
Abstract
For decades, taphonomists have dedicated their efforts to assessing the nature of the massive leporid accumulations recovered at archaeological sites in the northwestern Mediterranean region. Their interest lying in the fact that the European rabbit constituted a critical part of human subsistence during the late Pleistocene and early Holocene. However, rabbits are also a key prey in the food webs of Mediterranean ecosystems and the base of the diet for several specialist predators, including the Iberian lynx (Lynx pardinus). For this reason, the origin of rabbit accumulations in northwestern Mediterranean sites has proved a veritable conundrum. Here, we present the zooarchaeological and taphonomic study of more than 3000 faunal and 140 coprolite remains recovered in layer IIIa of Cova del Gegant (Catalonia, Spain). Our analysis indicates that this layer served primarily as a den for the Iberian lynx. The lynxes modified and accumulated rabbit remains and also died at the site creating an accumulation dominated by the two taxa. However, other agents and processes, including human, intervened in the final configuration of the assemblage. Our study contributes to characterizing the Iberian lynx fossil accumulation differentiating between the faunal assemblages accumulated by lynxes and hominins.
Similar content being viewed by others

Introduction
The Iberian lynx (Lynx pardinus) is a felid species that used to be present throughout the Iberian Peninsula and southern France. Over the last century, the population declined and today it is to be found in only very limited areas of the Iberian Peninsula, primarily in the south1. The main prey of the Iberian lynx is the European rabbit (Oryctolagus cuniculus), a diet that is supplemented by birds, such as the red-legged partridge (Alectoris rufa) and ducks (Anseriformes), small mammals, such as rodents, and sporadically by wild small ungulates, such as red deer fawns or young fallow deer2. However, actualistic studies of lynx food habits could be conditioned by the intensive anthropic pressure on the landscape, which means the food preferences of the lynx in the past are not readily evaluated. Insights should usefully be gained from the study of Pleistocene contexts characterised by low human pressure, such as that prevailing at the Cova del Gegant site.
Accumulations of rabbit remains, the main prey of lynx, are very common at archaeological sites. For decades, archaeologists have dedicated their efforts to assessing the nature of these massive leporid accumulations at prehistoric sites in the northwestern Mediterranean (Iberia and Southern France). This field of study is of particular importance for prehistorians given that the rabbit also constituted a critical part of human subsistence during the Upper Palaeolithic and Mesolithic3,4,5,6,7. Indeed, some researchers have proposed that the consumption of fast, small game (especially leporids, but also birds) was more common prior to the late Palaeolithic than previously thought and that archaic hominins from the northwestern Mediterranean, as early as Marine Isotope Stage (MIS) 13, had broader diets than those from adjacent regions8. However, the origin of rabbit accumulations at Iberian archaeological sites has proved a veritable conundrum.
The fact that the European rabbit (Oryctolagus cuniculus) is a key mammal prey species in the food webs of Mediterranean ecosystems9 greatly hinders attempts to unravel the taphonomic history of many assemblages. Indeed, most Iberian predators consume rabbit as a major part of their diets, being the main prey for almost 30 raptors and mammalian carnivores in Iberia10. Certain Iberian endemic predators, such as the Iberian lynx (Lynx pardinus) and the Iberian imperial eagle (Aquila adalberti), are even hyper-specialists, with rabbit constituting 100% of their diet for much of the seasonal cycle2,11. This means that all these rabbit predators are potential taphonomic agents and bone accumulators of assemblages dominated by their prey. At the same, however, although the European rabbit is highly adaptable, presenting considerable behavioural and ecological plasticity12,13, they often live in concentrations of dozens of individuals in large, deep communal warrens in which their populations are more prosperous and their breeding success greater than in shallow or small breeding stops14. As result, in large warrens natural dead are originators of large natural accumulations of skeletal remains of these animalse.g. 15.
Faced with such a variety of possible modifiers and accumulators of rabbit remains, neo-taphonomic studies have focused on characterizing the taphonomic signals of different rabbit predators16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33. Based on anatomical representations, percentage of adults, the fragmentation of bones and modifications on bone surfaces, great progress has been made. And, albeit to a lesser extent, some natural accumulations have also been characterized (based on the recovery of remains from rabbit warrens), in attempts at characterizing intrusive accumulations15. However, the results are often frustrating due to the ambiguity of some proxies (e.g. age of leporids)23 and, especially, because most Pleistocene assemblages are cumulative palimpsests in which several agents and processes are usually involved, including abundant natural intrusions8,34,35,36,37,38,39,40.
In this paper, we present a multiproxy study of the faunal assemblage from layer IIIa at the Cova del Gegant site (Catalonia, Spain) dated to MIS 3. We examined all macro-mammal, meso-vertebrate and coprolite remains in terms of their archaeological taphonomy and performed a multivariate statistical analysis. Our results provide new and valuable information for characterizing the leporid accumulations of the Iberian lynx (Lynx pardinus), an omnipresent carnivore in the Iberian faunal assemblages of the Pleistocene. Moreover, our study contributes to differentiating faunal assemblages accumulated by lynxes and hominins with a particular focus on rabbits.
Site stratigraphy
Cova del Gegant (Sitges, Barcelona) is a cave located on the seaward edge of the Garraf massif in the central part of the Catalan Coastal Range (1°46′27.33′′E, 41°13′24.75′′ N). The site consists of a principal chamber (GP), partially flooded by the sea and eroded by the ongoing coastal erosion, and an inner area (GP2 and GL-T), whose preserved sedimentation was the target of current fieldwork (Fig. 1).

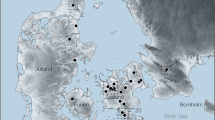
Location and map of Cova del Gegant site. (a–c) Location of Cova del Gegant in south-western Europe, NE of Iberian Peninsula and the Garraf massif. Data extracted from Map Tile 4_8–5 (CC BY-SA). OpenStreetMap© licensed under ODdL 1.0 (https://www.openstreetmap.org/copyright) by the OpenStreetMap Foundation (OSMF). ©OpenStreetMap contributors (https://www.openstreetmap.org/). (d) Panoramic view of Cova del Gegant and Punta de les coves (Photo by M. Sanz & J. Daura). (e, f) Site plan of Cova del Gegant indicating the position of the different galleries discussed in the text and the location of layer III.
