
Abstract
Animals have evolved adaptations to deal with environmental challenges. For instance, voluntarily releasing appendages (autotomy) to escape potential predators. Although it may enhance immediate survival, this self-imposed bodily damage may convey long-term consequences. Hence, compensatory strategies for this type of damage might exist. We experimentally induced autotomy in Prionostemma harvestmen. These arachnids are ideal to examine this topic because they show high levels of leg loss in the field but do not regenerate their legs. We video-recorded animals moving on a horizontal track and reconstructed their 3D trajectories with custom software tools to measure locomotor performance. Individuals that lost either three legs total or two legs on the same side of the body showed an immediate and substantial decrease in velocity and acceleration. Surprisingly, harvestmen recovered initial performance after 2 days. This is the quickest locomotor recovery recorded for autotomizing animals. We also found post-autotomy changes in stride and postural kinematics, suggesting a role for kinematic adjustments in recovery. Additionally, following leg loss, some animals changed the gaits used during escape maneuvers, and/or recruited the ‘sensory’ legs for locomotion. Together, these findings suggest that harvestmen are mechanically robust to the bodily damage imposed by leg loss.
Similar content being viewed by others

Introduction
Animals face a myriad of challenges during their lives, including predation, parasitism, navigating obstacles, and physiological stress. These challenges often lead to damage and many animals have evolved adaptations to compensate for these injuries. Compensation for damage often involves gradual improvements using developmental, morphological, or behavioral changes1,2. For instance, animals such as lizards, crickets and damselfly larvae reduce their mobility and become more cryptic after damage from potential predators3,4,5,6.
While bodily damage is often unintended, in some species injury is self-imposed and potentially adaptive. For example, many animals voluntarily lose appendages when grabbed by potential predators, a defensive strategy known as autotomy7. Although important for immediate survival5,7,8,9,10, the loss of body parts may compromise other aspects of organismal function and, by extension, an individual’s long term fitness11,12. Effects of autotomy include changes for locomotion, foraging, development, sensory biology, longevity, migration, and survival13,14,15.
With regards to locomotion, stability and maneuverability are altered by autotomy, as found for green anole lizards16 and leopard geckos17. Locomotor performance (i.e. acceleration or velocity), often interpreted as evidence of the ability to escape a potentially dangerous interaction, is also affected by autotomy7. Accordingly, wolf spiders missing legs are slower than intact individuals when running18,19. Besides performance metrics, another set of movement parameters (stride and posture kinematics) can change after autotomy. For instance, stride length and duty factor (the proportion of time during a stride that each leg is on the substrate instead of the air) changed in cellar spiders when walking on inclines20. Forelimb stance width and limb posture changed in Anolis lizards when running on narrow surfaces16. Additionally, the overall timing of all the legs used during locomotion (gaits) by arthropods has provided important insights on the effects of leg loss. Changes in the timing of using specific legs, as well as the multi-legged coordination to modify tetrapod or tripod gaits after autotomy has been recorded in tarantulas21, cockroaches22,23, crabs24, and ants25.
Given these negative impacts of autotomy on locomotion, a critical question is whether animals can compensate for this. Monitoring post-autotomy locomotor performance over time generates insights into the mechanisms for compensation and recovery. For example, in leopard geckos, body posture during locomotion changed immediately after tail autotomy but recovered to initial levels in 2–10 weeks, matching the time course for tail regeneration26. In contrast, tail autotomy in skinks caused a decrease in sprint speed, which did not recover after 4 weeks, even though endurance recovered over that timeframe27. Finally, among Anolis lizards, some individuals recovered in-air stability over the course of five weeks post tail autotomy28. Together, these findings suggest that some animals can recover locomotor performance over time. Additionally, wolf spiders undergo postural and kinematic adjustments aimed to maintain stability while moving after losing legs29. Some spiders changed their gait to a modified tripod gait or moved with an ablated tetrapod gait29. To maintain speed and stability after autotomy, spiders made small spatial changes in each leg position, as well as decreases in the proportion of time during a stride in which only two legs were on the ground as opposed to four29.
Here, we tested for the long-term consequences of autotomy of locomotion, as well as the potential kinematic and behavioral strategies harvestmen can use to mitigate the effect of bodily damage. This group of arachnids is ideal for exploring this topic because natural levels of leg autotomy are high, ranging between 33 and 58% of individuals in a population30,31,32,33. Additionally, harvestmen do not regenerate their legs even if lost before maturity34, contrary to other arthropods that regenerate legs after molting.
Despite having eight legs, harvestmen move using six legs in an alternate-tripod gait35,36, similar to terrestrial insects36,37,38,39. Legs of the second pair serve a sensory function40. Harvestmen use up to four different gaits to move (running, stotting, bobbing and walking), each of which differs in their kinematics, trajectory, and gait diagrams35. Previous work has shown that harvestmen missing legs experience immediate decreases in speed that are attributable to substrate properties and the number of legs lost32,33. To date, however, recovery from leg loss has not been examined, nor have the kinematic mechanisms associated with recovery.
The goal of this study was to experimentally test the hypothesis that autotomy affects locomotor performance, from which Prionostemma harvestmen can recover. We predicted that (1) locomotor performance would decline immediately after autotomy, but (2) harvestmen would recover performance over time, which would correlate with kinematic adjustments. Additionally, we expected that (3) negative consequences of leg loss on velocity and acceleration would be greater for animals missing more legs, missing locomotor (versus sensory) legs, and missing legs on the same (versus opposite) sides of the body. Finally, we predicted (4) changes in the gait performed and/or the legs used during locomotion. Altogether, we expected harvestmen to be robust to bodily damage, defined in this case as the ability to withstand and overcome the negative consequences of autotomy on locomotion.
