
Abstract
Coffee, after petroleum, is the most valuable commodity globally in terms of total value (harvest to coffee cup). Here, our bioeconomic analysis considers the multitude of factors that influence coffee production. The system model used in the analysis incorporates realistic field models based on considerable new field data and models for coffee plant growth and development, the coffee/coffee berry borer (CBB) dynamics in response to coffee berry production and the role of the CBB parasitoids and their interactions in control of CBB. Cultural control of CBB by harvesting, cleanup of abscised fruits, and chemical sprays previously considered are reexamined here to include biopesticides for control of CBB such as entomopathogenic fungi (Beauveria bassiana, Metarhizium anisopliae) and entomopathogenic nematodes (Steinernema sp., Heterorhabditis). The bioeconomic analysis estimates the potential of each control tactic singly and in combination for control of CBB. The analysis explains why frequent intensive harvesting of coffee is by far the most effective and economically viable control practice for reducing CBB infestations in Colombia and Brazil.
Similar content being viewed by others

Introduction
The coffee trade is, after petroleum, the most valuable commodity globally in terms of total value (harvest to coffee cup)1,2, and yet it faces severe issues with pests3. The coffee sector had a retail market value of USD 83 billion, providing jobs for 125 million people on 12.5 million farms worldwide, mostly smallholder farms including 22 Low Human Development Countries (LHDCs). In 2017, 70 per cent of total coffee production worth USD 19 billion was exported4. The coffee berry borer (CBB) Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) is the most important pest of coffee worldwide5,6 causing immense economic losses in the main production areas of Central and South America, Indonesia, South East Asia 7,8, Hawaii9,10, and Puerto Rico11 where infestation levels are variable but can reach up to 95%12.
Like many ambrosia beetles that form galleries for the development of progeny13, mated adult CBB females bore into coffee berries to form galleries. CBB is not effectively controlled by the action of parasitoids14, the establishment of which in new world coffee has proven difficult15,16,17,18. An early multitrophic study of the coffee agroecosystem using the mechanistic physiologically based demographic modeling (PBDM) approach19,20 demonstrated that CBB parasitoids alone were not efficient in controlling CBB21. Those findings were confirmed in subsequent restudy of the coffee system using PBDMs that introduced extensive new data from Colombia on the dynamics of coffee growth and development22,23,24, the effects of solar radiation on floral bud initiation; effects of leaf water potential on breaking dormancy in flower buds; effects of low temperature on photosynthesis and defoliation; enhanced CBB biology and population dynamics including the effects of intraspecific competition, temperature and rainfall on CBB adult emergence; the impact of baited traps for CBB control; and refinements of parasitoid biology and interactions (i.e. intra and inter competition). These PBDMs provided excellent fits to the field data on coffee growth and development, on CBB dynamics, and provide a solid base for evaluating the efficacy of the four parasitoids singly and in combinations in mitigating the impact of CBB (see review in the supplemental materials). In this paper, we add to the PBDM system the following factors: (1) Conventional cultural practices using intensive harvesting, cleanup of abscised berries, and insecticides, and (2) biopesticides based on two entomopathogenic fungal species (Beaveria bassiana and Metarhizium anisopliae), and two entomopathogenic nematodes (Steinernema sp. and Heterorhabditis sp.), and (3) the interaction of all control components. The scope of the coffee system components included in the analysis are depicted in Fig. 1. Field data are time consuming and prohibitively expensive to collect and are unlikely to yield global conclusion across time and geographic space. Realistic mechanistic weather driven PBDMs used as the objective function in our bio-economic analysis are not constrained by such limitations. The control components in the bio-economic analysis are described below.

Coffee system with all the components included (modified from Rodriguez et al.24). The complete system is embedded in a climate envelope which drives all the development and interaction variables using a PBDM (see text). Diagram of the plant model (A) including the link to the CBB model (B) and harvest and cleanup as part of CBB cultural management control (C). Effect of baited traps on capturing migrant adult females (not included in this paper based on results in Rodríguez et al.23) (D). The third trophic level is represented by four CBB parasitoids; the adult eulophid parasitoid Phymastichus coffee (E) and three bethylid parasitoids and their interactions, Cephalonomia stephanoderis, Cephalonomia hyalinipennis (not included in this paper based on results in Rodríguez et al.24) and Prorops nasuta (F). Three active ingredients (a.i.) of insecticides are include in rotation, affecting the whole system (G), as well as the entomopathogenic nematodes Steinernema sp. and Heterorhabditis sp. (H) and the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana (I). The dashed line indicates information flow.
Control components
- 1.
Intensive harvesting, cleanup, and insecticide.
There are two main periods of fruit ripening in the Colombian coffee region, April–June, and September–November25, but occasional dry periods occur in between that induce the presence of ripe fruits on the plant and abscised fruit on the ground25,26. Common cultural practices for CBB control are periodic harvesting of overripe and dry fruits on the tree and cleanup of abscised fruit on the ground (hereafter termed harvest and cleanup respectively)27,28,29,30.
Insecticides are commonly used for CBB control31,32,33, causing mortality of free-flying adult females before they bore into berries. The action of insecticides decreases over 15 days from the time of application. Benavides and Arévalo27 and Benavides et al.32 recommended, that chemical control should start 90 days after flowering, when berries have a dry matter content of approximately 20%, and that it should be continued if infestation levels are higher than 2%. However, because berries are often present year around in Colombia, it is difficult to apply this criterion for initiating chemical control25,26. A further drawback is that the chemicals are hazardous to farm workers and adversely affect biodiversity conservation, particularly natural enemies resulting in outbreaks of new pests34,35 and contamination of water and soils31, but also pollinators potentially resulting in decreased fruit set36. Due to these drawbacks, it has been suggested that farmers should increase resources for field sanitation37.
- 2.
Biopesticides of fungal pathogens (Beaveria bassiana and Metarhizium anisopliae).
Sprays infused with the fungus B. bassiana have been reported as an effective control agent of CBB adults38,39,40,41,42,43,44. Using B. bassiana as a stand-alone method of control was questioned by Hollingsworth et al.45 The infection rates in the field depend on climatic conditions and strain of the pathogen, with mortality rates ranging from 17.7%39 at a concentration of 106 conidia/ml to 40.6%40 at a concentration of 109 conidia per plant, though Bustillo46 reported mortality as high as 75%. The spores must come in contact with the beetles, and hence are most infective on new adults emerging from host berries or during the initial phases of fruit penetration47. Infection by Metarhizium anisopliae has a similar etiology, with reported infection rates ranging between 22.1%40 and 43.1%48.
- 2.
Biopesticides of nematodes (Steinernema and Heterorhabditis).
