
Abstract
Defective KLOTHO gene expression in mice led to a syndrome resembling human ageing. This study evaluated three KLOTHO polymorphisms, namely G395A, C1818T, and C370S, in an elderly population (mean age of 73 years) and their associations with ageing-related outcomes (cardiovascular events, kidney function, osteoporosis, sarcopenia) and mortality. Estimated glomerular filtration rates (eGFR) was lower in subjects with 1818TT (P = 0.047) and 370SS (P = 0.046) genotypes. The 1818TT genotype (P = 0.006) and 1818T allele were associated with higher frequency of myocardial infarction (MI) (CC:1.7% vs. CT + TT:7.0%; P = 0.002). The 370SS genotype was associated with lower stroke frequency (P = 0.001). MI (OR 3.35 [95% CI: 1.29–8.74]) and stroke (OR 3.64 [95% CI: 1.48–8.97]) were associated with mortality. Regarding MI, logistic regression showed 1818T allele was a risk factor for death-related MI (OR 4.29 [95% CI: 1.60–11.52]; P = 0.003), while 370C was protective (OR 0.03 [95% CI: 0.01–0.08]; P < 0.001). Regarding stroke, the 395A and 370C alleles were protective factors (respectively: OR 0.28 [95% CI: 0.20–0.80]; P = 0.018; OR 0.10 [95% CI: 0.05–0.18]; P < 0.001). This is the first study to determine potential associations between common ageing-related outcomes/mortality and KLOTHO polymorphisms. The 1818T allele was a risk factor for MI-related death. The 395A and 370C alleles were protective factors for stroke-related death in elderly from community.
Similar content being viewed by others

Introduction
The percentage of elderly people is increasing more than any other age group worldwide1. This trend comes with significant social consequences: communities with higher percentages of elderly people present higher disease rates (especially of chronic, noncommunicable diseases) and increased dependency of residential and hospital care1. Under these circumstances, several studies directed to basic health fields decided to look for a deeper comprehension of human aging to subsequently focus on intervention strategies2,3.
The KLOTHO gene was discovered in 1997, when a study revealed that its defective expression in mice led to a syndrome that resembles human ageing, including a short lifespan, arteriosclerosis, and osteoporosis4. Its products exist as a secreted protein5 or a transmembrane protein expressed predominantly in the distal renal tubules, choroid plexus, and pituitary gland4. Their function in humans, however, remains unknown. The secreted form suppresses oxidative stress and growth factor signalling, including insulin/IGF-1 signalling, all effects associated with longer life span6.
At least 10 KLOTHO mutations and single-nucleotide polymorphisms (SNPs) have been described in the humans. Nonetheless, the effects of each SNP in human ageing are still unclear. In fact, studies in human populations have sought for associations between aging, longevity, and some of such variants, including the most common SNPs G395A and C1818T7 and the functional Klotho variant KL-VS. This last variant comprises six polymorphisms in linkage disequilibrium, two of which resulting in amino acid substitutions: F352V and C370S6. Associations between these variants and age-related outcomes were demonstrated in different populations, mainly in Asians8,9.
There is only one publication regarding KLOTHO polymorphisms and age-related comorbidities in a Brazilian population10. Hence, the present study seeks to evaluate three KLOTHO gene polymorphisms in an elderly population from the São Paulo Ageing & Health Study (SPAH) and their potential association mainly with mortality, but also with the prevalence of major ageing-related outcomes, including osteoporosis, osteoporotic fractures (clinical and vertebral fractures), sarcopenia, kidney function (estimated glomerular filtration rate [eGFR]), and cardiovascular events (i.e. myocardial infarction and stroke).
Results
The demographic, anthropometric and clinical data of the 601 subjects are shown in Table 1, including genotype frequencies. The mean population age was 73.1 years. Body mass index varied from 17.2 to 47.4, with a mean of 28.1 kg/m².
Cardiovascular events
Of the 601 subjects, 80 (13.3%) experienced cardiovascular events, including angina pectoris, myocardial infarction or stroke.
Only 27 (4.5%) subjects had a history of MI. The analysis of the C1818T SNP revealed that the 1818TT genotype was associated with a higher frequency of MI (P = 0.006) (Table 2). Single-allele frequency analysis also showed association between 1818T and higher MI frequency (CC 1.7% vs. CT + TT 7.0%; P = 0.002). G395A and C370S had no statistically significant association with MI.
