
Abstract
A salt-tolerant denitrifying bacterium strain F2 was isolated from seawall muddy water in Dalian City, Liaoning Province, China. Strain F2 was identified by morphological observations, physiological and biochemical characteristics and 16 S rDNA identification. The salt tolerance of strain F2 was verified and the factors affecting the removal ability of strain F2 to nitrous nitrogen (NO2–N) and nitrate nitrogen (NO3–N) in saline conditions were investigated. Strain F2 was identified as Alishewanella sp., named Alishewanella sp. F2. Strain F2 can tolerate NaCl concentrations up to 70 g/L, and its most efficient denitrification capacity was observed at NaCl concentrations of 0−30 g/L. In the medium with NaCl concentrations of 0−30 g/L, strain F2 exhibited high removal efficiencies of NO2–N and NO3–N, with the removal percentages for both NO2–N and NO3–N of approximately 99%. In saline conditions with 30 g/L NaCl, the optimum culture pH, NaNO2 initial concentrations and inoculation sizes of strain F2 were 8−10, 0.4−0.8 g/L and 5−7%, respectively. Strain F2 was highly effective in removing NO2–N and NO3–N in saline conditions, and it has a good application potential in saline wastewater treatment.
Similar content being viewed by others

Introduction
In the past 40 years, with the rapid development of aquaculture industry in coastal areas of China, a large amount of coastal aquaculture wastewater has been discharged, which brought about various negative impacts on the environment1. The coastal aquaculture wastewater usually contains both inorganic salts (on average of 10−30 g/L NaCl) and many contaminants including nitrogen2. Large amounts of nitrogen pollutants (i.e., nitrous nitrogen (NO2–N) and nitrate nitrogen (NO3–N)) which are continuously released from the uneaten feed residue can result in the increase of inorganic nitrogen pollution in aquaculture water year by year1,3. Furthermore, with the rapid development of industrialization, a large number of saline wastewater sources, e.g., vegetable pickled wastewater4, textile wastewater5, and oily wastewater6, etc., with NaCl concentrations exceeding 30 g/L have been produced in various industrial processes. Most saline industrial wastewater also contains a large amount of nitrogen pollutants7,8. The increasing discharge of saline wastewater from aquaculture and industry without any treatment has threatened the aquatic, terrestrial and wetland ecosystems9,10. Therefore, the removal of nitrogen pollutants from saline wastewater has become an urgent problem.
Nitrite, an intermediate product of nitrification and denitrification, is frequently detected in water bodies and seriously threatens the aquatic organisms and human health11,12. The concentration of nitrite in wastewater is up to 50 mg/L or more11. High concentrations of nitrite seriously endanger the growth and normal metabolism of aquatic organisms. Humans accidentally drinking water containing high concentration of nitrite could easily lead to impaired intelligence and form strong carcinogens in the human body (e.g., nitrosamines), and even lead to death. As the most stable form of nitrogenous compounds in the aerobic environment, nitrate has a high solubility and can migrate and diffuse rapidly in water, resulting in secondary pollution13,14. The discharging of untreated and unqualified wastewater containing nitrate into surface water bodies will bring serious potential safety hazards and threaten the growth of plants, animals and human health15. Therefore, effectively reducing the NO2–N and NO3–N concentrations in water is of great practical significance for both ecosystem and human health.
The main technologies for removing nitrogen from wastewater include physical, chemical and biological methods16,17,18,19. Compared with physical and chemical methods, biological methods have the advantages of high efficiency, low energy consumption, low cost, easy implementation, etc., and have been widely used to remove NO2–N from wastewater. Denitrification can convert NO2–N and NO3–N into gaseous nitrogen (i.e., nitrogen (N2) and nitrous oxide (N2O)), and fundamentally solve the nitrogen pollution problem in water20. Several nitrite-type denitrifying bacteria, e.g., Acinetobacter baumannii21, Pseudomonas putida22, and Pseudomonas tolaasii23, etc., have been screened and were proved to be efficient in removing NO2–N from wastewater. However, saline wastewater contains not only high concentrations of nitrogen but also a large amount of soluble salts. The presence of salts can significantly and negatively affect the removal of pollutants by microorganisms24,25. When microorganisms are exposed to highly saline environment, osmotic pressure is increased, resulting in the excessive loss of water in general microbial cells, the separation of protoplasm, the inhibition of microbial growth and metabolism, and even death26. The denitrification capacity of most denitrifying bacteria is inhibited by high salinity27. Therefore, salt-tolerant denitrifying bacteria with efficient NO2–N and NO3–N removal ability are required to be isolated for the treatment of saline wastewater.
The objectives of this study are to: 1) isolate and identify a salt-tolerant denitrifying bacterium strain F2 from seawall muddy water; 2) validate the salt tolerance as well as the denitrification capacity of strain F2; and 3) evaluate the effects of initial pH, NaNO2 initial concentration and inoculation size on the denitrification capacity of strain F2 in saline conditions. The results of this study will provide efficient microbial resource and optimal process parameters for microbial denitrification of saline wastewater, which is of great significance for protecting the water environment safety and human health.
Results
Isolation of denitrifying bacteria
An anoxic condition was created by submerged culturing of denitrification medium with NaNO2 as the sole nitrogen source, but a small amount of NaNO2 was still oxidized and rapidly converted into NaNO328. After repeating three times of enrichment and isolation, three strains of denitrifying bacteria with nitrite as sole nitrogen source were obtained, and numbered F1, F2 and F3, respectively. The three strains became turbid during the enrichment and cultivation in the denitrification medium that was accompanied by different degrees of gas generation (i.e., N2, N2O and/or nitric oxide (NO))29. The enrichment cultivation of each strain cultivated for 5 d is shown in Fig. S1 (in supplementary material).