Materials and methods
Study animals
Research occurred in the Neotropical lowland rainforest at La Selva Biological Station, Costa Rica (10° 26′ N, 84° 00′ W, 50 m elevation), from June to August 2015. We studied a currently undescribed species of Prionostemma Pocock harvestmen (Opiliones: Sclerosomatidae, referred to as P. sp1. in Refs.35,41,42,43, which roosts during daytime in tree trunks, buttresses, and palm leaves44. At nighttime, they actively forage on the ground and foliage41,42. Harvestmen face a wide diversity of predators, including small mammals, lizards, frogs, spiders, centipedes, and insects45. We collected 135 eight-legged individuals and placed them in clear plastic deli containers (15 × 12 × 10 cm) 24 h before trials. Individuals were fed cucumber, apple, and cat food in captivity every 2 days.
Experimental setup
We recorded harvestmen moving during daytime across a horizontal arena (Supplementary Video S1) in the lab, using the same procedure as Ref.35. To simulate predation attempts, we grabbed individuals by their hind legs and then released them. Hence, we interpreted the animal’s subsequent movements as escape behaviors46. Trials were video recorded with a GoPro HERO4 camera (GoPro, San Mateo, CA, USA) at 120 frames/s. A mirror at 45° perpendicular to the ground on the opposite side of the arena to the camera allowed recording lateral and dorsal views.
The accurate 3D body’s location in the videos was obtained as follows. The focal length, optical center, and radial and tangential lens distortions (i.e. intrinsic camera parameters) for the GoPro camera and lens were obtained using the built-in checkerboard calibration app in MATLAB vR2016a (The Mathworks, Natick, MA, USA). Then, the translation and orientation of the lateral and dorsal views relative to the track (i.e. the extrinsic camera parameters) were estimated using an M-estimator sample consensus algorithm.
Experimental treatments and trials
We induced autotomy by gently grasping the femur of the target leg with forceps as Refs.20,31,33, which immediately resulted in the release of that leg at the coxa-trochanter joint. We measured locomotion for each animal at seven different times: once prior to autotomy, once immediately after, and then 2 h, 6 h, 24 h, and 2 days after autotomy.
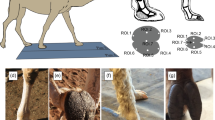
The experimental treatments used here varied in the number and type of legs missing (Fig. 1) to reflect the natural occurrence of autotomy. The treatments are named with a number representing the number of legs lost, and a letter representing the type of loss. Treatments were (1) C: intact control individuals with 8 legs. Legs were grabbed with forceps but without inducing autotomy, (2) 1L: missing the right locomotor leg I, (3) 2L: missing both of the locomotor legs I, (4) 2S: missing both sensory legs (legs II), (5) 2A: asymmetrical loss, missing legs I and III from the same side, and (6) 3L: missing three legs (both locomotor legs I and one leg II). Individuals were randomly assigned one treatment (n = 21–24 per treatment). We induced autotomy of hind legs, given that our field survey showed that missing hind legs was 53% more likely than missing rear legs.

Experimental comparisons. Body diagrams in the center column represent Prionostemma sp.1 harvestmen seen in dorsal view. The main prediction for each comparison is described in the right column. Figure was created in Microsoft Power Point version 16.39 (URL: https://www.microsoft.com/en-us/microsoft-365/microsoft-office).
Video analyses
We digitized videos using the custom scripts developed in Mathematica 10.4 (Wolfram Research, Inc., Champaign, IL, USA) described in Ref.35. Briefly, scripts automatically tracked the position of the animal’s body across each view. We then reconstructed its three-dimensional trajectory over time using built-in functions (i.e. estimateFundamentalMatrix and triangulate) and tools developed by Ref.47 for MATLAB. Using the XYZ trajectory of body position, we calculated the kinematics of the animal’s center of mass (CoM). Automatically tracking the movement of each leg was not possible, unfortunately. Harvestmen legs are very thin (Supplementary Video S1), which prevents a good contrast with the background.
We calculated nine performance, postural, or stride variables to describe harvestmen locomotion as in Escalante et al.35. For performance metrics, we used the XYZ positions over time to calculate (1) the average horizontal velocity, hereafter referred to as ‘velocity’, calculated as v_h = (x_final − x_initial)/(t_final − t_initial), where x_initial and x_final are the (x, y) coordinates of the body at the start and end of the trial, respectively. (2) Maximal horizontal acceleration, calculated as a_hmax = max_{t \in trial} a_h(t), where a_h(t) is the horizontal component of the acceleration calculated from a quintic smoothing spline fit to 3D position over time. We consider these variables reflect biologically relevant performance. We assumed that harvestmen would aim to sustain fast speed to avoid being captured (velocity), as well as a fast burst of speed to quickly move away from a potential predator (acceleration).
For postural variables, we calculated (3) the three-dimensional sinuosity normalized by time. This unitless measurement is the total path length of the trajectory divided by the linear distance between the endpoints and quantifies the lateral and vertical deviations from a straight path48. We also measured (4) the minimal and (5) the maximal height of the CoM.
For stride kinematics, we visually followed the movement of a focal leg (left leg I). We noted the time when each leg was on the ground (stance phase), and when it was lifted (aerial phase), which together represent one stride. For 2L and 3L treatments we followed the third left leg as the focal leg. We followed three strides to calculate (6) the average duty factor, the proportion of time during each stride that the focal leg was on the ground, (7) average stride frequency, the number of complete strides per second, (8) average stride period, the time to complete one stride, and (9) average stride length, the maximal distance along the x-axis the leg moved during one stride.
