
Abstract
We investigated the heritability of educational attainment and how it differed between birth cohorts and cultural–geographic regions. A classical twin design was applied to pooled data from 28 cohorts representing 16 countries and including 193,518 twins with information on educational attainment at 25 years of age or older. Genetic factors explained the major part of individual differences in educational attainment (heritability: a2 = 0.43; 0.41–0.44), but also environmental variation shared by co-twins was substantial (c2 = 0.31; 0.30–0.33). The proportions of educational variation explained by genetic and shared environmental factors did not differ between Europe, North America and Australia, and East Asia. When restricted to twins 30 years or older to confirm finalized education, the heritability was higher in the older cohorts born in 1900–1949 (a2 = 0.44; 0.41–0.46) than in the later cohorts born in 1950–1989 (a2 = 0.38; 0.36–0.40), with a corresponding lower influence of common environmental factors (c2 = 0.31; 0.29–0.33 and c2 = 0.34; 0.32–0.36, respectively). In conclusion, both genetic and environmental factors shared by co-twins have an important influence on individual differences in educational attainment. The effect of genetic factors on educational attainment has decreased from the cohorts born before to those born after the 1950s.
Similar content being viewed by others

Introduction
Education is one of the most important intellectual, social and material resources in modern societies. It typically precedes occupation, strongly affects occupation-based social position and lifetime income1, and is associated with health and mortality2. The education system is also the main institution reproducing social stratification3. Accordingly, the understanding of how genetic and environmental factors underlie the individual differences in educational attainment has not only scientific but also important societal implications. A meta-analysis of twin studies conducted in 10 countries found that genetic and environmental factors shared by co-twins explain about equal shares (around 40%) of the inter-individual differences in educational level4. The proportion of contribution by shared environment was substantially higher than found in a meta-analysis of performance in school subjects5. The shared environmental variation in intelligence is still substantial in childhood, but diminishes until adulthood6. Thus, in addition to cognitive skills, childhood family and other environmental factors seem to affect educational level. A previous study demonstrated the importance of social inheritance by showing that good parental socio-economic position is associated with academic success even after adjusting for cognitive ability7.
The role of genetic and environmental factors in educational differences can also vary between societies and changes over time because of differences between educational systems and societies in general4,5. For example, in more meritocratic and open societies, the role of individual differences derived from genetic factors may be more pronounced, whereas in socially more closed societies with less social mobility, childhood social factors may be more important. Information on how the macro-environment modifies the role of genetic and environmental factors behind inter-individual variation in education is important for the understanding of how socially and politically established barriers in the society modify the realization of genetic potential for educational outcomes. An early Norwegian twin study found that the role of genetic factors on education increased after World War II when the educational system was reformed8, and a study in Spain showed similar results on the increasing effect of genetic factors on education after an educational reform promoting equality in educational opportunities9. This is further supported by a re-analysis of the published heritability estimates of educational attainment4 finding that high intergenerational mobility in a society was associated with the higher proportion of genetic and the lower proportion of shared environmental factors of the total variation of educational attainment10. These twin study results correspond well with the findings that the effect of parental socioeconomic position on offspring education has diminished over the recent decades in Europe11. The previous literature thus suggests that a more open society, in which educational reforms make the education system more open and meritocratic, will strengthen the influence of genetic factors and decrease the influence of family background on educational attainment. An Estonian study supported this conclusion by showing that previously identified genetic polymorphisms associated with educational attainment explained a larger proportion of variation in educational attainment in the cohorts who received their education after rather than before the independence of Estonia from the Soviet Union12.
In this study, we analyzed the contribution of genetic and environmental factors to educational attainment in a large multinational twin database. Based on previous twin studies8,9, we expect that the role of genetic factors in educational variation has increased over the recent decades, especially after World War II when the educational system was expanded in many countries. We further expect that the increasing general level of education may increase the proportion of genetic variation since the decision for higher education is done later in adolescence when the children have become more independent from their parents13. Societies also differ whether education is free for students or whether tuitions are needed which may affect how parental financial resources affect possibilities for higher education. Thus, we expect to see macro-level differences between countries, which may reflect access to higher education. We classified the countries in accordance with the typology of welfare regimes presented by Esping-Andersen14; European countries follow the social democratic, conservative or mixed societal models whereas the USA and Australia follow the liberal model. The East Asian countries were difficult to classify by this typology, but they share many cultural similarities. Finally, we will analyze gender differences in the proportions of genetic and environmental variation in educational attainment, and whether they have changed over the twentieth century when opportunities for higher education have equalized for men and women.
