
Abstract
Large well-preserved crocodylomorph tracks from the Lower Cretaceous (? Aptian) Jinju Formation of South Korea, represent the well-known crocodylomorph ichnogenus Batrachopus. The Korean sample includes multiple, narrow-gauge, pes-only trackways with footprint lengths (FL) 18–24 cm, indicating trackmaker body lengths up to ~3.0 m. Surprisingly, the consistent absence of manus tracks in trackways, with well-preserved digital pad and skin traces, argues for bipedal trackmakers, here assigned to Batrachopus grandis ichnosp. nov. No definitive evidence, either from pes-on-manus overprinting or poor track preservation, suggests the trackways where made by quadrupeds that only appear bipedal. This interpretation helps solve previous confusion over interpretation of enigmatic tracks of bipeds from younger (? Albian) Haman Formation sites by showing they are not pterosaurian as previously inferred. Rather, they support the strong consensus that pterosaurs were obligate quadrupeds, not bipeds. Lower Jurassic Batrachopus with foot lengths (FL) in the 2–8 cm range, and Cretaceous Crocodylopodus (FL up to ~9.0 cm) known only from Korea and Spain registered narrow gauge trackways indicating semi-terrestrial/terrestrial quadrupedal gaits. Both ichnogenera, from ichnofamily Batrachopodidae, have been attributed to Protosuchus-like semi-terrestrial crocodylomorphs. The occurrence of bipedal B. grandis ichnosp. nov. is evidence of such adaptations in the Korean Cretaceous.
Similar content being viewed by others


Introduction
Crocodylomorph tracks are generally rare in the Mesozoic of Asia. It has been suggested that this is in part due to the lack of sedimentary facies representing suitable habitats for this group of ostensibly aquatic trackmakers1. However, the crocodylomorph ichnofamily Batrachopodidae2,3 appears to represent more terrestrially-adapted forms3,4. As currently defined, the batrachopodids5 include Batrachopus and Crocodylopodus, the former mostly known from small tracks (~2.0–8.0 cm long) from the Lower Jurassic of North America4, Europe5,6 and Africa7, the latter primarily from the Cretaceous of Europe8,9. The ichnofamily also includes Antipus known only from one well-described Lower Jurassic trackway from North America which is considered a synonym of Batrachopus by some3, but not all ichnologists10.
Recently Crocodylopodus was reported from the Lower Cretaceous (?Aptian)11 Jinju Formation of Korea12,13, where it represents the first Asian occurrence, and adds to the extraordinarily rich Jinju Formation, ichnofauna described as a Konservat-Lagerstätten14,15,16,17,18,19. Spanish8,9 and Korean12,13 Crocodylopodus represent relatively small animals (footprint lengths less than ~9.0–10.0 cm) with slender digit traces, which may reflect penetrative track preservation20. However, the trackway configuration is diagnostically crocodilian, with elongate tetradactyl pes and outwardly rotated pentadactyl manus, not unlike those of extant crocodilians21,22,23 (Supplementary Information: SI Fig. 1).
Here we report on a large, newly-discovered Jinju Formation assemblage with multiple trackways of large crocodylomorph tracks with footprint lengths up to 24.0 cm, from the Sacheon Jahye-ri tracksite, near Sacheon City (Fig. 1). These tracks are more than twice as large as any previously reported batrachopodid tracks and closely resemble Batrachopus with well-preserved pes footprints with clear digital pad impressions and localized skin traces. Surprisingly the trackways never include manus imprints and therefore appear to indicate exclusively bipedal progression, a gait not known or previously inferred from fossil crocodylomorph trackways, or argued convincingly from the functional morphology of potential trackmakers (SI).

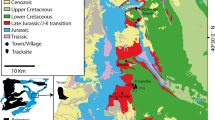
Location maps and stratigraphy for the Gyeongsang Supergroup. (A) location of the Gyeongsang Basin in southeast of Korean Peninsula, with Group-level geological map. (B) Formation-level geological map of the southwestern part of the Gyeongsang Basin showing study area (Sacheon Jahye-ri tracksite) west of Sacheon City. (C) Formation-level stratigraphy of region. Maps made by K-S K and M G L in Adobe Photoshop (version CS6 88) and Canvas X (version, 20 Build 390, http://www.canvasgfx.com/).
In contrast to the common Mesozoic crocodylomorph trackway Hatcherichnus24, representing swimming behavior, and it’s rare walking counterpart Mehliella25, Batrachopus has been attributed to a Protosuchus-like crocodylomorph3,4 with a narrow trackway interpreted to represent terrestrial progression. Evidence of bipedalism is consistent with such a terrestrial gait.
The unexpected discovery of trackways so suggestive of bipedal locomotion by Cretaceous crocodylomorphs has ichnological implications bearing directly on long-standing and controversial debates about the gait of pterosaurs. While, most pterosaurian trackways represent quadrupedal26 not bipedal progression27, multiple pes-only trackways, from the Haman Formation of Korea have been erroneously attributed to giant bipedal pterosaurs28. This unexpected evidence of apparently bipedal crocodylomorphs obliges us to investigate the alternative possibility that the trackway configurations represent an unusual mode of preservation, rather than bipedal locomotion, and underscores the need for a reexamination of the Batrachopodidae29 and other relevant, morphologically-similar ichnotaxa.
