
Abstract
Sebecosuchia was a group of highly specialized cursorial crocodyliforms that diversified during the Cretaceous and persist until the end of the Miocene. Their unique combination of cranial and post-cranial features indicates that they were active terrestrial predators that occupied the apex of the Late Cretaceous terrestrial ecosystems, even competing with theropod dinosaurs. Here, we report the discovery of the earliest sebecid worldwide, and the first from Eurasia, Ogresuchus furatus gen. et sp. nov., based on a semi-articulate specimen located in a titanosaurian sauropod nesting ground. The new taxon challenges current biogeographical models about the early dispersal and radiation of sebecid crocodylomorphs, and suggests an origin of the group much earlier than previously expected. Moreover, the new taxon suggests a potential convergent evolution between linages geographically isolated. Taphonomic evidences suggest that Ogresuchus died almost in the same place where fossilized, in a dinosaur nesting area. Biometric and morphologic observations lead to speculate that Ogresuchus could easily predate on sauropod hatchlings.
Similar content being viewed by others


Introduction
Late Cretaceous continental faunas from Europe are famed for showing a unique mosaic of taxa with both Gondwanan and Laurasian affinities1,2. During that time, the crocodomorph assemblage was characterized by the predominance of Laurasian eusuchian lineages, despite other minority groups like Gondwanan notosuchians were also present3. The enigmatic genus Doratodon is considered the only representative of the clade Notosuchia in the Late Cretaceous of Europe so far4,5, although the taxon is poorly known due to its scattered and fragmentary record3. On the other hand, the presence of isolated ziphodont teeth in several localities from Iberian Peninsula3,6 seems to indicate the presence of a more diverse notosuchian fauna than previously suspected.
Among notosuchians, the clade Sebecosuchia represents a group of highly specialized crocodyliforms characterized for their unique anatomical treats, showing triangular-shaped and laterally compressed skull, a significant reduction of the number of mandibular teeth, zhiphodont dentition, hypertrophied caniniform teeth, and cursorial limb morphology7,8,9,10 . The combination of these characters leads speculates that sebecosuchians were active terrestrial predators that could even compete with medium-size theropod dinosaurs as top predators during the Mesozoic8,9. Currently, two families can be recognised within Sebecosuchia: Baurusuchidae and Sebecidae11,12. While baurusuchids become especially diverse, abundant, and nearly exclusive from the Late Cretaceous of South America7,9,12, albeit putative remains of baurusuchids are mentioned in Asia13 and Africa14; sebecids diversified during the Palaeocene until the Miocene, also in South America11,12.
Here, we describe the first sebecid sebecosuchian of Eurasia on the based of a semi-articulate specimen, including cranial and post-cranial elements, which was discovered in a dinosaur nesting ground at the Southern Pyrenees (south-western Europe). This finding represents the oldest record of Sebecidae worldwide and sheds light on the early evolution of the group, and provides clues on the feeding behaviour of these terrestrial predators.
Results
Systematic palaeontology
Crocodylomorpha Walker, 1970 (sensu Clark15).
Crocodyliformes Hay, 1930 (sensu Clark 15).
Mesoeucrocodylia Whetstone and Whybrow, 1983.
Notosuchia Gasparini, 1971.
Sebecosuchia Simpson, 1937.
Sebecidae Simpson, 1937.
Ogresuchus furatus gen. et sp. nov.
Etymology
Genus name after Ogre- (French), in reference to the inferred feeding behaviour that included infant individuals, like the mythological creature from European folk tales; and –suchus, from the Greek Souchos meaning crocodile. Species name after furatus, from the Latin furari meaning to be stolen, in reference to the unfortunate event that took place during the fieldworks (see Supplementary Information S1).
Holotype
MCD-7149 (Museu de la Conca Dellà), a semi-articulate skeleton preserving the anterior part of the rostrum and several axial and appendicular elements (Fig. 1), and nine associate blocks containing large dinosaur eggshell fragments.

Skeletal remains of of Ogresuchus furatus (MCD-7149). (A,C) Photographic and (B,D) interpretative draws of the postcranial (A,B) and cranial (C,D) elements, and (E) silhouette showing preserved elements of Ogresuchus furatus. cv caudal vertebra, dv dorsal vertebra, fe femur, gr groves, lul left ulna, lt left tibia, m1-4 maxillary tooth, mc metacarpal, nvf neuro-vascular foramens, r rib, rt right tibia, sa sacral. Scale bar = 1 cm for (C,D) and 10 cm for (E).
Type locality and horizon
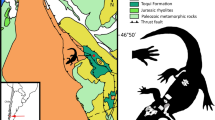
El Mirador site, (Coll de Nargó area, Lleida Province, Catalonia). High cemented grey marl level from the “lower grey unit” of the Tremp Formation; early Maastrichtian (near the C32n-C31r chrone boundary16,17).
Diagnosis
Small-sized sebecid diagnosed by the following autapomorphies: five maxillary tooth positions; teeth with smooth (unserrated) carinae; presence of apicobasal ridges on the enamel of the incisiviform and caniniform teeth; presence of apicobasal ridges on the enamel of posterior teeth; large and aligned neurovascular foramina on lateral surface of the maxilla; foramen in perinarial depression of the premaxilla; very large incisive foramen; absence of a large nutrient foramen on palatal surface of the premaxilla-maxilla contact; palatal surface of the maxilla without rugose surface; nasal-maxilary contacts remain parallel to each other (do not converge anteriorly or posteriorly); postzygapophyses located dorsally to the transverse processes in dorsal vertebrae.