Nematodes are not widely employed in commercial crops due to their high cost of production. Experimental evidence in laboratory suggests that they could be useful against CBB stages49,50,51. A field evaluation confirmed that both species can be effective in controlling CBB in abscised fruits in the soil with mortalities of as high as 80% sixty days after the application of 250,000 infective Steinernema sp. juveniles per tree. Similarly, applications of 125,000 infective Heterorhabditis sp. juveniles per tree can cause 75% mortality after 75 days52.
- 4.
The action of CBB parasitoids.
Several studies have reported the use of parasitoids for control of CBB, and their biology were well documented and modeled by Gutierrez et al.21 and Rodríguez et al.24. The bethylids Cephalonomia stephanoderis53,54,55,56,57,58, C. hyalinipennis18 and Prorops nasuta53,59,60,61 enter the CBB gallery to attack the immature stages. In contrast, the eulophid Phymastichus coffea62,63,64,65,66,67,68 parasitize female CBB adults as they are burrowing into the coffee berry. Results of Rodríguez et al.24 show that C. hyalinipennis, interacts negatively with the other betilids and provides poor control of CBB, and this argues against its introduction, and hence was not considered here. The present work explores only the performance of C. stephanoderis, P. nasuta and P. coffea.
Methods
Study sites
Geographical coordinates of the localities and weather data used in these simulation studies are listed in Table 1. The field studies for the plant and CBB were conducted for Colombia (coffee cv. Colombia) during 1990–199569 and 2009–201022,23, and during 1982–1986 for Londrina, PR, Brazil (coffee cv. Mundo Novo)23. Daily maximum and minimum temperatures (°C), relative humidity (%), precipitation (mm), and hours of sunshine for Colombia were obtained from data published in the Anuario Meterológico Cafetero70. Daily solar radiation (MJ∙m−2∙day−1) was estimated from recorded hours of sunshine using the relationship developed by Prescott71. Weather data for Londrina, Brazil were obtained from the Instituto Agronômico do Paraná.
Exploring alternatives for CBB management using the PBDM system
Cultural control
Simulated harvests (symbol H) and cleanup (CU) at three different time-intervals (T = 15 days, 30 days, and 60 days) were explored. Because the PBDM tracks the age structure of developing fruit, estimates of field harvest efficiency of fruit stages per branch reported by Baker72 were used in the model: 8.8% for unripe fruit, 67.3% for ripening fruit, 88.5% for ripe fruit and 53.4% for dry fruit, with harvest efficiency decreasing with tree age (i.e. 87% on 2-year old trees and 62% for 3- and 4-year trees). For cleanups, an efficiency of 53.4% was assumed72.
Chemical control
Mortality associated with insecticide sprays (C) occurs only to free-living adults, both CBB and parasitoids, as all immature stages and not free-living adults are inside fruits and are unaffected31,33,46. The active ingredients (ai = 1…3) of Chlorpyrifos, Fenitrothion and Phenthoate have been reported as effective for control of CBB females colonizing fruit with maximum mortalities γai = 0.93, 0.95 and 0.97, respectively33. Same mortality values were used for the parasitoid P. coffea. The realized mortality rate (\({\mu }_{ai}\)) of CBB and all parasitoids is assumed to decrease exponentially through time (t) in days from initial maximum mortality values (\(\gamma_{ai}\)) , Eq. (1):
The effect of Chlorpyrifos and Fenitrothion on the parasitoid P. nasuta was studied by Mejía et al.73 and provided estimates for \(\gamma\) of 0.96 and 0.97 respectively. These values were also used for the effect of the chemicals on free living C. stephanoderis. Data evaluating the effect of Phenthoate on the bethylids were not available, and hence the same value of \(\gamma\) for CBB (0.97) was assumed. In the simulations, a rotation scheme of the three sprays was used. The recorded 13 spray dates coinciding with simulated periods of high CBB female emergence and were used for the 5-year simulations for Colombia. Seven spray dates were used for Brazil during the 3-year study.
Entomopathogenic fungi
Logistic models assuming a logit link function74 were fit to data reported by De la Rosa et al.40 on the average proportion infection (mortality) by B. bassiana (strain Bb25), Eq. (2), and M. anisopliae (strain Ma4) Eq. (3), on cumulative daily degree days after spraying (i.e. dda) and daily relative humidity (RH). Daily degree days were computed with a nonlinear model75 using maximum and minimum temperatures and lower and upper thermal thresholds of 8 °C and 35 °C respectively for B. bassiana (BB)76, and 11 °C and 35 °C for M. anisopliae (MA)77. Several significant digits are given so the results can be accurately reproduced.
\({\mu}_{BB}\) and \({\mu }_{MA}\) are the proportion infection (i.e. mortality rate) of CBB adult females seeking or starting to colonize new fruits. The pathogens also affect the parasitoids, but data are available only for bethylid C. stephanoderis adults, and hence the same function was assumed for P. nasuta. Daily mortality rates for B. bassiana on parasitoids P. nasuta73, C. stephanoderis78 and P. coffea79 were estimated as a linear function of pathogen (dda). Infection and mortality of all developmental stages of P. coffea attacking CBB females was assumed79.
Entomopathogenic nematodes
Infection by nematodes is restricted to CBB in attacked berries and in abscised berries on the ground. Logit functions of infection with an application of 250,000 infective juveniles of Steinernema sp. (\({{\mu }}_{S}\)), Eq. (4) and 125,000 infective juveniles of Heterorhabditis sp. (\({{\mu }}_{H}\)), Eq. (5), were fit to data reported by Lara et al.52 as functions of cumulative daily degree days after spraying (dda) above 8 °C for Steinernema and 11 °C for Heterorhabditis starting from the beginning of the infection, Eqs. (4) and (5).
Humidity and ultraviolet radiation may also affect nematode survival and efficacy80, but data were not available on these effects and hence were not included in the analysis.
Data analysis
Combination and interactions of control components
The models for the different control factors are modular, and individual factors (independent variables) could be included in simulation runs using Boolean dummy variables (include = 1, exclude = 0), while the simulated cumulative number of CBB infested berries year−1 (I) was the dependent variable.
Two studies were done. In the first study, the ten combinations of cultural controls (H, CU) × time interval treatments widely used by coffee farmers81 (Table 2) and the use of chemical insecticides were analyzed. In the second study, 2,560 combinations of control factors were included: cultural control and times, insecticides, the four entomopathogen biopesticides and three parasitoids (C. stephanoderis, P. nasuta and P. coffea).
Negative binomial regression models assuming a log link function were used to summarize the simulation results, Eq. (\(6)\). Specifically, loge cumulative number of CBB infested berries⋅year-1 (loge I) was regressed on presence-absence values of the independent control variables xi.