Table 1 shows the removal percentages of NO2–N and NO3–N by each respective strain. After a 5 d cultivation, the removal percentages of NO2–N in denitrification medium by all the three strains were above 98%. However, the removal percentages of NO3–N by strain F2 were higher than that of other strains. The removal percentages of NO2–N and NO3–N by strain F2 were 98.38% and 96.13%, respectively, and gas was produced concomitantly in a 5 d cultivation. During the 5 d of cultivation, ammonia nitrogen (NH4+-N) concentration in the medium of strain F2 increased slightly, and the total nitrogen (TN) removal percentage by strain F2 was 20.99% (Fig. S2 in supplementary material), which indicated that some of the NO2–N and NO3–N in the medium were transformed into NH4+-N, and some were transformed into nitrogen-containing substances which are necessary for the growth of strain F2 through assimilation. Besides, according to the conservation of elements, the removal of TN in the medium was mainly due to the emission of gaseous nitrogen (i.e., N2, N2O and/or NO). After comprehensive analysis, strain F2, an ideal bacterium, was selected as the research object in the subsequent experiments.
Identification of strain F2
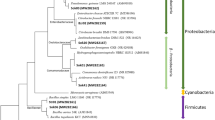
The physiological and biochemical characteristics of strain F2 are shown in Table 2. The extracted bacterial DNA was amplified using 16 S rDNA primers to a 1033 bp amplified fragment (Fig. S3 in supplementary material). The phylogenetic tree constructed by the MEGA 4.0 version software is shown in Fig. 1. The 16 S rDNA identification reveals that strain F2 was 99.81% homology genetic related with Alishewanella sp. N5 (GenBank accession no. EU287929.1). Therefore, according to the morphological observation and 16 S rDNA gene analysis, strain F2 was identified as Alishewanella sp., named Alishewanella sp. F2 (GenBank accession no. MN396708). It was deposited at the China General Microbiological Culture Collection Center (CGMCC) on March 25, 2019, numbered CGMCC No: 17433.

Molecular phylogenetic tree of strain F2 based on 16 S rDNA sequences.
Denitrification capacity of strain Alishewanella sp. F2
Strain Alishewanella sp. F2 was inoculated in a sterilized fresh denitrification medium containing NaNO2 as sole nitrogen source at a 5% inoculation size, and the concentrations of NO2–N and NO3–N in the culture solution were tested before and after the cultivation. The OD600 values of strain Alishewanella sp. F2 were measured, and the growth curve of strain Alishewanella sp. F2 was plotted (Fig. S4 in supplementary material). As shown in Fig. 2, the initial concentrations of NO2–N and NO3–N were 106.87 ± 4.58 mg/L and 45.60 ± 1.77 mg/L, respectively. There was basically no change in the concentrations of NO2–N and NO3–N within the first 24 h of cultivation, indicating that strain Alishewanella sp. F2 was in the growth adaptation period during the first 24 h. Strain Alishewanella sp. F2 exhibited inefficient denitrification capacity when it was in the stage of adapting to the new growth environment. After a 36 h cultivation, the concentrations of NO2–N and NO3–N in the medium decreased significantly (p < 0.05) to 27.45 ± 2.35 mg/L and 11.60 ± 5.35 mg/L with the removal percentages of 74.31% and 74.56%, respectively. Strain Alishewanella sp. F2 began to grow rapidly and entered the logarithmic growth phase (Fig. S4 in supplementary material) with obvious removal effects of NO2–N and NO3–N (Fig. 2). After a 48 h cultivation, the concentrations of NO2–N and NO3–N by strain Alishewanella sp. F2 decreased rapidly to 1.10 ± 0.14 mg/L and 2.37 ± 1.03 mg/L, with the corresponding removal percentages of 98.97% and 94.81%, respectively. During the 48−120 h cultivation, the removal percentages of NO2–N and NO3–N were above 98% and 94%, respectively. Strain Alishewanella sp. F2 entered a stable growth stage after 48 h and maintained high removal abilities of NO2–N and NO3–N until the end of 120 h. The efficient removal abilities of NO2–N and NO3–N of strain Alishewanella sp. F2 were achieved. During the cultivation process, the reduction of NO2–N and NO3–N concentrations was accompanied by gas generation, demonstrating that that strain Alishewanella sp. F2 can transform nitrites and nitrates into gas, thus achieving effective denitrification performance. For the control treatment without inoculating of strain Alishewanella sp. F2, the concentrations of NO2–N and NO3–N did not evidently change during the cultivation process (only 4% nitrite was transformed to nitrate).

Removal of NO2–N (a) and NO3–N (b) using strain Alishewanella sp. F2 in denitrification medium. Values represent the mean of three replicates and error bars represent standard deviations. Columns containing different letters indicate significant differences among treatments at p = 0.05.