Ichnological material and geological setting
In recent years the Lower Cretaceous (?Aptian) Jinju Formation, in the Jinju City area of South Korea has yielded an extraordinary volume and diversity of tetrapod tracks including “first” discoveries of small hopping mammal tracks, Koreasaltipes jinjuensis14, truly diminutive dromaeosaur tracks Dromaeosauriformipes rarus15, the first Korean examples of the theropod track Corpulentapus16, the first turtle tracks from Korea30, the aforementioned first report of Crocodylopodus12, the oldest known frog tracks18, diminutive theropod tracks (Minisauripus) with skin traces19, and the largest known Cretaceous lizard track assemblage17. While many other track assemblages and facies remain under active investigation, the present study focusses on the documentation of the large batrachopodid tracks and trackways from the Sacheon Jahye-ri tracksite (Figs. 1 and 2) from which large numbers of large Batrachopus tracks were recovered. This is the first global report of large Batrachopus and the first to indicate bipedal progression. They form the basis of the new ichnotaxon Batrachopus grandis ichnosp. nov., described in detail below.

Stratigraphy (left) of the Sacheon Jahye-ri tracksite with aerial view (top right), with elevation of surface (bottom right) showing cross section with main stratigraphic layers. Graphics made by K-S K and M G L in Adobe Photoshop (version CS6 88) and Canvas X (version, 20 Build 390, http://www.canvasgfx.com/). Photographs for the aerial view collected with a Sony A7R digital camera (25 mm lens) and the elevation images of surface using Agisoft PhotoScan Professional (v. 1.4.3) and esri ArcMap (v. 10.2.2.).
The Sacheon Jahye-ri tracksite represents an area of excavation approximately 40 × 20 meters (800 m2) in extent which has yielded four track-bearing levels from a outcrop and representing about 5 meters of stratigraphic section within the Jinju Formation (Fig. 2). Tracks are abundant at all levels, with high concentrations of crocodylomorph (Batrachopus) tracks and trackways at levels 2 and 4 (Figs. 3–6).

Photos and outline drawings of trackways and trackway segments of Batrachopus grandis ichnosp. nov. from in situ outcrops. T1-T10 and holotype trackway T16 (enlarged, with length and width rectangle indicating minimal rotation) represent level 2 and T11-T14 represents level 4. Compare with Figs. 4–6, SI Figs. 3 and 4 and SI Table 1. Photos and line drawings made and compiled by K-S K and M G L, in Adobe Photoshop (version CS6 88) and Canvas X (version, 20 Build 390, http://www.canvasgfx.com/).

Photographs of well-preserved in situ Batrachopus grandis ichnosp. nov. track impressions from surface 2. (A,B) Left pes tracks; (C,D) Right pes tracks. A, C = pes of T7 and T9, respectively. B shows skin impression of heel region (see SI Fig. 5). D = holotype IS 2 of Fig. 6. Photos by J-W K, S-M B and M G L, and compiled in Adobe photoshop (version CS6 88) and Canvas X (version, 20 Build 390, http://www.canvasgfx.com/).

Photos and 3D images of track casts and trackway segments. (A–D): Photos of SJ 117, 119, 138 and trackway 1; (a–d): 3D images of SJ 117, 119, 138 and trackway 1. Compare with Fig. 3 and SI Figs. 3 and 4. Photos and 3D images made and compiled by K-S K, S-M B and A R, in Adobe photoshop (version CS6 88) and Canvas X (version, 20 Build 390, http://www.canvasgfx.com/).

(A) Photograph of the surface from which tracings and 3D images of trackways T16 (the holotype) and T17 of Batrachopus grandis and isolated tracks IS 5-6 were obtained; (B–D) 3D images of pair of tracks including right pes track from holotype trackways T 16; (E) map of the surface shown in A, showing holotype trackway T 16 and paratype trackway T 17. Note well-defined digital pad traces especially in holotype. Photographs, line drawings and 3D images made and compiled by K-S K, M G L and A R. in Adobe Photoshop (version CS6 88) and Canvas X (version, 20 Build 390, http://www.canvasgfx.com/). Compare with Figs. 3–5, SI Figs. 3 and 4 and SI Table 1.
Standard methods of in situ, and laboratory track and trackway documentation, were used to document the large volume of ichnological material collected, the largest Batrachopus sample currently known (SI). The track-bearing layers were necessarily exposed by working from higher to lower stratigraphic levels, and resulted in removal of a large number of natural casts (convex hyporeliefs) that had filled the natural impressions (concave epireliefs) on successively lower surfaces including levels 2 and 4 from which many specimens, mostly casts were collected. Comprehensive methods of documentation and analysis described in the Supplementary Information, included 2D photography, 3D photogrammetry, tracing of track outlines and measurement of all standard track and trackway parameters (Figs. 3–6 and SI Figs. 3 and 4, SI Table 1).
Description of tracks and trackways
General observations
The tracks here formally referred to Batrachopus grandis ichnosp. nov., are all pes traces, occurring in narrow trackways (Fig. 3) and are consistently about twice as long as wide (Figs. 4–6) when including the heel trace which appears to be registered in all cases. Given the natural morphological division of the pes tracks into an anterior and posterior portion separated by a transverse crease (Figs. 4–6), it is possible to provide length and width measurements for both the entire track, including the heel and the anterior portion (SI Table 1). Providing both measurements is important in making comparisons with Jurassic Batrachopus, including the type material from North America, which only reveals sporadically or faintly preserved heel traces. In some cases the heel trace is absent and the distinctive bi-lobed posterior margin of the anterior portion of the tracks is clearly seen. In the following description, the lengths of digit traces and their inter-digital divarication angles are all measured from the posterior-most margin of the heel (SI Fig. 2).
Field observations indicated some differences in the quality of preservation of B. grandis ichnosp. nov. tracks from the level 2 and 4 surfaces. General tracks from the level 2 surfaces were shallower with clearly defined digit and digital pad impressions (Figs. 3–6), as well as transverse creases. They also yielded the majority of well-preserved natural casts. The well-defined digit and pad traces, rank these tracks as 2.5–3.0 on the four point preservation scale (0-1-2-3)31,32. One track shows well-defined skin (scale) impressions in the heel area that resemble those typical of modern crocodilians (Fig. 4B and SI Fig. 5)21,22,23. The tracks from level 4 were generally deeper and slightly more elongate, which we infer to be related to softer substrate conditions: see trackways T11–T14 (Fig. 3). In these trackways, which are slightly narrower, with corresponding higher pace angulation values, digit traces are still quite well-preserved, although digital pad traces are less well-defined.