Thirty (5%) stroke events occurred in the studied population. The 370SS genotype was associated with a lower outcome frequency (P = 0.001) while single-allele analysis detected no significant differences (Table 2).
Aortic Calcification
Three hundred and forty-six (57.6%) subjects presented with abdominal aortic calcification score >0 visible on radiographic assessment. However, none of the three polymorphisms were associated with this outcome (Table 2).
Estimated glomerular filtration rate (eGFR)
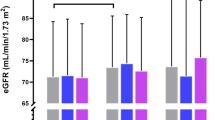
Subjects with the 1818TT and 370SS genotypes featured lower eGFR (P = 0.047 and P = 0.046, respectively) (Table 3). Furthermore, the presence of 1818C allele was associated with higher eGFR (TT: 55.5 vs. CC + CT: 58.1; P = 0.033). Single-allele frequency analysis directed to the C370S and G395A SNPs, however, detected no significant associations.
Mortality
From 2005 to 2012, there were 51 (8.5%) deaths among the 601 participants, mostly related to cardiovascular events (15 [29.4%, 9 from stroke and 6 from MI]), followed by pulmonary non-neoplastic causes (COPD, pneumonia, fibrosis) (10, 19.6%), non-ischemic heart failure (8, 15.7%), infections (excluding pneumonia) (7, 13.7%), cancer (4, 7.8%), and other causes, including pulmonary thromboembolism, Alzheimer’s Disease and kidney failure (7, 13.7%). The mean age of the deceased was 79.7 years, with a standard deviation of 6.8 years.
MI and stroke were the only variables associated with mortality. They were therefore analysed by logistic regression, which was adjusted by sex, BMI and age considering the average life expectancy of the Brazilian population during the data collection period. No association was observed concerning other outcomes, including eGFR.
MI was independently associated with death, with an Odds Ratio (OR) of 3.35 [95%CI: 1.29–8.74]). The presence of 1818T was a risk factor for MI-related death OR: 4.29 [95%CI: 1.60–11.52]; P = 0.003), whereas both the presence of 370C and the female sex were independent protective factors (respectively: OR: 0.4 [95%CI: 0.2–1.0], P = 0.041; and OR: 0.03 [95% CI: 0.01–0.08]; P < 0.001) (Table 4).
Stroke was also related to death (OR 3.64 [95%CI: 1.48–8.97]), and multiple logistic regression found that 395A and 370C alleles were protective factors (respectively: OR 0.28 [95%CI: 0.20–0.80]; P = 0.018, and OR 0.10 [95%CI: 0.05–0.18]; P < 0.001) (Table 4).
Other outcomes
No statistically significant associations between C1818T, G395A or C370S and lifestyle, including smoking history, alcohol intake and physical activity were found. Moreover, no polymorphism was independently associated with osteopenia/osteoporosis, clinical and vertebral fractures, and low appendicular muscle mass. (Supplementary Tables S1 and S2).
Discussion
To the best of our knowledge, this is the first study to investigate potential associations between the most common ageing-related outcomes and mortality with KLOTHO polymorphisms, namely C1818T, G395A, and C370S, in community-dwelling subjects.
In this community population, cardiovascular events were the main cause of death, and, as expected, the female sex was a protective factor for MI-related death. The 1818T allele was associated with a higher myocardial infarction frequency and, according to the logistic regression analysis, constitutes a risk factor for MI-related death. Most studies analysed this polymorphism regarding cardiovascular disease using coronary angiography8,11,12,13,14, myocardial scintigraphy15 or coronary artery calcification16, mainly in Asian population8,13,14,17,18. Rhee et al. showed that individuals that harbour the 1818T allele are associated with a lower prevalence of coronary artery disease (CAD) than those with the CC genotype, particularly in younger Korean subjects (<60 years)8. Analysis of an Iranian population, in turn, revealed a protective effect of the 1818CC genotype for age-related occurrence of CAD combined with hypertension in the subgroup older than 57 years12. The only study conducted in a Brazilian population so far, however, did not analyse the C1818T polymorphism in the mentioned setting10.