Salt tolerance of Alishewanella sp. F2
Strain Alishewanella sp. F2 was separated from seawater muddy water, which was a saline environment, therefore, it was suspected that strain Alishewanella sp. F2 was salt tolerant. To further verify the salt tolerance and its threshold of strain Alishewanella sp. F2, strain Alishewanella sp. F2 was cultivated in double-layer screening-isolation medium with different salinity gradients for 5 d. The growth of strain Alishewanella sp. F2 is shown in Table 3. After cultivation of 2 d, there was obvious colony growth on the double-layer screening-isolation medium with 0 g/L and 30 g/L NaCl concentrations, and there were trace colonies on the medium with 50, 70 and 100 g/L NaCl concentrations. After cultivation of 5 d, obvious colony growth was observed on the medium with 0, 30, 50, and 70 g/L NaCl concentrations, and trace colony growth was observed on the medium with 100 g/L NaCl concentration. The results indicate that strain Alishewanella sp. F2 can tolerate a NaCl concentration of up to 100 g/L, although extreme high salt stress (i.e., 100 g/L NaCl) lead to a slower growth of strain Alishewanella sp. F2 compared to lower salt stress (i.e., 0, 30, 50 and 70 g/L NaCl).
Denitrification capacity of strain Alishewanella sp. F2 in saline conditions
The efficiencies of NO2–N and NO3–N removal by strain Alishewanella sp. F2 under different salinity treatment is shown in Fig. 3. The initial concentrations of NO2–N and NO3–N in denitrification medium of all the salinity treatments were maintained at 124.00 ± 8.72 mg/L and 52.36 ± 2.51 mg/L, respectively. Strain Alishewanella sp. F2 maintained high removal percentages for NO2–N and NO3–N when the NaCl concentration was at 30 g/L and below, with the NO2–N and NO3–N removal percentages of 99.00−99.34% and 98.99−99.17%, respectively. However, the removal ability of strain Alishewanella sp. F2 to both NO2–N and NO3–N was significantly (p < 0.05) inhibited by higher salinity treatments, i.e., NaCl concentration of 50−100 g/L in this study. In the medium with 50, 70, and 100 g/L NaCl concentrations, the removal percentages of NO2–N and NO3–N by strain Alishewanella sp. F2 were only 26.21–30.34% and 0–37.78%, respectively. The above observation indicates that the NaCl concentration higher than 50 g/L has a significantly (p < 0.05) negative effect on the denitrification capacity of strain Alishewanella sp. F2. Additionally, there was no significant difference in NO2–N removal percentage among salinity treatments of 50, 70, and 100 g/L NaCl. However, when the NaCl concentration was increased to 100 g/L, the removal percentage of NO3–N by strain Alishewanella sp. F2 was significantly reduced (p < 0.05) compared to 50 and 70 g/L NaCl treatments.

Effluent concentrations and removal percentages of NO2–N (a) and NO3–N (b) in denitrification medium with different salinity levels. Values represent the mean of three replicates and error bars represent standard deviations. Columns containing different letters indicate significant differences among treatments at p = 0.05.
Effect of pH, NaNO2 initial concentration and inoculation size on denitrification capacity of strain Alishewanella sp. F2
The denitrification capacity of strain Alishewanella sp. F2 was affected by different pH value (Fig. 4a). When the pH was 3, 5 and 7, the removal percentages of NO2–N by strain Alishewanella sp. F2 was 5.76−8.65%, and there was no removal of NO3–N. When the pH was 8, 9 and 10, the removal percentages of NO2–N (96.85−100%) and NO3–N (96.41−100%) by strain Alishewanella sp. F2 were significantly increased (p < 0.05) compared to lower pH treatments (i.e., 3, 5 and 7). When the pH was 11, the NO2–N was not removed and only a small amount of NO3–N (6.74%) was removed.

Denitrification capacity of strain Alishewanella sp. F2 as affected by different pH values (a), NaNO2 initial concentrations (b) and inoculation sizes (c) after a 5 d cultivation. Values represent the mean of three replicates and error bars represent standard deviations. Columns containing different letters indicate significant differences among treatments at p = 0.05.
The influence of different NaNO2 initial concentration on denitrification capacity of strain Alishewanella sp. F2 is shown in Fig. 4b. When the NaNO2 initial concentrations were 0.4 g/L and 0.8 g/L, the removal percentages of NO2–N and NO3–N were above 98% and 94%, respectively. With an increase in the NaNO2 initial concentration, the NO2–N and NO3–N removal abilities of strain Alishewanella sp. F2 were significantly decreased (p < 0.05). To be specific, the removal percentages of NO2–N and NO3–N by strain Alishewanella sp. F2 were significantly (p < 0.05) decreased when NaNO2 initial concentrations were in the range of 1.6−3.2 g/L, as compared with treatments of 0.4 g/L and 0.8 g/L. The average removal percentages of NO2–N and NO3–N with NaNO2 initial concentrations of 1.6−3.2 g/L were 18.90−43.81% and 8.85−35.94%, respectively.
As shown in Fig. 4c, the denitrification capacity of strain Alishewanella sp. F2 varied with different inoculation size. When the inoculation sizes were 1% and 3%, the NO2–N and NO3–N removal percentages were 54.53−59.50% and 45.73−58.24%, respectively. With an increase in the inoculation size, the NO2–N and NO3–N removal percentages of strain Alishewanella sp. F2 were significantly increased (p < 0.05). When the inoculation sizes were 5% and 7%, the average removal percentages of NO2–N and NO3–N were 99.33–99.84% and 94.66–96.23%, respectively. There was no significant difference in NO2–N removal when further increase the inoculation size to 10%. However, when the inoculation size was 10%, the NO3–N removal percentage by strain Alishewanella sp. F2 was significantly (p < 0.05) reduced compared to 5% and 7% treatments.