Systematic description
Batrachopodidae Lull 1904, emended Lockley and Meyer 2004.
Batrachopus grandis ichnosp. nov.
Holotype: Trackway T 16 (Figs. 3, 4D and 6A. Table 1); with two replicas, CUE SJ IS 2 R and UCM 214.326
Paratypes: Trackways T1-T14 and T17 (Fig. 3, Table 1) and isolated (IS) tracks in Sacheon Jahye-ri (SJ) series SJ 001 – SJ 140 (SI Fig. 3).
Type locality: the Sacheon Jahye-ri tracksite, near Sacheon City, South Korea
Type horizon: Jinju Formation, Lower Cretaceous (?Aptian)11.
Differential diagnosis
B. grandis ichnosp. nov. is much larger than all previously described Batrachopus ichnospecies and morphotypes, mostly ‘lumped’ under the Lower Jurassic ichnogenus B. deweyi3, and differs from them in consistently displaying a well-defined heel making up on average ~40% of total pes length. B. grandis ichnosp. nov. is also less outwardly rotated than type Batrachopus (B. deweyi): i.e., in the former morphotype digit III parallels the trackway mid line, not digit II as in the latter ichnospecies with more outwardly rotated pes. No other Batrachopus ichnospecies has been described in such detail, from such a large and well preserved sample, even with skin traces, as Korean B. grandis ichnosp. nov. This allows for the definition of anatomy-consistent and diagnostic features such as the relative heel length and relative digit divarication angles with III-IV consistently greater than I-II and II-III (SI Fig. 3): i.e., ∠III-IV > ∠I-II and ∠II-III.
Description
Trackway of a tetrapod with tetradactyl pes. The manus traces were not found but register as pentadactyl traces in other Batrachopus ichnospecies. Pes twice as long as wide with narrow heel, separated from wider anterior portion, with digit traces, by a transverse, bi-lobed crease with anteriorly convex sulcus. Total track length averaging 20.7 cm (range 18.0–24.0 cm), along axis of longest digit III, with mean lengths of digits I, II and IV respectively as 14.7, 18.6 and 19.4 cm (from posterior margin of heel). Thus, III > IV > II > I. Digit traces up to ~2.0 cm wide, with well-defined pad impressions, generally in the better preserved specimens indicating a phalangeal pad formula of 1-2-3-4 corresponding to digits I, II, III and IV. However, the creases that differentiate the pad traces are not always clearly registered, especially in digit IV. Length of that portion of the track anterior to the transverse crease averaging 12.2 cm (range 11.0–15.1 cm): thus L/W ratio of anterior digital portion of the pes is 1.27 (12.2 cm/9.6 cm), with maximum track width situated anteriorly near distal margins of digits I and IV. Total I-IV digit divarication averaging 31.2° (range 28.0–38.0°) with mean divarication values for I-II, II-III and III-IV as 9.3°, 9.4° and 12.3° respectively: (∠III-IV > ∠I-II and ∠II-III). Skin traces consist of angular polygonal scale traces about 1.0 cm in diameter, and are presently known in the heel region.
Trackway narrow without manus traces on surfaces where pes traces registered. Step averaging 37.3 cm (range 29.8–47.0 cm); stride averaging 69.3 cm (range 50.8–81.0 cm); pace angulation averaging 148.5° (range 135–165°); outer trackway width (OTW) averaging 18.7 cm and inner trackway width averaging 0.6 cm and ranging from 5.5 to - 4.0 cm (SI Table 1).
Systematic discussion
Heel traces occur sporadically in type Batrachopus, i.e., B. deweyi from the Lower Jurassic of the Connecticut Valley region3, and are regarded as possibly incorporating a reduced digit V, never illustrated as a separate digit trace3,33. However they are inconsistently and incompletely registered, thus appearing in only two of seven consecutive tracks in the type trackway, and never with skin traces. In the case of the type of B. “gracilis” trackway3 there are no heel traces in this trackway, as is the case for most other known examples5,6,26.
Crocodylopodus described from the basal Cretaceous of Spain8 is based on a well-defined holotype trackway with tetradactyl pes, only 3.6 cm long with narrow digit traces that do not show pad impressions or any skin traces (SI Fig. 1B). The manus is pentadactyl and outwardly rotated, also with very narrow digit traces. Ostensibly the narrow digit traces distinguish Crocodylopodus from Batrachopus, with the former showing less outward rotation of the pes, but greater outward rotation of the manus tracks. If the difference in digit trace width is explained as a preservation feature, with the narrow traces being considered penetrative tracks20, the difference between Batrachopus and Crocodylopodus may be explained in part by differential preservation.
Recent studies of crocodylomorph tracks from the Cretaceous of Korea, notably from other Jinju Formation sites, have applied the term Crocodylopodus to small tracks (pes length less than 9.0 cm) with narrow digit traces that closely resemble Spanish Crocodylopodus12,13 (SI Fig. 6). These Korean Crocodylopodus trackways represent quadrupedal progression and none represent animals even half the size, in linear footprint dimensions, of the B. grandis ichnosp. nov. trackmaker. Although both the large and the small Korean crocodylomorphs tracks are considered batrachopodids, there is, as yet, no compelling case that their ichnogenus or ichnospecies level taxonomies are indistinguishable, or that the trackmakers (of B. grandis ichnosp. nov. and Korean Crocodylopodus) were the same. In order to make a case for strong similarity, cogent arguments for the role of preservational factors are needed, and discussed below.