The loge-linear model, Eq. (6), accounts for over dispersion and skewness and satisfies the assumptions of the parametric analysis82. The final model was selected using Akaike’s information criteria83 retaining only significant independent variables and interactions (p < 0.05). One model was fit to the combined data for the four Colombia localities and another for the single Brazilian locality.
To estimate the magnitude and direction of the impact of a management variable on CBB infestations, the derivative of Eq. (6) was with respects to xi , Eq. (7).
This yields the loge rate of change of infested berries given the action of xi and the average effects of the other independent variables. Taking the antilog of Eq. (7) we get the infestation rate as a proportion after the action of xi.
Results
Analysis of cultural and chemical controls
Colombian study
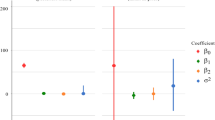
Results of the multiple regression model for cultural controls are summarized in Table 3. If H = harvest, CU = cleanup, C = chemical control and T = time is in days between cultural controls, then the marginal loge contributions of each factor, Eq. (7), in reducing I are: H(\({{\partial \log_{e} I} \mathord{\left/ {\vphantom {{\partial \log_{e} I} {\partial H}}} \right. \kern-\nulldelimiterspace} {\partial H}} = - 2.0942\)) > CU(\(= - 0.1641\)) > C(\(= - 0.1064\)), and show that harvesting has the greatest effect in reducing infestation levels. Note that, as time between harvests increases, infestation levels increase, as suggested by the positive marginal contributions for T (\(= 0.1929\)). The same notation (e.g., H(value)) will be used in the other sections for other control factors.
The antilog of marginal contribution of each factor (left super script A) is the proportion of fruit infested by CBB after the action of the factor given the average effect of the other independent variables: \(^{A} H = e^{( - 2.0842)} = 0.1231\), \(^{A} CU = e^{( - 0.1641)} = 0.8486\) and \(^{A} C = e^{( - 0.1064)} = 0.899\), which again highlights the importance of harvesting over cleanup and chemical control, suggesting that the impact of AH is 6.9 fold higher than CU (i.e., ACU/AH) and 7.3 higher than AC.
Brazilian study
In Brazil, periodic harvest (H) and time interval (T) were the only significant factors in the regression model (Table 4). The marginal loge contribution of harvest in reducing CBB infestation is H (− 2.0343), with the average proportion of berries infested by CBB being \(^{A} H = e^{( - 2.0343)} = 0.1307\). As in Colombia, the effect of T is positive, indicating that infestation levels increase as the time between treatments increases. Despite the very different growth forms of coffee in Brazil, the values are similar to those estimated for Colombia.
Analysis of all factors and interactions
Colombian study
The regression model of loge I on all independent variables and their combinations (Table 5) shows that all control variables singly reduce infestation levels (negative sign of the coefficient). On the other hand, positive signs for most interaction terms (except HˑPcˑBb) suggest the interactions increase infestation levels due to antagonistic effects among control components. Again, the positive sign of the regression coefficient for time interval T indicates that infestation levels increase with increasing time between implementation of cultural control practices. Harvest and cleanup reduce infestation to 13.48% 15 day intervals, but this percentage increases to 18.86% when the interval increases to 30 days, and rises markedly to 37.49% when 60 days.
The average marginal loge contributions of the various factors in decreasing CBB infestation (I) are: H(− 1.0454) > CU(− 0.6228) > Het(− 0.1388) > Stei(− 0.1215) > Bb(− 0.1016) > Pc (− 0.07564) > C(− 0.05907) > Ma(− 0.04196) > Cs(− 0.02566) > Pn(− 0.01148).
These results expressed as average proportion berry infestation rates (antilog) given the average effect of the other independent variables are:
AH = \({e}^{-1.0454}=0.3515\), ACU \(=0.5364\), AHet \(=0.8703\), AStei \(=0.8856\), ABb \(=0.9034\), APc \(=0.9271\), AC \(=0.9426\), AMa \(=0.9589\), ACs \(=0.9746\) and APn \(=0.9885\)). These results reinforce the notion that frequent harvesting (AH) is the most effective tactic for reducing loge I with an effect 1.5-fold > CU, 2.47-fold > Het, 2.51-fold > Stei, 2.57-fold > Bb, 2.63-fold > Pc, 2.68-fold > C, 2.72-fold > Ma, 2.77-fold > Cs and 2.81-fold > Pn.
Brazilian study
In Brazil, only a few single variables were significant contributors to the reduction of loge I (Table 6), and only the interactions PcˑC and HˑT had significant positive effects. Contrary to that obtained for Colombia, the interaction BbˑH has a negative effect on infestation levels. As in Colombia, the time interval (T) between cultural control practices was significant positive. The estimated infestation is 3.59% when the harvest and cleanup interval is 15 days, 9.69% for 30 days and 70.41% for 60 days.
Average marginal loge contribution to the reduction of I are as follows: H (− 2.1880) > Bb (− 0.1985) > Ma (− 0.1250) > Het (− 0.07821) > Stei (− 0.07379) > Pc (− 0.06540) > Cs (− 0.03636) > C(− 0.03140). The results expressed as average proportion of berries infested are: AH(\({e}^{-2.1880}=0.1121\)), ABb(\({e}^{-0.1985}=0.8200\)), AMa(\({e}^{-0.1250}=0.8825\)), AHet(\(=0.9248\)), AStei(\(=0.9288\)), APc(\(=0.9367\)), ACs(\(=0.9643\)) and AC( \(=0.9690\)).
The order of impact of the control practices differ in some cases from the results for Colombia, although harvest (AH) is again the most effective control method to reduce infestations, with its affect being 7.3-fold > Bb, 7.9-fold > Ma, 8.2-fold > Het, 8.3-fold > Stei, 8.3-fold > Pc, 8.6-fold > Cs and 8.4-fold > C.
Discussion
Bioeconomics is the study of the economics of renewable resource acquisition and allocation applicable to all trophic levels. In human economies, harvesting of renewable resources occur via the economic system84. Econometric marginal analysis is best done with extensive field data85, but such data may be difficult to collect and is prohibitively expensive (e.g., coffee). However, simulation data generated by a well parameterized, field-validated mechanistic models can provide a highly suitable alternative because the results can be compared to limited field data86,87. Our PBDM system developed to simulate the growth and development of coffee, the dynamics and infestation levels of coffee berry borer (CBB) and the action of four parasitoids of CBB is based on extensive data21,22,23,24 and provide a very suitable platform for including the effects of cultural practices such as harvest (H), cleanup of abscised berries (CU) and the time intervals between these activities, and the effects of sprays of insecticide and of biopesticides of two fungal pathogens and two nematode parasites.