Results
Table 1 presents the descriptive statistics in the whole data as well as stratified by cultural–geographic region (the corresponding statistics by twin cohort are available in Supplementary Table 1). The educational level increased over the birth cohorts in men and women: when the most recent birth cohorts (1980–1989), where education may not yet have been finished, were removed, the increase was 0.55 (95% confidence interval (CI) 0.53–0.57) educational years per decade in men and 0.92 (95% CI 0.90–0.94) educational years per decade in women. Mean education was virtually the same in men and women (12.5 years) in the whole dataset, but the educational level increased more rapidly in women than in men (0.37 educational years more per decade in women than in men; 95% CI 0.35–0.40). In all birth cohorts, the educational level was higher in North America and Australia than in Europe or East Asia.
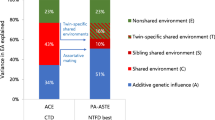
Table 2 presents the proportions of educational variation explained by genetic and environmental factors. In the whole data, the largest proportion of individual differences in education was explained by genetic factors (heritability: a2 = 0.43; 95% CI 0.41–0.44), but environmental factors shared by co-twins were also substantial (c2 = 0.31; 95% CI 0.30–0.33). When stratified by gender, we found that the proportion of variation in educational attainment attributable to genetic factors was larger in men than in women (a2 = 0.47; 95% CI 0.45–0.50 vs. a2 = 0.38; 95% CI 0.36–0.40) – and correspondingly the proportion explained by shared environmental factors smaller (c2 = 0.26; 95% CI 0.24–0.29 vs. c2 = 0.36; 95% CI 0.34–0.38). The highest heritability was found in the latest birth cohort born in 1980–1989 (a2 = 0.60; 95% CI 0.50–0.71). This result remained when we restricted the analyses only to those of 30 years of age or older to confirm that that this was not affected by unfinished education in this birth cohort (Supplementary Table 2).
We then conducted birth-cohort-specific analyses by the cultural–geographic region (Table 3). The three cultural–geographic regions did not show systematic differences in the proportions of genetic and environmental factors. When pooling all birth cohorts together, genetic factors explained virtually the identical proportions of individual differences in educational attainment in Europe (a2 = 0.40; 95% CI 0.37–0.42) as in North America and Australia (a2 = 0.39; 95% 0.36–0.41). In East Asia, the heritability point estimate was lower but did not differ statistically significantly from the other cultural–geographic regions either (a2 = 0.32; 95% 0.19–0.48); because of the smaller sample size, the confidence interval was wide. When we stratified the results by gender, there were no differences between the cultural–geographic regions (Supplementary Table 3 for men and 4 for women).
Table 4 presents the analyses stratifying cohorts born in 1900–1949 and 1950–1989 in participants at 30 years of age or older from European, North American and Australian twin cohorts together. The role of genetic factors decreased (a2 = 0.44; 95% CI 0.41–0.46 vs. a2 = 0.38; 95% CI 0.36–0.40) and the role of shared environment increased over the birth cohorts (c2 = 0.31; 95% CI 0.29–0.33 vs. c2 = 0.34; 95% CI 0.32–0.36). When we stratified these analyses into Europe and North America and Australia, we saw a similar decrease in the proportion of additive genetic factors and increase in the proportion of shared environmental factors in North America and Australia. In the gender-stratified analyses, women showed only minor decrease in the role of additive genetic factors. Thus, the difference between men and women in the proportion of variation explained by additive genetic factors was greater in the cohorts born in 1900–1949 than in 1950–1989 (a2 = 0.49; 95% CI 0.46–0.52 vs. a2 = 0.41; 95% CI 0.37–0.44 in men and a2 = 0.38; 95% CI 0.35–0.41 vs. a2 = 0.36; 95% CI 0.32–0.39 in women).
Finally, we applied the genetic models separately to each twin cohort to explore whether there was a consistent, regular pattern within the countries not captured by our classification of the three cultural–geographic regions (Supplementary Table 5). These analyses showed no systematic differences in the estimates of proportions of genetic and environmental variation between the cohorts. Confidence intervals, however, were wide in many cohorts making it difficult to draw firm conclusions. When stratifying the analyses by gender (Supplementary Tables 6 and 7), these cohort specific analyses confirmed the higher heritability estimates in men than in women: only in six out of 24 cohorts including both genders, the point estimates of heritability were higher in women than in men.