At least eight site formation episodes, formed by several stratigraphic layers, from the Upper Pleistocene (Episodes 0–3) to the Holocene (Episodes 4–7) have been recognized in the Cova del Gegant stratigraphic sequence. Five Neandertal specimens have been identified41,42 and two of these fossils have been recovered in stratigraphic context (layer V), at the base of this sequence (GP2). The archaeological remains discussed in this study come from layer IIIa preserved at the back of the main gallery (GP2). This layer corresponds to Episode 3 and can be chronologically located in MIS 3, a period between 59 and 29 calendar (cal) kyr BP (ca. 56 to 2514C kyr BP). At this point in our study, the chronological framework of this layer is between ca. 31.1 and 30.5 ka cal. BP.
Layer IIIa corresponds to Episode 3a (Fig. 1). It is subjacent to layer II and is located in the inner area of the main gallery (GP2) and formed by light reddish-brown (5YR 6/4) sandy silt containing faunal remains and charcoals43. The layer is 10 to 20 cm thick and is located 100–120 cm below datum. Preliminary studies of this layer focused on the archaeological materials from the first excavation (GP2) (years 2007–2010), located between rows 22 and 2644.
According to this work, small carnivores were proposed as the principal agents involved in the formation of layer IIIa and that geological and diagenetic processes were primarily responsible for bone modification. No biological modifications were observed, although only a small proportion of the macromammals were examined. In addition, human activity, in the form of hearths, and bioturbation were identified as being responsible for the final configuration of the assemblage. These processes and agents have been chronologically sequenced: thus, the first event can be identified as “the use of the cave as a den/shelter by a small felid or canid, resulting in the accumulation of carnivore bones and coprolites. The second event was both a sporadic, and probably single, occupation of the cave by human groups that left behind the few burned remains and hearths found to date. The third event was the partial reworking of the sediments produced by carnivore activity, as suggested by fragments of coprolites observed in thin section, and by burrowing animals, as evidenced during the excavation. The mixing, and the following sedimentary diagenesis, produced the current patchy distribution presented by the uppermost layers in the combustion structure”44 p.112. Our latest study (reported here) examines this interpretation in greater depth and sheds new light on the taphonomic history of the site’s fossil accumulation. By including all the macro- and meso-vertebrate remains and all the coprolites in our taphonomic analysis, we can provide fresh information on the intervention of biological and natural processes.
Results
Archaeological-taphonomy
In this study, we analysed 3640 specimens (NSP). The good state of preservation of these fossil remains meant we were able to determine a high number of them. Indeed, 3608 specimens have been identified at the anatomical and taxonomic levels (NISP = 99.1%). The fossils belong to large and medium-sized ungulates, carnivores, leporids and birds (Table 1). Despite this apparent diversity, the faunal association is composed primarily of the remains of leporids (NISP = 84.1%), lynxes (NISP = 8.2%) and birds (NISP = 6.5%), these three taxonomic groups making up more than 98% of the NISP. Thirty-two specimens were classified according to size, the majority being categorized as small-sized mammals. Based on the NISP, these probably correspond in the main to bone fragments of small carnivores (Table 1). Simpson’s diversity index (D = 0.72) points to the predominance of one taxon (the leporids), while Shannon’s evenness index indicates an uneven distribution of the species represented (E = 0.24).
The estimate of the assemblage’s MNE is high at 2987, most belonging to the dominant taxa (MNE = 98.9%). Based on these elements, we calculate a minimum number of 89 individuals, distributed as follows leporids: 56, birds: 16 and lynxes: 6, and accounting for 87.64% of the total MNI.
Due to the differences in the respective methodologies employed to study macro-mammals and meso-vertebrates (leporids and birds in this work), we present the data in two different sub-sections.
Macro-mammals
The total number of macro-mammals remains (NSP) in our analysis is 358 (337 NISP). This includes all ungulate, carnivore and taxonomically indeterminable remains attributed to weight sizes that are not compatible with animals considered as meso-vertebrates in our study.
The anatomical representation of the macro-mammals in level IIIa of the sequence is heavily biased, with the except of the lynx that shows a high degree of integrity. Cranial elements include isolated teeth of horse and red deer, and a fragment of the horn core of an indeterminate bovid (Bos/Bison/Capra). Postcranial bones include one unfused distal tibia of an indeterminate small ungulate; one complete metacarpal and one capitatum-trapezoid of an adult red deer, probably corresponding to the same individual; one complete patella of an adult horse; and two fragments of trunk elements: the corpus of a thoracic vertebra and one rib fragment from an indeterminate medium-sized ungulate. Among the 32 remains classified by size, nine – in the main trunk specimens – are compatible with that of the ungulates represented. Ten elements have been estimated by association with the ungulate specimens identified, belonging to a minimum of four individuals (Table 1). At least two of these are young individuals (one horse and one deer) and another (deer) is a prime adult.
As in the case of the ungulates, the skeletal completeness of the carnivores is very low. Specimens of large canid (probably Canis lupus) and hyena correspond mainly to distal foot bones (tarsal, metapodials and phalanges). These bones correspond to one adult canid and two hyenas – one young and one a prime adult. One red fox adult individual is represented by an extraordinarily well-preserved skull, one fragment of hemi-mandible and one fragment of innominate. Ten specimens (NISP = MNE) from cranial, appendicular and axial post-cranial segments indicate the presence of two adult wild-cat individuals in the assemblage.