Intensive international efforts to achieve biological control of CBB have failed. Our simulation results for Colombia explain the average reduction of < 15% by parasitoids, entomopathogens, and chemical control. Additionally, antagonistic effects among these control tactics were found. For example, harvesting and cleanup affected the action of CBB parasitoids, P. nasuta (Pn) and P. coffea (Pc) increasing CBB infestation levels. Specifically, positive coefficients for the interactions PnˑH, PcˑH and PcˑCU indicate detrimental effects to parasitoid efficacy of harvesting and cleanup because parasitoid life stages are also removed from the system, resulting in lower future CBB parasitization rates. The incompatibility between cultural control and parasitoids was also found by Gutiérrez et al.21 and Aristizábal et al.88.
Similar antagonistic effects were found for harvests and cleanup with sprays of pathogenic fungi (B. bassiana, Bb) and cultural practices as indicated by the positive sign interactions of BbˑH and BbˑCU. Bustillo47 reports that sustained efficacy of the pathogen B. bassiana in the field is strongly associated with the production of spores from field infected CBB, but harvest and cleanup remove these inoculum sources. Another significant antagonistic effect identified for Colombia was the interaction of the eulophid parasitoid P. coffea and B. bassiana (i.e. PcˑBb). In laboratory studies, Castillo et al.79 found that exposure to B. bassiana caused mortality rates of 100% in P. coffea immature stages and a reduction of 22% in adult longevity which reduces parasitoid efficacy. Chemical control (C) also affects the efficacy of P. coffea as indicated by the positive interaction PcˑC. This occurs because unlike the bethylid parasitoids that enter the berry, P. coffea female are entirely free living and attacks CBB females initiating penetration of coffee berries making them susceptible to insecticides89.
Despite some detrimental effects on biological control agents, periodic harvest of fruit and clean up were found to be the major control practice reducing CBB infestation levels (I) in both Colombia and Brazil, with the efficacy of the practice decreasing as the time (T) between harvests (H) and cleanup (CU) increased from 15 to 60 days. The analysis for Colombia suggests that cleanup is the second most important control strategy for reducing the level of infestations. These simulation result agrees with Johnson et al.90, who found that ground and tree raisins (dry overripe fruit) left after harvest, could be the main CBB reservoir in the inter-crop season in Hawaii. The results for these cultural practices also agree with field studies of Duque and Cháves91 who found that > 94% of Colombian farmers participating in a survey considered cultural control to be the most important method for reducing CBB populations. Bustillo et al.92 found that periodic harvesting reduced CBB populations up to 80%, with Benavides et al.32 and Aristizábal et al.29 in Colombia and Aristizábal in Hawaii30, reporting that periodic harvests at 15 day was the main method for reducing CBB populations, and for generating higher yield and income. Unfortunately, producers have a checkered record of implementation cultural control tactics, as Aristizábal et al.93 found that only 45% were applying periodic harvest according to the criteria proposed by Bustillo et al.92.
Gutierrez et al.21 found for Brazil that harvesting and cleanup (only twice a year) had little impact on control because at harvest most berries were infested, the females inside fruits were near the end of their reproductive life, and most adult progeny had emerged. However, as in Colombia, harvesting was the most important factor reducing CBB infestation.
In summary, harvesting and cleanup at 15-day intervals is the only control tactic that significantly reduces CBB infestation level in Colombia and Brazil. Aristizabal et al.30 analyzing the cost of harvesting and cleanups (“sanitation picks") in Hawaii, remarked that while initially the cost appears to be high, in the final analysis, sanitation pays the cost of labor and processing, while reducing the source of the pest. That study for Hawaii and the study of Benavides et al.32 for Colombia, shows that harvesting and cleanup can be economically feasible. However, it may not be economic in Londrina, Brazil which is at the southern climatic limits of coffee production, with short dry periods followed by short periods of rain throughout the year, resulting in the production of susceptible berries over a longer period than in Colombia. This fruiting phenology has a strong impact on the dynamics of the system and on CBB control as shown by our PBDM results22,23.
The socio-economic conditions differ in various coffee growing regions, the fluctuation of prices in the international market can vary widely (including for premium quality coffee), and infestation levels have an important impact on coffee yield and price94. Hence, in economic analyses, control tactics must enter not only as cost, but also as price enhancing attributes. For example, effective CBB control based on sustainable periodic harvesting could be an important element in promoting and positioning select coffees on the international markets as unique, organic, and highest quality. To this end, an in-farm mixture of shade grown, and sun grown coffees using organic cultural practices to control CBB has been proposed as a sustainable option for coffee production on small to medium properties95.
In conclusion, our model is a realistic virtual crop system that provides a very useful general tool for investigating aspects not readily amenable to field experimentation and has the capacity to integrate more layers such as a socio-economic one. This tool can also be used to examine new technological opportunities prior to their wide adoption. For example, CBB control may be affected by disrupting the symbiotic bacteria in CBB’s microbiome responsible for caffeine breakdown96. Another tactic is the development of attractants that are more competitive with the activeness of coffee berries96,97; a tactic that could be especially important because coffee flowering phenology varies widely throughout the world in response to regional climate patterns that influences the phenology and dynamics of CBB infestation and the success of progeny development.
Climate change, including climate variability, must be considered as this may change extant regional dynamics of both coffee and CBB, and their interactions. Increased temperature may generate conditions favorable (or unfavorable) for coffee and CBB allowing range extensions to new areas, and changes in CBB damage levels in its current geographical distribution. Increased dry “El Niño” climatic events in some countries could increase CBB populations, while “La Niña” events with prolonged wet seasons would limit CBB populations. The effects of such phenomena differ across geographical region7, and the coffee/coffee berry borer system model provides a framework for analyzing the potential effect of variation in weather, climates and of climate change on coffee yield, and the dynamics of CBB across diverse bio-geographical zones23.
As an aside, the high pest status of this species in monocultures is a consequence of an evolutionary background, similar to what have been observed in other systems (e.g., between fruiting in sylvan cotton and cotton weevil, Anthonomus grandis Boh.20). From the prospective of the ecological theory, the large female bias appears to have had high adaptive value in the African tropical forest where it evolved so that large numbers of the small females with low searching rates could find scattered patches of suitable age berries23. This adaptation would appear to occur at the expense of reduction in genetic variability caused by sib-mating and reported pseudo-arrhenotoky98,99,100,101.
As a final note, our C. arabica PBDM can easily be modified to include other species of coffee (e.g., C. robusta), and has transferability enabling its use in a bio-economic analysis on larger, albeit global scale, and in the face of climate change.