Discussion
In this dataset, pooling twin cohorts from 16 countries, we found that genetic factors explained 43% of the individual differences in educational attainment. The role of genetic factors affecting differences in educational attainment is not surprising. Intelligence, having an important effect on educational attainment, shows a substantial influence of genetic factors6, but there are also other genetic factors, which may affect educational attainment, for example, through personality aspects such as conscientiousness15. Genetic factors also contribute to the choice of subject at school, which can affect further opportunities for higher education16. The latest meta-analysis of genome-wide association (GWA) studies, including 1.1 million participants, identified 1271 genome-wide significant loci associated with education, but even together, they explained only 3.86% of the variation of education17. When using all common genetic variants (known as SNP-heritability), 12% of educational variation could be explained18. This lower proportion of genetic variation estimated by measured genetic polymorphisms than in our estimates based on twin design reflects the multifactorial and polygenic background of educational attainment; the current GWA technology does not capture all existing genetic variation.
In addition to genetic factors, our results show the importance of shared environmental factors explaining 31% of educational variation. This variation can include the effect of home environment but also the influence of school, common peers and other environmental factors shared by co-twins. This result is interesting since many previous studies have reported that shared environmental influences on intelligence6 or personality factors are small or not existing in adulthood19. Further, studies have not shown shared environmental effects for other socio-economic traits, such as income20. It is, however, noteworthy that disentangling genetic and shared environmental effects needs considerable statistical power21, which may have led to ignoring shared environmental effects in some smaller studies. A large Swedish twin study of young adult men found that shared environmental factors explained 20% of variation of intelligence22. Furthermore, differences in intelligence in childhood, influenced by greater shared environmental effect, may be more important for education than differences in adult intelligence. However, we found the substantial shared environmental variance components in nearly all twin cohorts, which were larger than generally found for intelligence, even in childhood6. These estimates were also higher than found in previous studies for academic performance at school5. Previous results have shown that parental socio-economic position affects offspring education, even after adjusting the results for cognitive ability7. A recent study demonstrated that the alleles of the educational polygenic score of parents that were not transmitted to their offspring were associated with school performance at age 1723 as well as the educational years in adulthood in offspring24. This genetic nurture effect was replicated in a Dutch study, where in adults, both transmitted and non-transmitted alleles included in educational polygenic scores were associated with offspring education25. In a UK study, polygenic education scores predicted school performance in adolescence better between individuals from different families than within dizygotic (DZ) co-twins, suggesting that a part of the genetic effect on education was because of family environment created by parental genotype26. Together, these results support the conclusion that even when family environment has a limited effect on many psychological and social outcomes27, it may be important for education. However, more research is needed to reveal the mechanisms between family background and education.
We found that genetic factors explained a larger proportion, and shared environmental factors a smaller proportion, of educational variation in the earlier born cohorts in 1900–1949 than in the later born cohorts in 1950–1989. These results, thus, do not support our initial hypothesis that the role of genetic factors declined over the birth cohorts because of higher educational level, especially in women, reflecting the expansion of higher education after World War II, which may have led to higher meritocracy. There have been somewhat contrasting results on the intergenerational transmission of education, one study suggesting diminishing effect of parental social-position in eight European countries11 and another study showing no change in the correlation between parental and offspring education in 42 countries28. The previous meta-analysis on the heritability of education, including several twin cohorts included also in our study, also found decreasing shared environmental variation in the cohorts born after 1950, but this effect was not statistically significant4. We confirmed these results by limiting these analyses to those of 30 years of age or older having thus virtually finished their education. Thus, unfinished education does not affect the results, which may happen, for example, if those with higher intelligence have finished their education in a shorter period.
Our region-specific results are also against of our prior hypothesis. We expected that shared environment would be especially large in countries following the liberal model of typology by Esping-Andersen14, i.e. North America and Australia in this study, leading to lower heritability, because in these countries parental economic resources may be more important for higher education than in the countries where higher education is free and organized by the government. However, this was not the case: the heritability estimates were at the same level as in the European countries including in this study countries following the social-democratic or conservative models. We analyzed this in more detail in individual cohorts to assess whether there would be a systematic pattern not captured by our classification of countries. However, we did not find any evidence on systematic differences in the heritability estimates between the countries. Our results suggest that shared environment has an important role in educational differences regardless of the model of society. A study showing a higher genetic effect in Estonia after re-independence from the Soviet Union indicates that the societal structure can play a role in the genetic variation of education12. However, it may be apparently limited to specific societal systems, such as former Soviet Union, or specific times in a societal development.