The other two KLOTHO polymorphisms analysed (G395A and C370S) were not associated with MI. Regarding G395A SNP, previous studies showed an association with CAD mainly in Asian population13,14,17,19, and a Spanish study20 found that it was a predictor of diabetes. On the other hand, regarding C370S, studies from Massachusetts, USA revealed no association between C370S and cardiovascular disease (premature acute coronary artery disease and vascular calcification) in young11 and older patients16. Moreover, there are other evidences tying Klotho to cardiovascular events, even though not related to its polymorphisms, but to its serum and tissue expressions. In fact, a Brazilian community-dwelling elderly population showed association between serum Klotho levels and history of myocardial infarction10. A Spanish study found association between the vascular levels of Klotho expression and presence of cardiovascular disease and cardiovascular risk factors. Indeed, individuals with coronary artery disease had a significantly lower expression of Klotho in aortic samples20.
In the present study, stroke was an important cause of cardiovascular death. The multivariate analysis indicated that the S allele of the C370S SNP and the A allele of the G395A SNP were protective factors for stroke-related death, whereas sedentary behaviour was identified as a risk factor for this outcome. Prior data have demonstrated that homozygous 370CC or 370SS individuals have a higher risk for stroke, while heterozygotes display a lower risk21,22. The C1818T SNP was also studied in Indian population, but was not associated with ischemic stroke21. Furthermore, in Korean women, the G395A SNP A allele was related to cardioembolic stroke23.
The present study found no associations between vascular calcification and Klotho polymorphisms in our community-dwelling elderly subjects. Of note, this issue had never been investigated in humans.
Renal dysfunction has been related to Klotho protein, but there are few studies correlating kidney function to KLOTHO polymorphisms in subjects without chronic kidney disease. Our results showed that 1818TT and 370CC genotypes were associated with lower eGFR. Analysis of two Chinese populations (Uygur and Kazak) with different longevities revealed that such genotypes were associated with higher serum creatinine in the Kazak population, which has shorter longevity5.
No association between Klotho polymorphisms and BMD and fractures were found in the present study. In contrast, data from literature have speculated that Klotho may have an effect on osteocytes24,25 and osteoblasts26, in addition to promoting osteoclast differentiation24. Data from previous studies are controversial, showing that the CS/SS genotypes (C370S SNP) were associated with higher BMD, but not with osteoporotic fractures in Spanish women27 and young/middle-aged Spanish men28. Moreover, postmenopausal white and Japanese women with the G395A SNP GG genotype tended to have lower BMD29, while A allele carriers among Japanese women had higher BMD9,30. The same association was found with T allele carriers of the C1818T SNP in postmenopausal women30, a finding not observed in another report9.
Potential associations between sarcopenia and KLOTHO polymorphisms had not been previously studied before, so the lack of such associations revealed by our study consists in negative but novel information.
Klotho polymorphisms have been previously associated with longevity (defined, depending on the study, as people over the age of 66, 75 or 93 years) in different populations6,31,32. Such findings prompted research about those variants and mortality itself, however only in chronically ill populations, such as patients in haemodialysis33,34 and IgA nephropathy35. Our study is the first one to examine KLOTHO SNPs and mortality in elderly community-dwelling subjects and two polymorphisms were associated with the main events that led to death, i.e., stroke and myocardial infarction.
Materials and Methods
Subjects
This study was based on data from the São Paulo Ageing & Health (SPAH) Study, which surveyed 65-year-old and older individuals living in a community in the Butantã district, located on the western zone of the city of São Paulo. The first survey took place from 2005 to 2007, and provided the baseline data36. The subjects were invited to participate in a second survey from 2010 to 2012, which provided data regarding mortality37. Full details pertaining to the study population, assessments, and procedures have been previously reported elsewhere36,37.
One hundred and twenty-five participants were interviewed on the first survey; 725 (70.73%) attended the second evaluation, and 601 of them (381 women and 220 men) had blood samples collected for DNA extraction and were included in the genotype-based analyses. Fifty-one of the analysed subjects (8.49%) died from 2005 to 2012.
This study was approved by the local ethics committee (Comissão de Ética para Análise de Projetos de Pesquisa do HCFMUSP [CAPPesq] #426/06), and all the participants provided written informed consent. All the methods were carried out in accordance with relevant guidelines and regulations stated by Declaration of Helsinki.
Data collection and assessments
Each subject was interviewed by a physician and responded to a standard questionnaire designed to gather information about lifestyle, health behaviours, and medical history, including age, smoking status, physical activity (ability to perform daily chores), alcohol consumption (at least 3 units per day), previous clinical fragility fractures, history of falls during the past year, and history of myocardial infarction or stroke.