References
Talbot, J. M. Grounds for Agreement. The Political Economy of the Coffee Commodity Chain 1–250 (Rowman & Little field Publishers Inc, Washington, DC, 2004).
Igami, M. Market power in international commodity trade: The case of coffee. J. Ind. Econ. 63, 225–248 (2015).
Waller, J. M., Bigger, M. & Hillocks, R. J. Coffee Pests, Diseases and their Management 1–400 (CABI, Wallingford, 2007).
Sustainable Commodities Marketplace Series. Global Market Report: Coffee. International Institute for Sustainable Development (IISD). https://www.iisd.org/sites/default/files/publications/ssi-global-market-report-coffee.pdf (2019).
Vega, F. E., Infante, F., Castillo, A. & Jaramillo, J. The coffee berry borer, Hypothenemus hampei (Ferrari) (Coleoptera:Curculionidae): A short review, with recent findings and future research directions. Terr. Arthropod. Rev. 2, 129–147 (2009).
Vega, F. E., Infante, F. & Johnson, A. J. In The Genus Hypothenemus, Biology and Ecology of Native and Invasive Species with Emphasis on H. hampei, The Coffee Berry Borer in Bark Beetles (eds Vega, F. E. & Hofstetter, R. W.) 427–494 (Elsevier, London, 2015).
International Coffee Organization. IPM of Coffee Berry Borer (CFC/ICO/02). Impact Assessment. https://www.ico.org/project_pdfs/CBB_ICO02.pdf (2009).
CABI. Stopping the coffee berry borer in its tracks. Development projects. https://www.cabi.org/uploads/projectsdb/documents/2734/Coffee-berry-borer_HR.pdf (2012).
Burbano, E., Wright, M., Bright, D. E. & Vega, F. E. New record for the coffee berry borer, Hypothenemus hampei, in Hawaii. J. Insect. Sci. 11, 117 (2011).
State of Hawaii Department of Agriculture. Coffee berry borer Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae: Scolytinae). Hawaii Department of Agriculture. New Pest Advisory 10–01. https://hdoa.hawaii.gov/pi/files/2013/01/Hypothenemus-hampei-NPA-MASTER.pdf (2014).
NAPPO. North American Plant Protection Organization. Detection of the coffee berry borer, Hypothenemus hampei, in Puerto Rico. Official Pest Alerts. https://www.pestalerts.org/es/official-pest-report/detecciones-de-la-broca-del-caf-hypothenemus-hampei-en-puerto-rico-estados (2007).
Mariño, Y. A. et al. The coffee berry borer (Coleoptera: Curculionidae) in Puerto Rico: Distribution, infestation, and population per fruit. J. Insect Sci. 17(2), 58 (2017).
Wood, S. L. Bark and Ambrosia Beetles of South America. 1–900 (Monte L. Bean Sci. Mus., Provo, Utah, 2007).
Damon, A. A review of the biology and control of the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae). Bull. Entomol. Res. 90, 453–465 (2000).
Batchelor, T. P., Hardy, I. C. W. & Barrera, J. F. Interactions among bethylid parasitoid species attacking the coffee berry borer, Hypothenemus hampei (Coleoptera:Scolytidae). Biol. Control 36, 106–118 (2006).
Benassi, V.L.R.M. Biologia em diferentes temperaturas e ocorrência de Prorops nasuta Wat. e Cephalonomia stephanoderis Betr. (Hymenoptera:Bethylidae) parasitando Hypothenemus hampei (Ferr.) (Coleoptera:Scolytidae). Biblioteca digital Brasileira de teses e dissertações, https://bdtd.ibict.br/vufind/Record/UNSP_921d283e6a2f9fc4271784147e404a27 (2007).
Maldonado, C. E. & Benavides, P. Evaluación del establecimiento de Cephalonomia stephanoderis y Prorops nasuta, controladores de Hypothenemus hampei, en Colombia. Revista cenicafé (Colombia) 58, 333–339 (2007).
Pérez-Lachaud, G., Batchelor, T. P. & Hardy, I. C. W. Wasp eat wasp: Facultative hyperparasitism and intra-guild predation by bethylid wasps. Biol. Control 30, 149–155 (2004).
Gutierrez, A. P. & Baumgärtner, J. U. Multitrophic level models of predator-prey energetics: II: A realistic model of plant-herbivore-parasitoid-predator interactions. Can. Entomol. 116, 933–949 (1984).
Gutierrez, A. P. Applied Population Ecology: A Supply-Demand Approach 1–300 (Wiley, Hoboken, 1996).
Gutierrez, A. P., Villacorta, A., Cure, J. R. & Ellis, C. K. Tritrophic analysis of the coffee (Coffea arabica)-coffee berry borer Hypothenemus hampei (Ferrari)-parasitoid system. Anais da sociedade entomológica do brasil 27(3), 357–385 (1998).
Rodríguez, D., Cure, J. R., Cotes, J. M., Gutierrez, A. P. & Cantor, F. A coffee agroecosystem model. I. Growth and development of the coffee plant. Ecol. Model. 222, 3626–3639 (2011).
Rodríguez, D., Cure, J. R., Cotes, J. M., Gutierrez, A. P. & Cantor, F. A coffee agroecosystem model. II. Dynamics of coffee berry borer. Ecol. Model. 248, 203–214 (2013).
Rodríguez, D., Cure, J. R., Cotes, J. M. & Gutierrez, A. P. A coffee agroecosystem model. III. Parasitoids of the coffee berry borer (Hypothenemus hampei). Ecol. Model. 363, 96–110 (2017).
Arcila, J., Jaramillo, A., Baldión, V. & La Bustillo, A. E. floración del cafeto y su relación con el control de la broca. Avances Técnicos Cenicafé 193, 6 (1993).
Vélez, B. E., Jaramillo, A., Cháves, B. & Franco, M. Distribución de la floración y la cosecha de café en tres altitudes. Avances Técnicos Cenicafé 272, 4 (2000).
Benavides, P. & Arévalo, H. Manejo integrado: una estrategia para el control de la broca del café en Colombia. Revista Cenicafé (Colombia) 53(1), 39–48 (2002).
Benavides, P., Bustillo, A. E., Cárdenas, M. R. & Montoya, E. C. Análisis biológico y económico del manejo integrado de la broca del café en Colombia. Revista Cenicafé (Colombia) 54(1), 5–23 (2003).
Aristizábal, L. F., Jiménez, M., Bustillo, A. E. & Arthurs, S. P. Monitoring cultural control practices for coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) management in a small coffee farm in Colombia. Fla. Entomol. 94(3), 685–687 (2011).