Our most consistent result was the larger proportion of genetic variation and consequently the smaller proportion of shared environmental variation of education in men than in women. This supports a previous meta-analysis, based mainly on published results4, but is extended with a greater number of countries, thus showing the universality of the result. Importantly, there is no evidence on a gender difference in the role of genetic and shared environmental factors in intelligence29. This suggests that family background and other environmental factors shared by co-twins are more important for women than for men in choosing to continue education. Interestingly, the differences in the heritability estimates between men and women were larger in the cohorts born in 1900–1949 than in the later cohorts born in 1950–1989 when the role of shared environment increased in men. Thus, in the later birth cohorts, the role of genetic and environmental factors converged in men and women. Parallel results have been found in molecular genetic research: A US study found that the polygenic risk score of education predicted educational years more strongly in men than in women and this difference has diminished from the cohorts born in the 1930s to 1950s30. This may be associated to more equal opportunities for women to get a higher education in the later birth cohorts, which also this study showed by an increasing education in women.
One factor which may have inflated the estimates of shared environmental variation is assortative mating. There is a well-known spousal correlation in education, which may itself change over time31. If this induces a genetic correlation between spouses, the genetic correlation between DZ twins becomes higher than 0.5 as assumed by the twin model. In a sub-cohort, we had information on maternal and paternal education (23,705 families). The spousal correlation in the parents of twins was 0.57 after adjusting for the twin cohort and the birth year of twin children used as a proxy of the missing information on the birth years of parents. If applied to the estimates of genetic and environmental variation using the formula presented by Martin32, this spousal correlation was too high even if all common environmental variation is caused by assortative mating, i.e., it should have produced even higher shared environmental variance component than we found. However, in addition to phenotypic assortment, social homogamy, i.e. similar social environment of spouses, can affect a spousal correlation, which does not generate a correlation between genotypes of spouses33. A US study found that the correlation of polygenic risk score of education was 0.13 between spouses, which was much lower than the spousal educational correlation34. Correcting shared environmental variation by this genetic correlation reduced them somewhat, but they remained substantial (c2 = 0.19 in men and c2 = 0.30 in women). However, the polygenic risk score explains only a fraction of educational variation17, and so we should make an assumption that the spouses share the same amount of unknown genetic variants affecting education than they share the known genetic variants. In a UK study using all genetic loci affecting education, the genetic correlation between spouses was even higher (rA = 0.65) than the educational trait correlation35. When applied to our estimates, this genetic correlation should have produced even higher shared environmental variation than found in this study. The higher genetic than trait correlation in education would also expect a different mechanism behind assortative mating, i.e. selecting a spouse because of genetic liability rather than the trait itself, than previously assumed. Thus, it is too early to argue how much assortative mating has inflated shared environmental variation in this study because we do not know the mechanism behind assortment well enough to be able to estimate the genetic spousal correlation. However, we found that there were no differences in spousal correlations between cohorts born 1900–1949 and 1950–1989 (r = 0.59 vs. r = 0.54, respectively) or, as expected, between the parents of males and females (r = 0.58 vs. r = 0.55, respectively) which could explain the differences in the genetic architecture between birth cohorts or between men and women. In contrast, the spousal correlations were thus even lower in the latter birth cohorts and in women, which thus cannot explain the larger shared environmental variation in these groups than in the earlier birth cohorts and men.
Our data have important strengths but also weaknesses. As compared to the previous meta-analysis of twin studies of education based mainly on published results4, our study based solely on individual level data offered several important strengths. With access to individual data, we could conduct more flexible analyses than could be done when based on published estimates. Further, our data are free of publication bias, which may lead to tendency to publish results in line of previously published heritability estimates, and we also have more data from a larger number of countries, including Asian countries, than in this previous meta-analysis4. The progress of GWA studies have allowed to estimate the role of genetic factors based on individual level data with information on common genetic variants17; GWA studies, however, lack information on rarer variants and the more complex genetic variation that is captured by whole genome sequencing. A strength of the classical twin design is that it allows separating the effects of all genetic and of environmental factors shared by co-twins, such as family background. Thus, twin studies allow for a more comprehensive understanding of how genetic and social inheritance affect the transmission of education across generations. A weakness of our data is that we have information only on accomplished education. It would be important to collect also information on school performance, intelligence, motivation and other factors affecting education. This would help to understand mechanisms behind genetic and environmental components of educational variation.
