this suggests that attenuated MMP-2-mediated ECM breakdown facilitates the build-up of the ECM, thereby promoting the emergence of a state of limited plasticity. In relation to the cognitive phenotype we observe at the long-term, it is of interest that MMP-2 and MMP-9 activity are downregulated in serum of patients with recurrent depression, where higher MMP-levels were correlated with better cognitive performance83. On the other hand, we could not detect changes in activity of either enzyme shortly after stress cessation (cf. Fig. 4). This could indicate the involvement of other metalloproteinases, or an effect at the level of de novo ECM transcription and translation. Moreover, as we analyzed the whole dorsal hippocampus, we cannot exclude the presence of a subregion-specific increase in MMP-activity shortly after stress, as observed previously84.
Conclusion
Collectively, our results show that while SDPS elicits enduring social avoidance and cognitive impairment, these phenotypes progress differently, revealing a temporal dichotomy between affective behavior and cognitive function as often observed in MDD patients85. Moreover, our results reveal that SDPS induced a biphasic regulation of cognitive function, highlighting the dissociation of the sustained depressive-like state from the acute effects of stress. Furthermore, as we show that hippocampal ECM remodeling accompanies both early and long-lasting SDPS effects, targeting ECM levels may provide novel therapeutic opportunities against depression and other stress-related neuropsychiatric disorders. Together, from a translational point of view it is important to prioritize on the temporal and lasting aspects of affective and cognitive disturbances in animal models to elucidate the underlying causes of depression.
References
Malhi, G. S. & Mann, J. J. Depression. Lancet 392, 2 (2018).
McIntyre, R. S. & Lee, Y. Cognition in major depressive disorder: A ‘systemically important functional index’ (SIFI). Curr. Opin. Psychiatry 29, 48 (2016).
Papakostas, G. I. Cognitive symptoms in patients with major depressive disorder and their implications for clinical practice. J. Clin. Psychiatry 75, 8–14 (2014).
Rock, P. L., Roiser, J. P., Riedel, W. J. & Blackwell, A. D. Cognitive impairment in depression: A systematic review and meta-analysis. Psychol. Med. 44, 2029–2040 (2013).
Trivedi, M. H. & Greer, T. L. Cognitive dysfunction in unipolar depression: Implications for treatment. J. Affect. Disord. 152, 19–27 (2014).
Gonda, X. et al. The role of cognitive dysfunction in the symptoms and remission from depression. Ann. Gen. Psychiatry 14, 27 (2015).
Hammen, C. Stress and depression. Ann. Rev. Clin. Psychol. 1, 293–319 (2005).
McEwen, B. S. Stress and hippocampal plasticity. Annu. Rev. Neurosci. 22, 105–122 (1999).
Kim, J. J., Song, E. Y., Kim, J. J., Song, E. Y. & Kosten, T. A. Stress effects in the hippocampus: Synaptic plasticity and memory. Stress 9, 1–11 (2009).
Marazziti, D., Consoli, G., Picchetti, M., Carlini, M. & Faravelli, L. Cognitive impairment in major depression. Eur. J. Pharmacol. 626, 83–86 (2010).
Milne, A. M. B., MacQueen, G. M. & Hall, G. B. C. Abnormal hippocampal activation in patients with extensive history of major depression: An fMRI study. J. Psychiatry Neurosci. JPN 37, 28–36 (2012).
McEwen, B. S., Nasca, C. & Gray, J. D. Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 41, 3 (2016).
Riga, D. et al. Hippocampal extracellular matrix alterations contribute to cognitive impairment associated with a chronic depressive-like state in rats. Sci. Transl. Med. 9, 8753 (2017).
Ruis, M. A. W. et al. Housing familiar male wildtype rats together reduces the long-term adverse behavioural and physiological effects of social defeat. Psychoneuroendocrinology 24, 285–300 (1999).
Lupien, S. J., McEwen, B. S., Gunnar, M. R. & Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 10, 434–445 (2009).
Charles, S. T., Piazza, J. R., Mogle, J., Sliwinski, M. J. & Almeida, D. M. The wear and tear of daily stressors on mental health. Psychol. Sci. 24, 733–741 (2012).
Dityatev, A. & Schachner, M. Extracellular matrix molecules and synaptic plasticity. Nat. Rev. Neurosci. 4, 456–468 (2003).
Ferrer-Ferrer, M. & Dityatev, A. Shaping synapses by the neural extracellular matrix. Front. Neuroanat. 12, 40 (2018).
Härtig, W., Brauer, K. & Brückner, G. Wisteria floribunda agglutinin-labelled nets surround parvalbumin-containing neurons. NeuroReport 3, 869 (1992).
Pizzorusso, T. et al. Reactivation of ocular dominance plasticity in the adult visual cortex. Science 298, 1248–1251 (2002).