Baseline measurements included height and weight, and were carried out using standard protocols to calculate the body mass index (BMI) and the appendicular skeletal muscle mass (ASM). Baseline laboratory and imaging evaluations included thoracic and lumbar spine radiographs, bone mineral density (BMD) measurement36,38, and serum creatinine concentration, determined using standard automated laboratory methods. Estimated glomerular filtration rate (eGFR) was calculated according to the Modification of Diet in Renal Disease (MDRD) formula39,40. The second interview provided data concerning mortality.
BMD measurement and Osteoporosis classification
Bone Mineral Density was measured using dual-energy X-ray absorptiometry (DXA, Hologic Inc. Bedford, MA, USA) at the lumbar spine, femoral neck, and total hip. All BMD measurements were performed by the same experienced technologist, following the recommendations by the International Society for Clinical Densitometry (ISCD)41. The least significant change was calculated with 95% confidence to be 0.033 g/cm2 for anteroposterior spine, 0.047 g/cm2 for femoral neck, and 0.039 g/cm2 for total hip. BMD characterization was performed according to the WHO classification: normal, T-score ≥ −1; osteopenia, T-score < −1 and > −2.5; and osteoporosis, T-score ≤ −2.5 in the spine, femoral neck, or total hip41.
Clinical osteoporotic fractures and assessment of vertebral fracture
Osteoporotic fracture was defined as a fracture occurring at sites characteristic of bone fragility (e.g., the rib, spine, forearm, humerus, and femur) on fifty-year-olds and older. Fractures occurring in the face, skull, ankles, elbows, and fingers were not considered osteoporotic fractures42.
Standard lateral thoracic and lumbar spine radiographs were taken using a 40-in tube-to-film distance centered at T7 and L2. All images provided good visibility of all vertebrae from T4 to L4, and vertebrae could be reliably identified. Two experienced readers independently performed the identification of vertebral fractures. Each of the T4–L4 vertebral images was evaluated to detect fractures, and nonvisible vertebrae were excluded. The agreement rate between the readers, which was established on a random subsample of 60 radiographs, was 96% with a kappa coefficient of 0.82. A consensus was reached between the readers for any difference of interpretation. Vertebral fractures were classified using the Genant semiquantitative approach43.
Appendicular skeletal muscle mass (ASM)
Appendicular skeletal muscle mass (ASM) was measured by DXA: ASM relative to height and total fat mass were assessed. Relative ASM was derived by adjusting for fat mass in addition to height44. Linear regression was used to determine the relationship between ASM relative to height (in meters) and fat mass (in kilograms). The residuals of the regression were used to identify those with ASM lower than the predicted value (given by an equation derived from the model), for an individual fat mass. A positive residual would indicate a relatively muscular individual, whereas negative values would indicate relatively sarcopenic individuals. In men, the equation resultant from the model was as follows: ASM (kg) = −20.67 + 22.48 × height (m) + 0.177 × fat mass (kg); and, in women: ASM (kg) = −14.51 + 17.27 × height (m) + 0.20 × fat mass (kg). The 20th percentile of the distribution of residuals was used as the cut point for sarcopenia according to ASM adjusted for fat. The cut-off corresponded to a residual of −2.06 in men45, and −1.45 in women46 in our Brazilian sample.
Assessment of abdominal aortic calcification
Lateral lumbar spine radiographs were acquired to quantify aortic calcification, besides assessing vertebral fractures. Calcium deposits were considered to be present if the densities were visible in an area that was parallel to the lumbar spine and anterior to the lower part of the spine. These calcific densities were graded on a 0 to 3 scale at each lumbar vertebrae segment: 0 denotes no aortic calcific deposits; 1 denotes small scattered calcific deposits that occupies less than one third of the longitudinal wall of the aorta; 2 indicates that one third or more but less than two thirds of the longitudinal wall of the aorta is calcified; and 3 indicates that two thirds or more of the longitudinal wall of the aorta is calcified47. This resulted in an abdominal aortic calcium score (AACS) ranging from 0 to 24. The reproducibility of AACS blind assessment was analysed using the Wilcoxon test for sum score as continuous and categorized data. In both cases, the P values were higher than 0.10 (P = 0.129 for the continuous data analysis and P = 0.99 for the categorized data analysis), confirming appropriate level of reproducibility.