In conclusion, our international twin data demonstrated that genetic factors are important for educational attainment regardless of the society. We also found environmental variation shared by co-twins behind inter-individual differences in educational attainment, but its estimate depends how we assume assortative mating to affect the genetic correlation between spouses. The role of shared environmental variation was higher in women than in men, especially in the cohorts born before World War II. This suggests that the influence of environmental factors on education differs between men and women and this gender difference itself may change over time.
Data and methods
Sample
The data for this study were derived from the international COllaborative project of Development of Anthropometrical measures in Twins (CODATwins) database. The CODATwins project aimed to pool data from all twin cohorts in the world having information on height and weight36. However, additional information was also collected on own education37. Together, 28 twin cohorts representing 16 countries provided data on education to the CODATwins database and were included in the present study. The footnote of Table 1 shows the names of these cohorts. Among these cohorts, 13 come from Europe, 8 from the USA, two from Australia and single cohorts from South Korea, Japan, China, Brazil and Sri Lanka (Supplementary Table 1). We classified the countries in accordance with the typology of welfare regimes presented by Esping-Andersen14: European countries follow the social democratic (Finland, Norway and Sweden), conservative (German, Hungarian, Italy and Spain) or mixed (Netherlands and Belgium) societal models whereas the USA and Australia follow the liberal model. The East Asian countries (China, Japan and South Korea) were difficult to classify by this typology, but they share many cultural similarities. Sri Lanka and Brazil were not included in these region-specific analyses, because they are geographically and culturally distinct from the other regions.
All participants were volunteers and gave informed consent when participating in their original study. Only a limited set of observational variables and anonymized data were delivered to the data management center at University of Helsinki. The pooled analysis was approved by the ethical committee of Department of Public Health, University of Helsinki, and the methods were carried out in accordance with the approved guidelines.
We transformed the different educational classifications used in the original surveys from each cohort into educational years by using the mean levels of educational years in each category. The original classifications and the corresponding numbers of educational years have been described elsewhere37. To ensure that the educational attainment variable captured the participants’ final highest level of education, we removed those under 25 years of age from the sample. We used this age limit because a higher age limit would have led to a substantial loss of data in the latest birth cohorts. However, when making comparisons between birth cohorts, we used a stricter age limit, i.e. 30 years, to confirm that unfinished education would not affect the results. Further, we replicated the heritability analyses by birth cohort using this stricter age limit to analyze whether it would change the results systematically. Together, we had educational data from 193,518 twins (54% women) including 81,894 complete twin pairs; 40% of the complete twin pairs were monozygotic (MZ), 39% same-sex dizygotic (SSDZ) and 21% opposite-sex dizygotic (OSDZ). From these participants, 39,235 were under 30 years of age, and we thus removed them when using this stricter age limit. The birth years of the twins ranged from 1889 to 1989. However, we had data for only 70 men and 137 women born earlier than 1900, whom we removed from the analyses because the number was too small for birth cohort specific analyses. The largest number of twins were from Europe (N = 100,272) and North America and Australia (N = 88,106), while the number of twins was much smaller for East Asia (N = 2728). In the region-specific analyses, East Asians born before 1950 (113 men and 106 women) were removed because the number was too small for the analyses stratified by birth cohort and region.
Statistical analyses
The data were analyzed using quantitative genetic twin modeling based on structural equation models38. The twin modeling uses the different genetic relatedness of MZ and DZ twins: DZ twins share, on average, 50% of genetic variation, whereas MZ twins are virtually identical at the DNA sequence level. This principle allows decomposing the educational variation into variance attributable to additive genetic factors (A: correlated by 1.0 for MZ and 0.5 for DZ pairs), environmental factors shared by co-twins (C: by definition correlated by 1.0 for both MZ and DZ pairs), and environmental factors unique to each twin individual (E: by definition uncorrelated for MZ and DZ pairs). The unique environmental component also includes measurement error. The models were estimated using the OpenMx package (version 2.0.1) of R statistical software39. As we have reported previously37, MZ twins had slightly higher education than DZ twins, and therefore we allowed different means for MZ and DZ twins in the models.