Description
Cranial skeleton
The right premaxilla, the left maxilla, some teeth, the palatine and the palpebral are the best-preserved cranial remains, although a fragmentary right prefrontal could be also present. Complete descriptions for these bones were possible after a micro CT-scanning (Fig. 2).

3D reconstruction of the skull of Ogresuchus furatus (MCD-7149) in (a) lateral, (b) medial, (c) dorsal, (d) palatal, and (e) cranial view. (f) Volume rendering of the segmented neurovascular network of the trigeminal nerve overlaid on the articulated premaxilla and maxilla. app anterior palpebral, ch choana, dn dentary notch, en external naris, f neuro-vascular foramen, if inferior foramen, l-mx lacrimal-maxilla contact, m1-5 maxillary tooth, mes medial shelf, mx maxilla, paf palatal foramen, pd paramedian depressions, pfr prefrontal, plt palatine, pltf palatine foramen, pm1-4 premaxillary tooth, pmx premaxilla, pmx-mx premaxilla-maxilla contact, poas posantral strut, s apicobasal sulcus, snv-tgn V supranarial vessels and the trigeminal nerve V (ophthalmic branch), mv-tgn V maxillary vessels and the trigeminal nerve V (maxillary branch). Scale bar = 2 cm.
The right premaxilla is exposed on the rock in lateral view. It is a medio-laterally thin bone, and dorso-ventrally higher than rostro-caudally wide. The caudal margin is sinuous, making a dorso-caudal projection of the premaxilla for the contact with the nasals, and articulating with the lost right maxilla in a sigmoid suture (see specular image of Fig. 2). This margin is larger than the rostral premaxillary margin, making a sharp snout. The premaxilla makes the ventral, lateral and part of the dorsal margins of the external naris (Fig. 2a,b,e), which opens directly rostrally in the lower part of the snout. Except for the sloping wall of the naris and the lateral side of the tooth row, the lateral surface of the premaxilla is ornamented by a shallow pit-and-bulge pattern. Four premaxillary tooth positions are present (Fig. 2a,b,d). All the alveoli are of similar length and elliptical shape, although the third is slightly larger than the others. There is also a large foramen between the first alveolus and the naris. The third premaxillary tooth is preserved. It is conical with a very sharp crown and very labiolingually compressed. The crown is curved lingually and mesially. The mesial and distal margins of the crown are rounded and do not bear carinae. The enamel is ornamented with few apico-basal ridges that cross the crown continuously (Fig. 2a). In palatal view, the premaxilla makes a large incisive foramen separated from the tooth row. The premaxilla-maxilla suture is oriented anteromedially. At least two paramedial depressions are visible mesially and distally to the second tooth position of the premaxilla.
The left maxilla is also exposed in its lateral side, but on the opposite side of the rock respect to the premaxilla (Fig. 1C). It is latero-medially compressed, dorso-ventrally large and rostro-caudally short. The lateral surface is gently rugose, ornamented with the same pit-and-bulge pattern as the premaxilla. The maxilla is subpentagonal in outline. The anterior margin of the maxilla is oblique, because the premaxilla-maxilla suture is located into a notch for the reception of the dentary caniniform (Fig. 2a). The dorsal surface for the contact with the nasals is straight and reduced, and then, the dorsal border of the maxilla slopes ventro-caudally for contacting the lacrimal and, ventrally, the jugal. There is no evidence for an anteorbital fenestra. The maxilla projects medially from its ventral margin, making the secondary palate. In medial view, a septum appears on the lateral wall of the maxilla and turns caudally over the palatal portion of the bone, covering the internal breathing chamber. Caudally to the origin of this septum in the lateral wall, a big foramen opens to trigeminal passage. In palatal view, the maxillae branches meet completely anteriorly to the palatines (Fig. 2d). The maxilla makes the anterior border of the suborbital fenestra, precluding ectopterygoid-palatine contact in this margin.
Only five maxillary tooth positions are present (Fig. 2a,b,d). The maxilla preserves the second, third and fourth erupted teeth. The fist is partially preserved unerupted within the alveolus. The third maxillary tooth is the largest, whereas the fourth is the smallest of the three, although the fifth might be even smaller. The alveolar margin is ventrally arched, reaching the greatest depth at the third maxillary position. After the third maxillary tooth, the alveolar margin turns dorsally making a small notch, where the fourth alveolus is located. A row of eight foramina is present in the lateral side over the alveoli. The crowns are curved lingually and distally. The cross section is labio-lingually compressed. The mesial margins of the crowns are rounded, but the distal margins bear unserrated carinae. The enamel is ornamented with several conspicuous ridges that cross the crowns continuously from the base to the apex (Fig. 2a).
The palatine is an elongated bone rostro-caudally oriented, forming part of the narial passage. It is almost straight, though the caudal end is slightly wider than its rostral one. The anteriormost edge is not preserved, but the maxillary outline reveals a sharp anterior margin of the palatine, exceeding the anterior end of the suborbital fenestra and extending between the maxillae (Fig. 2d). The palatine forms the medial margin of the suborbital fenestra. The posterior ends of the palatines define the anterior and lateral margins of a large choanal opening. The anterior margin of the choana is situated between the suborbital fenestrae. Another D-shaped fenestra opens in the middle of the palatal shaft, anteriorly to the choana.
The anterior palpebral is large, and it is not sutured to the adjacent bones. The bone is subtriangular with a wider anterior end, and its major axis oriented antero-posteriorly (Fig. 2c). The anteromedial border is projected medially, forming a sharp crest for the articulation with the prefrontal. The bone is elongate posteriorly and forms the lateral margin of the supraorbital fenestra. The contact of palpebrals is not preserved, but the preserved portion suggests an oval supraorbital fenestra with an antero-posterior major axis.