To investigate patterns of leg use, we visually followed all legs during three strides and constructed gait diagrams. We did this for five individuals in each experimental treatment, before autotomy, immediately after, as well as 2 days after autotomy. Finally, we visually scored each video based on the type of gait performed. We grouped the type of gaits into “fast gaits” (running and stotting) and “slow gaits” (bobbing and walking). We grouped gaits this way because performance (velocity and acceleration) is similar within gait groups35. Additionally, our focus here was on understanding the consequences of autotomy regardless of gait type.
Body measurements
We measured the length of the left leg IV for each individual to the nearest 0.05 mm using digital calipers. Leg IV length (see Supplementary Table S2) is a good proxy of body size since leg IV was never autotomized, and it correlated with leg I and III lengths (r = 0.39, 0.49, respectively. P < 0.02 for both. See Supplementary Fig. S1). Leg morphology is not sexually dimorphic40 and thus we included both adult females and males in this study. Voucher specimens of all individuals are preserved in 70% ethanol in the Essig Museum of Entomology, University of California Berkeley.
Data analyses
To test the influence of autotomy on the locomotor performance we performed generalized linear mixed models (GLMMs) using velocity or acceleration as the response variable. Predictor variables included as fixed effects were treatment, time since autotomy (treated as categorical, hereafter referred to as ‘time’), gait group (fast or slow), and all possible interactions (Table 1). Individual identity was included as a random effect to account for repeated measurements. Preliminary analyses revealed that neither the leg length nor sex affected locomotor variables (GLM: leg length, sex, and leg length × sex interaction all P > 0.42). Hence, these variables were excluded from the final models. None of the nine kinematic variables were normally distributed (Shapiro tests P < 0.05). However, GLMMs and GLMs are robust to deviations from normality, so they were useful for between treatment and time comparisons.
To identify the variables (and interactions) that affected locomotor performance we ran likelihood ratio tests49 for each model. Three sets of GLMMs (Fig. 1, Table 1) compared for the effect of (1) the number of missing legs (comparing C, 1L, 2L, and 3L treatments), (2) the type of missing leg (C, 2L and 2S), and (3) the side of the body where harvestmen lost legs (C, 2L, and 2A). Post hoc Tukey comparisons examined for differences within treatments over time, as well as differences between treatments at a given time after autotomy. We defined locomotor recovery as the first time point at which the mean locomotor performance was statistically indistinguishable from pre-autotomy levels following experimental leg loss.
Lastly, to examine potential changes in the gait use after autotomy, we compared the number of individuals in each treatment that performed each gait type before and immediately after autotomy using an independence chi-square. To examine gait type changes over time we compared the numbers before and two days after autotomy with an independence chi-square for each treatment. Statistical analyses were run in R (R Development Core Team 2018). A small portion of this dataset (the before autotomy trials of all individuals) were collected as part of another study describing gait kinematics35. Hence, information collected from some of the same individuals in Ref.35 were included in the current dataset, along with five other time points.
Results
Autotomy in the field
The levels of autotomy are high in this population. Of 399 surveyed individuals, 69% were missing at least one leg, 38% of animals were missing one leg, 23% missing two, 8% missing three, and 1% were missing four legs. We also observed that 2% of animals were missing the two sensory legs, 4% were missing one locomotor leg on each side of the body, and 3% were missing two locomotor legs on the same side of the body.
Locomotor performance
Number of legs lost
We found that velocity and acceleration differed between leg conditions and over time (GLMM: both factors P < 0.0001, Table 1, Supplementary Table S1). Post hoc comparisons revealed that locomotor performance did not differ among treatments before autotomy (P = 0.17, Fig. 2), but differed immediately after autotomy (P < 0.0001, Fig. 2). Specifically, animals missing three locomotor legs (3L treatment) decreased in both measures of performance (P < 0.05 for both comparisons). No changes were observed in the Control, 1L or 2L treatments (all tests P > 0.05). There was a significant treatment × time interaction (P = 0.02) in velocity, but not for acceleration (Table 1). Velocity decreased 40% on average and acceleration decreased 54% on average immediately after autotomy in 3L individuals (Fig. 2). However, animals in the 3L treatment recovered pre-autotomy velocity two days after leg loss (Fig. 2, Supplementary Video S1). At that point, velocity was indistinguishable from pre-autotomy levels (P > 0.05 for all Tukey tests) (Fig. 2, Supplementary Fig. S3 and Supplementary Tables S1, S3). Additionally, velocity was indistinguishable from pre-autotomy levels two days after autotomy in all treatments (see Supplementary Fig. S3). The recovery of acceleration occurred even earlier, 6 h after leg loss (Fig. 2). These findings suggest that harvestmen that lost three locomotor legs completely recovered locomotor performance two days after autotomy.

Locomotor performance over time for different levels of leg loss. Mean velocity (top) and acceleration (bottom) (± standard error) over time in Prionostemma sp.1 harvestmen with different levels of leg loss. Treatments here test for the effect of the number of legs lost. Dotted circles represent statistically significant differences from pre-autotomy levels of that treatment. Major effects of time and treatment are based on the GLMMs tests (Table 1) and the dotted circles reflect post hoc tests (see Supplementary Table S3). Graph shows all gaits pooled. Plot shows data up to 2 days after autotomy. Sample size (N) is included on the top of each treatment’s diagram. Figure was created in Microsoft Excel version 16.39 and Microsoft Power Point version 16.39 (URL: https://www.microsoft.com/en-us/microsoft-365/microsoft-office).
Type of missing legs
The type of leg lost did not affect locomotor performance after autotomy. Individuals that lost two locomotor or two sensory legs did not experience declines in either velocity or acceleration after autotomy. Further, there were no significant treatment × time interactions (P > 0.05 for both comparisons) (Fig. 3, Table 1, Supplementary Table S1).