Cardiovascular event assessment
During the interview, when a past history of myocardial infarction (MI) or stroke was identified, the participant’s medical records (completed by the family doctor or hospital) were reviewed. Stroke was defined as a sudden focal neurological deficit of presumed vascular origin that persisted for more than 24 hours. MI was defined as ischemic symptoms, development of diagnostic Q waves on the electrocardiogram, electrocardiographic changes indicative of ischaemia (ST segment elevation or depression), or coronary artery intervention.
Mortality data
Mortality data of the original SPAH cohort was gathered from the date of the baseline examination through December 31, 2012. They were collected from the publicly available death certificates ascertained by the Programa de Aprimoramento das Informações de Mortalidade no Município de São Paulo (PRO-AIM, Improvement Program of Information on Mortality in São Paulo). The underlying causes of death were coded according to the 10th revision of the International Classification of Diseases (ICD-10). Cardiovascular mortality was defined as ICD-10 codes I00 to I99 and non-cardiovascular mortality as all other causes of death.
Genotyping
Genomic DNA was isolated from peripheral blood leukocytes using the salting-out methodology48, and was stored at −70 °C prior to analysis. The TaqMan allelic discrimination method was employed for genotyping, using specific probes labelled with VIC and FAM (Applied Biosystems, Foster City, CA, USA).
The analysed SNPs were chosen based on previous literature data that suggested possible associations with ageing and functional significance5,7,8,9,49,50. Such SNPs comprised C1818T (rs564481), a silent mutation30 located in exon 4 of KLOTHO7,30; G395A (rs1207568), located in the promoter region of KLOTHO7,30 and possibly associated with its expression and function30; and C370S (also named G1117C; rs9527025), located in exon 251. C370S is one of the SNPs that compose the functional Klotho variant KL-VS, which impairs the protein function by influencing its trafficking and catalytic activity51,52.
All SNPs were genotyped using commercial TaqMan assays (assay IDs: rs1207568: C_7604792_10; rs9527025: C_2983036_20; rs564481: C_592739_10) with TaqMan Genotyping Master Mix and were measured using a StepOne plus detection system (Applied Biosystems). Cycling conditions included a denaturation step at 95 °C for 10 minutes followed by 40 cycles of denaturation at 92 °C for 15 seconds and annealing for 1 minute at 60 °C. To assess the genotyping quality, the polymorphisms were analysed twice in 5% of all samples, yielding a 98% concordance.
Rationale for outcome selection
Following the evidence-based choice for studying each of these three KLOTHO SNPs, we selected the most clinically relevant ageing-related outcomes associated to them. As discussed above, there lacks data correlating mortality to KLOTHO SNPs, thus mortality is a hard outcome that was chosen to be one of this studies cornerstones. Potential causes for death, as cardiovascular events, and outcomes related to healthy ageing that were previously studied in relation to these and other KLOTHO SNPs (CKD, BMD, etc) were chosen according to previous literature, also debated on Discussion.
Statistical analysis
Categorical data were expressed as absolute frequency (n) and relative frequency and analysed by Pearson χ2. Continuous variables were expressed as mean ± standard deviation for parametric data or median and quartiles for non-parametric data. When comparing three or more groups, nonparametric data were analysed using the Kruskal-Wallis test adjusted by the Bonferroni post-test, while the Mann-Whitney test was applied for comparisons between two groups. In the multivariate analysis, Logistic Regression, results were expressed as odds ratio and 95% confidence interval. Hosmer–Lemeshow goodness-of-fit test was applied to evaluate the calibration of logistic regression model and to check the importance of the discrepancy between observed and expected for Myocardial Infarction and Stroke. Forward stepwise selection procedure was used to select the variables significantly related to Myocardial Infarction and Stroke, as assessed by the Likelihood Ratio test. Odds ratios with 95% confidence intervals were calculated for statistically significant variables. A P-value <0.05 was considered statistically significant. The genotype distribution and allele frequencies of the C1818T and G395A SNPs were consistent with the Hardy-Weinberg equilibrium (χ²= 0.16 with P = 0.68, and χ² = 3.49 with P = 0.06, respectively), whereas the ones associated with C370S were not (χ² = 3.90 with P = 0.05). However, since the number of subjects that would have to be excluded from the analysis to adjust it was higher than 5 to 10%, all the data were analysed to avoid additional bias53. Furthermore, for the allelic analyses, the number of patients was not duplicated as not to create bias from the duplication of phenotypic traits.