Discussion
Evidence for bipedalism of the B. grandis ichnosp. nov. trackmaker
Various crocodylomorph tracks have been reported from the Mesozoic and Cenozoic13,22 and assigned to at least eleven ichnogenera (Table 1) of which six occur exclusively in the Jurassic and Cretaceous. The discovery of B. grandis ichnosp. nov. was made soon after the discovery and description of Korean Crocodylopodus12,13 and significantly enhances our understanding of the morphology, size range, abundance and preservational factors affecting Korean crocodylomorph tracks: i.e., Batrachopodidae footprints (Table 1), and more generally to the range of morphotypes attributable to this ichnofamily.
From an ichnotaxonomic viewpoint it has already been argued that B. grandis ichnosp. nov. can be differentiated from all other reports of Batrachopus. It differs from other morphotypes not only in size, but in being more plantigrade: i.e., always registering heel traces both in shallower and deeper tracks and in showing less outward rotation of the pes (Fig. 6). It also has very well-preserved digital pad traces and even some scale impressions. If the trackmaker was truly bipedal (either an obligate or facultative biped) as seems persuasive based on the consistent biped configuration in trackways, a case could be made for a higher level of taxonomic differentiation, say at the ichnogenus level: i.e. bipedalism would be another diagnostic difference. We first examine the possibility of bipedality and, second, consider the alternate possibilities.
It is well known that in addition to obligate bipeds and obligate quadrupeds, there are also various facultative bipeds and quadrupeds. Among the archosaurs we recognized that some sauropodomorphs (“prosauropods”) were facultative bipeds, as shown by the occurrence of both pes-only and manus-and-pes trackways of Otozoum34,35. It is also well known that various ornithischians were facultative quadrupeds, as demonstrated by the various examples of trackways assigned to the ichnogenera Anomoepus, Moyenisauropus36 and Caririchnium indicate37. In all these cases the manus tracks are much smaller than those of the pes, which appears to have been universally the case with terrestrial archosaurs. This is true not only of those described as “small manus” forms with greater heteropody, but also among “large manus” morphotypes with less heteropody38,39,40,41. From this it is possible to infer that most forms carried most of their weight and over the pelvic girdle, and it has been noted that an elongate heel trace is often associated with bipedal forms: i.e., as an expression of posture with posterior emphasis41. However, as discussed for sauropods41 the force exerted on the substrate by manus and pes is a function of both weight and foot size: i.e., weight distribution. Thus, in the case of sauropods, it was shown, both in theory and in the rock record that the manus could penetrate more deeply that the pes (Fig. 7), creating different impacts on different sedimentary layers, in some cases creating “manus only” trackways42. This largely dispelled speculations that manus-only trackways were the result of sauropod swimming activity. Other, compelling lines of evidence suggesting that the Korean B. grandis trackways are attributable to bipedal crocodylomorphs come from the known occurrence of large bipedal crocodylomorphs from the early Mesozoic43,44. However, to date no such bipedal crocodylomorphs have been reported from the Cretaceous.

How differential depth of manus and pes may affect trackway configurations. Note manus only and manus dominated trackways are common in the sauropod track record (left). The possibility of a pes deeper than manus scenario to explain apparent evidence of a bipedal crocodilian (right) is considered a possibly, but highly unlikely. See text for details. Original graphics made and compiled by M G L and A R, in Adobe Photoshop (version CS6 88) and Canvas X (version, 2017 Build 160, http://www.canvasgfx.com/).
Possible arguments against bipedalism of the B. grandis trackmaker
Although the sauropod swim tracks debate need not be reviewed in detail here (Fig. 7), it is pertinent to note that the pes-only B. grandis ichnosp. nov. trackways can only logically be attributed to deeper penetration of the pes. If the manus had penetrated more deeply, or as deeply, as in the case of some sauropods, one would expect to see deeper manus traces on the same surfaces as the pes tracks. There is no evidence for this at the Sacheon Jahye-ri tracksite.
This argument leads logically to only a few possible conclusions. The first is that the manus did register footprints as the trackmaker progressed quadrupedally, but that the manus did not penetrate as deeply as the pes, because it exerted less force: i.e., weight was distributed mostly over the pes, beneath the pelvic, rather than the pectoral girdle (Fig. 7). This inference, which implies that the manus registered on stratigraphically higher surfaces, which perhaps had different compositions, consistencies and preservation potential, is potentially consistent with the evidence that all other batrachopodid trackways reveal a smaller manus than pes. But such a scenario assumes that the large track sinks deeper, and does not factor in size and weight distribution differences. There is no documented evidence to suggest that the manus was less clearly or deeply impressed than the pes in small quadrupedal batrachopodid trackways. So there is no direct evidence that the pes tracks penetrated to register on surfaces buried beneath the surfaces on which the manus tracks are inferred to have registered. Therefore, this interpretation, implying that all manus tracks registered on surfaces other than those on which the pes is consistently found, is also very difficult to support in the light of the recognition of other tracksites, discussed below, with similar evidence of similar ostensibly bipedal large batrachopodids.
Analysis of the limb, foot and weight distribution of possible crocodylomorph trackmakers known from Mesozoic body fossils has the potential to shed light on this issue. Study of trackways registered by modern crocodilians during terrestrial walking progression is also informative21, and indicates that pes and manus track morphologies are quite similar to those of Mesozoic crocodylomorphs. However, the trackways of well-known extant species such as Alligator mississippiensis and Crocodylus acutus are generally wider than those reported from the Mesozoic21,45 (Fig. 8). Moreover, while it is known that modern species can run bipedally for short distances, trackways produced during such running progression are rare and to the best of our knowledge not recorded or mapped to scales from which morphometric data can be obtained. On the other hand there are videos of modern crocodilian progressing subaqueously with only the pes contacting the substrate in a regular right-left-right cycle46,47. Again, the substrate expressions of such subaqueously-registered tracks, corresponding to video footage, have not been recorded. However, such subaqueous progression does potentially provide a plausible mechanism for producing pes only trackways.