Discussion
In recent years, many bacteria with denitrification capacity have been studied. Denitrifying bacteria do not have a specific taxonomy in microbial taxonomy and they are scattered among many genera of prokaryotes. In a highly saline environment, the growth and metabolism of most bacteria are apt to be inhibited and even lead to death26. General marine microorganisms grow in a saline environment of 10−30 g/L NaCl, and belong to slightly halophilic bacteria, while moderately halophilic bacteria can grow in a saline environment of 30−145 g/L NaCl30. A few halophilic denitrifying bacteria were found, mainly distributed in Bacillus and Halomonas bacteria, and Halobacterium and Haloarcula of archaea, etc.31. In particular, the halophilic bacteria that can survive in salt-free conditions are defined as salt-tolerant bacteria32. For example, a salt-tolerant bacterium strain Bacillus hwajinpoensis SLWX2 was screened from sea water, and the removal percentages of NO2–N and NO3–N at 24 h in a concentration of 30 g/L NaCl by a strain of Bacillus hwajinpoensis SLWX2 were 99.5% and 85.6%, respectively33. In this study, a salt-tolerant denitrifying bacterium strain Alishewanella sp. F2 was isolated from seawall muddy water and its salinity tolerance ranged from 0 to 70 g/L NaCl. It is reported for the first time that the Alishewanella sp. has the characteristic of salt tolerance. Strain Alishewanella sp. F2 is different from the previous reported salt-tolerant denitrifying bacteria, which indicates the diversity of denitrifying bacteria in nature, and is of great significance for enriching the knowledge of denitrifying bacterial ecology.
In anoxic conditions, nitrites and nitrates mainly undergo denitrifying processes, which are converted to gaseous products, e.g., N2, N2O or other biochemical reduction products12,28. Most available studies used the anaerobic incubator (e.g., Gas-pak Anaerobic Jar) to create an anaerobic environment34,35. However, an anoxic environment is necessary for the culturing of facultative anaerobes. Therefore, the submerged culture and sandwiching plate method, which can create an anoxic environment, was developed in this study, and the ideal target strain Alishewanella sp. F2 exhibiting excellent NO2–N and NO3–N removal ability in saline conditions was obtained.
Environmental conditions not only affect the growth and metabolism of bacteria, but also affect the denitrification capacity36,37,38. Optimal environmental conditions and nutrients are necessary for promoting microbial growth and metabolism and improving biological nitrogen removal39. A variety of factors (i.e., salinity, pH, initial nitrite concentration) was considered during the screening of denitrifying bacteria in this study. Exploring the optimal condition for growth and denitrification of strain Alishewanella sp. F2 will provide important guidance for practical application in environmental conditions.
The growth and reproduction of bacteria in a medium go through four phases, i.e. lag phase, exponential phase, stationary phase and death phase40. The denitrification process of strain Alishewanella sp. F2 mainly occurred between 24 to 48 h after cultivation, for both NO3–N and NO2–N (Fig. 2). Combining with the growth curve of strain Alishewanella sp. F2 (Fig. S4 in supplementary material), it can be concluded that the denitrification mainly occurred in the exponential phase (12−48 h), which is similar to the strains reported in previous studies41,42. There was no obvious change in the NO2–N and NO3–N concentrations during the lag phase (0−12 h), stationary phase (48 h later) and death phase (Fig. S4 in supplementary material and Fig. 2), indicating that no obvious denitrification process occurred during these periods. During the exponential phase, bacteria grow fastest and metabolize most vigorously and the energy and reducing power needed for cell synthesis are mainly consumed at this stage43. Therefore, the highest denitrification efficiency of strain Alishewanella sp. F2 was observed during the exponential phase.
Alishewanella is a genus of the class Gammaproteobacteria in the phylum Proteobacteria44. Proteobacteria is the largest phylum in the bacterial domain and is commonly found in a variety of environments. Proteobacteria is salt tolerant, and it is dominant under 0−18 g/L salinity, and the relative abundance increases with the increasing salinity levels45. In addition, Gammaproteobacteria can tolerate highly saline environments45,46. In this study, strain Alishewanella sp. F2 showed a high ability to remove NO2–N and NO3–N in medium with 0−30 g/L NaCl concentrations, reflecting the high salt tolerance of strain Alishewanella sp. F2 in the experimental system. Besides its high salt tolerance, strain Alishewanella sp. F2 can also adapt to a high alkali condition (pH of 8−10). In the condition of pH = 10 and 30 g/L NaCl concentration, the removal percentages of strain Alishewanella sp. F2 to NO2–N and NO3–N with initial concentrations of 123.17 ± 1.01 mg/L and 52.07 ± 1.16 mg/L were 99.00% and 99.17%, respectively (Fig. 3). Strain Alishewanella sp. F2 has the characteristics of strong adaptability, fast growth rate, high nitrogen removal efficiency, etc. Therefore, the discovery of strain Alishewanella sp. F2 and the evaluation of its nitrogen removal characteristics in highly saline environments can provide reference for the microbial nitrogen removal process. The denitrification capacity of strain Alishewanella sp. F2 under salt-alkali stress indicates that strain Alishewanella sp. F2 has a promising application prospect in the treatment of saline wastewater. The culture conditions, denitrification characteristics and large-scale applications in the application of strain Alishewanella sp. F2 are recommended for further study.
Conclusion
Three denitrifying bacteria were isolated from seawall muddy water in Dalian city, Liaoning province, China. Strain F2 proved to be more effective in NO2–N and NO3–N removal than other strains, and was identified as Alishewanella sp., named Alishewanella sp. F2 (GenBank accession no. MN396708). Strain Alishewanella sp. F2 was deposited at the CGMCC on March 25, 2019, numbered CGMCC No: 17433.