We started the analyses by estimating the genetic and environmental variances in educational attainment in the whole dataset as well as in 10-year birth cohorts by first pooling data from all cohorts together and then stratifying them by the cultural–geographic region. In these analyses, we adjusted education by twin cohort because otherwise different educational systems between the countries and different education classifications between the surveys might have affected the results. Further, we adjusted for the effect of birth year in order to account for the effect of increasing educational level during the twentieth century in these cohorts37. The adjustments were done by calculating regression residuals of education by using birth year and the dummy variables of twin cohorts as independent variables in the regression models. We then analyzed temporal trends by pooling the birth cohorts into two broad time spans, 1900–1949 and 1950–1989, to increase power to detect differences in the genetic and environmental variation over the birth cohorts. We selected these cohorts because of the much higher education in the later born cohorts indicating extension in educational systems. We restricted these analyses to Europe and North America and Australia since older birth cohorts were not available in East Asia and in other countries. After these analyses by birth cohort, we conducted a second set of analyses and estimated genetic and environmental variance components separately in each twin cohort to confirm that the used classification did not hide any existing pattern. In these twin cohort specific analyses, we adjusted education for birth year separately in each twin cohort.
Data availability
This study is based on the re-analyses of original datasets from third parties. The data are available based on request if following the same rules and principles of CODATwins project followed also in this study. More information is available from the corresponding author.
References
Ishida, I., Müller, W. & Ridge, J. M. Class origin, class destination, and education: a cross-national study of ten nations. Am. J. Sociol. 101, 145–193 (1995).
Mackenbach, J. P. et al. Socioeconomic inequalities in health in 22 European countries. N. Engl. J. Med. 358, 2468–2481 (2008).
Hout, M. & DiPrete, T. A. What we have learned: RC28’s contributions to knowledge about social stratification. Res. Soc. Stratif. Mobility 24, 1–20 (2006).
Branigan, A. R., McCallum, K. J. & Freese, J. Variation in the heritability of educational attainment: an international meta-analysis. Soc. Forces 92, 109–140 (2013).
de Zeeuw, E. L., de Geus, E. J. C. & Boomsma, D. I. Meta-analysis of twin studies highlights the importance of genetic variation in primary school educational achievement. Trends Neurosci. Educ. 4, 69–76 (2015).
Plomin, R. & Deary, I. J. Genetics and intelligence differences: five special findings. Mol Psychiatry 20, 98–108 (2015).
Bukodi, E., Erikson, R. & Goldthorpe, J. H. The effects of social origin and cognitive ability on educational attainment: evidence from Britain and Sweden. Acta Sociol. 57, 293–310 (2014).
Heath, A. C. et al. Education policy and the heritability of educational attainment. Nature 314, 734–736 (1985).
Colodro-Conde, L., Rijsdijk, F., Tornero-Gómez, M. J., Sánchez-Romera, J. F. & Ordoñana, J. R. Equality in educational policy and the heritability of educational attainment. PLoS ONE 10, e0143796 (2015).
Engzell, P. & Tropf, F. C. Heritability of education rises with intergenerational mobility. PNAS 116, 25386–25388 (2019).
Breen, R., Luijkx, R., Müller, W. & Pollak, R. Nonpersistent inequality in educational attainment: evidence from eight European countries. Am. J. Sociol. 114, 1475–1521 (2009).
Rimfeld, K. et al. Genetic influence on social outcomes during and after the Soviet era in Estonia. Nat. Hum. Behav. 2, 269–275 (2018).
Kilford, E. J., Garrett, E. & Blakemore, S. J. The development of social cognition in adolescence: An integrated perspective. Neurosci. Biobehav. Rev. 70, 106–120 (2016).
Esping-Andersen, G. The Three Worlds of Welfare Capitalism (Polity Press, Cambridge, 1990).
Krapohl, E. et al. The high heritability of educational achievement reflects many genetically influenced traits, not just intelligence. Proc. Natl. Acad. Sci. USA 111, 15273–15278 (2014).
Rimfeld, K., Ayorech, Z., Dale, P. S., Kovas, Y. & Plomin, R. Genetics affects choice of academic subjects as well as achievement. Sci. Rep. 6, 26373 (2016).
Lee, J. J. et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat Genet 50, 1112–1121 (2018).
Broad Institute. LD Hub: a centralised database of summary-level GWAS results and a web interface for LD score regression. 2016; Available at: https://ldsc.broadinstitute.org/. Accessed 8/12, 2019.
Sanchez-Roige, S., Gray, J. C., MacKillop, J., Chen, C. H. & Palmer, A. A. The genetics of human personality. Genes Brain Behav. 17, e12439 (2018).
Hyytinen, A., Ilmakunnas, P., Johansson, E. & Toivanen, O. Heritability of lifetime earnings. J. Econ. Inequal. https://doi.org/10.1007/s10888-019-09413-x (2019).
Visscher, P. M., Gordon, S. & Neale, M. C. Power of the classical twin design revisited: II detection of common environmental variance. Twin Res. Hum. Genet. 11, 48–54 (2008).