The c. 300 lynx specimens merit more detailed attention. The skeletal representation of lynx shows a high degree of completeness (Table 2 and Fig. 2). All skeletal elements are represented, the most abundant in NISP being phalanges, vertebrae, metacarpal and carpal bones. The 299 specimens identified have allowed us to estimate a minimum of 273 elements. Both the most and least represented elements are the same as for the NISP, which again is indicative of the completeness of the lynx skeletons. Based on the different degrees of development of the upper teeth, we estimate a minimum of six individuals. One of them is an infant, just a few weeks old; another a juvenile of less than a year; and four individuals correspond to prime adults (Table 2 and Fig. 2). Most of the specimens correspond to the adults or the young juvenile individual, while the lynx kitten is represented by just a few specimens. In relative terms (%ISU), the anatomical representation of lynxes shows high frequencies for the high-survival elements, including skulls, isolated teeth and the main limbs, and low frequencies for small bones, such as sesamoids and tarsals. Some of the low-survival elements, above all ribs, are poorly represented but vertebrae are relatively well preserved. The mean %ISU is 41, suggesting the survival of almost half the skeletons.

Skeletal reconstruction of Iberian lynx from different individuals. Example of the completeness of skeletal representation of Iberian lynx in the layer IIIa of Cova del Gegant.
Fragmentation of the macro-mammal remains is rare with 257 elements remaining complete (69% of MNE), including 24 of the main long bones of lynx (19), wild cat (4), large canid (1), and one red deer metapodial. In the specific case of the lynx, the percentage is higher (79.5%). Considering that the lynx represents 90% of the NISP of macro-mammals, the low rate of breakage of skeletal elements suggests very little post-mortem disturbance of the bones. During fieldwork, several anatomical connections were recorded among the lynx elements, including almost complete right forepaw (Fig. 2). Likewise, the two almost complete skulls and the anatomical refitting implemented among the fragments broken under dry conditions reinforce this observation.
Our analysis of long limb bone breakage included only a small number of specimens (20)–40% correspond to bones that preserve a 1/4 or 1/2 of the length of the diaphysis and the whole section and 60% correspond to small fragments i.e. a 1/4 or less of the length of the diaphysis and less than 1/3 of their section. Only eleven fracture outlines have been analysed. More than 63% present right angles, while 100% present irregular surfaces indicating dry post-depositional breakage. Fractures produced during the excavation process affect 24 specimens; diagenetic breakage 32 and only one fragment has been attributed to fresh (green) breakage generated by carnivores in association with a type A notch. Unfused bones (12) and bones with indeterminate fractures (9) complete the assemblage. According to the data presented, the scarce fracturing of the sub-set of macro-mammals can be said to have occurred when the bones were in a dry/fossilized state.
Meso-vertebrates
A total of 3271 specimens (NISP) correspond to mesofauna, primarily leporids and small birds. As noted above, leporids are the most abundant with 3035 identified remains (NISP) (Table 1). Based on left tibia records, a total of 56 individuals (MNI) has been estimated. The mortality pattern shows a predominance of adults > 9 months (MNI 36 or 63%), followed by young individuals (MNI 36%). Among the latter, at least six are perinatal, 13 infantile <3 months and one is juvenile of 3–5 months.
The skeletal elements have all been recovered in different percentages. Phalanges, metatarsus, isolated lower molars and vertebrae are the best represented anatomical elements in absolute terms, while patellae, scapulae, astragals and articular bones appear in smaller numbers. The MNE is 2492, dominated by phalanges (Table 2). The relative abundance (%RA) of the skeletons presents a predominance of tibiae, innominate, calcaneus, mandibles, femurs and metatarsus (up to 60% of the RA) and a lower presence of ribs, vertebrae, articular bones (carpal/tarsal) and metacarpal (less than 10% of the RA). The best represented element in relative terms is the tibia (Table 2).
The relative proportions of skeletal remains indicate a greater representation of the postcranial than of the cranial skeleton (Supplementary Table S1). However, if we evaluate the accuracy of this test, the evenness of proportions is less pronounced. Thus, the TA/MD index that considers two elements, ranks mineral density highly45, and reveals higher valance proportions in the cranial/postcranial representation (159.2), especially if we consider the MNE (TAE/MDE = 127.5). In the case of limb bones, the lower limbs (metapodials) are better represented than the intermediate limbs (250.4), which, in turn, are better represented than the upper limb bones (131.9). Finally, the forelimb is slightly better represented than the hindlimb (54.2) in the Cova del Gegant assemblage. This relationship becomes more pronounced as we proceed from proximal to distal elements (Supplementary Table S1).
A study of bone breakage shows that 55.6% of the skeletal remains of leporids are complete. However, there is no statistically significant correlation between the size of the complete element in comparative specimens and their completeness in the Cova del Gegant assemblage (rho = 0.1, p > 0.05). Yet, the largest elements in a complete skeleton, including the main long bones, skull and girdles, are usually broken while the shortest, including phalanges, metacarpal and articular bones, are usually complete. No complete ulnae, scapulae or ribs have been recovered. The size of the leporid remains ranges between 1.2 and 93.56 mm (mean = 16.03 mm), with more than 61% being greater than 10 mm in length. In the case of limb bones, the upper and intermediate (UILB) bones are highly fragmented with only 43 complete bones (7.8% of the UILB), represented primarily by femurs (39.53% of the complete UILB) and humerus (30.23% of the complete UILB), while the lower limbs (metacarpal and metatarsal) are mainly complete (71 and 53%, respectively). The fragmentation of long bones indicates the same frequencies for proximal (PE + PES) and distal (SDE + DE) parts (~30%). The breakage categories included in Supplementary Table S2 show that the cranium is represented mainly by NC and M fragments and the mandibles by MBI. Complete elements in skulls are more abundant among mandibles (9.71%) than they are in crania (1%). Innominate are represented by fragments that retain the acetabulum (AIS, AISIL, AIL), while scapulae are represented by GCN fragments. Ribs are mainly shaft (S) or shaft retaining epiphyseal fragments (PES). Articular bones, in situ teeth and isolated teeth are represented mainly by complete elements (Supplementary Table S2). Some anatomical connections have been recorded, such as tarsal with metatarsal bones and carpal with metacarpal elements.