Axial skeleton
Most of the dorsal series and few caudal vertebrae are identified. Preserved dorsal series includes seven complete and three fragmentary vertebrae, almost in articulation. These vertebrae are tentatively identified as 5th to 14th dorsal vertebrae. They are exposed in dorsal view, except 6th and 14th vertebrae that show their caudal view. Vertebral centra are amphicoelous. Prezygapophyses and postzygapophyses are well developed, with rounded margins, and laterally oriented. However, no variation in their orientation is observed along the dorsal series. The matrix partially hides the vertebrae, therefore some additional characters (i.e., orientation of articular facets; presence and morphology of a suprapostzygapophyseal lamina) cannot be assessed. The prezygapophyses seem to fuse with the transversal processes from the 7th dorsal vertebra on, as described in other related taxa as Notosuchus terrestris18, Baurusuchus albertoi19, Pissarrachampsa sera10 and Campinasuchus dinizi20. However this condition must be taken with caution, because it is only based on the 7th and 11th vertebrae. Transversal processes are hidden by the matrix in the rest of the series. Neural spines are broken in all the vertebrae except in the 14th dorsal. This spine is well developed and high, corresponding to half of the total height of the vertebra. However, based on the broken basis of neural spines along the series, the spine is medio-posteriorly located on the neural arch, as in B. albertoi19 and Campinasuchus20. A few distal caudal vertebral centra are also preserved, without association with neural spines and transverse processes.
In addition, some dorsal ribs are also identified. These elements are flattened. The proximal end shows the capitulum and the tuberculum for articulating with the associated vertebrae. Capitulum and tuberculum are separated by a well-marked U-shaped depression. The shaft is ventrally curved and shows a median longitudinal depression, unlike Campinasuchus20. At middle length the shaft makes torsion, being antero-posteriorly flattened at proximal half and medio-laterally fattened at distal half.
Forelimb
Only the right ulna, and the metacarpals I, II, III and IV are well identified. The proximal epiphysis of the right radius is also probably preserved (Fig. S9).
The ulna is an elongated and latero-medially flattened bone, as in other sebecids, baurusuchids, and notosuchians10,19,20. It is exposed in lateral side. In lateral view, the bone is arquated, displaying a concave anterior margin and a concave posterior one. The bone becomes shaper on its distal portion. The distal condyles are lost. The proximal end is cranio-caudally expanded. The proximal articular surface is concave, with the caudal olecranon process more developed than the cranio-lateral one. The lateral face bears a shallow longitudinal groove for the insertion of M. extensor carpi radialis brevis pars ulnaris, delimited caudally by a ridge for the insertion of M. flexor ulnaris10,21.
The proximal epiphysis of the radius is not well preserved. In proximal view it is a sub-squared bone with wide condyles, but it is strongly damaged hampering the assessment of detailed morphology.
Metacarpals were identified based on it general outline. The metecarpals I and II are almost complete, but the II, IV and the probable V are distally broken. Metacarpals decrease in width and robustness from the I to the V, being the first the largest. Each of them has an expanded proximal portion for articulating with the next metacarpal. The width of this expansion also decreases in size accordingly. In MI and III, the distal condyles bear a circular central depression for the attachment of M. interossei is observed21. These bones are similar to those referred to other baurusuchids10,19.
Hindlimb
A partial left femur, both tibiae and an indeterminate metatarsal were identified.
The femur is broken in two parts. The shaft seems almost cylindrical, but both proximal and distal ends are lost hindering any accurate morphological description.
Both tibiae are exposed in posterior view. They are long and medially curved bones, as in B. albertoi16, Sebecus22, Stratiotosuchus23 and Mariliasuchus8, differing from the straight condition in Crocodylia. Left tibia is preserved only in its distal portion, but the right tibia is almost complete. The tibial shaft is bowed posteriorly and medially, as in Sebecus22. This tibia is expanded at both ends, although the proximal articular surface is not well preserved. The distal end of the tibia is divided into lateral and medial portions. The medial portion is mesio-distally projected, forming an oblique distal margin. The lateral portion is well developed. This condition is present in other notosuchians as Stratiotosuchus, Notosuchus, Araripesuchus, Yacarerani, Pissarrachampsa and Sebecus8,10,22,24,25.
A left metatarsal is well preserved. It is a long and slender bone, compressed cranio-caudally. The shaft is almost straight with expanded proximal and distal ends. The proximal end shows well-marked lateral and medial condyles separated by a shallow concavity. The distal condyles are rounded, making a squared epiphysis. A lateral circular concavity is observed in both sides for the attachment of the M. interossei, as in the metacarpals. Based on the moderate expansion of the proximal end, this bone is tentatively considered as the metatarsal I.
Remarks
Based on the reconstructed 3D model, the general outline of the skull (Fig. 2), especially the lateromedially compressed and dorsoventrally high premaxillae and maxillae and the reduced dental formula (four or five maxillary teeth), resemble the typical doggy-shaped baurusuchid skulls7,26. The maxilla of Ogresuchus specially resembles that of Gondwanasuchus27, but although both taxa show apicobasal sulci on their teeth, Gondwanasuchus bears serrated dentition. The ornamentation pattern of Ogresuchus is also similar to Caipirasuchus teeth, although Caipirasuchus shows a highly specialized dentition composed by three serrated morphotypes in a continuous tooth row, not separated by a premaxillary-maxillary notch12. The anterior palpebral of Ogresuchus is unusually elongated. This bone differs from the morphology observed in most basal notosuchians, baurusuchids and sebecids; although comparisons are hindered because of the palpebral is not preserved in several species. On the other hand, the shape of the anterior palpebral is reminiscent to Gondwanasuchus27 and Araripesuchus tsangtsangana24. Finally, The absence of antorbital fenestra in Ogresuchus differs from many basal notosuchians and some baurusuchids12,26,27,28,29,30,31,32. This condition is similar to those observed in some basal notosuchians, a few baurusichids, and sebecids7,31,33.