Left to right: Comparison of trackways of modern Crocodylus acutus45, Batrachopus grandis ichnosp. nov., Mehliella jeffersonensis (with tail trace)25 and the pterosaur trackway Haenamichnus uhangriensis (right)56, showing difference in trackway width. Note that B. grandis ichnosp. nov. trackway indicates a biped and that the trackway is narrow with inner trackway width negative. Original line drawings made and compiled by M G L, in Adobe Photoshop (version CS6 88) and Canvas X (version, 20 Build 390, http://www.canvasgfx.com/). Compare with SI Fig. 1. See text for details.
While said video footage46,47 of modern crocodilians progressing subaqueously shows that they may make tracks with the pes while the manus makes no contact with the substrate, the trackway patterns produced by such ‘punting’ modes of locomotion48,49,50 have not been recorded. We may infer that any punting pes tracks would be incomplete pes toe tracks of the “swim track” variety49,51. Therefore, we cannot infer that B. grandis ichnosp. nov. tracks and regular trackway patterns represent punting progression.
Modern crocodilian trackways made during normal walking are wide gauge. While similar in overall shape and digit trace proportions, they do not show clear digital pads, but conversely, unlike B. grandis ichnosp. nov., they show interdigital web traces. Thus, we can infer that B. grandis ichnosp. nov. tracks are different from those of extant crocodilians. This conclusion points again to the likelihood that the B. grandis ichnosp. nov. trackmaker was a terrestrial or semi-terrestrial form, with the weight of evidence supporting bipedality43,44.
Distinguishing crocodilian from pterosaurian trackways
An unexpected result of the discovery of B. grandis trackway has been to shed light on a the controversial issue of pterosaur locomotion debated since the 1980s27 and 1990s26: were pterosaurs bipedal or quadrupedal? These debates, mainly concerning relatively small pterosaurian tracks, have largely been resolved in favor of quadrupedalism52. However, some uncertainty remained regarding tracks of purported ‘giant’ pterosaurians that were described as ‘enigmatic’ and inferred to have progressed bipedally28. These trackways from the Lower Cretaceous, Haman Formation, at the Gain-ri tracksite, Korea were named Haenamichnus gainensis28 and inferred to represent, large, plantigrade pterodactyloid pterosaurs that might have walked bipedally so that the long wings did not become mired in the substrate. It was further inferred they may have been wading in shallow water.
We can now confirm confidently, that these tracks from the Gain-ri tracksite and others from Adu Island28: (SI Fig. 7) are identical to poorly preserved large Batrachopus trackways. Thus, they should be removed from Haenamichnus28,50 and regarded as large poorly preserved batrachopodid tracks. The type specimen then technically becomes Batrachopus gainensis (comb nov.). Thus, H. gainensis becomes a footnote to ichnotaxonomic history, shown to be an extramorphological expressions large of Batrachopus, only recognizable retrospectively after comparison with B. grandis. Therefore ichnologists may retrospectively choose to regard H. gainensis as a nomen dubium, and find little value in the trival name (gainensis). Alternatively they may simply refer to the Haman Formation tracks as Batrachopus cf. grandis.
Note that the Gain-ri and Adu island trackways are from the Haman Formation and so these occurrences indicate a widespread distribution in space (three sites) and time (two formations) of this distinctive apparently bipedal morphotype. The pes tracks from the two Haman Formation sites are also larger (27.5–39.0 cm long), but with trackway proportions (step, stride, pace angulation etc.,) quite similar to those from the Jinju Formation.
The identification of the Haman Formation trackways as poorly preserved large batrachopodid tracks apparently suggests that the trackmakers habitually progressed bipedally. Alternatively the same speculative arguments for apparent rather than real bipedalism would have to be invoked as was the case with the Jinju material. Moreover, in almost all cases the trackways are very narrow gauge with a narrower straddle than seen in modern crocodylians (Fig. 8). It is also of interest that least five sub parallel more or less equally spaced trackways53 were registered on the level 4 surface. This suggests either that the trackmakers may have been gregarious, or that they were following a physically controlled route, such as a shoreline, defined by the paleoenvironment54.
The overall length of extant crocodylians can be estimated from their tracks using the pes, body length ratio 1:1255. Based on this proportion the largest B. grandis ichnosp. nov. tracks from the Jinju Formation indicate a trackmaker with length of up to about 3.0 m (pes length 0.25 m × 12) and no less than about 2.16 m (0.18 m × 12). If we include the large poorly preserved tracks from the Haman Formation the overall length of the trackmaker could have been up to 4.68 m (0.39 m × 12).
Conclusions
The Lower Cretaceous Jinju Formation has yielded the largest known Batrachopus track morphotype, and the largest known assemblage, including many well-preserved tracks and trackways with clearly defined digital pad impressions and localized skin traces. These rank high (2–3) on the four point 0-1-2-3 quality of preservation scale31,32, and form the basis of the new ichnospecies Batrachopus grandis ichnosp. nov.
Surprisingly, the trackways appear to represent bipedal progression which is atypical of all known smaller batrachopodid trackways. This suggests gaits atypical of large crocodylomorphs, except for a few early Mesozoic reports43,44. Less well-preserved large batrachopodid tracks from the overlying Haman Formation, previously described as “enigmatic” and incorrectly assigned the pterosaurian ichnogenus Haenamichnus (as H. gainensis) are reinterpreted as Batrachopodidae tracks, also representing large crocodylomorphs. These apparently also indicate bipedal progression, and increase the database for this trackway morphotype in space and time.