The bone breakage study revealed a mixture of green and dry fractures with a predominance of the former. Sixty-two rich-marrow long limb bones (humerus, femur and tibia) retain the epiphysis plus a fragment of diaphysis presenting a helical or V-shaped fracture that can be classified as morphotypes of carnivore breakage20,29,45. They include 19 distal humerus, 27 distal tibiae and 2 distal and 14 proximal femurs. Although in Supplementary Table S1, 86 remains are classified as shaft (S), only the main long bones (humerus, femur and tibia) contain enough features to establish the state (dry or green) in which the fracture occurred. Thus, only one diaphyseal cylinder presenting green fractures (tubes) has been observed, although at least 30 remains display midshaft cylinder morphology (fake tubes) (Fig. 3i). All the evidence points therefore to a high incidence of diagenetic and recent (excavation-storing) breakage.

Bone surface modifications attributed to post-depositional agencies. (a) Trampling marks at different magnifications, (a1) x35, (a2) x100, (a3) x140, (b,c), rodent gnawing, (d) scratching, (e) scratching and cementations (white arrow), (f) scratching (white arrow indicates fissures), (g) trampling in bird scapula, (h) biochemical alteration (black arrow) and trampling (white arrow), (i) fake-tubes. Scale bar is 20 mm (in A4, 1:1).
Avian remains comprise the third most abundant taxonomic group in the Cova del Gegant IIIa assemblage. Of the 235 remains, 127 (54.04%) have been identified taxonomically. The rest have been identified anatomically and categorised as small birds. The specimens identified correspond to Pyrrhocorax graculus – the yellow-billed chough (NISP 61, MNE 50, MNI 7), Pyrrhocorax pyrrhocorax – the red-billed chough (NISP 61, MNE 45, MNI 6), Columba livia/oenas – the rock/stock dove (NISP 2, MNE 2, MNI 1) and Alectoris cf. rufa – the red-legged partridge (NISP 2, MNE 2, MNI 1). One piece of proximal tarsometatarsal bone from Accipiter gentilis – the northern goshawk, represents the only medium-sized bird specimen.
We estimated 189 of the elements, 31 of which correspond to young corvid individuals (13.19% of the NISP). The ulnae and tibiae indicate the presence of a minimum of 13 corvids (MNI). These, together with the remaining three individuals give a total of 16 MNI for birds in the assemblage. All anatomical elements are represented except for the cranium and ribs. The most represented in relative terms (%RA) are the ulna, tibia and coracoid, while the furcula, sternum and vertebrae are the least represented (Table 2). Relative proportions of skeletal elements and portions are shown in Supplementary Table S1. Our results indicate a degree of balance between wing and leg elements and between proximal and distal elements, while those of the limbs are more abundant than those of the core.
Breakage is high among the bird remains, especially in the case of limb bones that preserve proximal (38.9%) or distal parts (36.6%). A total of 90 elements are complete, of which 51 are complete phalanges (mainly pedal) and cervical vertebrae. In six cases, limb bones are represented by shaft fragments. However, as with the leporids, these shafts present features indicative of dry/post-depositional breakage (Supplementary Table S3). All the limb fragments analysed show transversal delineation in their fracture planes and shatter surfaces (97.3%). The few fracture planes that present smooth surfaces (3.7%) display right angles. This combination of features indicates dry/post-depositional breakage as the norm in the avian assemblage in layer IIIa of Cova del Gegant.
Bone surface modifications (BSMs)
The cortical surface of the bones in the assemblage is very good allowing us to present the percentage of modifications as a proportion of total remains (NSP). The most abundant modifications in terms of frequency, for both the macro- and meso-fauna, are those of geological/post-depositional origin (Supplementary Table S4). These modifications affect all the remains with a similar frequency and intensity, regardless of whether they are macro- or meso-vertebrate remains. Biological (mainly carnivore) modifications in contrast differ in their occurrence between macro-mammals, leporids and birds, although the frequency is similar (see below).
The modifications associated with the karstic environment, i.e. concretions, manganese coating, humic staining and humidity-related surface alterations such as exfoliations and fissures, are the most frequently documented. Manganese coating affects 27% of the remains in the assemblage, while root and organic staining affects 10.4%. Exfoliation, cracking and fissures resulting from changes in humidity (referred to here as degree 1 weathering) affect 9.4% of the specimens, the same proportion than concretions (CaCO3 cementations). Deformations due to sediment pressure affect only five macro-mammal remains. Classic trampling marks, identified as groups of striae, straight grooves and parallel organization, are recorded in four macro-mammal and in five avian remains, while groups of scratches of undetermined origin (albeit like trampling marks) affect seven leporid remains. Of the leporid specimens, just 0.2% present concentrations of very shallow grooves like trampling marks, referred to here as scratchings (Fig. 3). Classic trampling affects 1.1% of the macro-mammal remains and 2.1% of the bird bones. Biochemical modifications on bone surfaces in the form of irregular grooves and pits with corrosion around the borders and dissolution in the form of rounded cupules with rounded edges and a curved bottom are rare, affecting just 1% of NSP. Rodent gnawing was recorded on seven leporid remains.
In addition, we have observed modifications generated by carnivores in the form of tooth marks, digestion marks and carnivore breakage. Macro-mammal remains were only affected by carnivore modifications in two cases (0.5%). One shaft fragment from an indeterminate long bone (small mammal) presents a type A notch associated with tooth pits; however, the moderate digestion of the fragment prevents us from taking the pit measurements. Furthermore, a corpus fragment of an immature ungulate vertebra presents a score that is 1.07 mm in width
Forty leporid specimens display tooth marks in the form of pits (14 NISP), punctures (16 NISP) and scores (10 NISP). This, however, is a very small fraction of the assemblage of leporid bones (1.32% of the NISP). The marks appear mainly on innominate (32.5%), humerus (20%), femur and tibia (12.5%). In absolute terms, the most frequently gnawed element is the innominate and humerus (15.66 and 8.08% of NISP, respectively). In the case of the long limb bones, tooth marks mainly appeared on shaft portions and were normally isolated pits, although punctures appeared on fracture edges (4 NISP) and opposite pits/punctures (6 NISP) too. The tooth marks on innominate are mainly punctures (Supplementary Table S5 and Fig. 4). The size of the pits in the cortical tissue (shaft of long bones) point to tooth marks that are slightly larger than the experimental data recovered for both lynxes and foxes on leporid carcasses (breadth mean of 1.67 mm vs. 1.38 mm and 1,62 mm recorded in experimental works conducted with Iberian lynx on rabbit carcasses20,29 (Supplementary Fig. S1)). In the case of the scores, we documented nine bones with multiple small scores similar to those made by lynx cubs observed in experimental studies29 (Fig. 4b). Among the carnivore modifications, note should be made of the signals of digestion affecting 40 leporid remains, mainly on humerus (50%), phalanges (15%), and metatarsus (7.5%). In relative terms, the elements of the hindlimbs, especially the humerus (20.2% of the humerus NISP) and scapula (6.25% of scapula NISP), were affected. The degrees of digestion were mainly light (48.9%) and moderate (40.4%), and heavy in just 10.6% of the remains (Fig. 4). No single leporid bone presents extreme digestion damage.