Latvala, A., Kuja-Halkola, R., D’Onofrio, B. M., Larsson, H. & Lichtenstein, P. Cognitive ability and risk for substance misuse in men: genetic and environmental correlations in a longitudinal nation-wide family study. Addiction 111, 1814–1822 (2016).
Bates, T. C. et al. Social competence in parents increases children’s educational attainment: replicable genetically-mediated effects of parenting revealed by non-transmitted DNA. Twin Res. Hum. Genet. 22, 1–3 (2019).
Kong, A. et al. The nature of nurture: Effects of parental genotypes. Science 359, 424–428 (2018).
de Zeeuw EL, Hottenga JJ, Ouwens KG, Dolan CV, Ehli EA, Davies GE, et al. Intergenerational transmission of education and ADHD: effects of parental genotypes. Behav Genet 50, 221–232. (2020).
Selzam, S. et al. Comparing within- and between-family polygenic score prediction. Am. J. Hum. Genet. 105, 351–363 (2019).
Polderman, T. J. et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat. Genet. 47, 702–709 (2015).
Hertz, T., Jayasundera, T. & Piraino, P. The inheritance of educational inequality: international comparisons and fifty-year trends. Adv. Econ. Anal. Policy 7, 10 (2007).
Knopik, V. S. & DeFries, J. C. A twin study of gender-influenced individual differences in general cognitive ability. Intelligence 26, 81–89 (1998).
Herd, P. et al. Genes, gender inequality, and educational attainment. Am. Sociol. Rev. 84, 1069–1098 (2019).
Blossfeld, H. Educational assortative marriage in comparative perspective. Ann. Rev. Sociol. 35, 513–530 (2009).
Martin, N. G. Genetics of social and sexual attitudes. In Twin Research: Psychology and Methodology (ed. Nance, W. E.) 13–23 (Alan R. Liss Inc, New York, 1978).
Heath, A. C. & Eaves, L. J. Resolving the effects of phenotype and social background on mate selection. Behav. Genet. 15, 15–30 (1985).
Conley, D. et al. Assortative mating and differential fertility by phenotype and genotype across the 20th century. Proc. Natl. Acad. Sci. USA 113, 6647–6652 (2016).
Robinson, R. et al. Genetic evidence of assortative mating in humans. Nat. Hum. Behav. 1, 0016 (2017).
Silventoinen, K. et al. The CODATwins Project: The cohort description of collaborative project of development of anthropometrical measures in twins to study macro-environmental variation in genetic and environmental effects on anthropometric traits. Twin Res. Hum. Genet. 18, 348–360 (2015).
Silventoinen, K. et al. Education in twins and their parents across birth cohorts over 100 years: an individual-level pooled analysis of 42 twin cohorts. Twin Res. Hum. Genet. 20, 395–405 (2017).
Posthuma, D. et al. Theory and practice in quantitative genetics. Twin Res. 6, 361–376 (2003).
Neale, M. C. et al. OpenMx 2.0: Extended structural equation and statistical modeling. Psychometrika 81, 535–549 (2016).
Acknowledgements
This study was conducted within the CODATwins project. We thank Wendy Cozen, University of Southern California, for sharing data with us. Funding for collaborators: Netherlands Twin Register acknowledges the Netherlands Organization for Scientific Research (NWO) and MagW/ZonMW grants 904-61-090, 985-10-002, 912-10-020, 904-61-193,480-04-004, 463-06-001, 451-04-034, 400-05-717, Addiction-31160008, Middelgroot-911-09-032, Spinozapremie 56-464-14192; VU University’s Institute for Health and Care Research (EMGO+); the European Research Council (ERC - 230374), the Avera Institute, Sioux Falls, South Dakota (USA). Data collection and analyses in Finnish twin cohorts have been supported by ENGAGE – European Network for Genetic and Genomic Epidemiology, FP7-HEALTH-F4-2007, Grant Agreement Number 201413, National Institute of Alcohol Abuse and Alcoholism (Grants AA-12502, AA-00145, and AA-09203 to R J Rose, the Academy of Finland Center of Excellence in Complex Disease Genetics (Grant Numbers: 213506, 129680), and the Academy of Finland (Grants 100499, 205585, 118555, 141054, 265240, 263278 and 264146 to J Kaprio). Since its origin the East Flanders Prospective Survey has been partly supported by grants from the Fund of Scientific Research, Flanders and Twins, a non-profit Association for Scientific Research in Multiple Births (Belgium). Anthropometric measurements of the Hungarian twins were supported by Medexpert Ltd., Budapest, Hungary. The Murcia Twin Registry is supported by Fundación Séneca, Regional Agency for