The possibility that B. grandis ichnosp. nov. represents quadrupedal progression where manus tracks are not recognized because they registered on a different higher sedimentary surface is not supported by such evidence. Likewise the possibility that ‘unrecognized’ B. grandis ichnosp. nov. manus tracks were overprinted by the pes is also not supported by the evidence. Both scenarios, while intriguing and necessary to consider, are based only on inference and negative evidence, rather than the abundant trackway evidence.
The scenario that the trackmakers were progressing by punting through shallow water, only using their hind feet is considered implausible due to the fully plantigrade registration of pes tracks in narrow regular trackways rather that the partial toe-tip tracks that result from punting or swim tracks.
The trackway evidence of a large crocodylomorph with bipedal or facultative bipedal gait in the Lower Cretaceous is surprising, but consistent with terrestrial or semi-terrestrial adaptation, reported from the early Mesozoic trackmakers, and could potentially be supported by future body fossil evidence from the Cretaceous record.
References
Lockley, M. G. et al. Tracking crocodiles and turtles in the Cretaceous: comparisons between North America and east Asia in The 12th Symposium on Mesozoic Terrestrial, Ecosystems, Abstracts vol., Shenyang China, Aug, 16–20th 2015 (eds. Zhang, Y., Wu, S. Z. & Sun, G.) 193–195 (2015).
Lull, R. S. Fossil Footprints of the Jura-Trias of North America. Memoirs of the Boston Society of Natural History 5, 461–557 (1904).
Olsen, P. E. & Padian, K. Earliest records of Batrachopus from the southwestern United States, and revision of some early Mesozoic Crocydilomorph ichnogenera in The Beginning of the Age of Dinosaurs (ed. Padian, K.) 259–273 (Cambridge University Press, 1986).
Colbert, E. H. & Mook, C. C. The ancestral crocodile Protosuchus. American Mus. Nat. Hist. Bull 97, 143–182 (1951).
Lockley, M. G. & Meyer, C. Crocodylomorph trackways from the Jurassic to Early Cretaceous of North America and Europe: implications for ichnotaxonomy. Ichnos 11, 167–178 (2004).
Lapparent, A. Fde & Montenat, G. Les empreintes des pas de retiles de l’infralias du Veillon (Vendée). Bulletin Société Géologique de France, nouvelle série, Mémoire 107, 1–41 (1967).
Olsen, P. E. & Galton, P. M. A review of the reptile and amphibian assemblages from the Stormberg of southern Africa, with special emphasis on the footprints and the age of the Stormberg. Paleontologica Africana 25, 87–110 (1984).
Fuentes Vidarte, C. & Meijide Calvo, M. Primeras Huellas de Cocodrilo en el Weald de Cameros (Sria, Espana) Nueva Familia Crocodilopodidae: Nuevo icnogenero: Crocodylopodus Nueva icnoespecie: C. meijidei. Actas de las jornadas internacionales sobre paleontologia de dinosairios y su entorno. Sala de los infantes (Burgos, Espana). Collectivo Arqueologico- Paleontologico de Salas, 329–338 (1999).
Pascual Arribas, C., Hernandez Madrano, N., Latorre Macarron, P. & Sanzperez, E. Nuevo rastro de ichnitas de cocodrilo en la aloformacion Herteles de la Cuanca de Cameros Yacimiento del Barranco de Valdelavilla (Valdelavilla, Soria, Espana). Studia Geologica Salmanticensia 41, 77–91 (2005).
Coombs, W. P. Jr. Re-description of the ichnospecies Antipus flexiloquus Hitchcock, from the Early Jurassic of the Connecticut Valley. Journal of Paleontology 70, 327–331 (1996).
Kang, H. C. & Paik, I. S. Review on the geological ages of the formations in the Gyeongsang Basin, Korea. Journal of the Geological Society of Korea 49, 17–29 (2013).
Park, H. D., Kim, K. S., Lim, J. D. & Kim, D. H. A preliminary study on crocodile tracks from the Cretaceous Jinju Formation of Jinju City, Gyeongnam in Spring Meeting of the Korean Earth Science Society and Gyeongnam Goseong International Symposium (eds. Kim, K. S., Kim, D. H. & Lim, J. D.) 143 (The Korean Earth Science Society & Goseong County, 2016) (in Korean).
Lockley, M. G. et al. First reports of Crocodylopodus from Asia: implications for the paleoecology of the Lower Cretaceous. Cretaceous Research (2020) (online, March 2020).
Kim, K. S. et al. Korean trackway of a hopping, mammaliform trackmaker is first from the Cretaceous of Asia. Cretaceous Research 74, 188–191 (2017).
Kim, K. S. et al. Smallest known raptor tracks suggest microraptorine activity in lakeshore setting. Scientific Reports 8, 16908, https://doi.org/10.1038/s41598-018-35289-4 (2018).
Kim, K. S. et al. First reports of a distinctive theropod track assemblage from the Jinju Formation (Lower Cretaceous) of Korea provides strong correlations with China. Cretaceous Research 81, 26–35 (2018).
Kim, K. S. et al. Largest Cretaceous lizard track assemblage, new morphotypes and longest trackways comprise diverse components of an exceptional Korean Konservat-Lagerstätten ichnofauna. Scientific Reports 9, 1327816908, https://doi.org/10.1038/s41598-019-49442-0 (2019).
Kim, K. S., Lockley, M. G., Lim, J. D. & Kim, D. H. The oldest known Anuran (frog) trackways from the Jinju Formation, Lower Cretaceous, Korea. Cretaceous Research 96, 142–148 (2019).
Kim, K. S., Lockley, M. G., Lim, J. D. & Xing, L. Exquisitely-preserved, high-definition skin traces in diminutive theropod tracks from the Cretaceous of Korea. Scientific Reports 9, 2039, https://doi.org/10.1038/s41598-019-38633-4 (2019).