Strain Alishewanella sp. F2 has a promising salt-tolerant denitrification capacity. The removal percentages of NO2–N and NO3–N by strain Alishewanella sp. F2 in a saline condition of 30 g/L NaCl were all above 99%. Besides, strain Alishewanella sp. F2 has an efficient denitrification capacity under high alkali conditions (pH of 8−10) and high initial nitrogen concentrations (NaNO2 of 0.4−0.8 g/L). In summary, strain Alishewanella sp. F2 is an efficient salt-tolerant denitrifying bacterium, which can be potentially applied in denitrification of saline wastewater in the future.
Materials and Methods
Sample collection and culture media description
Muddy water samples were collected from the estuary of an aquaculture wastewater stream in Dalian City, Liaoning Province, China (39°38′31′′ N, 122°58′19′′ E), and stored at 4 °C before further treatment in the Key Laboratory of Wetland Ecology and Environment, Chinese Academy of Sciences, China.
The culture media were described as follows: denitrification medium was composed of (g/L) CH3COONa 5, K2HPO4 1, NaNO2 0.8, CaCl2 0.03, NaCO3 1, FeSO4•7H2O 0.06, MgSO4•7H2O 0.2 and PH = 10. It was noteworthy that FeSO4•7H2O was added after the addition of deionized water to avoid oxidation of divalent iron (Fe2+) to ferric iron (Fe3+)47. Ingredients for the screening-isolation medium (pH = 10) and oblique tube preservation medium (pH = 10) were same as the denitrification medium but with 2% agar (m/v) added. All media were autoclaved at 121 °C for 30 min before applying.
Enrichment, isolation and screening of bacterial strains
Two mL seawall muddy water samples were added to the 250 mL Erlenmeyer flasks containing 200 mL denitrification medium with three replicates, cultivated in a constant temperature incubator at 30 °C for 5 d. The flasks were sealed with parafilm to avoid gas exchange. Three successive transfers were carried out in fresh denitrification medium by subculturing 2 mL inoculum and incubating for 5 d for each time.
Following the last transfer of 5 d, the cultures were diluted 1000-fold. A 0.5 mL of each dilution was spread on screening-isolation medium in the glass Petri dish. Then an unsolified screening-isolation medium (<40 °C) was poured on the culture for air isolation. The Petri dishes were sealed, turned over and placed at 30 °C in a constant temperature incubator until clear colonies appeared. Isolated colonies were streaked onto new double-layer screening-isolation medium dishes, purified by repeated streaking.
Denitrification capacity test of the isolated strains
The strains preserved on the preservation medium were inoculated into the denitrification medium and incubated for 5 d in hypoxic conditions at 30 °C. Each strain was inoculated into a fresh denitrification medium at a 5% inoculation size, and the flask mouth was sealed and statically cultivated at 30 °C for 5 d. During the incubation, the turbidity and gas production of the denitrification medium were observed and recorded. Finally, 10 mL liquid sample was taken periodically, 5 mL cultivated solution was taken at regular intervals, and centrifuged at 5000 r/min for 10 min, and the supernatant was then diluted and measured for NO2–N and NO3–N concentrations. The concentrations of NO2–N and NO3–N in water samples were determined by N-(1-naphthyl)-ethylenediamine spectrophotometry and naphthyl ethylenediamine hydrochloride spectrophotometry, respectively. The removal percentages of NO2–N and NO3–N were calculated to determine the denitrification capacity of strains.
In order to clarify the growth of bacteria and ensure the denitrification capacity, the test experiment was carried out when the bacteria were in logarithmic phase. A single bacterial colony was selected from the oblique tube preservation medium, inoculated using sterilizing inoculator, and cultivated in denitrification medium at 30 °C under anoxic condition. The turbidimetric method (OD600) was used to determine the growth of bacteria. The OD600 value of bacteria was determined every 12 h, and the growth curve of bacteria was drawn.
Identification of strain F2
According to the results of denitrification capacity test (see Result), the denitrification capacity of strain F2 was more efficient than other strains. Therefore, we focused on only strain F2 in the following experiments including the salt tolerance test and the analysis of influencing factors. The morphology and the physiological and biochemical experiments of strain F2 were carried out according to the Manual for Systematic Identification of Common Bacteria48 and the Bergey’s manual of systematic bacteriology49. Using an Ezup column bacterial genomic DNA extraction kit (Sangon Co. Ltd., China), the total genome DNA of strain F2 was extracted by a conventional method. Using total DNA as a template, the genomic DNA of strain F2 was amplified by the polymerase chain reaction (PCR) thermal cycler (Mastercycler, Eppendorf Co. Ltd., German). The primers for PCR reaction were 16 S rDNA amplification universal primers. The forward primer was 27 F 5′-AGAGTTTGATCCTGGCTCAG-3′ and the reverse primer was 1492 R 5′-GGTTACCTTGTTACGACTT-3′. The composition of the PCR reaction system is as follows: 0.5 μL of template DNA, 1 μL of dNTP (mix), 2.5 μL of Taq Buffer (with MgCl2), 0.2 μL of Taq enzyme, 0.5 μL of primer F (10 μM), 0.5 μL of primer R (10 μM), and double distilled water to 25 μL. Under the following conditions, the amplification of PCR was completed: requires at 94 °C for 4 min, denaturation at 94 °C for 45 s, annealing at 55 °C for 45 s, extension at 72 °C for 1 min, 30 cycles, repair extension at 72 °C for 10 min and reaction was terminated at 4 °C. The PCR amplification product of 5 μL was detected by 1% agarose gel electrophoresis. The 16 S rDNA sequencing of recycled PCR products was carried out in Sangon Biotech (Shanghai, China) Co., Ltd. The sequence obtained by 16 S rDNA sequencing was compared and analyzed by using the BLAST at the National Center for Biotechnology Information (NCBI) (http://:/www.ncbi.nlm.nih.gov/). The phylogenetic tree of strain F2 was constructed by Neighbor-Joining method in MEGA 4.0 (Arizona State University, 2007) software, and the species of strain F2 were determined.