Locomotor performance over time for different combinations of leg loss and loss of different leg types. Mean velocity (top) and acceleration (bottom) (± standard error) for different patterns of leg loss in Prionostemma sp.1 harvestmen. Treatments here show the effects of (1) the type of leg missing (C-2L-2S), and (2) the symmetry of loss (C-2L-2A). Dotted circles around the means represent statistically significant differences with pre-autotomy levels performance of that treatment. Major effects of time and treatment are based on the GLMMs tests (Table 1) and the dotted circles reflect post hoc tests (see Supplementary Table S3). Graph shows all gaits pooled. Plot shows data up to 24 h after autotomy . Sample size (N) is included on top of each treatment’s diagram. Figure was created in Microsoft Excel version 16.39 and Microsoft Power Point version 16.39 (URL: https://www.microsoft.com/en-us/microsoft-365/microsoft-office).
Asymmetry of loss
Immediately after autotomy, individuals that lost two locomotor legs on the same side (2A treatment, Fig. 1) experienced decreases in mean velocity and acceleration of 43% and 46%, respectively (Fig. 3, GLMM post hoc tests: P < 0.05). When comparing Control, 2L and 2A treatments we found significant treatment × time interactions for both velocity and acceleration (P = 0.02 and P = 0.04, respectively, Table 1, Supplementary Table S2). Despite these decreases, 2A individuals recovered performance to pre-autotomy levels after two hours (Figs. 3, 4, Supplementary Video S1, post hoc tests: P > 0.05).

Summary of kinematic changes after leg loss in Prionostemma sp.1 harvestmen. (A) Possible patterns of kinematic changes. Different colors correspond to different patterns. (B) Post hoc Tukey results of the GLMs showing within treatment comparisons. Color codes corresponding to (A) reflect statistical analysis (see Supplementary Table S3). (C) schematic representation of the experimental treatments. X = leg loss. Figure was created in Microsoft Excel version 16.39 and Microsoft Power Point version 16.39 (URL: https://www.microsoft.com/en-us/microsoft-365/microsoft-office).
Stride and postural kinematics
Number of legs lost
The number of legs lost affected stride length and frequency. Immediately after autotomy, harvestmen that lost three legs moved with fewer strides (i.e. with a reduced stride frequency). However, stride frequency increased back to pre-autotomy levels after 2 days for 3L harvestmen (Fig. 4, Supplementary Tables S1, S3). Stride period and duty factor showed the opposite pattern: increased immediately after autotomy, but decreased to pre-autotomy levels over time (Fig. 4). Harvestmen that lost one or two locomotor legs (1L and 2L, respectively) also increased their duty factor (Fig. 4). Additionally, 2L and 3L harvestmen moved with a lower minimal height (Fig. 4), suggesting a more crouched posture after autotomy. Contrary to 3L harvestmen, 1L and 2L individuals had longer strides length after autotomy (Fig. 4). Together, these patterns suggest that modifications in stride and postural kinematics are dependent on the intensity of autotomy, and become more acute with the number of legs lost.
Type of missing leg
Harvestmen that lost two sensory legs (2S) displayed changes in stride period, stride length, and minimal height after autotomy (Fig. 4, Supplementary Table S1). Thus, after leg loss, 2S individuals moved with slower and longer strides, as well as with a more crouched posture. These changes resemble those of 2L animals (see above), with the exception of the changes in stride period, which recovered to pre-autotomy levels 2 days later.
Asymmetry of loss
Animals that lost two locomotor legs on the same side (2A) displayed changes in all stride and postural variables examined (Fig. 4, Supplementary Table S1). These harvestmen started walking with longer, slower and fewer strides, as well as a more crouched posture with a more sinuous trajectory, compared with their pre-autotomy movement patterns. Over time, only some of these variables went back to pre-autotomy levels (Fig. 4). However, no clear pattern of recovery occurred for stride and postural kinematics for 2A individuals.
Gait types
Locomotor performance consistently differed between gait types. Running and stotting had higher velocity and acceleration than bobbing and walking. These differences occurred both before and after autotomy, as well as in every time frame analyzed (see Supplementary Fig. S2 and Table S3). The six treatments analyzed showed the same pattern of consistent differences between fast and slow gaits over time, as there was no significant interaction regarding gait pair in any model (Table 1).
Across trials, running was the most commonly performed gait, whereas stotting was the least common (Table 2). The frequency of individuals performing a given gait type did not vary after autotomy for Control, 1L, or 2L animals. However, in the 2A and 3L treatments, the number of individuals walking increased after autotomy, and decreased for running (Fig. 5, Table 2). Gait frequency did not return to pre-autotomy levels (Fig. 5, Table 2). On the other hand, harvestmen that lost two sensory legs showed a different pattern, in which the proportion of individuals bobbing increased over time and did not decrease to pre-autotomy levels (Table 2).

Frequency of gait types performed over time by Prionostemma sp.1 harvestmen in individuals that lost three legs (3L treatment) (n = 23). See “Methods” and Escalante et al.35 for further description of the types of gaits. P values are from independence chi-squares (Table 2). Similar patterns were found for 2S and 2A treatments (see “Results” and Supplementary Table S3). Figure was created in Microsoft Excel version 16.39 (URL: https://www.microsoft.com/en-us/microsoft-365/microsoft-office).