Discussion
The phylogenetic position of Ogresuchus furatus was assessed based on the character-taxa matrices and methodology of Pol et al.12. The tree search strategy resulted in 1,150 most parsimonious trees of 1613 steps, before the second round of TBR; and exceeded the maximum number of trees retained in memory (99,999) after that. Therefore, the analysis was rerun excluding 3 unstable taxa pruned in the analysis reported by Pol et al.12—Pabwheshi, Pehuenchesuchus and Coringasuchus. The analysis resulted in 3,456 most parsimonious trees of 1604 steps, after the final round of TBR (CI = 0.3137, RI = 0.7338). As a result, Ogresuchus was recovered as a member of Sebecidae, within Sebecosuchia (Fig. 3). The main topology of the strict consensus tree (Fig. S3) agrees with other previous phylogenetic hypotheses12,32,34.

Time-calibrated evolutionary tree for Sebecosuchia. Reduced Consensus tree produced in TNT, with additional sebecosuchia taxa incorporated (see Supplementary text). The circles at each node represent the relative probabilities for the ancestral areas inferred using the Statistic Divergence-Vicariance Analysis method (S-DIVA; see Supplementary text). Global paleogeographic reconstructions from the Paleobiology Database (https://www.paleobiodb.org).
Ogresuchus possess the following unequivocal synapomorphies that support its inclusion within Sebecosuchia: a narrow orienorostral skull, one enlarged caniniform maxillary tooth, and a strong reduction of the dental formula (see Supplementary Information S1). The new taxon also exhibits some features that rule out its inclusion in the family Baurusuchidae, such as possessing an anteromedial margin of the palatine exceeding the anterior margin of the palatal fenestra; a broad palatine that is close to the half of the width of the maxillary palate; and the absence of a groove located on the premaxillary lateral surface. On the other hand, the inclination of the maxillary lateral surface, the large incisive foramen, and the posteriorly bowed tibial shaft group Ogresuchus within the clade formed by “Iberosuchus + Bergisuchus + Sebecidae”; whereas the small extension of the perinarial fossa is shared between Ogresuchus, Barinasuchus and Sebecus; and the size of the notch at the premaxilla-maxilla contact relates Ogresuchus and Sebecus. On the other hand, Ogresuchus convergently shares with Baurusuchus and Campinasuchus the conditions of five maxillary teeth and aligned large neurovascular foramina on lateral maxillary surface, respectively. Because of all the aforementioned synapomorphies plus the autopormophies exhibited by the new taxon, Ogresuchus furatus can be distinguished from any other sebecosuchian.
Biogeographic implications
Besides rare exceptions35, sebecids have been traditionally considered an exclusive Cenozoic group of cursorial mesoeucrocodylians from South America11. It is though that the first members of the family appeared during the middle Palaeocene, but the group diversified along over 45 Ma, from the Eocene to the Miocene (Fig. 3). However, the occurrence of Ogresuchus in the early Maastrichtian of Europe not only seems to pre-date the origin of the group, but also forces to review its biogeographic history.
Historically, the occurrence of similar fauna between Europe and southern landmasses during the Cretaceous has promoted the idea of a biogeographic connection between these regions1,2. This hypothesis, sometime referred as the Eurogondwana model5,36, suggests that some Gondwanan groups could migrate to the Late Cretaceous European archipelago by using intermittent land bridges1,5. This seems to be the case of the notosuchian Doratodon, which could target Europe as a part of a Turonian-Coniacian immigration wave that could follow a temporary route connecting Eastern Europe and Northern Africa1,5. In this regard, Ogresuchus could take similar routes than Doratodon to reach the European archipelago, but the advanced phylogenetic position of the new taxon within Sebecidae, closely related to Palaeogene Zulmasuchus and Sebecus spp. of South America, seems to depict a more complex evolutionary scenario.
From a broad palaeobiogeographic perspective, sebecosuchians originated in Gondwana at the beginning of Jurassic, and spread across southern continents (i.e. South America, Africa, India) until the Early Cretaceous34,37. The full opening of the Southern Atlantic and the complete separation of the Gondwana landmasses at the middle Albian (Fig. 3) prevented any terrestrial connections between southern landmasses from that time onwards. According to our Statistic Divergence-Vicariance Analysis (S-DIVA, see Supplementary Information S8 and Fig. S9), it is likely to suggest that sebecids were already present in both South American and African continent prior to the middle Albian, and that ancestral representatives of this group of terrestrial crocodyliforms would have evolve independently by allopatric speciation in separate continents (Fig. 3). Although sebecosuchian remains have been mentioned from the Cenomainan14 and the Eocene38 of Western and Northern Africa, the absence of conclusive evidences of sebecids from this continent humps building up a solid hypothesis about the evolution of a purported African linage during the Cretaceous, and the pathways of its radiation through Europe that eventually gave rise to Ogresuchus and Doratodon. In any case, what it seems likely is that Ogresuchus could belong to a distinct linage than that of Zulmasuchus and Sebecus from the Palaeogene of South America, and therefore a key question arise: why do they appear phylogenetically grouped?