Bone surface modifications attributed to biological agencies. (a) Carnivore tooth marks, puncture (white arrow), and pits at different magnifications, (a1) x35, (a2) measurements by HIROX KH-8700 3D software, (b) carnivore scoring, (b1-b2) at different magnifications x35, (c) digested bones, (d) burning damage (left) vs. non-burned leporid calcaneus, (e) carnivore pits in innominate, (e1) and detail at x35 magnifications, (f) cut marks in femur of leporid at different magnifications, (f1) x35, (f2) x50, (f3) x140. Scale bar is 20 mm (in A4, 1:1).
Anthropogenic modifications of bone are largely testimonial but, nevertheless, present (Fig. 4). The most frequent are thermo-alterations (58 NSP or 1.6%), associated, it can be presumed, with the combustion structures documented previously in layer IIIa44. This burning has affected the main taxa (lynx, birds and especially leporids) and some indeterminate remains to different degrees, ranging from rubefaction (grades 1/2 = 3) and cremation (grade 4/5 = 40) to calcination (grade 6 = 5). Butchering marks are totally absent except for five slicing marks located on the midshaft of the femur of an adult leporid. The cuts are clustered in the proximal meta-diaphysis on the anterior to medial side. All the slices show straight delineation, oblique orientation and a parallel correlation. The cuts are presumably the result of defleshing (Fig. 4f), while the femur has lost its distal epiphysis as the result of a modern fracture. During the analysis undertaken here and during the process of reviewing the manuscript, we (and 2 reviewers) have called into question the diagnosis of these marks more than once. Indeed, the assemblage contains other marks that are not readily diagnosed (see Figs. 3a and 4f), but which are presumably the result of geological processes, such as trampling. Observed at a lower magnification (e.g. hand lens), or under oblique light, these marks can easily be interpreted as cut marks. However, these difficulties, once again46, highlight the challenge of diagnosing certain taphonomic marks and stress the need to use recently developed methods (e.g47.) to solve equifinality problems. The fact that we have found a single remain with what appear to be cuts, but on which other anthropogenic marks of butchering are absent, means we have to be cautious in our interpretation of these marks. However, two conclusion can be drawn: i) these putative cut marks can be related to the ephemeral activities conducted in the cave in relation to fireplaces and ii) the scarce or non-existent human modifications of the bones analysed here suggest a fairly homogeneous assemblage and one that is, therefore, optimal for analysing lynx signatures.
The comparative multivariate analysis conducted between the leporid assemblage in layer IIIa of Cova del Gegant and several actualistic and archaeological assemblages of reference was based on two principal component analyses. The first examined the anatomical composition in terms of minimal animal units (%MAU) of the leporid remains in the assemblages (Fig. 5a) drawing on the data in Supplementary Table S6. The second combined taphonomic (% cut marks, % burned bones, % digested bones and % tooth/beak marks) with breakage data (% complete long limb bones) and mortality (% adults) (Supplementary Table S7) (Fig. 5b). The cluster analysis uses five taphonomic criteria (% cut marks, % burned, % digested, % tooth marks and % midshaft cylinders or tubes) (Supplementary Table S8). Results indicate the influence of two main taphonomic agencies, terrestrial carnivores, probably the lynx, and natural depositions in the assemblage of Cova del Gegant (Fig. 5c).

Comparative multivariate analysis of taphonomic features of layer IIIa at Cova del Gegant. (a) Principal component analysis of anatomical representation of leporid remains in several experimental and archaeological assemblages, (b) Principal component analysis of several taphonomic attributes of leporid assemblages generated by different taphonomic agents and processes, (c) Cluster analysis of several taphonomic attributes of leporid assemblages generated by different taphonomic agents and processes. To see reference data Supplementary Tables S6 to S8.
Coprolites
Two different morphotypes can be identified in layer IIIa at Cova del Gegant. However, morphotype 1 is represented by a single fragment, making morphotype 2 by far the most abundant (N = 147). Coprolites are dominated by shapeless specimens (N = 131. 89%), followed by fragments (N = 15.1%) and, finally, complete coprolites (N = 2.1%).
Their internal structure – comprising a spiral fabric and friable hardness – is the most frequently occurring feature allowing us to ascribe the vast majority of coprolites to morphotype 2. No aggregates are observed by the naked eye in the interior, which are spongy and present voids.
The morphometry of five coprolites (morphotype 2) was analysed. Morphotype 2 ranges in length from 17 to 50.46 mm and from 17.71 to 23.68 mm in diameter. The latter being the most significant dimension for discriminating between carnivore faeces. Cova del Gegant coprolites are clearly distinct from those of the hyena (morphotype 1) and the scats of the large felids (morphotype 2), including the mountain lion (Puma concolor) and jaguar (Panthera onca) (Supplementary Fig. S2). The five specimens are similar in size to the scats of the modern coyote (Canis latrans, mean of 21 ± 0.2 mm), wolf (Canis lupus lupus, mean of 19.70 ± 2.45 mm) and lynx (Lynx pardinus, mean of 21.8 mm). However, the scats of other small carnivores, such as those of the red fox (Vulpes vulpes, mean of 14 ± 0.2 mm) could overlap, given that their range is between 8 and 20 mm48.