Gatesy, S. M. & Falkingham, P. Hitchcock’s Leptodactyli, penetrative tracks, and dinosaur footprint diversity. Journal of Vertebrate Paleontology (in review).
Farlow, J. O. & Elsey, R. M. Footprints and trackways of the American alligator, Rockefeller Wildlife Refuge, Louisiana. New Mexico Museum of Natural History and Science Bulletin 51, 31–39 (2010).
Milàn, J., Lucas, S. G., Lockley, M. G. & Spielmann, J. A. Crocodyle tracks and traces. (New Mexico Museum of Natural History and Science Bulletin 51, (2010).
Milàn, J. & Hedegaard, R. Interspecific variation in tracks and trackways from extant crocodylians. New Mexico Museum of Natural History and Science Bulletin 51, 15–29 (2010).
Foster, J. R. & Lockley, M. G. Probable crocodilian tracks and traces from the Morrison Formation (Upper Jurassic) of eastern Utah. Ichnos 5, 121–129 (1997).
Lockley, M. G. A solution to the Mehliella mystery: tracking, naming, identifying and measuring the first crocodylian trackway reported from the Cretaceous (Dakota Group, Colorado). New Mexico Museum of Natural History and Science Bulletin 51, 157–164 (2010).
Lockley, M. G. et al. The fossil trackway Pteraichnus is pterosaurian, not crocodilian: implications for the global distribution of pterosaurs tracks. Ichnos 4, 7–20 (1995).
Padian, K. & Olsen, P. E. The fossil trackway Pteraichnus: not pterosaurian but crocodilian. Journal of Paleontology 58, 178–184 (1984).
Kim, J. Y. et al. Enigmatic giant pterosaur tracks, and associated ichnofauna from the Cretaceous of Korea: implications for bipedal locomotion of pterosaurs. Ichnos 19, 50–65 (2012).
Le Loeuff, J. et al. Late Early Cretaceous crocodyliform trackways from northeastern Thailand. New Mexico Museum of Natural History and Science Bulletin 51, 175–178 (2010).
Kim, J. Y. & Lockley, M. G. First report of turtle tracks from the Lower Cretaceous of Korea. Cretaceous Research 64, 1–6 (2016).
Belvedere, M. & Farlow J. O. A numerical scale for quantifying the quality of preservation of vertebrate tracks in Dinosaur tracks: the next steps (eds. Falkingham, P. L., Marty, D. & Richter, A.) 92–98 (Indiana University Press, 2016).
Marchetti, L. et al. Defining the morphological quality of fossil footprints. Problems and principles of preservation in tetrapod ichnology with examples from the Palaeozoic to the present. Earth Science Reviews 193, 109–145 (2019).
Hunt, A. P., Lucas, S. G. & Klein, H. Late Triassic nonmarine vertebrate and invertebrate trace fossils and the pattern of the Phanerozoic record of vertebrate trace fossils. In The Late Triassic World (pp. 447–544). Springer, (2018).
Rainforth, E. C. Revision and re–evaluation of the Early Jurassic dinosaurian ichnogenus Otozoum. Palaeontology 46, 803–838 (2003).
Lockley, M. G., Lucas, S. G. & Hunt, A. P. Evazoum and the renaming of northern hemisphere “Pseudotetrasauropus”: implications for tetrapod ichnotaxonomy at the Triassic-Jurassic boundary. New Mexico Museum of Natural History of Science Bulletin 37, 199–206 (2006).
Lockley, M. G. & Gierlinski, G. Diverse vertebrate ichnofaunas containing Anomoepus and other unusual trace fossils from the Lower Jurassic of the western United States: implications for paleoecology and palichnostratigraphy. New Mexico Museum of Natural History and Science Bulletin 37, 175–191 (2006).
Lockley, M. G. & Wright, J. L. The trackways of large quadrupedal ornithopods from the Cretaceous: a review in Mesozoic Vertebrate Life: new research inspired by the Paleontology of Philip J. Currie (eds. Carpenter, K. & Tanke, D.) 428–442 (Indiana University Press, 2001).
Peabody, F. Reptile and amphibian trackways from the Lower Triassic Moenkopi Formation of Arizona and Utah. University of California publications, Bulletin of the Dept. of Geological Sciences 27, 295–468 (1948).
Lockley, M. G. Trackways-dinosaur locomotion in Paleobiology II (eds. Briggs, D. E. G. & Crowther, P.) 412–416 (Blackwell, Oxford, 2001).
Lockley, M. G. The morphodynamics of dinosaurs, other archosaurs and their trackways: holistic insights into relationships between feet, limbs and the whole body in Ichnology at the crossroads: a multidimensional approach to the science of organism - substrate interactions (eds. Bromley, R. & Melchor, R.) Society of Economic Paleontologists and Mineralogists Special Publication 88, 27–51 (2007).
Lockley, M. G. & Rice, A. Did Brontosaurus ever swim out to sea? Ichnos 1, 81–90 (1990).
Lockley, M. G., Pittman, J. G., Meyer, C. A. & Santos, V. F. On the common occurrence of manus-dominated sauropod trackways in Mesozoic carbonates. Gaia 10, 119–124 (1994).
Gautier, J. A., Nesbitt, S. J., Schachner, E. R., Bever, G. S. & Joyce, W. G. The bipedal stem crocodilian Poposaurus gracilis: inferring function in fossils and innovation in archosaur locomotion. Bull. Peabody Mus. Nat. Hist 52, 107–126 (2011).
Zanno, L. E., Drymala, S., Nesbitt, S. J. & Schneider, V. P. Early crocodylomorph increases top tier predator diversity during rise of dinosaurs. Scientific Reports 5, 9276, https://doi.org/10.1038/srep09276 (2015).