Aristizábal, L. F. Frequent and efficient harvesting practices to reduce coffee berry borer populations. CBB Notes Number 05, Kailua-Kona, Hawaii https://www.researchgate.net/publication/340502340_CBB_Notes_Frequent_and_Efficient_Harvesting_Practices_to_Reduce_Coffee_Berry_Borer_Populations (2020).
Villalba, D. A., Bustillo, A. E. & Cháves, B. Evaluación de insecticidas para el control de la broca del café en Colombia. Revista Cenicafé (Colombia) 46(3), 152–163 (1995).
Benavides, P., Bustillo, A. E., Montoya, E. C., Cárdenas, M. R. & Mejía, C. G. Participación del control cultural, químico y biológico en el manejo de la broca del café. Rev. Colomb. Entomol. 28, 161–165 (2002).
Tabares, J. E., Villalba, D. A., Bustillo, A. E. & Vallejo, L. F. Eficacia de insecticidas para el control de la broca del café usando diferentes equipos de aspersión. Revista Cenicafé (Colombia) 59(3), 227–237 (2008).
Beers, E. H. et al. Nontarget effect of orchard pesticides on natural enemies: Lessons from the field and the laboratory. Biol. Control 102, 44–52 (2016).
Bueno, A. F., Carvalho, G. A., Santos, A. C., Sosa-Gómez, D. R. & Silva, D. M. Pesticide selectivity to natural enemies: Challenges and constrains for research and field recommendation. Ciência Rural (Santa María, Brasil) https://doi.org/10.1590/0103-8478cr20160829 (2017).
Klein, A. M., Steffan-Dewenter, I. & Tscharntke, T. Bee pollination and fruit set of Coffea arabica and C. canephora (Rubiaceae). Am. J. Bot. 90, 153–157 (2003).
Woodill, A. J., Nakamoto, S. T., Kawabata, A. M. & Leung, P. S. To spray or not to spray: A decision analysis of coffee berry borer in Hawaii. Insects 8(4), 116 (2017).
González, M. T., Posada, F. J. & Bustillo, A. E. Bioensayo para evaluar la patogenicidad de Beauveria bassiana (Bals.) Vuill. sobre la broca del café, Hypothenemus hampei (Ferrari). Rev. Colomb. Entomol. 19, 123–130 (1993).
Tobar, S. P., Vélez, P. E. & Montoya, E. C. Evaluación en campo de un aislamiento del hongo Beauveria bassiana seleccionado por resistencia a la luz ultravioleta. Revista Cenicafé (Colombia) 50(3), 195–204 (1999).
De la Rosa, W., Alatorre, R. R., Barrera, J. F. & Toriello, C. Effect of Beauveria bassiana and Metarhizium anisopliae (Deuteromycetes) upon the Coffee Berry Borer (Coleoptera: Scolytidae) under field conditions. J. Econ. Entomol. 93, 1409–1413 (2000).
Posada, F. J., Osorio, E. & Velásquez, E. T. Evaluación de la patogenicidad de Beauveria bassiana sobre la broca del café empleando el método de aspersión foliar. Rev. Colomb. Entomol. 28, 139–144 (2002).
Cárdenas, A. B., Villalba, D. A., Bustillo, A. E., Montoya, E. C. & Góngora, C. E. Eficacia de mezclas de cepas del hongo Beauveria bassiana en el control de la broca del café. Revista Cenicafé (Colombia) 58, 293–303 (2007).
Aristizábal, L. F. et al. Integrated pest management of coffee berry borer in Hawaii and Puerto Rico: Current status and prospects. Insects 8, 123 (2017).
Greco, B. E., Wright, M. G., Burgueño, J. & Jaronski, S. TEfficacy of Beauveria bassiana applications on coffee berry borer across an elevation gradient in Hawaii. Biocontrol Sci. Technol. 28, 995–1013 (2018).
Hollingsworth, R. G. et al. Incorporating Beauveria bassiana into an integrated pest management plan for coffee berry borer in Hawaii. Front. Sustain. Food Syst. 4, 22 (2020).
Bustillo, A. E. Una revisión sobre la broca del café, Hypothenemus hampei, en Colombia. Rev. Colomb. Entomol. 32(2), 101–116 (2006).
Bustillo, A. E. El manejo de cafetales y su relación con el control de la broca del café en Colombia 2nd edn, 1–40 (Cenicafé, Chinchiná, 2007).
Bernal, M. G., Bustillo, A. E. & Posada, F. J. Virulencia de aislamientos de Metarhizyum anisopliae y su eficacia en campo sobre Hypothenemus hampei. Rev. Colomb. Entomol. 20(4), 225–228 (1994).
Molina, J. P. A. & López, N. J. C. Efeito da aplicação de nematoides entomopatogênicos sobre frutos infestados com broca-do-café, Hypothenemus hampei (Coleoptera:Scolytidae). Nematol. Bras. (Brazil) 33(2), 115–122 (2009).
Benavides-Machado, P., Quintero, J. C. & López, J. C. Evaluación en el laboratorio de nematodos entomopatógenos nativos para el control de la broca del café. Cenicafé (Colombia) 61(2), 119–131 (2010).
Guide, B. A. et al. Selection of entomopathogenic nematodes and evaluation of their compatibility with cyantraniliprole for the control of Hypothenemus hampei. Semina: Ciências Agrárias, Londrina 39(4), 1489–1502. https://doi.org/10.5433/1679-0359.2018v39n4p1489 (2018).
Lara, J. C., Lopez, J. N. & Bustillo, A. E. Efecto de entomonemátodos sobre poblaciones de la broca del café, Hypothenems hampei (Coleoptera, Scolytidae) en frutos en el suelo. Rev. Colomb. Entomol. 30(2), 179–185 (2004).
Abraham, Y. J., Moore, D. & Goodwin, G. Rearing and aspects of biology of Cephalonomia stephanoderis and Prorops nasuta (Hymenoptera:Bethylidae) parasitoid of the coffee berry borer Hypothenemus hampei (Coleoptera: scolytidae). Bull. Entomol. Res. 80, 121–128 (1990).
Barrera, J. F. Dynamique des populations du scolyte des fruits du caféier, Hypothenemus hampei (Coleoptera: Scolytidae), et lutte biologique avec le parasitoide Cephalonomia stephanoderis (Hymenoptera: Bethylidae), au Chiapas, Mexique. Ph.D. Thesis. Université Paul Sabatier, Toulouse III. France. https://agritrop.cirad.fr/312449/ (1994).
Aristizábal, L. F., Baker, P. S., Orozco, H. J. & Orozco, G. L. Determinación de las horas del día convenientes para la liberación del parasitoide Cephalonomia stephanoderis (Betrem) (Hymenoptera: Bethylidae). Rev. Colomb. Entomol. 2, 99–104 (1996).