Salt tolerance test of strain F2
The growth of strain F2 was observed while changing the salinity levels of screening-isolation medium. Five salinity levels of the screening-isolation medium were designed, i.e., NaCl concentrations of 0, 30, 50, 70, and 100 g/L, respectively. Strain F2 from the same concentration was streaked on each plate of the same area with designated salinity level and placed in an incubator at 30 °C for 5 d. Within each plate, the number of lines and the line spacing were ensured to be consistent. Each treatment was repeated three times to avoid the error caused by manual operation. The colony of strain F2 on the medium plate was determined for evaluating the tolerance of strain F2 to different salinity levels.
Evaluation of the denitrification capacity of strain F2 under different salinity levels
The denitrification medium with different salinity levels was prepared by setting NaCl concentrations at 0, 30, 50, 70, and 100 g/L, respectively. Strain F2 was inoculated in the denitrification medium at a 5% inoculation size, and the flask mouth was sealed and statically cultivated at 30 °C for 5 d. The removal percentages of strain F2 to NO2–N and NO3–N under each respective salinity level were determined and calculated as described above.
Effects of initial pH values, NaNO2 initial concentrations, and inoculation sizes on denitrification capacity of strain F2 in saline conditions
Strain F2 was cultivated in a fresh denitrification medium with 30 g/L NaCl concentration for 5 d until the culture solution was cloudy and there was gas produced. As influencing factors, initial pH (i.e., 3, 5, 7, 9, 10 and 11), NaNO2 initial concentrations (i.e., 0.4, 0.8, 1.6, 2.4 and 3.2 g/L) and inoculation sizes (v/v) (1, 3, 5, 7 and 10%) were observed for the effects on the NO2–N and NO3–N removal of strains. In this experiment, only each respective factor that tested was changed while the other conditions remained constant. After cultivation for 5 d, samples were taken to determine the concentrations of NO2–N and NO3–N.
Statistical analysis
All results were presented as the average of three independent experiments. The data presented in the figures were expressed as means ± standard deviation. Means between different treatments were compared by one-way ANOVA with Tukey’s test at the significance level of 0.05. All statistical analyses were performed by using Microsoft Office Excel 2007 and SPSS 22.0 for Windows system. All graph design was carried out by Origin 9.1 for Windows system.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Liang, Y. et al. Historical Evolution of Mariculture in China During Past 40 Years and Its Impacts on Eco-environment. Chinese Geographical Science 28, 363–373, https://doi.org/10.1007/s11769-018-0940-z (2018).
Li, C. et al. Fast start-up strategies of MBBR for mariculture wastewater treatment. J. Environ. Manage. 248, 109267–109267, https://doi.org/10.1016/j.jenvman.2019.109267 (2019).
Chen, Y., Dong, S. L., Wang, F., Gao, Q. F. & Tian, X. L. Carbon dioxide and methane fluxes from feeding and no-feeding mariculture ponds. Environmental Pollution 212, 489–497, https://doi.org/10.1016/j.envpol.2016.02.039 (2016).
Abou-Elela, S. I., Kamel, M. M. & Fawzy, M. E. Biological treatment of saline wastewater using a salt-tolerant microorganism. Desalination 250, 1–5, https://doi.org/10.1016/j.desal.2009.03.022 (2010).
Lin, J. Y. et al. Sustainable Management of Textile Wastewater: A Hybrid Tight Ultrafiltration/Bipolar-Membrane Electrodialysis Process for Resource Recovery and Zero Liquid Discharge. Ind. Eng. Chem. Res. 58, 11003–11012, https://doi.org/10.1021/acs.iecr.9b01353 (2019).
Pendashteh, A. R. et al. Evaluation of membrane bioreactor for hypersaline oily wastewater treatment. Process Safety and Environmental Protection 90, 45–55, https://doi.org/10.1016/j.psep.2011.07.006 (2012).
Xie, D. H. et al. Ion-exchange membrane bioelectrochemical reactor for removal of nitrate in the biological effluent from a coking wastewater treatment plant. Electrochem. Commun. 46, 99–102, https://doi.org/10.1016/j.elecom.2014.06.020 (2014).
Yu, H., Li, J., Dong, H. Y. & Qiang, Z. M. Nitrogen removal performance of marine anammox bacteria treating nitrogen-rich saline wastewater under different inorganic carbon doses: High inorganic carbon tolerance and carbonate crystal formation. Bioresour. Technol. 288, 8, https://doi.org/10.1016/j.biortech.2019.121565 (2019).
Liang, Y. X. et al. Constructed wetlands for saline wastewater treatment: A review. Ecol. Eng. 98, 275–285, https://doi.org/10.1016/j.ecoleng.2016.11.005 (2017).
Zhao, C., Zhang, H., Song, C., Zhu, J.-K. & Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. The Innovation, https://doi.org/10.1016/j.xinn.2020.100017 (2020).
Kroupova, H., Machova, J. & Svobodova, Z. Nitrite influence on fish: a review. Vet. Med. 50, 461–471, https://doi.org/10.17221/5650-vetmed (2005).
Kroupova, H. K., Valentova, O., Svobodova, Z., Sauer, P. & Machova, J. Toxic effects of nitrite on freshwater organisms: a review. Reviews in Aquaculture 10, 525–542, https://doi.org/10.1111/raq.12184 (2018).