Changes in leg use
Autotomy affected which legs were used during locomotion (Fig. 6). Immediately after autotomy, leg II was used to move, which did not occur in intact or control individuals, which alternated between the 1R-3L-4R and 1L-3R-4L tripods (Fig. 6). Recruitment of leg II for locomotion occurred across all gait types (Fig. 6). After autotomy, 1L individuals started using the leg II (sensory leg) located next to the leg that was lost, matching the original tripod-gait (Fig. 6). Additionally, 2L individuals (missing both first legs) used one or both legs II during locomotion. The resulting alternating tripod gait for 2L individuals was 2L-3R-4L and 2R-3L-4R (Fig. 6). Harvestmen with an asymmetric loss (2A) used the leg II of the autotomized side to move. In that case, leg II replaced the lost leg III, instead of replacing leg I as in the 1L and 2L treatments. The new leg pattern for 2A individuals was 1R-2L-4R and 3R-4L (Fig. 6). Finally, 3L individuals showed major changes in their tripod gait after autotomy. They started using the remaining leg II to move, which replaced the missing leg I on the same side. Their new gait pattern was 3R-4L and 2R-3L-4R. Control and 2S animals did not display changes in the legs used to move over time.

Gait diagrams of the four gaits performed by Prionostemma sp.1 harvestmen. Bars represent when each leg (1, 2, 3 or 4; from either the left or right side of the body) was on the ground. Each panel represents the same individual before and immediately after autotomy. These are representative harvestmen from different treatments that performed the same type of gait in both time frames. Note the changes in using legs II. Treatment nomenclature: the number represents the number of legs animals lost and the letter represents the type of loss (L: missing locomotor legs, S: missing sensory legs, and A: missing two locomotor legs in the same side of the body). Leg groups in parenthesis represent the two groups of leg tripods used during locomotion after leg loss. Black and grey color coding represent the original tripods when animals have an intact leg condition (Sensenig and Shultz36; Escalante et al.35). Legs II are not shown in the left side panels because harvestmen do not use them to move before autotomy in any treatment or gait. Control or 2S individuals are not shown as they never used legs II to move. Figure was created in Microsoft Power Point version 16.39 (URL: https://www.microsoft.com/en-us/microsoft-365/microsoft-office).
Discussion
Consequences of leg loss and locomotor recovery
Overall, our findings demonstrate that harvestmen are robust to perturbations imposed by bodily damage, specifically leg loss. We found negative effects of autotomy on locomotor performance (velocity and acceleration) only in individuals that lost three legs total or two locomotor legs on the same side of the body. In contrast, performance was not affected by the loss of one or two locomotor legs or two sensory legs. Therefore, we found an effect of autotomy based on the number of legs lost and the asymmetry of autotomy, but not on the type of leg lost. It is possible that the immediate decrease in locomotor performance was due to balance disruptions of the standard alternating tripod gait used by harvestmen35,36, only after surpassing a certain threshold of damage. Autotomy can impede the ability for the remaining legs to alternate between stance and aerial phase, potentially by decreasing the torque produced by the power stroke from the body side with missing legs18. Ultimately, these changes will prevent animals from quickly moving away from potentially agonistic stimuli.
For situations in which performance decreased immediately after autotomy, recovery of initial levels of velocity and acceleration occurred within two days of leg loss. Overall, our findings indicate that harvestmen are capable of compensating for the permanent loss of legs. Additionally, our findings represent the quickest recovery of locomotor performance recorded in animals to date7.
Despite the observed rapid recovery of locomotor performance after autotomy, differences in the magnitude of leg loss may affect the survival of Prionostemma harvestmen. For instance, missing a large number of legs, or missing many legs on one side of the body only could impact locomotion severely, as found here. Individuals missing three or more legs or more than two legs on the same side were rare in our study population as well as in other species (I. Escalante, unpublished), suggesting that such levels of autotomy negatively affect survival. Consequently, the 2 days immediately after autotomy—i.e., before recovery—could be a particularly vulnerable period for these arachnids. The specific body location where loss happens might also influence the effects of autotomy on locomotion. Losing rear legs can modify gaits more substantially since legs IV showed a higher contact during stance phase when compared to hind legs. Our design did not test for this possibility. However, future research will address this topic.
Compensation for leg loss
Kinematics of locomotion
Our findings suggest that recovery of locomotor performance is associated with fine-scale adjustments in stride and postural kinematics. Individuals that experienced extensive leg loss (i.e. 2A and 3L) reorganized their kinematics through subtle adjustments. These harvestmen increased stride frequency and decreased stride period and duty factor. In particular, duty factor shifts after leg loss occurred in all treatments except in individuals that lost the two sensory legs (2S), suggesting its importance for compensation. Changes in duty factor have important implications to the interchange of potential and kinetic energy (as well as elastic energy), the ground reaction force exerted on the legs50, and the energetics of locomotion, as51 found an increase in oxygen consumption after leg loss in crabs.
Additionally, changes in gait type, as well as the legs used for locomotion played a role in recovery. Similar mechanisms for compensation following autotomy have been reported for green anoles16 and leopard geckos17,26. Changes in the position and time of leg stance phases after autotomy in wolf spiders allowed them to avoid negative consequences of autotomy on running stability and performance29.
Given the absence of regeneration, harvestmen are expected to quickly adapt to leg loss. For species that regenerate appendages, their relative robustness to damage and the ability to recover has been found to correlate with the time needed to fully regenerate. These patterns have been found after tail regeneration in skinks27, lizards28, and geckos26. Our findings suggest that, in the absence of limb regeneration, kinematic and behavioral compensatory mechanisms might assist the recovery and improve robustness to bodily damage, as found for ants during the homing navigation after leg loss25.