Aristizábal, L. F., Baker, P. S., Orozco, S. J. & Chaves, B. Parasitismo de Cephalonomia stephanoderis Betrem sobre una población de Hypothenemus hampei (Ferrari) con niveles bajos de infestación en campo. Rev. Colomb. Entomol. 23, 157–164 (1997).
Aristizábal, L. F., Bustillo, A. E., Baker, P. F., Orozco, H. J. & Chaves, C. B. Efecto depredador del parasitoide Cephalonomia stephanoderis (Hymenoptera: bethylidae) sobre los estados inmaduros de Hypothenemus hampei (Coleoptera: scolytidae) en condiciones de campo. Rev. Colomb. Entomol. 24, 35–41 (1998).
Damon, A. & Valle, J. Comparison of two release techniques for the use of Cephalonomia stephanoderis (Hymenoptera: Bethylidae), to control the coffee berry borer Hypothenemus hampei (Coleoptera: Scolytidae) in Soconusco, southeastern Mexico. Biol. Control 24, 117–127 (2002).
Hempel, A. Combate à broca do café por meio da Vespa de Uganda. Bol. Agric. Zoot. Vet. 6(9), 551–555 (1933).
Hempel, A. A Prorops nasuta Waterston no Brasil. Arquivos do Instituto Biológico de São Paulo (Brasil) 5, 197–212 (1934).
Infante, F. Biological Control of Hypothenemus hampei(Coleoptera:Scolytidae) in Mexico, Using the Parasitoid Prorops nasuta (Hymenoptera:bethylidae) Dissertation, Imperial College, University of London (1998).
Infante, F. et al. Cría de Phymastichus coffea, parasitoide de la broca del café y algunas notas sobre su historia de vida. Southw. Entomol. 19, 313–315 (1994).
Aristizábal, L. F., Salazar, H. M., Mejía, C. G. & Bustillo, A. E. Introducción y evaluación de Phymastichus coffea (Hymenoptera: eulophidae) en fincas de pequeños caficultores, a través de investigación participativa. Rev. Colomb. Entomol. 30, 219–234 (2004).
Castillo, A., Infante, F., Vera-Graziano, J. & Trujillo, J. Host-discrimination by Phymastichus coffea, a parasitoid of the coffee berry borer (Coleoptera: Scolytidae). Biocontrol 49, 655–663 (2004).
Castillo, A. et al. Laboratory parasitism by Phymastichus coffea (Hymenoptera: Eulophidae) upon non-target bark beetles associated with coffee plantations. Fla. Entomol. 87, 274–277 (2004).
Castillo, A. Análisis Post-introductorio Del Parasitoide Africano Phymastichus coffea LaSalle (Hymenoptera: Eulophidae) a México. Dissertation 1–150 (Colegio de Postgraduados, Montecillo, Estado de México, 2005).
Jaramillo, J., Bustillo, A. E., Montoya, E. C. & Borgemeister, C. Biological control of the coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae) by Phymastichus coffea (Hymenoptera: Eulophidae) in Colombia. Bull. Entomol. Res 95, 1–6 (2005).
Jaramillo, J., Borgemeister, C. & Setamou, M. Field superparasitism by Phymastichus coffea, a parasitoid of adult coffee berry borer, Hypothenemus hampei. Entomol. Exp. Appl. 119, 231–237 (2006).
Riaño, N., Arcila, J., Jaramillo, A. & Chaves, B. Acumulación de materia seca y extracción de nutrimentos por Coffea arabica L. cv. Colombia en tres localidades de la zona cafetera central. Revista Cenicafé (Colombia) 55, 265–276 (2004).
CENICAFE Anuario Meteorológico Cafetero 1990–1995. Chinchiná, Colombia, https://books.google.com.co/books?id=6QNdAAAAMAAJ&hl=es&source=gbs_book_other_versions (Centro Nacional de Investigaciones del Café “Pedro Uribe Mejía”, 1995).
Prescott, J. A. Evaporation from water Surface in relation to solar radiation. Trans. R. Soc. South. Aust. 64, 114–118 (1940).
Baker, P.S. The Coffee Berry Borer in Colombia. Final report of the DFID-Cenicafé-CABI Bioscience IPM for coffee Project (CNTR 93/1536A), https://www.sidalc.net/cgi-bin/wxis.exe/?IsisScript=CAFE.xis&method=post&formato= 2&cantidad=1&expresion=mfn=019976 (Chinchiná, Colombia DFID-Cenicafé, 1999).
Mejía, J. W., Bustillo, A. E., Orozco, H. J. & Cháves, B. Efecto de cuatro insecticidas y de Beauveria bassiana sobre Prorops nasuta (Hymenoptera:Bethylidae), parasitoide de la broca del café. Rev. Colomb. Entomol. 26(3–4), 117–123 (2000).
McCullagh, P. & Nelder, J. A. Generalized Linear Models 1–532 (Chapman and Hall/CRC, Boca Raton, 1989) (ISBN 9780412317606).
Lactin, D. J., Holliday, N. J., Johnson, D. L. & Craigen, R. Improved rate model of temperature dependent development by arthropods. Environ. Entomol. 24, 68–75 (1995).
Fargues, J., Goettel, M. S., Smits, N., Ouedraogo, A. & Rougier, M. Effect of temperature on vegetative growth of Beauveria bassiana from different origins. Mycologia 89(3), 383–392 (1997).
Ouedraogo, A., Fargues, J., Goettel, M. S. & Lomer, C. J. Effect of temperature on vegetative growth among isolates of Metarhizium anisopliae and M. flavoviride. Mycopathologia 137, 37–43 (1997).
Reyes, I. C., Bustillo, A. E. & Chaves, B. Efecto de Beauveria bassiana y Metarhizium anisopliae sobre el parasitoide de la broca del café, Cephalonomia stephanoderis. Rev. Colomb. Entomol. 21(4), 199–204 (1995).
Castillo, A., Gómez, J., Infante, F. & Vega, F. E. Susceptibilidad del parasitoide Phymastichus coffea LaSalle (Hymenoptera: Eulophidae) a Beauveria bassiana en condiciones de laboratorio. Neotrop. Entomol. 38, 665–670 (2009).
Banu, J.G. Efficacy of entomopathogenic nematodes against coleopteran pests. In Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes (eds Mahfouz, M.M. et al.) 174–192 (CABI, Oxfordshire, 2017).
Aristizábal, L. F., Salazar, E. H. & Mejía, M. C. Cambios en la adopción de los componentes del manejo integrado de la broca del café Hypothenemus hampei (Coleoptera: Scolytidae) a través de metodologías participativas. Rev. Colomb. Entomol. 28(2), 153–160 (2002).