Although being a preliminary speculation, two potential scenarios might explain such results. On one hand, the phylogenetic features that define the clade grouping Ogresushus + Zulmasuzhus + Sebecus spp. might, indeed, to be considered common synapomorphic characters shared by both South American and African linages, remaining invariable for several millions of years. Most of these characters are related to the contact area between maxilla and premaxilla (see Supplementary Information), and it might suggest that the basic configuration of the anterior part of the snout that defines the group was already present in the early members of the family.
On the other hand, the phylogenetic treats that group Ogresuchus with Palaeogene South American sebecids could result as a convergent evolution phenomenon. In this case, the alleged allopatric speciation of the South American and African linages could drive some taxa to acquire similar morphological features. In fact, convergent evolution among crocodyliforms is well known, and several evidences suggest that ecomorphological convergent adaptation (i.e. occupation of similar dietary niche) can contribute more signal than phylogenetic relatedness39. If so, this could imply that the real phylogenetic position of Ogresuchus could be obscured by its ecomorphological convergence with other taxa.
Estimation of body size and body length
Body size is one of the most important biological features because its intimate relationship to the ecology of any organism40,41,42. Based on recent allometric equations20, the body length of the Ogresuchus was estimated in 1.09 m and its body mass in only 9.04 kg of weight (see Supplementary information S1), making Ogresuchus one of the smallest and lightest sebecosuchids ever discovered. Other medium-sized predator sebecosuchians from the Late Cretaceous (i.e. Baurusuchus and Pissarrachampsa) and the Palaegene (i.e. Sebecus) doubled the size of Ogresuchus (2–3 m in length) but notably exceeded its weight (c. 70 kg)9,22.
Taphonomy and preservation
From a taphonomic point of view, the skeleton of Ogresuchus is incomplete and not fully articulated although the bones are preserved in good condition and show certain anatomic-like arrangement (Supplementary Information S1 and Fig. S5). The arrangement of bones resembles those of current crocodile carcasses undergoing sub-aerial and subaqueous decay without burial43. Several well-preserved eggshell fragments occur at the right side of the skeleton, and most of them are oriented perpendicular to the bedding plant, indicating low or no transport44. The combination of these taphonomic evidences lead to suggest an autochthonous deposition, with Ogresuchus most likely buried at the same place where it died.
Paleoecology and feeding behaviour
Notosuchians are known for exhibiting a large diversity of tooth morphology adapted to many types of feeding behaviours, and their cranio-mandibular anatomy indicates that they were capable to perform complex jaw movements45,46. Fossil evidences indicate that Late Cretaceous sebecosuchians occupied a dominant position in the terrestrial trophic web, competing with theropod dinosaurs as top predators9. Although it has been hypothesized that some sebecosuchians could feed on young dinosaurs8,9,10, no direct or indirect evidence are provided to support such ecological inference so far. Fossil record offers indeed rare examples about predation upon eggs, neonate or very young dinosaur individuals47,48,49,50. In this regard, the in situ occurrence of Ogresuchus in a dinosaur nesting-site offers a worthy opportunity to explore possible end-Cretaceous prey-predator relationships.
As a first approach, it can be stated that the small body size of Ogresuchus and its dental morphology suggest some basic biometric and biomechanical limitations for directly predating upon a 19–23 cm-in-diameter thick-shelled egg (see Supplementary Information). Although it has been stated that the jaw of sebecosuchians could produce orthal movements with a wide opening46, the small cranial size of Ogresuchus probably could not load an efficient bite force upon the thick megaloolithid eggshell. In addition, none of the 30 eggshells surrounding the skeleton nor any of the 1,000 fragments discovered in the El Mirador site16 shown evidences of the characteristics hole marks and cracking associated to predation51. These observations are consistent with previous studies pointing out that sebecosuchians had no specific adaptations for opening eggs47.
On the contrary, it is likely that Ogresuchus could produce some kind of predation pressure on a 3-kg-weigh, 40-cm-long neonate titanosaur52,53. One of the most highlighting features of Ogresuchus is its labio-lingual compressed dentition without serration and marked apicobasal ridges on the tooth crown, a dental condition shared with some theropod dinosaurs like Buitretraptor and Compsognathus54. Although non-serrated grooved teeth are commonly referred to purported piscivorous tetrapods55, the blade-like morphology of caniniform teeth of Ogresuchus could likely pierce the soft flesh of a baby titanosaur that, in addition, hatched with no defensive elements (i.e. osteoderms56). Thus, a feeding behaviour that incorporates the ingestion of hatchling dinosaurs should be in the scope of further palaeobiological analyses of Ogresuchus.
In conclusion, the discovery of Ogresuchus furatus in Late Cretaceous deposits from southern Europe “pushes back” the origination time of the once top terrestrial predators sebecid sebecosuchians. According to our interpretations, sebecids would have originated the previous to the full break of Gondwana (middle Albian). If so, a remainder group of sebecids could evolve in Africa for several millions of years until some taxa could reached the Late Cretaceous European archipelago. As occurs with many other taxonomic Cretaceous groups from Europe, a cornerstone in this hypothesis relies in future discoveries of terrestrial fossils from the Early Cretaceous of the African continent. Anyway, whatever the factor drifting the evolution of the group, phylogenetic and morphological evidences might suggest certain degree of ecomorphological convergence between early members of the family and more derived sebecid taxa. Furthermore, taphonomic and biometric evidences provide evidences that lead speculating that Ogresuchus furatus could stalk neonate titanosaure sauropods, incorporating them as a part of diet.