Science and Technology, Murcia, Spain (08633/PHCS/08, 15302/PHCS/10 & 19479/PI/14) and Ministry of Science and Innovation, Spain (PSI2009-11560 & PSI2014-56680-R). K Silventoinen is supported by Osaka University's International Joint Research Promotion Program. We acknowledge The Swedish Twin Registry for access to data. The Swedish Twin Registry is managed by Karolinska Institutet and receives funding through the Swedish Research Council under the grant no 2017-00641. This research was facilitated through access to Twins Research Australia, a national resource supported by a Centre of Research Excellence Grant (ID: 1079102), from the National Health and Medical Research Council. California Twin Program was supported by The California Tobacco-Related Disease Research Program (7RT-0134H, 8RT-0107H, 6RT-0354H) and the National Institutes of Health (1R01ESO15150-01). The Carolina African American Twin Study of Aging (CAATSA) was funded by a grant from the National Institute on Aging (grant 1RO1-AG13662-01A2) to K. E. Whitfield. Colorado Twin Registry is funded by NIDA funded center grant DA011015, & Longititudinal Twin Study HD10333; Author Huibregtse is supported by 5T32DA017637 and 5T32AG052371. Washington State Twin Registry (formerly the University of Washington Twin Registry) was supported in part by grant NIH RC2 HL103416 (D. Buchwald, PI). Vietnam Era Twin Study of Aging was supported by National Institute of Health grants NIA R01 AG018384, R01 AG018386, R01 AG022381, and R01 AG022982, and, in part, with resources of the VA San Diego Center of Excellence for Stress and Mental Health. The Cooperative Studies Program of the Office of Research & Development of the United States Department of Veterans Affairs has provided financial support for the development and maintenance of the Vietnam Era Twin (VET) Registry. The content of this manuscript is solely the responsibility of the authors and does not necessarily represent the official views of the NIA/NIH, or the VA. The NAS-NRC Twin Registry acknowledges financial support from the National Institutes of Health grant number R21 AG039572. Korean Twin-Family Register was supported by the Global Research Network Program of the National Research Foundation (NRF 2011-220-E00006). Osaka University Aged Twin Registry is supported by grants from JSPS KAKENHI JP (23593419, 24792601, 26671010, 24590695, 26293128, 16K15385, 16K15978, 16K15989, 16H03261).
Author information
Authors and Affiliations
Contributions
K.S., J.K., D.I.B., T.S. and Y.Y. planned the study design of the CODATwins project. J.H., J.T., A.B., C.K., L.C.F., V.O., P.F., A.H., T.M., T.L.N., K.W., R.C., B.H., C.D., R.V., R.L., J.K., A.T., D.T., E.M., L.N., V.T., J.S., H.P., J.L., S.L., H.M., J.L.S., R.K., M.M., S.P., J.O., J.S.R., L.C.C., M.G., D.A.B., D.I.B., G.W., C.B., M.B., J.H., I.B., T.N., R.T., F.I., C.H., N.S., M.W., Q.T., Z.P., D.Z., N.M., S.M., G.M., S.S., M.H., F.Ri., A.S., P.M., N.P., A.D.A., F.R., P.T., C.F., W.K., M.L., G.D. and D.B. collected the data. K.S. and A.J. were in charge of data management. K.S. wrote the first draft of the manuscript and conducted the analyses. All authors have commented the manuscript and read and approved the final version of manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Silventoinen, K., Jelenkovic, A., Sund, R. et al. Genetic and environmental variation in educational attainment: an individual-based analysis of 28 twin cohorts. Sci Rep 10, 12681 (2020). https://doi.org/10.1038/s41598-020-69526-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-69526-6
This article is cited by
-
Parental education and children’s depression, anxiety, and ADHD traits, a within-family study in MoBa
npj Science of Learning (2024)
-
More than nature and nurture, indirect genetic effects on children’s academic achievement are consequences of dynastic social processes
Nature Human Behaviour (2024)
-
How Can Genetically Informative Research Contribute to Life Course Research?
KZfSS Kölner Zeitschrift für Soziologie und Sozialpsychologie (2024)
-
Paternal intelligence affects school grades in children with and without ADHD - a register-based study
European Child & Adolescent Psychiatry (2024)
-
Using DNA to Predict Education: a Meta-analytic Review
Educational Psychology Review (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