Bhatnagar, A. & Sillanpaa, M. A review of emerging adsorbents for nitrate removal from water. Chem. Eng. J. 168, 493–504, https://doi.org/10.1016/j.cej.2011.01.103 (2011).
Tyagi, S., Rawtani, D., Khatri, N. & Tharmavaram, M. Strategies for Nitrate removal from aqueous environment using Nanotechnology: A Review. J. Water Process. Eng. 21, 84–95, https://doi.org/10.1016/j.jwpe.2017.12.005 (2018).
Sevda, S., Sreekishnan, T. R., Pous, N., Puig, S. & Pant, D. Bioelectroremediation of perchlorate and nitrate contaminated water: A review. Bioresour. Technol. 255, 331–339, https://doi.org/10.1016/j.biortech.2018.02.005 (2018).
Lochmatter, S., Maillard, J. & Holliger, C. Nitrogen Removal over Nitrite by Aeration Control in Aerobic Granular Sludge Sequencing Batch Reactors. Int. J. Environ. Res. Public Health 11, 6955–6978, https://doi.org/10.3390/ijerph110706955 (2014).
Wang, W. F., Cao, L. X., Tan, H. M. & Zhang, R. D. Nitrogen removal from synthetic wastewater using single and mixed culture systems of denitrifying fungi, bacteria, and actinobacteria. Appl. Microbiol. Biotechnol. 100, 9699–9707, https://doi.org/10.1007/s00253-016-7800-5 (2016).
Awual, M. R., Asiri, A. M., Rahman, M. M. & Alharthi, N. H. Assessment of enhanced nitrite removal and monitoring using ligand modified stable conjugate materials. Chem. Eng. J. 363, 64–72, https://doi.org/10.1016/j.cej.2019.01.125 (2019).
Yun, L. et al. Ammonia nitrogen and nitrite removal by a heterotrophic Sphingomonas sp. strain LPN080 and its potential application in aquaculture. Aquaculture 500, 477–484, https://doi.org/10.1016/j.aquaculture.2018.10.054 (2019).
Gong, Y. K., Peng, Y. Z., Yang, Q., Wu, W. M. & Wang, S. Y. Formation of nitrous oxide in a gradient of oxygenation and nitrogen loading rate during denitrification of nitrite and nitrate. Journal of Hazardous Materials 227, 453–460, https://doi.org/10.1016/j.jhazmat.2012.05.002 (2012).
Cao, H. P. et al. Isolation and characterization of a denitrifying Acinetobacter baumannii H1 using NO2–N as nitrogen source from shrimp farming ponds. Afr. J. Microbiol. Res. 6, 2258–2264, https://doi.org/10.5897/ajmr11.814 (2012).
He, T., Li, Z. & Xu, Y. Denitrification characteristics of a hypothermia nitrite-denitrifier Pseudomonas putida Y-12. Acta Scientiae Circumstantiae 35, 3071–3077 (2015).
He, T., Xu, Y. & Li, Z. Identification and characterization of a hypothermia nitrite bacterium Pseudomonas tolaasii Y-11. Wei sheng wu xue bao = Acta microbiologica Sinica 55, 991–1000 (2015).
Ng, K. K., Shi, X. Q., Ong, S. L., Lin, C. F. & Ng, H. Y. An innovative of aerobic bio-entrapped salt marsh sediment membrane reactor for the treatment of high-saline pharmaceutical wastewater. Chem. Eng. J. 295, 317–325, https://doi.org/10.1016/j.cej.2016.03.046 (2016).
Chen, L. J. et al. Shifts in soil microbial metabolic activities and community structures along a salinity gradient of irrigation water in a typical arid region of China. Science of the Total Environment 598, 64–70, https://doi.org/10.1016/j.scitotenv.2017.04.105 (2017).
Liu, C., Yamamoto, T., Nishiyama, T., Fujii, T. & Furukawa, K. Effect of salt concentration in anammox treatment using non woven biomass carrier. J. Biosci. Bioeng. 107, 519–523, https://doi.org/10.1016/j.jbiosc.2009.01.020 (2009).
Leung, J. Y. S., Cai, Q. & Tam, N. F. Y. Comparing subsurface flow constructed wetlands with mangrove plants and freshwater wetland plants for removing nutrients and toxic pollutants. Ecol. Eng. 95, 129–137, https://doi.org/10.1016/j.ecoleng.2016.06.016 (2016).
Philips, S., Laanbroek, H. J. & Verstraete, W. Origin, causes and effects of increased nitrite concentrations in aquatic environments. Reviews in Environmental Science &. Biotechnology 1, 115–141 (2002).
Zhang, Z., Zhang, Y. & Chen, Y. Recent advances in partial denitrification in biological nitrogen removal: From enrichment to application. Bioresour. Technol. 298, https://doi.org/10.1016/j.biortech.2019.122444 (2020).
Ventosa, A., Nieto, J. J. & Oren, A. Biology of moderately halophilic aerobic bacteria. Microbiol. Mol. Biol. Rev. 62, 504–+ (1998).
Zumft, W. G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 61, 533 (1997).
Tindall, B. J. et al. The family Halobacteriaceae. The Prokary-otes 1, 768–808 (1992).
Cheng, Y., Li, Q.-F., Fei, Y.-T. & Zhang, Y. Screening and Nitrogen Removing Characteristics of Heterotrophic Nitrification-Aerobic Denitrification Bacteria SLWX2 from Sea Water. Huan jing ke xue=. Huanjing kexue 37, 2681–2688, https://doi.org/10.13227/j.hjkx.2016.07.035 (2016).