Bone and tooth content are abundant in most of the coprolites (Supplementary Table S9). The bones are dominated by leporid remains (NR = 46.66%), followed by small mammal remains (NR = 7.1%) and shaft fragments from leporid or small mammals (NR = 9.13%). Larger mammals are represented by just five remains (6%). The four ulnae identified are represented by the proximal portion and the radius by the distal part.
Discussion
Several of the features detected in the assemblage point to the presence of a lynx den in layer IIIa of Cova del Gegant. Based on the biostratinomic alterations, our results strongly indicate that the Iberian lynx was the main bone modifier and accumulator of the fossil assemblage. While the assemblage is undoubtedly an accumulation of mixed origin, in which different agents and processes have intervened in the formation of the deposit, layer IIIa allows us to characterize it as the taphocoenosis of the Iberian lynx. Moreover, the BSMs generated by fossildiagenesis are abundant and have played a key role in the final configuration of the layer IIIa assemblage. However, in this sense, the data presented here contribute to support previous interpretations44.
Most of the taphonomic modifications recorded in layer IIIa are related to the feeding behaviour of mammalian carnivores. Tooth marks and digestion corrosion are especially prominent in the bones of the rabbit, the main prey here of the taphonomic accumulator. The frequency of the tooth marks, their location, the number of marks per element, the morphology and dimensions of the pits on the rabbit bones are all compatible with the neo-taphonomic record of the Iberian lynx18,20. While it is true that the dimensions of the tooth pits on the cortical tissue are slightly larger than those recorded in actualistic studies of current Iberian lynx20,29, recall that L. pardinus comes from a long anagenetic lineage (L. issidorensis- L. pardinus spelaeus- L. pardinus pardinus) characterized by the progressive reduction in body size49. This reduction in body size of the post-glacial lynx could explain the differences observed between the fossil assemblage and current records. Likewise, the number of pits is very low, and the methods employed appear to have changed, and despite the comparative analyses conducted here to illustrate our results, with a sample of fewer than 30 tooth pits, our statistical results are not fully representative. Nevertheless, even when we take into account the differences in tooth pit size and the differences in the methods employed, our results still indicate a considerable overlap between the means (95% CI) of the breadth of the tooth pits in the cortical bone found in experimental studies conducted with Iberian lynx and those found in our data from layer IIIa at Cova del Gegant. As such, these dimensions could be used as a reference for the tooth marks of medium-sized felines in archaeo-paleontological assemblages.
Moreover, small tooth scoring marks, associated with the presence of lynx cubs29, have been recorded too. Based on actualistic data, the gnawing damage caused by the fox on rabbit bones is very similar in terms of location, morphology and size of the pits to that generated by the Iberian lynxe.g. 24. Having said that, tooth marks are slightly more frequent in the case of canids and the occurrence of multiple pits/scores per element is rare in assemblages modified by felids yet common in those modified by foxes20,24,25,29,32,50,51,52.
Modifications generated by large raptors, such as beak/talon marks, and complete or near-complete long bones presenting marks of digestion are totally absent in our assemblage, which allows us to rule them out as modifying agents.
Digested bones are extremely scarce here compared with the high percentages recorded in actualistic studies for different leporid accumulators, especially compared to the high frequencies reported for nocturnal and diurnal raptors5,16,17,19,21,23,27,28,31,53,54,55. However, the analysis of the scats of small carnivores, such as lynx and fox, indicates high proportions of digested remains, especially those presenting heavy or extreme damage18,24,26,32. Taking into account the large presence of coprolites and fragments of coprolites in layer IIIa, the scarce presence of digested bones, and the total absence of extreme grades of digestion, may be related to the differential destruction of poorly preserved bones in archaeological sites56; however, our own errors in identifying the correct degree of digestion cannot be entirely ruled out here57. The morphology, dimensions and content of the coprolites associated with the faunal assemblage all point to the cave having served as a den for Iberian lynx. When denning, low mobility cubs tend to defecate inside the den, and it is the mothers, as in other fields, that are responsible for burying and removing droppings to avoid attracting the attention of predators. This has been detected before in the archeo-paleontological record of at least two sites with Iberian lynx accumulations56.
The variety of species represented and their frequency allows us to discard the large carnivores as the main bone accumulators of the assemblage58,59,60,61,62,63 and points rather to medium-small predators. The taxonomic representation is fully compatible with the diet of the Iberian lynx2,64. However, in Mediterranean ecosystems, both small carnivorous mammals and several nocturnal and diurnal raptors base their diet on rabbits10,64. In addition, in this region during the terminal Pleistocene, humans generated accumulations in which rabbits usually represented more than 90% of the remains7,8. For this reason, the taxonomic composition of the assemblage has only relative utility for the reconstruction of the taphonomic history.
The notable skeletal completeness of the lynx, and the abundance of remains reinforces its role as an accumulating agent65,66,67,68. In addition, the anatomical connections are compatible with the in situ deaths of individuals69 while the presence of infantile and young individuals is common in natal dens70,71,72. Both in our study and in other accumulations attributed totally or partially to the Iberian lynx38,56,73, the abundance of adults is frequent and difficult to explain. In other nearby Pleistocene sites, where lynxes were also involved in the accumulations, some lynx bones present clear evidence of carnivore damage74,75, but this is not the case at Cova del Gegant. Thus, the parsimonious explanation at our site would appear to be that the remains are associated with the natural death of adult individuals.
We cannot rule out the possibility that some elements – in particular, those of the ungulates – might have been sporadically introduced by large carnivores. In fact, the presence of at least one coprolite of morphotype 1 and some large bones with digestion corrosion are compatible with the contribution of hyenas to the accumulation59. However, the predominant contribution by the Iberian lynx is also quite compatible with the occasional introduction of elements by other carnivores. Finally, we cannot rule out altogether the natural origin of the few ungulate remains and the probable intrusion of some of the rabbit and chough remains, as the high frequency of complete long bones, the low incidence of BSMs and the high representation of immature individuals indicate15,76,77,78. For the same reason, the breaking patterns and anatomical composition of the meso-vertebrates are difficult to interpret. Although only a small fraction of the meaty, marrow-rich bones present breaking patterns identical to those observed in actualistic assemblages modified by small terrestrial carnivores20,24,25,79, most of them are complete-intact bones or bones fragmented by diagenesis. Moreover, the incidence of natural intrusions distorts taphonomic patterns by mixing an assemblage modified by carnivores with rabbits and corvids that have died naturally, thus invalidating the use of certain proxies, such as anatomical representation, breakage and mortality patterns, as our multivariate analysis has demonstrated. Moreover, the presence of burned bones indicates a secondary input of humans as biostratinomic agents, albeit our analysis of five robust taphonomic criteria rule out any major human contribution.