O’Hara, R. B. & Kotze, D. J. Do not log-transform count data. Methods Ecol. Evol. 1, 118–122 (2010).
Akaike, H. Information theory and an extension of the maximum likelihood principle. In Second International Symposium on Information Theory (eds. Petrov, V.N. & Csaki, F.) 267–281 (Academiai Kiado, Budapest, 1973).
Georgescu-Roegen N. Energy and economic myths: institutional and analytical economic essays. Pergamon, New York, 1976) https://www.elsevier.com/books/energy-and-economic-myths/georgescu-roegen/978-0-08-021027-8.
Neuenschwander, P. et al. Impact assessment of the biological control of the cassava mealybug, Phenacoccus manihoti Matile-Ferrero (Hemiptera: Pseudococcidae), by the introduced parasitoid Epidinocarsis lopezi (De Santis) (Hymenoptera: Encyrtidae). Bull. Entomol. Res. 79, 579–594 (1989).
Gutierrez, A. P., Pitcairn, M. J., Ellis, C. K., Carruthers, N. & Ghezelbash, R. Evaluating biological control of yellow starthistle (Centaurea solstitialis) in California: a GIS based supply–demand demographic model. Biol. Control 34, 115–131 (2005).
Gutierrez, A. P. & Ponti, L. Deconstructing the control of the spotted alfalfa aphid Therioaphis maculata. Agric. For. Entomol. 15, 272–284 (2013).
Aristizábal, L. F. et al. Establishment of exotic parasitoids of the coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) in Colombia through farmer participatory research. Int. J. Trop. Insect Sci. 32, 24–31 (2012).
Vergara, J. D., Bustillo, A. E. & Cháves, B. Biología de Phymastichus coffea en condiciones de campo. Revista Cenicafé (Colombia) 52, 97–103 (2001).
Johnson, M. A., Fortna, S., Hollingsworth, R. G. & Manoukis, N. C. Postharvest population reservoirs of coffee berry borer (Coleoptera: Curculionidae) on Hawai’i Island. J. Econ. Entomol. 112, 2833–2841 (2019).
Duque, H. & Cháves, C.B. Estudio Sobre la Adopción de Tecnología en Manejo Integrado de la Broca del café. Centro Nacional de Investigaciones del Café-Cenicafé, Chinchiná. https://books.google.com.co/books/about/Estudio_sobre_adopcion_del_manejo_integr.html?id=pDpGMwEACAAJ&redir_esc=y (2000).
Bustillo, A. E. et al. Manejo integrado de la broca del café Hypothenemus hampei (Ferrari) en Colombia. Chinchiná, Colombia. Cenicafé, https://biblioteca.cenicafe.org/handle/10778/848 (1998).
Aristizábal, L. F., Vélez, J. C. & León, C. A. Diagnóstico del manejo integrado de la broca del café, Hypothenemus hampei (Coleoptera: Curculionidae) con caficultores caldenses. Rev. Col. Entomol. 32, 117–124 (2006).
Atallah, S. S., Gómez, M. I. & Jaramillo, J. A bioeconomic model of ecosystem services provision: Coffee Berry Borer and shade-grown coffee in Colombia. Ecol. Econ. 144, 129–138 (2018).
Hernandez-Aguilera, J. N., Conrad, J. M., Gómez, M. I. & Rodewald, A. D. The economics and ecology of shade-grown coffee: A model to incentivize shade and bird conservation. Ecol. Econ. 159, 110–121 (2019).
Infante, F. Pest management strategies against the coffee berry borer (Coleoptera:Curculionidae: Scolytinae). J. Agric. Food Chem. 66, 5275–5280 (2018).
Mendesil, E. et al. Semiochemicals used in host location by the coffee berryborer, Hypothenemus hampei. J. Chem. Ecol. 35(8), 944–950 (2009).
Borsa, P. & Gingerich, D. P. Allozyme variation and an estimate of the inbreeding coefficient in the coffee berry borer, Hypothenemus hampei (Coleoptera:Scolytidae). Bull. Entomol. Res. 85(1), 21–28 (1995).
Gingerich, P. D., Borsa, P., Suckling, D. M. & Brun, L. O. Inbreeding in the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae) estimated from endosulfan resistance phenotype frequencies. Bull. Entomol. Res. 86, 667–674 (1996).
Andreev, D., Breilid, H., Kirkendall, L., Brun, L. O. & Ffrench-Constant, R. H. Lack of nucleotide variability in a beetle pest with extreme inbreeding. Insect. Mol. Biol. 7(2), 197–200 (1998).
Benavides, P., Aspectos genéticos de la broca del Café. In La broca del café en América Tropical: Hallazgos y Enfoques. (eds Barrera J. F. et al.) 101–110 (Sociedad Mexicana de Entomología and El Colegio de la Frontera Sur, México, 2007).
Acknowledgements
We thank two reviewers for extensive suggestions and uncited references. The authors are grateful for the support from the Fund for Research of the Universidad Militar Nueva Granada, Bogotá, Colombia (CIAS 1569, 2530) and CASAS Global NGO, Kensington CA, USA. We extend our acknowledgements to the MED-GOLD project that has received funding from the European Union’s Horizon 2020, reserach and innovation programme, under grant agreement No. 776467, for funding the continuation of our research on coffee. We thank Ramón Vargas and Jorge Iván Valencia for the insights on the economy of coffee marketing.
Author information
Authors and Affiliations
Contributions
J.R.C., D.R., A.P.G. and L.P. contributed equally to this work. J.R.C. and D.R. elaborated Fig. 1.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cure, J.R., Rodríguez, D., Gutierrez, A.P. et al. The coffee agroecosystem: bio-economic analysis of coffee berry borer control (Hypothenemus hampei). Sci Rep 10, 12262 (2020). https://doi.org/10.1038/s41598-020-68989-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-68989-x
This article is cited by
-
Preparing Colombian coffee production for climate change: Integrated spatial modelling to identify potential robusta coffee (Coffea canephora P.) growing areas
Climatic Change (2024)
-
Effects of classical biocontrol agents on indigenous natural enemies: reproduction in pomegranate butterfly Deudorix livia eggs by native and imported parasitoids
Journal of Pest Science (2024)
-
Neem-Extract Formulation on Hypothenemus hampei Preference and Performance in Arabica Coffee Fruits and Artificial Diet
Neotropical Entomology (2024)
-
Tritrophic analysis of the prospective biological control of brown marmorated stink bug, Halyomorpha halys, under extant weather and climate change
Journal of Pest Science (2023)
-
Predation of Coffee Berry Borer by a Green Lacewing
Neotropical Entomology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