Materials and methods
Phylogenetic analysis
The data matrix and the phylogenetic methodology were preformed following Pol et al.12. The new taxon was coded for this study and included in the dataset, resulting in a final matrix of 412 characters and 110 OTUs. According to methods described by Pol et al.9, the character 5 was excluded from the analysis and the characters 1, 3, 6, 10, 23, 37, 43, 44, 45, 49, 65, 67, 69, 71, 73, 77, 79, 86, 90, 91, 96, 97, 105, 116, 126, 140, 142, 143, 149, 167, 182, 187, 193, 197, 226, 228, 279, 339, 356, 357, 364, 368 were set as additive because represent nested sets of homologies and/or entail present and absence information. The character matrix was analysed using a maximum parsimony approach (traditional search method) in TNT 1.557,58. An heuristic tree search of 10,000 replicates of Wagner trees with random addition sequences was performed followed by TBR branch-swapping, collapsing zero-length branches. According to Pol et al.12, all most parsimonious trees recovered in this search were subjected to a final round of TBR branch-swapping.
CT scan and virtual reconstruction
A virtual three-dimensional (3D) cranial and appendicular reconstructions of O. furatus were performed based on the holotype MCD-7149 (Figs. 1, 2, Video 1). The specimen was scanned by microfocus X-ray computed tomography (Multitom CORE X-ray CT; voxel size of 25 µm, and 120 kV) at the CORELAB Laboratory of the CRG Marine Geosciences of the Faculty of Earth Sciences at the University de Barcelona (Barcelona Spain). Three separated acquisitions were necessary to cover the whole specimen. Raw data from each scanning were imported (as stack of TIFF 16-bit files) to Avizo 7.1 and Blender 2.79b for segmentation, repositioning, mirroring and visualization. Each bone or bone fragment was segmented virtually removing the surrounding matrix using semiautomatic thresholding tools and obtaining individual 3D digital models. As a result, eleven 3D models were generated prior to repositioning and mirroring them to assemble partial skull and forelimb. The 3D models were repositioned and mirrored using Blender 2.79b based on bilateral symmetry.
Change history
07 January 2021
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Csiki-Sava, Z., Buffetaut, E., Ősi, A., Pereda-Suberbiola, X. & Brusatte, S. L. Island life in the Cretaceous-faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. ZooKeys 469, 1–161 (2015).
Blanco, A. Igdabatis marmii sp. Nov. (Myliobatiformes) from the lower Maastrichtian (Upper Cretaceous) of northeastern Spain: An Ibero-Armorican origin for a Gondwanan batoid. J. Syst. Palaeontol. 17, 865–879 (2019).
Puértolas-Pascual, E., Blanco, A., Brochu, C. A. & Canudo, J. I. Review of the Late Cretaceous-early Paleogene crocodylomorphs of Europe: Extinction patterns across the K-Pg boundary. Cretac. Res. 57, 565–590 (2016).
Company, J., Pereda-Suberbiola, X., Ruíz-Omeñaca, J. I. & Buscalioni, ÁD. A new species of Doratodon (Crocodyliformes: Ziphosuchia) from the Late Cretaceous of Spain. J. Verteb. Paleontol. 25, 343–353 (2005).
Rabi, M. & Sebök, N. A. Revised Eurogondwana model: Late Cretaceous notosuchian crocodyliforms and other vertebrate taxa suggest the retention of episodic faunal links between Europe and Gondwana during most of the Cretaceous. Gondw. Res. 28, 1197–1211 (2015).
Blanco, A., Puértolas-Pascual, E. & Marmi, J. Assessing the crocodylomorph diversity in the Maastrichtian (Late Cretaceous) of Ibero-Armorica, based on tooth qualitative traits. Abstract book of the XVI Annual Meeting of the European Association of Vertebrate Palaeontologists, 28 (2018).
Montefeltro, F. C., Larsson, H. C. E. & Langer, M. C. A new baurusuchid (Crocodyliformes, Mesoeucrocodylia) from the Late Cretaceous of Brazil and the phylogeny of Baurusuchidae. PLoS ONE 6, e21916 (2011).
Riff, D. & Kellner, A. W. A. Baurusuchid crocodyliforms as theropod mimics: Clues from the skull and appendicular morphology of Stratiotosuchus maxhechti (Upper Cretaceous of Brazil). Zool. J. Linn. Soc. 163, S37–S56 (2011).
Godoy, P. L., Montefeltro, F. C., Norell, M. A. & Langer, M. C. An additional baurusuchid from the Cretaceous of Brazil with evidence of interspecific predation among Crocodyliformes. PLoS ONE 9, e97138 (2014).
Godoy, P. L. et al. Postcranial anatomy of Pissarrachampsa sera (Crocodyliformes, Baurusuchidae) from the Late Cretaceous of Brazil: Insights on lifestyle and phylogenetic significance. PeerJ 4, e2075 (2016).
Kellner, A. W. A., Pinheiro, A. E. P. & Campos, D. A. A new sebecid from the Paleogene of Brazil and the crocodyliform radiation after the K-Pg boundary. PLoS ONE 9, e81386 (2013).
Pol, D. et al. A new notosuchian from the Late Cretaceous of Brazil and the phylogeny of advanced notosuchians. PLoS ONE 9, e93105 (2014).
Wilson, J. A., Malkane, M. S. & Gingerich, P. D. New crocodyliform (Reptilia, Mesoeucrocodylia) from the Upper Cretaceous Pab Formation of Vitakri, Balochistan (Pakistan). Cont. Mus. Paleontol. Univ. Michingan 30, 321–336 (2001).