Apart from layer IIIa of the Cova del Gegant, few other Iberian Peninsula assemblages have previously been interpreted as accumulations generated almost exclusively by Iberian lynx. Two other cases, however, stand out, in which the main taxon is the rabbit (Oryctolagus cuniculus): Canyars (layer MLU)56 on the Mediterranean coast and the Navalmaíllo Rock Shelter (Layer F)80 on the Iberian plateau. A third case is provided by Buraca Escura (layer 2) on the Iberian Atlantic coast, which has been proposed as a lynx-modified accumulation of small-sized ungulates, mainly young ibex (Capra aff. pyrenaica), in which rabbits are scarce81. Here, the cave-dwelling ibex could have been the prey of lynx. Lynx remains are also very abundant in terms of NISP and MNI at the Canyars and Buraca Escura sites, but totally absent from Navalmaíllo. The Iberian lynx can be recognised as a species in the Iberian record for more than a million years82 and scholars believe that, in all probability, the origin of the species and the morphological changes observed during its anagenetic evolution are closely related to its highly specialized82,83,84,85. The trophic behaviour we have observed in Cova del Gegant, and which has also been recorded at the Canyars and Navalmaíllo sites, corroborate this point. However, local variations can be identified due, it seems, to changes in ecosystems, which may have obliged the lynx to prey on other animals, the case of Buraca Escura. More work is needed in this regard since we have no experimental references on the damage caused by Iberian lynx on ungulate carcasses, nor on other prey other than rabbit (except for partridges)51.
The Iberian lynx is the most frequently represented carnivore in the Iberian fossil record86,87,88. Their Pleistocene and early Holocene range extends into present-day southern France and the north of the Italic peninsula89, regions in which it was probably also abundant in the past. Their fossils are located above all in caves and rock shelters, dating back in its current form at least 1.6–1.7 million years or as Lynx pardinus spelaeus, an ancestor already specialized in predation on rabbits82. All in all, five conditions make the Iberian lynx an ideal candidate for generating large bone accumulations. These can be taphonomically identified as the following: (i) the Iberian lynx use caves and crevices as reproductive dens, (ii) they systematically transport their prey to these shelters to feed their cubs, (iii) they defecate in latrines, (iv) they are highly specialised predators and their diet is very narrow, and, (v), in common with other carnivores, the Iberian lynx is a great bone modifier and their taphonomic signal is readily tracked.
Based on the results presented in this study, the taphocoenosis of the Iberian lynx can be characterized by:
- (1)
the prevalence of rabbits in the assemblages, followed by other prey such as birds;
- (2)
the high representation of lynx remains in NISP, MNE and MNI, and of both adults and juveniles (cubs included);
- (3)
the presence of the taphonomic signal of the Iberian lynx on rabbit remains, characterized by the greater representation of the posterior appendicular skeleton (especially distal parts), the high breakage of long bones and relative shortage of tooth marks;
- (4)
The small size of the tooth marks (pits and punctures), usually confined to bones from natal dens, and the presence of tooth scoring on long bones attributed to kitten chewing; and
- (5)
The abundance of coprolites (morphotype 2) as a source of variable quantities of digested bones presenting high degrees of digestion but a relative absence of extreme degrees.
Methods
Excavation methodology
The current assemblage is the result of fieldwork undertaken by the Grup de Recerca del Quaternari (GRQ-SERP, University of Barcelona). In 2007, GRQ renewed fieldwork at the site, performed chronometric dating, established the stratigraphic framework and reanalysed the site’s archaeological record. The current phase in excavations is focused on the back of the main gallery and covers an area of ~7 m2. The location of all the remains excavated (including coprolites) was recorded in three-dimensions, according to the 1-m2 grid system, and identified by letter and number. Remains were mapped in situ prior to removal, whereas unidentified small fragments (<2 cm), postcranial bones of leporids, birds and small vertebrates were bagged by 1-m2 units of provenience. The sediments were water-screened using superimposed 5.2- and 0.5-mm mesh screens. The layer was affected by cross cuttings of rodent burrows and storage pits. Thus, burrow trails of fossorial animals were observed at the time of excavation, and the sediment contained in these burrows was excavated separately from that of layer IIIa to avoid bioturbated elements. The archaeological materials analysed here included all the remains recovered during field seasons 2007–2017, including the materials previously described44 and the materials between grid rows 22 and 28.
Archaeological taphonomy
A total of 3640 fossil faunal remains was studied from layer IIIa. The zooarchaeological analysis involved the identification of animal bones to genus or species level (where possible). Their anatomical region of origin (cranial, axial and appendicular) was determined, along with the skeletal element, the portion of that element, side and age (perinatal, juvenile, adult or senile) following standard zooarchaeological methods90. The taxonomic composition comprises mainly macro-mammals, leporids and small birds and, so, on occasion specific methods have been used. The number of specimens (NSP), number of identified specimens (NISP), minimum number of elements (MNE), minimum number of individuals69,91,92, skeletal survivorship in the form of relative abundance (%RA) for leporids and birds93 and the index of survival for macro-mammals (%ISu)63 were estimated. To calculate the MNI, we considered complete dental series and isolated teeth, taking into account their grade of eruption and wear94,95,96 and skeletal development patterns, fusion and ossification in line with Barone97. The state of development and fusion of the skeletal elements and dental eruption and replacement for lynx were estimated using a combination of several studies98,99. The epiphysis fusion pattern is the most commonly used method for establishing the age of rabbits at death. At about 9 or 10 months of age, all the epiphyses of the long bones are fused26,100,101,102,103. The distal epiphysis of the humerus begins to fuse at about 2 months of age. Therefore, the relative presence of the distal epiphyses of unfused humerus helps to identify the proportion of unweaned rabbits in the assemblages. The distal tibia begins this process at about 3 months. Therefore, the proportion of unfused distal tibiae reflects the proportion of very young individuals, although it is less precise than the identification of unweaned individuals by means of unfused distal humeri103.