Gogolla, N., Caroni, P., Lüthi, A. & Herry, C. Perineuronal nets protect fear memories from erasure. Science 325, 1258–1261 (2009).
Slaker, M. et al. Removal of perineuronal nets in the medial prefrontal cortex impairs the acquisition and reconsolidation of a cocaine-induced conditioned place preference memory. J. Neurosci. 35, 4190–4202 (2015).
Lubbers, B. R. et al. The extracellular matrix protein Brevican limits time-dependent enhancement of cocaine conditioned place preference. Neuropsychopharmacology 41, 1907–1916 (2016).
Berretta, S. Extracellular matrix abnormalities in schizophrenia. Neuropharmacology 62, 1584–1597 (2012).
Morawski, M. et al. Involvement of perineuronal and perisynaptic extracellular matrix in Alzheimer’s disease neuropathology. Brain Pathol. 22, 547–561 (2012).
Schindelin, J. et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Spijker, S. Dissection of Rodent Brain Regions. In Neuroproteomics (ed. Li, K. W.) 13–26 (Humana Press, Totowa, 2011).
Szklarczyk, A., Lapinska, J., Rylski, M., McKay, R. D. G. & Kaczmarek, L. Matrix metalloproteinase-9 undergoes expression and activation during dendritic remodeling in adult hippocampus. J. Neurosci. 22, 920–930 (2002).
Akkerman, S., Prickaerts, J., Steinbusch, H. W. M. & Blokland, A. Object recognition testing: Statistical considerations. Behav. Brain Res. 232, 317–322 (2012).
Riga, D., Schmitz, L. J. M., Hoogendijk, W. J. G., Smit, A. B. & Spijker, S. Temporal profiling of depression vulnerability in a preclinical model of sustained depression. Sci. Rep. 7, 8570 (2017).
Song, I. & Dityatev, A. Crosstalk between glia, extracellular matrix and neurons. Brain Res. Bull. 136, 101–108 (2018).
Reinhard, S. M., Razak, K. & Ethell, I. M. A delicate balance: Role of MMP-9 in brain development and pathophysiology of neurodevelopmental disorders. Front. Cell. Neurosci. 9, 280 (2015).
Ethell, I. M. & Ethell, D. W. Matrix metalloproteinases in brain development and remodeling: Synaptic functions and targets. J. Neurosci. Res. 85, 2813–2823 (2007).
Syed, S. A. & Nemeroff, C. B. Early life stress, mood, and anxiety disorders. Chron. Stress 1, 2470547017694461 (2017).
Kober, H. & Ochsner, K. N. Regulation of emotion in major depressive disorder. Biol. Psychiat. 70, 910–911 (2011).
Kupferberg, A., Bicks, L. & Hasler, G. Social functioning in major depressive disorder. Neurosci. Biobehav. Rev. 69, 313–332 (2016).
Cacioppo, J. T., Hughes, M. E., Waite, L. J., Hawkley, L. C. & Thisted, R. A. Loneliness as a specific risk factor for depressive symptoms: Cross-sectional and longitudinal analyses. Psychol. Aging 21, 140 (2006).
Douglas, L. A., Varlinskaya, E. I. & Spear, L. P. Rewarding properties of social interactions in adolescent and adult male and female rats: Impact of social versus isolate housing of subjects and partners. Dev. Psychobiol. 45, 153–162 (2004).
Golden, S. A., Berton, O. & Russo, S. J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 6, 1183 (2011).
Berton, O. et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 311, 864–868 (2006).
Niesink, R. J. M. & Van Ree, J. M. Short-term isolation increases social interactions of male rats: A parametric analysis. Physiol. Behav. 29, 819–825 (1982).
de Jong, J. G., van der Vegt, B. J., Buwalda, B. & Koolhaas, J. M. Social environment determines the long-term effects of social defeat. Physiol. Behav. 84, 87–95 (2005).
Bortolato, B. et al. Cognitive remission: A novel objective for the treatment of major depression?. BMC Med. 14, 9 (2016).
Pan, Z. et al. Cognitive impairment in major depressive disorder. CNS Spect. https://doi.org/10.1017/s1092852918001207 (2018).
Bhalla, R. K. et al. Persistence of neuropsychologic deficits in the remitted state of late-life depression. Am. J. Geriatr. Psychiatry 14, 419–427 (2006).
Zuckerman, H. et al. Recognition and treatment of cognitive dysfunction in major depressive disorder. Front. Psychiatry 9, 655 (2018).
Park, C. et al. Predicting antidepressant response using early changes in cognition: A systematic review. Behav. Brain Res. 353, 154–160 (2018).