References
World Population Prospects: The 2015 Revision. (2015).
Shetty, A. K., Kodali, M., Upadhya, R. & Madhu, L. N. Emerging Anti-Aging Strategies - Scientific Basis and Efficacy. Aging. Dis. 9, 1165–1184 (2018).
Eline Slagboom, P., van den Berg, N. & Deelen, J. Phenome and genome based studies into human ageing and longevity: An overview. Biochim. Biophys. Acta - Mol. Basis Dis. 1864, 2742–2751 (2018).
Kuro-o, M. et al. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature 390, 45–51 (1997).
Xu, X., Liang, X., Hu, G., Zhang, J. & Lei, H. Renal function and klotho gene polymorphisms among Uygur and Kazak populations in Xinjiang, China. Med. Sci. Monit. 21, 44–51 (2015).
Di Bona, D., Accardi, G., Virruso, C., Candore, G. & Caruso, C. Association of Klotho polymorphisms with healthy aging: a systematic review and meta-analysis. Rejuvenation Res. 17, 212–6 (2014).
Shimoyama, Y. et al. KLOTHO gene polymorphisms G-395A and C1818T are associated with low-density lipoprotein cholesterol and uric acid in Japanese hemodialysis patients. Am. J. Nephrol. 30, 383–388 (2009).
Rhee, E. J. et al. The differential effects of age on the association of KLOTHO gene polymorphisms with coronary artery disease. Metabolism. 55, 1344–1351 (2006).
Shimoyama, Y., Nishio, K., Hamajima, N. & Niwa, T. KLOTHO gene polymorphisms G-395A and C1818T are associated with lipid and glucose metabolism, bone mineral density and systolic blood pressure in Japanese healthy subjects. Clin. Chim. Acta 406, 134–138 (2009).
Paula, R. et al. Serum Klotho (but not haplotypes) associate with the post-myocardial infarction status of older adults. Clinics 71, 725–732 (2016).
Low, A. F. et al. Aging syndrome genes and premature coronary artery disease. BMC Med. Genet. 6, 38 (2005).
Akbari, H. et al. Association of Klotho gene polymorphism with hypertension and coronary artery disease in an Iranian population. BMC Cardiovasc. Disord. 18, 237 (2018).
Jo, S.-H. et al. KLOTHO gene polymorphism is associated with coronary artery stenosis but not with coronary calcification in a Korean population. Int. Heart J. 50, 23–32 (2009).
Imamura, A. et al. Klotho gene polymorphism may be a genetic risk factor for atherosclerotic coronary artery disease but not for vasospastic angina in Japanese. Clin. Chim. Acta 371, 66–70 (2006).
Arking, D. E. et al. KLOTHO Allele Status and the Risk of Early-Onset Occult Coronary Artery Disease. Am. J. Hum. Genet 72 (2003).
Tangri, N., Alam, A., Wooten, E. C. & Huggins, G. S. Lack of association of Klotho gene variants with valvular and vascular calcification in Caucasians: a candidate gene study of the Framingham Offspring Cohort. Nephrol. Dial. Transplant. 26, 3998–4002 (2011).
You, N. & Liu, X. L. Tian Xiao Chun & Ma, H. X. Correlation between single nucleotide polymorphisms at three sites of Klotho gene with coronary heart disease. J. Chongqing Med. Univ. 37, 579–584 (2012).
He, G.-P. et al. KLOTHO gene promoter G-395A polymorphisms of acute coronary syndrome in the Chinese Han population of Suwan region. Nat. Sci. (2010).
Zhai, C. et al. Review Article Polymorphism of klotho G-395A and susceptibility of coronary artery disease in East-Asia population: a meta-analysis. Int. J. Clin. Exp. Med. 8 (2015).
Donate-Correa, J. et al. Influence of Klotho gene polymorphisms on vascular gene expression and its relationship to cardiovascular disease. J. Cell. Mol. Med. 20, 128–133 (2016).
Majumdar, V., Nagaraja, D. & Christopher, R. Association of the functional KL-VS variant of Klotho gene with early-onset ischemic stroke. Biochem. Biophys. Res. Commun. 403, 412–416 (2010).