Gait type
We found that the gait-specific variation in performance35 did not affect recovery. After autotomy, however, harvestmen did change the frequency of gait types used to move. Particularly, walking became more frequent after autotomy in animals that lost more than one leg. Additionally, bobbing became more common only in animals that lost both sensory legs. These changes in gait types used may reflect biomechanical and/or physiological constraints imposed by a new leg combination. Running gaits are strongly associated with elastic energy recovery36. Consequently, leg loss might affect the functionality and use of the spring-mass dynamics, as the symmetry of that recovery is disrupted. Regardless of this, individuals that experienced high levels of leg loss were less able to move using the conventional alternating tripod gait for harvestmen36, which may increase the frequency of non-running gaits. Additionally, losing legs decreased the number of spring-mass monopods acting on the total stiffness50 of the system. This makes more dynamic gaits (i.e. running and stotting) less favorable in terms of energetics and ground reaction forces50. Therefore, decreases in the stiffness of the overall locomotor system of harvestmen may explain why gaits such as bobbing and walking became more common after autotomy.
Alternatively, changing the gait type performed may be adaptive. The different gaits of Prionostemma harvestmen have been suggested to confer distinct benefits in terms of predator avoidance35 or represent different optima in the physiological costs of locomotion. Consequently, post-autotomy modifications in gait may reflect adaptive adjustments to changes in predation risk or costs of moving. Post-autotomy changes in anti-predatory behavior have also been interpreted as compensation in snails52, dragonfly larvae53, and lizards16,54. Whether changes in the gait type use are due solely to mechanical constraints or if bodily damage promotes behavioral plasticity is unclear and can be addressed in future research.
Legs used for locomotion
Plasticity in leg function can allow harvestmen to compensate for the effects of leg loss. The ‘sensory’ legs acquired a locomotor function in this study. In animals whose performance did not change post-autotomy (1L, 2L, and 2S), individuals began using the sensory legs in locomotion immediately after leg loss, which does not occur in intact eight-legged animals35. These findings are novel in arthropods and suggest a redundant locomotory function to the sensory legs. Sensory legs thus not only serve an “antenniform” function55 but also serve as backup locomotor legs in case of leg loss. Having this redundant function is only possible since the ‘sensory appendages’ of harvestmen retain the same leg shape, as opposed to differently-shaped appendages in insects or the “antenniform” legs of whip spiders. Hence, maintaining that shape over evolutionary time will benefit the robustness for leg loss in these arthropods. Previous research has also reported the recruitment of food-handling appendages for locomotion after leg loss, including the pedipalps in tarantulas21 and the chela in crabs24.
The specific kinematic mechanisms promoting recovery remain unknown for these harvestmen. Changes in the use of the recently-recruited leg II for locomotion could contribute to the locomotor recovery observed here. Additionally, after leg loss harvestmen could modify the stance phase of that leg, as well as of other legs. Along with changes in minimal height during locomotion, we found changes in duty factor over time for one leg. Future studies can address the quantitative variation across treatments, time, and gaits of postural and stride kinematics of all legs. This could elucidate the fine-scale mechanisms allowing locomotor recovery. This will require higher resolution information about what each leg is doing over time. However, since Prionostemma harvestmen legs are so thin we were not able to automatically track their movement.
Leg multi-functionality raises the possibility of movement and sensory trade-offs after autotomy, suggesting that compensation may have other consequences. In this experiment, harvestmen that lost both sensory legs were the only group that increased the proportion of bobbing, even though they did not show other changes in performance. This suggests that bobbing can trade-off with sensory performance, and 2S harvestmen are relying on this gait more often potentially to gather information from the environment. However, we do not have evidence for this claim, but future research can address the interactive effects of autotomy on locomotion and foraging, sensory abilities, survival, or reproduction.
Conclusions
Our findings suggest that harvestmen are robust to bodily damage. For most conditions of leg loss tested here, harvestmen rapidly altered features of their locomotion to maintain and/or regain performance. In extreme cases (asymmetric leg loss, loss of three legs), locomotor performance decreased after leg loss, but harvestmen were still able to recover relatively quickly. We found that adjustments in the kinematics of locomotion (including changes in duty factor, stride frequency, stride period, and body height) and changes in the legs used to move were correlated with recovery of locomotor performance. Bodily damage due to autotomy is widespread across taxa and we suggest that many species may have evolved mechanisms to compensate for the diminished performance caused by injuries. Despite the fact that our analyses did not allow us to unravel the kinematic mechanisms promoting recovery, we suggest that compensation is important, yet often overlooked facet of fitness. Accordingly, comparative studies of animals with different body plans, natural histories and physiologies are important to understanding the evolution of responses to limb loss and other forms of bodily damage.
Data availability
The complete dataset included in this study is available on Dryad here.
References
Bauman, J. M. & Chang, Y.-H. Rules to limp by: Joint compensation conserves limb function after peripheral nerve injury. Biol. Lett.9, 20130484–20130484 (2013).
Suter, E., Herzog, W., Leonard, T. R. & Nguyen, H. One-year changes in hind limb kinematics, ground reaction forces and knee stability in an experimental model of osteoarthritis. J. Biomech.31, 511–517 (1998).
Cooper, W. E. Jr. Compensatory changes in escape and refuge use following autotomy in the lizard Sceloporus virgatus. Can. J. Zool.85, 99–107 (2007).
Wilson, D. & Cooper, J. W. How to stay alive after losing your tail. Behaviour145, 1085–1099 (2008).
Formanowicz, D. R. The antipredator efficacy of spider leg autotomy. Anim. Behav.40, 400–401 (1990).
Bateman, P. W. & Fleming, P. A. Sex, intimidation and severed limbs: The effect of simulated predator attack and limb autotomy on calling and emergence behaviour in the field cricket Gryllus bimaculatus. Behav. Ecol. Sociobiol.59, 674–681 (2006).
Emberts, Z., Escalante, I. & Bateman, P. W. The ecology and evolution of autotomy. Biol. Rev.94, 1881–1896 (2019).