Sereno, P. C., Wilson, J. A. & Conrad, J. L. New dinosaurs link southern landmasses in the mid-Cretaceous. Proc. R. Soc. Bio. Sci. 271, 1325–1330 (2004).
Clark, J.M. Phylogenetic Relationships of the Crocodylomorph Archosaurs. PhD thesis, (University of Chicago, 1986).
Sellés, A. G. et al. Dinosaur eggs in the Upper Cretaceous of the Coll de Nargó area, Lleida Province, south-central Pyrenees, Spain: Oodiversity, biostratigraphy and their implications. Cretac. Res. 40, 10–20 (2013).
Fondevilla, V. Registre geològic, paleoambients i successió dels darrers dinosaures del sud-oest europeu. PhD thesis, (Univeristat Autònoma de Barcelona, 2017).
Pol, D. Postcranial remains of Notosuchus terrestris (Archosauria: Crocodyliformes) from the Upper Cretaceous of Patagonia, Argentina. Ameghiniana 42, 1–17 (2005).
Nascimento, P. M. & Zaher, H. A new species of Baurusuchus (Crocodyliformes, Mesoeucrocodylia) from the Upper Cretaceous of Brazil, with the first complete postcranial skeleton described for the family Baurusuchidae. P. Avu. Zool. 50, 323–361 (2010).
Cotts, L., Piacentini Pinheiro, A. E., de Silva Marinho, T., de Souza, C. I. & Di Dario, F. Postcranial skeleton of Campinasuchus dinizi (Crocodyliformes, Baurusuchidae) from the Upper Cretaceous of Brazil, with comments on the ontogeny and ecomorphology of the species. Cretac. Res. 70, 163–188 (2017).
Meers, M. B. Crocodylian forelimb musculature and its relevance to Archosauria. Anatom. Rec. Part A 274, 891–916 (2003).
Pol, D., Leardi, J. M., Lecuona, A. & Krause, M. Postcranial anatomy of Sebecus icaeorhinus (Crocodyliformes, Sebecidae) from the Eocene of Patagonia. J. Vertebr. Paleontol. 32, 328–354 (2012).
Campos, D. A., Suarez, J. M., Riff, D. & Kellner, A. W. Short note on a new Baurusuchidae (Crocodyliformes, Metasuchia) from the Upper Cretaceous of Brazil. Bol. Museu Nacional Nova Série Geologia 57, 1–7 (2001).
Turner, A. H. Osteology and phylogeny of a new species of Araripesuchus (Crocodyliformes: Mesoeucrocodylia) from the Late Cretaceous of Madagascar. Hist. Biol. 18, 255–369 (2006).
Fiorelli, L. E. & Calvo, J. O. New remains of Notosuchus terrestris Woodward, 1896 (Crocodyliformes: Mesoeucrocodylia) from Late Cretaceous of Neuquen, Patagonia Argentina. Arquivos do Museu Nacional 66, 83–124 (2008).
Carvalho, I. S. et al. Campinasuchus dinizi gen. et sp. nov., a new Late Cretaceous baurusuchid (Crocodyliformes) from the Bauru Basin, Brazil. Zootaxa 2871, 19–42 (2011).
Da Silva Marinho, T., Vidoi Iori, F., Carvalho, I. S. & Mesquita de Vasconcellos, F. Gondwanasuchus scabrosus gen. et sp. nov., a new terrestrial predatory crocodyliform (Mesoeucrocodylia: Baurusuchidae) from the Late Cretaceous Bauru Basin of Brazil. Cretac. Res. 44, 104–111 (2013).
Gomani, E. M. A crocodyliform from the Early Cretaceous Dinosaur Beds, Northern Malawi. J. Vertebr. Paleontol. 17, 280–294 (1997).
Sereno, P. C., Sidor, C. A., Larsson, H. C. E. & Gado, B. A new notosuchian from the Early Cretaceous of Niger. J. Vertebr. Paleontol. 23, 477–482 (2003).
Carvalho, I. S., Mesquita de Vasconcellos, F. & Simionato Tavares, S. A. Montealtosuchus arrudacamposi, a new peirosaurid crocodile (Mesoeucrocodylia) from the Late Cretaceous Adamantina Formation of Brazil. Zootaxa 1607, 35–46 (2007).
Nobre, P. H. & Carvalho, I. S. Adamantinasuchus navae: A new Gondwanan Crocodylomorpha (Mesoeucrocodylia) from the Late Cretaceous of Brazil. Gondw. Res. 10, 370–378 (2006).
Fiorelli, L. E. et al. A new Late Cretaceous crocodyliform from the western margin of Gondwana (La Rioja Province, Argentina). Cretac. Res. 60, 194–209 (2016).
Zaher, H. et al. Redescription of the cranial morphology of Mariliasuchus amarali, and its phylogenetic affinities (Crocodyliformes, Notosuchia). Am. Museum Novitates 3512, 1–40 (2006).
Dal Sasso, C., Pasini, G., Fleury, G. & Maganuco, S. Razanandrongobe sakalavae, a gigantic mesoeucrocodylian from the Middle Jurassic of Madagascar, is the oldest known notosuchian. PeerJ 5, e3481 (2017).
Golz, D. J. & Lillegraven, J. A. Summary of known occurrences of terrestrial vertebrates from Eocene strata of southern California. Cont. Geol. Univ. Wyoming 15, 43–65 (1977).
Ezcurra, M. D. & Agnolín, F. L. A new global palaeobiogeographical model for the Late Mesozoic and Early Tertiary. Syst. Biol. 61, 553–566 (2012).