Arking, D. E., Atzmon, G., Arking, A., Barzilai, N. & Dietz, H. C. Association between a functional variant of the KLOTHO gene and high-density lipoprotein cholesterol, blood pressure, stroke, and longevity. Circ. Res. 96, 412–8 (2005).
Kim, Y. et al. Klotho is a genetic risk factor for ischemic stroke caused by cardioembolism in Korean females. Neurosci. Lett. 189–194 (2006).
Kawaguchi, H. et al. Independent impairment of osteoblast and osteoclast differentiation in klotho mouse exhibiting low-turnover osteopenia. J. Clin. Invest. 104, 229–237 (1999).
Kawaguchi, H., Manabe, N., Chikuda, H., Nakamura, K. & Kuroo, M. Cellular and molecular mechanism of low-turnover osteopenia in the klotho-deficient mouse. Cell. Mol. Life Sci. 57, 731–7 (2000).
Suzuki, H. et al. Histological evidence of the altered distribution of osteocytes and bone matrix synthesis in klotho-deficient mice. Arch. Histol. Cytol. 68, 371–81 (2005).
Riancho, J. A. et al. Association of the F352V variant of the Klotho gene with bone mineral density. Biogerontology 121–127, https://doi.org/10.1007/s10522-006-9039-5 (2007).
Zarrabeitia, M. T. et al. Klotho Gene Polymorphism and Male Bone Mass. Calcif. Tissue Int. 80, 10–14 (2007).
Yamada, Y., Ando, F., Niino, N. & Shimokata, H. Association of polymorphisms of the androgen receptor and klotho genes with bone mineral density in Japanese women. J. Mol. Med. 83, 50–57 (2005).
Kawano, K. et al. Klotho gene polymorphisms associated with bone density of aged postmenopausal women. J. Bone Miner. Res. 17, 1744–1751 (2002).
Zhu, Z. et al. Klotho gene polymorphisms are associated with healthy aging and longevity: Evidence from a meta-analysis. Mech. Ageing Dev. 178, 33–40 (2019).
Revelas, M. et al. Review and meta-analysis of genetic polymorphisms associated with exceptional human longevity. Mech. Ageing Dev. 175, 24–34 (2018).
Ko, G. J. et al. The association of Klotho gene polymorphism with the mortality of patients on maintenance dialysis. Clin. Nephrol. 80, 263–269 (2013).
Friedman, D. J. et al. Klotho Variants and Chronic Hemodialysis Mortality. J. Bone Miner. Res. 24, 1847–1855 (2009).
Ko, G. J. et al. The Association of Klotho Polymorphism with Disease Progression and Mortality in IgA Nephropathy. Kidney Blood Press. Res 36, 191–199 (2012).
Lopes, J. B. et al. Prevalence and risk factors of radiographic vertebral fracture in Brazilian community-dwelling elderly. Osteoporos. Int. 22, 711–719 (2011).
Santana, F. M. et al. Association of Appendicular Lean Mass, and Subcutaneous and Visceral Adipose Tissue With Mortality in Older Brazilians: The São Paulo Ageing & Health Study. J. Bone Miner. Res. 34, jbmr.3710 (2019).
Lopes, J. B. et al. Osteoporotic Fractures in the Brazilian Community-Dwelling Elderly: Prevalence and Risk Factors. J. Clin. Densitom. 14, 359–366 (2011).
Levey, A. S. et al. Using standardized serum creatinine values in the modification of diet in renal disease study equation for estimating glomerular filtration rate. Ann. Intern. Med. 145, 247–54 (2006).
Go, A. S., Chertow, G. M., Fan, D., McCulloch, C. E. & Hsu, C. Chronic kidney disease and the risks of death, cardiovascular events, and hospitalization. N. Engl. J. Med 351, 1296–305 (2004).
Binkley, N. et al. Official Positions of the International Society for Clinical Densitometry and Executive Summary of the 2005 Position Development Conference. J. Clin. Densitom. 9, 4–14 (2006).
Seeley, D. G. et al. Which fractures are associated with low appendicular bone mass in elderly women? The Study of Osteoporotic Fractures Research Group. Ann. Intern. Med. 115, 837–42 (1991).