Daniels, C., Flaherty, S. & Simbotwe, M. Tail size and effectiveness of autotomy in a lizard. J. Herpetol.20, 93–96 (1986).
Johnson, S. A. & Jakob, E. M. Leg autotomy in a spider has minimal costs in competitive ability and development. Anim. Behav.57, 957–965 (1999).
Ramsay, K., Kaiser, M. & Richardson, C. Invest in arms: Behavioural and energetic implications of multiple autotomy in starfish (Asterias rubens). Behav. Ecol. Sociobiol.50, 360–365 (2001).
Amaya, C. C., Klawinski, P. D. & Formanowicz, D. R. J. The effects of leg autotomy on running speed and foraging ability in two species of wolf spider (Lycosidae). Am. Midl. Nat.145, 201–205 (2001).
Taylor, P. W., Roberts, J. A., Wignall, A. E. & Uetz, G. W. Foreleg autotomy reduces mating success of male Schizocosa ocreata wolf spiders. J. Insect Behav.21, 193–202 (2008).
Roth, V. D. & Roth, B. M. A review of appendotomy in spiders and other arachnids. Bull. Br. Arachnol. Soc.6, 137–146 (1984).
Fleming, P. A., Muller, D. & Bateman, P. W. Leave it all behind: A taxonomic perspective of autotomy in invertebrates. Biol. Rev. Camb. Philos. Soc.82, 481–510 (2007).
Maginnis, T. L. The costs of autotomy and regeneration in animals: A review and framework for future research. Behav. Ecol.17, 857–872 (2006).
Hsieh, S. T. Tail loss and narrow surfaces decrease locomotor stability in the arboreal green anole lizard (Anolis carolinensis). J. Exp. Biol.219, 364–373 (2016).
Jagnandan, K. & Higham, T. E. Lateral movements of a massive tail influence gecko locomotion: An integrative study comparing tail restriction and autotomy. Sci. Rep.7, 1–8 (2017).
Brown, C. A. & Formanowicz, D. R. The effect of leg autotomy on terrestrial and aquatic locomotion in the wolf spider Pardosa valens (Araneae: Lycosidae). J. Arachnol.40, 234–239 (2012).
Fleming, P. A. & Bateman, P. W. Just drop it and run: The effect of limb autotomy on running distance and locomotion energetics of field crickets (Gryllus bimaculatus). J. Exp. Biol.210, 1446–1454 (2007).
Gerald, G. W., Thompson, M. M., Levine, T. D. & Wrinn, K. M. Interactive effects of leg autotomy and incline on locomotor performance and kinematics of the cellar spider, Pholcus manueli. Ecol. Evol.7, 1–7 (2017).
Wilson, B. Y. D. M. Stepping patterns in tarantula spiders. J. Exp. Biol.47, 133–151 (1967).
Delcomyn, F. The effect of limb amputation on locomotion in the cockroach Periplaneta americana. J. Exp. Biol.54, 453–469 (1971).
Hughes, G. M. The co-ordination of insect movements: I the walking movements of insects. J. Exp. Biol.29, 267–285 (1957).
Barnes, W. J. P. Leg co-ordination during walking in the crab, Uca pugnax. J. Comp. Physiol. A96, 237–256 (1975).
Wittlinger, M. & Wolf, H. Homing distance in desert ants, Cataglyphis fortis, remains unaffected by disturbance of walking behaviour and visual input. J. Physiol. Paris107, 130–136 (2013).
Jagnandan, K., Russell, A. P. & Higham, T. E. Tail autotomy and subsequent regeneration alter the mechanics of locomotion in lizards. J. Exp. Biol.217, 3891–3897 (2014).
Chapple, D. G. & Swain, R. Effect of caudal autotomy on locomotor performance in a viviparous skink, Niveoscincus metallicus. Funct. Ecol.16, 817–825 (2002).
Kuo, C. Y., Gillis, G. B. & Irschick, D. J. Take this broken tail and learn to jump: The ability to recover from reduced in-air stability in tailless green anole lizards [Anolis carolinensis (Squamata: Dactyloidae)]. Biol. J. Linn. Soc.107, 583–592 (2012).
Wilshin, S. et al. Limping following limb loss increases locomotor stability. J. Exp. Biol.221, jeb.174268 (2018).
Guffey, C. Leg autotomy and its potential fitness costs for two species of harvestmen (Arachnida, Opiliones). J. Arachnol.26, 296–302 (1998).
Houghton, J. E., Townsend, V. R. & Proud, D. N. The ecological significance of leg autotomy for climbing temperate species of harvestmen (Arachnida, Opiliones, Sclerosomatidae). Southeast. Nat.10, 579–590 (2011).
Escalante, I., Albín, A. & Aisenberg, A. Lacking sensory (rather than locomotive) legs affects locomotion but not food detection in the harvestman Holmbergiana weyenberghi. Can. J. Zool.91, 726–731 (2013).
Domínguez, M. et al. Losing legs and walking hard: Effects of autotomy and different substrates in the locomotion of harvestmen in the genus Prionostemma. J. Arachnol.44, 76–82 (2016).
Gnaspini, P. & Hara, M. R. Defense mechanisms. In Harvestmen: The Biology of Opiliones (eds Pinto-Da-Rocha, R. et al.) 374–399 (Harvard University Press, Harvard, 2007).
Escalante, I., Badger, M. A. & Elias, D. O. Variation in movement: Multiple locomotor gaits in Neotropical harvestmen. Biol. J. Linn. Soc.127, 493–507 (2019).
Sensenig, A. T. & Shultz, J. W. Mechanical energy oscillations during locomotion in the harvestman Leiobunum vittatum (Opiliones). J. Arachnol.34, 627–633 (2006).