Sandi, C. & Pinelo-Nava, M. T. Stress and memory: Behavioral effects and neurobiological mechanisms. Neural Plast. 2007, 1–20 (2007).
Schwabe, L., Joëls, M., Roozendaal, B., Wolf, O. T. & Oitzl, M. S. Stress effects on memory: An update and integration. Neurosci. Biobehav. Rev. 36, 1740–1749 (2012).
de Quervain, D., Schwabe, L. & Roozendaal, B. Stress, glucocorticoids and memory: Implications for treating fear-related disorders. Nat. Rev. Neurosci. 18, 7–19 (2017).
Sapolsky, R. M. Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch. Gen. Psychiatry 57, 925–935 (2000).
Kim, E. J., Pellman, B. & Kim, J. J. Stress effects on the hippocampus: A critical review. Learn. Mem. 22, 411–416 (2015).
Roozendaal, B., Griffith, Q. K., Buranday, J., de Quervain, D.J.-F. & McGaugh, J. L. The hippocampus mediates glucocorticoid-induced impairment of spatial memory retrieval: Dependence on the basolateral amygdala. Proc. Natl. Acad. Sci. 100, 1328–1333 (2003).
Meerlo, P., Sgoifo, A., De Boer, S. F. & Koolhaas, J. M. Long-lasting consequences of a social conflict in rats: Behavior during the interaction predicts subsequent changes in daily rhythms of heart rate, temperature, and activity. Behav. Neurosci. 113, 1283–1290 (1999).
Folha, O. A. et al. Effect of chronic stress during adolescence in prefrontal cortex structure and function. Behav. Brain Res. 326, 44–51 (2017).
Ueno, H. et al. Region-specific impairments in parvalbumin interneurons in social isolation-reared mice. Neuroscience 359, 196–208 (2017).
Pesarico, A. P. et al. Chronic stress modulates interneuronal plasticity: Effects on PSA-NCAM and perineuronal nets in cortical and extracortical regions. Front. Cell. Neurosci. 13, 197 (2019).
Gomes, F. V., Zhu, X. & Grace, A. A. The pathophysiological impact of stress on the dopamine system is dependent on the state of the critical period of vulnerability. Mol. Psychiatry https://doi.org/10.1038/s41380-019-0514-1 (2019).
Yu, Z. et al. Decreased density of perineuronal net in prelimbic cortex is linked to depressive-like behavior in young-aged rats. Front. Mol. Neurosci. 13, 4 (2020).
Kwok, J. C. F., Warren, P. & Fawcett, J. W. Chondroitin sulfate: A key molecule in the brain matrix. Int. J. Biochem. Cell Biol. 44, 582–586 (2012).
Testa, D., Prochiantz, A. & Nardo, A. A. D. Perineuronal nets in brain physiology and disease. Semin. Cell Dev. Biol. https://doi.org/10.1016/j.semcdb.2018.09.011 (2018).
Yamaguchi, Y. Lecticans: organizers of the brain extracellular matrix. Cell. Mol. Life Sci. CMLS 57, 276–289 (2000).
Dityatev, A., Schachner, M. & Sonderegger, P. The dual role of the extracellular matrix in synaptic plasticity and homeostasis. Nat. Rev. Neurosci. 11, 735 (2010).
Saghatelyan, A. K. et al. The extracellular matrix molecule tenascin-R and its HNK-1 carbohydrate modulate perisomatic inhibition and long-term potentiation in the CA1 region of the hippocampus. Eur. J. Neurosci. 12, 3331–3342 (2000).
Montag-Sallaz, M. & Montag, D. Severe cognitive and motor coordination deficits in Tenascin-R-deficient mice. Genes Brain Behav. 2, 20–31 (2003).
Brakebusch, C. et al. Brevican-deficient mice display impaired hippocampal CA1 long-term potentiation but show no obvious deficits in learning and memory. Mol. Cell. Biol. 22, 7417–7427 (2002).
Favuzzi, E. et al. Activity-dependent gating of parvalbumin interneuron function by the Perineuronal net protein Brevican. Neuron 95, 2 (2017).
Dityatev, A. et al. Activity-dependent formation and functions of chondroitin sulfate-rich extracellular matrix of perineuronal nets. Dev. Neurobiol. 67, 570–588 (2007).
Lensjø, K. K., Lepperød, M. E., Dick, G., Hafting, T. & Fyhn, M. Removal of perineuronal nets unlocks juvenile plasticity through network mechanisms of decreased inhibition and increased gamma activity. J. Neurosci. 37, 1269–1283 (2016).
Donato, F., Rompani, S. B. & Caroni, P. Parvalbumin-expressing basket-cell network plasticity induced by experience regulates adult learning. Nature 504, 272 (2013).
Hu, H., Gan, J. & Jonas, P. Fast-spiking, parvalbumin+ GABAergic interneurons: From cellular design to microcircuit function. Science 345, 1255263 (2014).