Farlow, J. O. et al. Trackways of the American crocodile (Crocodylus acutus) in northwestern Costa Rica: implications for crocodylian ichnology. Ichnos 25, 30–65 (2017).
Mustoe, G. E. Lower Eocene footprints from northwest Washington USA, Part 1: Reptile Tracks. Geosciences 9, 321, https://doi.org/10.3390/geosciences9070321 (2019).
Lockley, M. G. et al. Crocodile waterways and dinosaur freeways: implications of multiple swim track assemblages from the Cretaceous Dakota Group, Golden area, Colorado. New Mexico Museum of Natural History and Science Bulletin 51, 137–156 (2010).
Lee, Y. N. et al. Unusual locomotion behavior preserved within a crocodyliform trackway from the Upper Cretaceous Bayanshiree Formation of Mongolia and its paleobiological implications. Palaeogeography Palaeoclimatology Palaeoecology 533, 109239 (2019).
Kukihara, R. & Lockley, M. G. Fossil footprints from the Dakota Group (Cretaceous) John Martin Reservoir, Bent County, Colorado: new insights into the paleoecology of the dinosaur freeway. Cretaceous Research 33, 165–182 (2012).
Kubo, T. In quest of the Pteraichnus trackmaker: comparisons to modern crocodilians. Acta Palaeontologica Polonica 53, 405–412 (2008).
Lockley, M. G. Tracks and traces: new perspectives on dinosaurian behavior, ecology and biogeography in The Age of Dinosaurs (eds. Padian, K. & Chure, D. J.) 134–145 (Short courses in Paleontology #2, Paleontological Society; Knoxville, Tennessee, 1989).
Ostrom, J. H. Were some dinosaurs gregarious. Palaeogeography Palaeoclimatology Palaeoecology 11, 287–311 (1972).
Tompson, W. R. Field method for estimating the overall length of the crocodile. Journal of Southern Africa Wildlife Management Association 2, 27 (1972).
Hwang, K. G., Huh, M., Lockley, M. G., Unwin, D. M. & Wright, J. L. New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, S. W. Korea. Geological Magazine 139, 421–435 (2002).
McCrea, R. T., Pemberton, S. G. & Currie, P. J. New ichnotaxa of mammal and reptile tracks from the Upper Paleocene of Alberta. Ichnos 11, 323–339 (2004).
Eriksen, B. R. Crocodile and arthropod tracks from the Late Paleocene Wannagan Creek Fauna of North Dakota. Ichnos 12, 303–308 (2005).
Rajkumar, H. S., Mustoe, G. E., Khaidem, K. S. & Soiban, I. Crocodylian tracks from the Lower Oligocene flysch deposits of the Barail Group, Manipur, India. Ichnos 22, 122–131 (2015).
Falkingham, P. L., Milan, J. & Manning, P. L. A crocodylian trace from the Lance Formation (Upper Cretaceous) of Wyoming. New Mexico Museum of Natural History and Science Bulletin 51, 171–174 (2010).
Lockley, M. G. et al. New tetrapod ichnotaxa from the Blackhawk Formation (Upper Cretaceous), Utah. New Mexico Museum of Natural History and Science, Bulletin 79, 469–480 (2018).
Simpson, E. L. et al. A crocodylomorph track in the Upper Cretaceous capping sandstone member of the Wahweap Formation, Grand Staircase-Escalante National Monument, Utah, USA. New Mexico Museum of Natural History and Science Bulletin 51, 165–170 (2010).
McCrea, R. T. et al. A review of vertebrate track-bearing formations from the Mesozoic and earliest Cenozoic of western Canada with a description of a new theropod ichnospecies and reassignment of an avian ichnogenus. New Mexico Museum of Natural History and Science Bulletin 62, 5–93 (2014).
Mehl, M. G. Additions to the vertebrate record of the Dakota Sandstone. American Journal of Science 21, 441–452 (1931).
Avanzini, M. et al. Crocodylomorph tracks from the late Jurassic of Asturias (Spain). Ichnos 14, 143–153 (2007).
Abbassi, N., D’Orazi, P. S., Wagensommer, A., Dahnavi, M. G. Dinosaur and crocodylomorph footprints from the Hojhedk Formation (Bajocian, Middle Jurassic of North Kerman, central Iran. Italian Journal of Geoscience 134, 86–94 (2015).
Young, C. C. Note on some fossil footprints in China. Bulletin of the Geological Society of China 13, 151–154 (1943).
Lockley, M. G., Cart, K., Foster, J. & Lucas, S. G. Early Jurassic Batrachopus-rich track assemblages from interdune deposits in the Wingate Sandstone, Dolores Valley, Colorado, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 491, 185–195 (2018).
Acknowledgements
Authors would like to express our gratitude to volunteers who participated as intern researchers during the excavation of tracksite. We also thank Dr. Lorenzo Marchetti (Urweltmuseum, Thallichtenberg, Germany) and an anonymous reviewer, as well as Dr. J. O. Farlow (Indiana University) for helpful reviews of previous versions of this manuscript.
Author information
Authors and Affiliations
Contributions
K.-S.B., M.G.L. J.-D.L. and S.-M.B. all engaged in on-site field work, collecting, measuring and photographing of tracks. K.-S.B. and M.G.L. coordinated manuscript preparation and illustration design. J.-D.L. and S.-M.B. assisted with literature search and specimen cataloging. A.R. created 3D images and assisted with illustration design.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, K.S., Lockley, M.G., Lim, J.D. et al. Trackway evidence for large bipedal crocodylomorphs from the Cretaceous of Korea. Sci Rep 10, 8680 (2020). https://doi.org/10.1038/s41598-020-66008-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-66008-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