Maganuco, S., Dal Sasso, C. & Pasini, G. A new large predatory archosaur from the Middle Jurassic (Bathonian) of Madagascar, with remarks on its affinities and paleobiology. A. Soc. Ital. Sci. Nat. Mus. Civ. Stor. Nat. Milano. 147, 19–51 (2006).
Buffetaut, E. A zhiphodon mesosuchian crocodile from the Eocene of Algeria and its implications for vertebrate dispersal. Nature 300, 176–178 (1982).
Erickson, G. M. et al. Insights into the ecology and evolutionary success of crocodilians revealed through bite-force and tooth-pressure experimentation. PLoS ONE 7, e3178 (2012).
Peters, P. The Ecological Implications of Body Size (Cambridge University Press, Cambridge, 1983).
Calder, W. A. III. Size, Function, and Life History (Harvard University Press, Cambridge, 1984).
Bonner, J. T. Why Size Matters: From Bacteria to Blue Whales (Princeton University Press, Princeton, 2006).
Syme, C. E. & Salisbury, S. W. Patterns of aquatic decay and disarticulation in juvenile Indo-Pacific crocodiles (Crocodylus porosus), and implications for the taphonomic interpretation of fossil crocodyliform material. Palaeogeo. Palaeocli. Palaeoeco. 412, 108–123 (2014).
Hayward, J. L., Dickson, K. M., Gamble, S. R., Owen, A. W. & Owen, K. C. Eggshell taphonomy: Environmental effects on fragment orientation. Hist. Biol. 23, 5–13 (2011).
O’Connor, P. M. et al. The evolution of mammal-like crocodyliforms in the Cretaceous period of Gondwana. Nature 466, 748–751 (2010).
Ősi, A. The evolution of jaw mechanism and dental function in heterodont crocodyliforms. Hist. Biol. 26, 279–414 (2014).
Kirkland, J. I. In Dinosaur eggs and babies (eds Carpenter, K. et al.) 124–133 (Cambridge University Press, Cambridge, 1998).
Wilson, J. A., Mohabey, D. M., Peters, S. E. & Head, J. J. Predation upon hatchling dinosaurs by a new snake from the Late Cretaceous of India. PLoS Biol. 8, e1000322 (2010).
Hu, Y., Meng, J., Wang, Y. & Li, C. Large Mesozoic mammals fed on young dinosaurs. Nature 433, 149–152 (2005).
Boyd, C. A., Drumhelles, S. K. & Gates, T. A. Crocodyliform feeding traces on juvenile ornithischian dinosaur from the Upper Cretaceous (Campanian) Kaiparwits Formation, Utah. PLoS ONE 8, e57605 (2013).
Hirsch, K. J., Kihm, A. J. & Zelenitsky, D. K. New eggshell of ratite morphotype with predation marks from the Eocene of Colorado. J. Vertebr. Paleontol. 17, 360–369 (1997).
Chiappe, L. M., Salgado, L. & Coria, R. A. Embryonic skulls of titanosaur sauropod dinosaurs. Science 293, 2444–2446 (2001).
Curry Rogers, K., Whitney, M., Demic, M. & Bagley, B. Precocity in a tiny titanosaur from the Cretaceous of Madagascar. Science 352, 450–453 (2016).
Hendrickx, C., Mateus, O. & Araújo, R. A proposed terminology of theropod teeth (Dinosauria, Saurischia). J. Vertebr. Paleontol. 35, e982797 (2015).
Gianechini, F. A., Makovicky, P. J. & Apesteguía, S. The teeth of the unenlagiine theropod Buitreraptor from the Cretaceous of Patagonia, Argentina, and the unusual dentition of the Gondwanan dromaeosaurids. Acta Palaeontol. Polonica 56, 279–290 (2011).
Coria, R. A. & Chiappe, L. M. Embryonic skin from Late Cretaceous sauropods (Dinosauria) from Auca Mahuevo, Patagonia, Argentina. J. Paleontol. 81, 1528–1532 (2007).
Goloboff, P. A. & Catalano, S. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32, 221–238 (2016).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786 (2008).
Acknowledgements
The fossil preparation have been performed by Irina Fernandez in the Dinosfera (Coll de Nargó, Lleida) and Museu Conca Dellà (Isona, Lleida) equipments. CRG Marine Geosciences is recognized within the Generalitat de Catalunya excellence research groups program (reference 2017SGR315). JF and MC acknowledge support from the EU FEDER program for co-funding the purchase of the Multitom CORE X-ray CT instrument through the Spanish Ministry for Economy and Competitiveness, and from Generalitat de Catalunya autonomous government to GRC Geociències Marines (ref. 2017 SGR 315). The ICP researchers are part of the CERCA program of Generalitat de Catalunya.
Author information
Authors and Affiliations
Contributions
A.G.S., B.V., and À.G. collected the specimen of study. A.G.S., A.B, and B.V. designed the study. A.G.S. and A.B. performed the systematic and phylogenetic study. J.M. carry out the taphonomic analyses. A.G.S., F.J.L-S., and B.V. accomplished the palaeobiological study. J.F. and M.C. conducted the scanning and compilation of digital data. S.L. performed the digital segmentation and the 3D reconstruction of the specimen. All authors collected data, discussed the results, and wrote and reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
I declare that the authors have no competing interests as defined by Nature Publishing Group, or other interests that might be perceived to influence the results and/or discussion reported in this paper.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sellés, A.G., Blanco, A., Vila, B. et al. A small Cretaceous crocodyliform in a dinosaur nesting ground and the origin of sebecids. Sci Rep 10, 15293 (2020). https://doi.org/10.1038/s41598-020-71975-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-71975-y
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