Luscher, B. & Fuchs, T. GABAergic Control of Depression-Related Brain States. In Advances in Pharmacology 97–144 (Elsevier, Amsterdam, 2015).
Ferguson, B. R. & Gao, W.-J. PV interneurons: Critical regulators of E/I balance for prefrontal cortex-dependent behavior and psychiatric disorders. Front. Neural Circ. 12, 37 (2018).
Aguayo, F. I. et al. Matrix metalloproteinase 9 displays a particular time response to acute stress: Variation in its levels and activity distribution in rat hippocampus. ACS Chem. Neurosci. 9, 945–956 (2018).
Ganguly, K. et al. Matrix metalloproteinase (MMP) 9 transcription in mouse brain induced by fear learning. J. Biol. Chem. 288, 20978–20991 (2013).
Baba, Y. et al. Timp-3 deficiency impairs cognitive function in mice. Lab. Invest. 89, 1340–1347 (2009).
Nagy, V., Bozdagi, O. & Huntley, G. W. The extracellular protease matrix metalloproteinase-9 is activated by inhibitory avoidance learning and required for long-term memory. Learn. Memory 14, 655–664 (2007).
Lombard, C., Saulnier, J. & Wallach, J. Assays of matrix metalloproteinases (MMPs) activities: A review. Biochimie 87, 265–272 (2005).
Nakamura, H. et al. Brevican is degraded by matrix metalloproteinases and aggrecanase-1 (ADAMTS4) at different sites. J. Biol. Chem. 275, 38885–38890 (2000).
Howell, M. D. & Gottschall, P. E. Lectican proteoglycans, their cleaving metalloproteinases, and plasticity in the central nervous system extracellular microenvironment. Neuroscience 217, 6–18 (2012).
Hoffman, A. N. et al. Recovery after chronic stress within spatial reference and working memory domains: Correspondence with hippocampal morphology. Eur. J. Neurosci. 34, 1023–1030 (2011).
Ortiz, J. B. & Conrad, C. D. The impact from the aftermath of chronic stress on hippocampal structure and function: Is there a recovery?. Front. Neuroendocrinol. https://doi.org/10.1016/j.yfrne.2018.02.005 (2018).
Bobińska, K., Szemraj, J., Gałecki, P. & Talarowska, M. The role of MMP genes in recurrent depressive disorders and cognitive functions. Acta Neuropsychiatrica 28, 221–231 (2016).
van der Kooij, M. A. et al. Role for MMP-9 in stress-induced downregulation of nectin-3 in hippocampal CA1 and associated behavioural alterations. Nat. Commun. 5, 4995 (2014).
Darcet, F., Gardier, A., Gaillard, R., David, D. & Guilloux, J. Cognitive dysfunction in major depressive disorder a translational review in animal models of the disease. Pharmaceuticals 9, 9 (2016).
Acknowledgements
DR and ABS received support from NBSIK PharmaPhenomics grant LSH framework FES0908; MKK, DR, and SS were supported by an NWO VICI grant (ALW-Vici 016.150.673/865.14.002); DR was supported by the EU Research and Innovation Framework Programme Horizon 2020: Marie Skłodowska Curie Actions, Individual Fellowship (MSCA-IF-EF-ST; 793106).
Author information
Authors and Affiliations
Contributions
M.K.K., D.R., A.B.S., S.S. designed the experiments. D.R., M.K.K. executed behavioral experiments. D.R. analyzed behavioral data. Y.M. performed all decapitations for molecular analyses. M.K.K., D.R. executed and analyzed molecular data. M.K.K., S.S. made the figures. M.K.K., D.R., S.S. wrote the manuscript. A.B.S. commented during the study and on the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Koskinen, MK., van Mourik, Y., Smit, A.B. et al. From stress to depression: development of extracellular matrix-dependent cognitive impairment following social stress. Sci Rep 10, 17308 (2020). https://doi.org/10.1038/s41598-020-73173-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-73173-2
This article is cited by
-
Upregulated ECM genes and increased synaptic activity in Parkinson’s human DA neurons with PINK1/ PRKN mutations
npj Parkinson's Disease (2024)
-
Enriched environmental exposure reduces the onset of action of the serotonin norepinephrin reuptake inhibitor venlafaxine through its effect on parvalbumin interneurons plasticity in mice
Translational Psychiatry (2023)
-
Differential effects of stress-related and stress-unrelated humor in remitted depression
Scientific Reports (2022)
-
Benchmarking brain organoid recapitulation of fetal corticogenesis
Translational Psychiatry (2022)
-
Neurogenesis mediated plasticity is associated with reduced neuronal activity in CA1 during context fear memory retrieval
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