Domiciano, D. S. et al. Vertebral Fracture Assessment by Dual X-Ray Absorptiometry: A Valid Tool to Detect Vertebral Fractures in Community-Dwelling Older Adults in a Population-Based Survey. Arthritis Care Res. (Hoboken) 65, 809–815 (2013).
Newman, A. B. et al. Sarcopenia: Alternative Definitions and Associations with Lower Extremity Function. J. Am. Geriatr. Soc 51, 1602–1609 (2003).
Figueiredo, C. P. et al. Prevalence of sarcopenia and associated risk factors by two diagnostic criteria in community-dwelling older men: the São Paulo Ageing & Health Study (SPAH). Osteoporos. Int. 25, 589–596 (2014).
Domiciano, D. S. et al. Discriminating sarcopenia in community-dwelling older women with high frequency of overweight/obesity: the São Paulo Ageing & Health Study (SPAH). Osteoporos. Int. 24, 595–603 (2013).
Kauppila, L. I. et al. New indices to classify location, severity and progression of calcific lesions in the abdominal aorta: a 25-year follow-up study. Atherosclerosis 132, 245–50 (1997).
Bonfá, A. C., Seguro, L. P. C., Caparbo, V., Bonfá, E. & Pereira, R. M. R. RANKL and OPG gene polymorphisms: associations with vertebral fractures and bone mineral density in premenopausal systemic lupus erythematosus. Osteoporos. Int. 26, 1563–71 (2015).
Riancho, J. A. Role of the Klotho Gene in Bone and Mineral Metabolism. Clin. Rev. Bone Miner. Metab 6, 31–36 (2008).
Ogata, N. et al. Association of klotho gene polymorphism with bone density and spondylosis of the lumbar spine in postmenopausal women. Bone 31, 37–42 (2002).
Arking, D. E. et al. Association of human aging with a functional variant of klotho. Proc. Natl. Acad. Sci 99, 856–861 (2002).
Xuan, N. T. & Hai, N. Van. Changes in expression of klotho affect physiological processes, diseases, and cancer. Iran. J. Basic Med. Sci. 21, 3–8 (2018).
Dong, Y. & Peng, C. Y. J. Principled missing data methods for researchers. Springerplus 2, 1–17 (2013).
Acknowledgements
This study was not sponsored by any pharmaceutical company. The SPAH project was supported by grants from the Fundacao de Amparo e Pesquisa do Estado de Sao Paulo (FAPESP #03/09313‐0, #04/12694‐8, and #09/15346‐4; FAPESP #11/00411‐5 to DSD); Conselho Nacional de Ciencia e Tecnologia (CNPQ#300559/2009‐7 and #301805/2013‐0 to RMRP), Federico Foundation (to RMRP), and Coordenacao de Aperfeicoamento de Pessoal de Nivel Superior (CAPES to DSD, and CPF). We further cordially thank Jackeline Couto Alvarenga for collecting the blood samples from the subjects; Vera Lucia Barbosa and Maria de Lourdes Floriano for the recruitment of study subjects, including door‐to‐door visits; and Maria Luiza Santilli for assistance in data entry.
Author information
Authors and Affiliations
Contributions
Conception and design of the study: R.M.R.P., P.R.M. Study Conduct: R.M.R.P., V.F.C., L.T., D.S.D., L.G.M., C.P.F. Data Collection: D.S.D., L.G.M., C.P.F., P.R.M. Data Analysis: R.M.R.P., T.Q.F., A.S.F., L.F.O., I.C. Data Interpretation: R.M.R.P., T.Q.F., A.S.F., L.F.O., I.C. Drafting Manuscript: R.M.R.P., T.Q.F., A.S.F. Revising Manuscript: R.M.R.P., T.Q.F., A.S.F., L.F.O., I.C. Approving Final Version of Manuscript: R.M.R.P., T.Q.F., A.S.F., L.T., V.F.C., D.S.D., L.G.M., C.P.F., P.R.M., L.F.O., I.C.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pereira, R.M.R., Freitas, T.Q., Franco, A.S. et al. KLOTHO polymorphisms and age-related outcomes in community-dwelling older subjects: The São Paulo Ageing & Health (SPAH) Study. Sci Rep 10, 8574 (2020). https://doi.org/10.1038/s41598-020-65441-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-65441-y
This article is cited by
-
Role of Klotho Protein in Neuropsychiatric Disorders: A Narrative Review
Indian Journal of Clinical Biochemistry (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