Full, R. J. & Tu, M. S. Mechanics of six-legged runners. J. Exp. Biol.148, 129–146 (1990).
Mongeau, J. M. et al. Rapid inversion: Running animals and robots swing like a pendulum under ledges. PLoS ONE7, e38003 (2012).
Seipel, J. E., Holmes, P. J. & Full, R. J. Dynamics and stability of insect locomotion: A hexapedal model for horizontal plane motions. Biol. Cybern.91, 76–90 (2004).
Shultz, J. W. & Pinto-da-Rocha, R. Morphology and functional anatomy. In Harvestmen: The Biology of Opiliones (eds Pinto-da-Rocha, R. et al.) 14–61 (Harvard University Press, Harvard, 2007).
Wade, R. R., Loaiza-Phillips, E. M., Townsend, V. R. & Proud, D. N. Activity patterns of two species of neotropical harvestmen (Arachnida: Opiliones) from Costa Rica. Ann. Entomol. Soc. Am.104, 1360–1366 (2011).
Proud, D. N. et al. Diversity and habitat use of neotropical harvestmen (Arachnida: Opiliones) in a Costa Rican Rainforest. ISRN Zool.2012, 1–16 (2012).
Grether, G. F. et al. Species differences and geographic variation in the communal roosting behavior of Prionostemma harvestmen in Central American rainforests. J. Arachnol.42, 257–267 (2014).
Grether, G. F., Levi, A., Antaky, C. & Shier, D. M. Communal roosting sites are potential ecological traps: Experimental evidence in a Neotropical harvestman. Behav. Ecol. Sociobiol.68, 1629–1638 (2014).
Cokendolpher, J. C. & Mitov, P. G. Natural enemies. In Harvestmen: The Biology of Opiliones (eds Pinto-da-Rocha, R. et al.) 339–373 (Harvard University Press, Harvard, 2007).
Wheatley, R., Niehaus, A. C., Fisher, D. O. & Wilson, R. S. Ecological context and the probability of mistakes underlie speed choice. Funct. Ecol.32, 990–1000 (2018).
Hedrick, T. L. Software techniques for two- and three-dimensional kinematic measurements of biological and biomimetic systems. Bioinspir. Biomim.3, 1–6 (2008).
Mistick, E. A., Mountcastle, A. M. & Combes, S. A. Wing flexibility improves bumblebee flight stability. J. Exp. Biol.219, 3384–3390 (2016).
Bolker, B. M. et al. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol.24, 127–135 (2009).
Blickhan, R. & Full, R. J. Similarity in multilegged locomotion: Bouncing like a monopode. J. Comp. Physiol. A173, 509–517 (1993).
Herreid, C. F. & Full, R. J. Energetics of hermit crabs during locomotion: The cost of carrying a shell. J. Exp. Biol.120, 297–308 (1986).
DeWitt, T. J., Sih, A. & Hucko, J. A. Trait compensation and cospecialization in a freshwater snail: Size, shape and antipredator behaviour. Anim. Behav.58, 397–407 (1999).
Mikolajewski, D. J. Morphological and behavioral defenses in dragonfly larvae: Trait compensation and cospecialization. Behav. Ecol.15, 614–620 (2004).
Kuo, C. Y., Irschick, D. J. & Lailvaux, S. P. Trait compensation between boldness and the propensity for tail autotomy under different food availabilities in similarly aged brown anole lizards. Funct. Ecol.29, 385–392 (2015).
Willemart, R. H., Farine, J. P. & Gnaspini, P. Sensory biology of Phalangida harvestmen (Arachnida, Opiliones): A review, with new morphological data on 18 species. Acta Zool.90, 209–227 (2009).
Acknowledgements
Funding for this project to I.E. came from the Fulbright Foreign Student Program, a Short-term Fellowship by the Smithsonian Tropical Research Institute (STRI), a Starter Grant from the Department of Environmental Science, Policy, and Management, University of California—Berkeley, and the Margaret C. Walker Fund for teaching and research in systematic entomology from UC Berkeley. M.A.B was supported by a National Science Foundation CiBER-IGERT under Award DGE-0903711 and a National Science Foundation Graduate Research Fellowship under Grant No. DGE-1106400. We are grateful to B Matarrita, D Brenes, and R Boza, who assisted with in field work. E Lewis, S Miller, K Foster, and W Liu helped with the video analysis and T Daluro, C Christensen and Winston Vo with morphological measures. Insightful comments on early versions of this manuscript were provided by one anonymous reviewer, R Gillespie, A Kamath, E Brandt, M Raboin, B Kessler, M Rosenthal, S O’Brien, E Lacey, and UC Berkeley’s Animal Behavior seminar.
Author information
Authors and Affiliations
Contributions
I.E. and D.O.E. design the study. I.E. conducted the trials. M.A.B. wrote the computing tools to analyze the videos. I.E. and M.A.B. digitized and analyzed the videos. I.E. analyzed the data, made the figures and wrote most of the manuscript. All authors contributed equally to the editing and proofreading of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Escalante, I., Badger, M.A. & Elias, D.O. Rapid recovery of locomotor performance after leg loss in harvestmen. Sci Rep 10, 13747 (2020). https://doi.org/10.1038/s41598-020-70557-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-70557-2
This article is cited by
-
Different interaction effects between two invasive harvestmen and a native relative in the laboratory
Biological Invasions (2023)
-
Robustness in action: Leg loss does not affect mating success in male harvestmen
Behavioral Ecology and Sociobiology (2022)
-
Leg loss decreases endurance and increases oxygen consumption during locomotion in harvestmen
Journal of Comparative Physiology A (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