Bonin, P. C., Michotey, V. D., Mouzdahir, A. & Rontani, J. F. Anaerobic biodegradation of squalene: Using DGGE to monitor the isolation of denitrifying Bacteria taken from enrichment cultures. FEMS Microbiol. Ecol. 42, 37–49, https://doi.org/10.1016/s0168-6496(02)00304-5 (2002).
Zhao, F. et al. Characterization and Evaluation of a Denitrifying and Sulfide Removal Bacterial Strain Isolated From Daqing Oilfield. Pet. Sci. Technol. 33, 694–701, https://doi.org/10.1080/10916466.2014.1003941 (2015).
Silva, C. F. L. et al. Heterotrophic nitrifying/aerobic denitrifying bacteria: Ammonium removal under different physical-chemical conditions and molecular characterization. J. Environ. Manage. 248, 109294, https://doi.org/10.1016/j.jenvman.2019.109294 (2019).
Han, H., Song, B., Song, M. J. & Yoon, S. Enhanced Nitrous Oxide Production in Denitrifying Dechloromonas aromatica Strain RCB Under Salt or Alkaline Stress Conditions. Front. Microbiol. 10, 11, https://doi.org/10.3389/fmicb.2019.01203 (2019).
Hang, Q. Y. et al. Application of plant carbon source for denitrification by constructed wetland and bioreactor: review of recent development. Environ. Sci. Pollut. Res. 23, 8260–8274, https://doi.org/10.1007/s11356-016-6324-y (2016).
Chen, S. H., He, S. Y., Wu, C. J. & Du, D. Y. Characteristics of heterotrophic nitrification and aerobic denitrification bacterium Acinetobacter sp. T1 and its application for pig farm wastewater treatment. J. Biosci. Bioeng. 127, 201–205, https://doi.org/10.1016/j.jbiosc.2018.07.025 (2019).
Lilleorg, S. et al. Bacterial ribosome heterogeneity: Changes in ribosomal protein composition during transition into stationary growth phase. Biochimie 156, 169–180, https://doi.org/10.1016/j.biochi.2018.10.013 (2019).
Zhou, D.-D., Ma, F., Wang, H.-Y. & Dong, S.-S. Study on screening methord of aerobic denitrifiers. Wei sheng wu xue bao = Acta microbiologica Sinica 44, 837–839 (2004).
Shi, X., Li, Y., Xing, G. & Kang, X. Screening and Primary Identification of an Aerobic Denitrifier Isolate with Salt Tolerance. Biotechnology Bulletin, 175-180 (2013).
Zwietering, M. H., Rombouts, F. M. & Vantriet, K. Comparison of definitions of the lag and exponential phase in bacterial growth. Journal of Applied Bacteriology 72, 139–145, https://doi.org/10.1111/j.1365-2672.1992.tb01815.x (1992).
Euzeby, J. P. List of bacterial names with standing in nomenclature: A folder available on the Internet. Int. J. Syst. Bacteriol. 47, 590–592, https://doi.org/10.1099/00207713-47-2-590 (1997).
Fu, G., Han, J., Yu, T., Huangshen, L. & Zhao, L. The structure of denitrifying microbial communities in constructed mangrove wetlands in response to fluctuating salinities. J. Environ. Manage. 238, 1–9, https://doi.org/10.1016/j.jenvman.2019.02.029 (2019).
Elifantz, H., Horn, G., Ayon, M., Cohen, Y. & Minz, D. Rhodobacteraceae are the key members of the microbial community of the initial biofilm formed in Eastern Mediterranean coastal seawater. FEMS Microbiol. Ecol. 85, 348–357, https://doi.org/10.1111/1574-6941.12122 (2013).
Wu, S., Vymazal, J. & Brix, H. Critical Review: Biogeochemical Networking of Iron in Constructed Wetlands for Wastewater Treatment. Environmental science &. technology https://doi.org/10.1021/acs.est.9b00958 (2019).
Dong X Z, C. M. Y. Manual for Systematic Identification of Common Bacteria. (Science Press, 2001).
MichaelGoodfellow et al. Bergey’s Manual® of Systematic Bacteriology. (2012).
Acknowledgements
This work was supported by Open Project of State Key Laboratory of Urban Water Resource and Environment, Harbin Institute of Technology (NO. ESK 201802); the National Key Research and Development Program of China (No. 2016YFC0500404-4); and the Youth Innovation Promotion Association CAS (No. 2017274).
Author information
Authors and Affiliations
Contributions
R.C. and X.W. carried out all experiments and drafted the manuscript. H.Z. (corresponding author) is responsible for this study, participated its design and help to draft the manuscript. B.Y., B.S., Y.X. and B.F. provided technical and theoretical support. H.W. executed the experiments, analyzed all data and produced figures. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cheng, R., Wang, X., Zhu, H. et al. Isolation and characterization of a salt-tolerant denitrifying bacterium Alishewanella sp. F2 from seawall muddy water. Sci Rep 10, 10002 (2020). https://doi.org/10.1038/s41598-020-66989-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-66989-5
This article is cited by
-
Microbial diversity and ecological interactions of microorganisms in the mangrove ecosystem: Threats, vulnerability, and adaptations
Environmental Science and Pollution Research (2022)
-
A heterotrophic nitrification-aerobic denitrification bacterium Halomonas venusta TJPU05 suitable for nitrogen removal from high-salinity wastewater
Frontiers of Environmental Science & Engineering (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