Following Stiner104, we clustered the estimated individuals into three age groups: young, prime-adults and old individuals. To these three age classes, we incorporate perinatal when we note the absence of epiphysis, scarce ossification and very small elements. In order to include the non-identified remains with the identified specimens, we established four weight categories: large (>300 kg), medium (100–300 kg), small (10–100 kg) and very small (<10 kg).
The proportions of leporid skeletal elements were also evaluated using several indices:17,18 postcranial in relation to cranial (PCRT/CR, PCRAP/CR, PCRLB/CR, TA/MD, TAE/MDE, HU + FM/CR + MD), loss of distal limb elements (AUT/ZE, Z/E) and the ratio of forelimb to hindlimb elements (AN/PO, HU/FM, RDU/TA, MCP/MTT). In the case of birds, the ratios used were: the representation of wings versus legs (W/L), relation between proximal and distal parts of elements (P/D) and the proportion of core to limb elements (CO/LB) all expressed as a percentage21,51,105,106 (see Supplementary Table S1 for abbreviations). We calculated both Simpson and Shannon’s indices to assess taxonomic diversity following the recommendations of Grayson and Delpech107.
Breakage patterns were described in terms of the maximum length of all identified skeletal elements. Percentages were calculated for complete remains and isolated teeth. For immature individuals, the diaphysis of long bones with unfused epiphyses were considered complete elements. Breakage categories used for leporids and birds are fully described and illustrated in Lloveras et al. 17, see Fig. 1,106,108. Different “breakage categories were used depending on bone type: patellae, carpals and tarsals were classified as complete or fragmented; phalanges were recorded as complete, proximal, distal or fragmented, vertebrae were recorded as complete, vertebral body, vertebral epiphysis or spinous processes; breakage of teeth was calculated separately for isolated and in situ elements and they were classified as complete or fragmented”20, p.3030. Long bone cylinders also called “tubes” in the literature (fragments of long bones with snapped ends resulting from consumption), and V-shaped and helical fractures109 were also recorded. In this study, in the case of leporids, we only consider the elements containing an appreciable amount of marrow as humerus, tibia, and femur4. We considered as “fake tubes”, the long bone cylinders with sign of dry breakage (jagged or right-angle-smooth ends). Following Bunn110 and Villa and Mahieu109, we also recorded shaft circumference and shaft length along with fracture outline, angle and edge to explore the nature of fragmentation observed in the assemblage. Notches considered as “semi-circular- to arcuate-shaped indentations on fracture edges with corresponding negative flake scars on medullary surfaces [of limb bones]”111, p. 724) were also identified and analysed according to the typological classifications proposed by Egeland112, although we only recorded one notch (“complete or A type” defined by its having two inflection points on the cortical surface and a non-overlapping negative flake scar).
All skeletal remains were examined both macro- and microscopically. For microscopic observations, a binocular lens with variable magnifications (6.5 to 40) with an oblique cold light source was used. Punctures, pits, scores, crushing and helical fractures, crenulated edges and digestion marks57,113,114,115 were identified and documented. Punctures and pits were also classified by their number (isolated or multiple) and distribution (unilateral or bilateral)116. We also noted the size measurements of the pits, punctures and scores using the criteria outlined by Andrés et al.117 and compared them with experimental data from Andrés et al.117, Massigogue et al.52,and Rodríguez-Hidalgo et al.20,29. Cut marks were identified on the basis of the criteria outlined by Potts and Shipman118, Shipman and Rose119 and Domínguez-Rodrigo et al.120. Damage attributable to burning was described by colour (naked eye) using the six-grade scale proposed by Stiner et al.121: (1) slightly burned, (2)>half carbonized, (3) fully carbonized, (4) slightly calcined, (5)>half calcined and (6) fully calcined (completely white). Other modifications associated with the depositional history of the bone assemblage such as the presence of roots etching, chemical corrosion, biochemical marks, manganese oxide staining and trampling have been identified following the criteria detailed in several works122,123,124. Detailed pictures of some of the markings were taken using a HIROX KH-8700 3D Digital Microscope with an MXG-5000REZ triple objective revolving lens.
Statistical analysis
Principal component analyses (PCA) were performed with R125. The cluster analysis was generated with PAST126 using a Manhattan pairwise similarity matrix. For these analyses, we used data from leporid assemblages in actualistic, experimental and archaeological studies. In the case of the actualistic and experimental studies, all data refer to leporid predators. The archaeological studies are of deposits and layers of the Upper Palaeolithic and Epipalaeolithic, where anthropic activity was especially intense. Anatomical data have been standardized using the minimal animal unit (MAU), expressed as a percentage127. Taphonomic and age of death data are expressed as percentages. References for all data used for comparative purposes can be consulted in Supplementary Tables S6–S8.
Coprolite analysis
A total of 148 coprolite remains was studied from layer IIIa. The Cova del Gegant coprolites were analysed adapting the standardized method proposed by Jouy-Avantin et al.128 and Sanz et al.129. The analysis focused on the following aspects: preservation (complete, fragmentary and shapeless), morphotype (morphotype 1 to 3), morphometric on single droplets (recording maximum length and width or diameter in well preserved specimens) and bone content. The statistics derived from these data were compared with measurements reported in several studies on both fossil and modern scats88,129,130. For bone content analysis, the outer (external) surface or the sections of fragmented coprolites were examined to identify bone or tooth inclusions and where possible they were assigned to taxonomic and anatomical groups. Coprolites were not disaggregated in order to identify these items. Due to the high degree of fragmentation we used the taxonomic categories: large mammals (larger than leporids), mesofauna (leporids) and small mammals (smaller than leporids). Undetermined remains were recorded as skeletal tissues, such as shaft (long bone) or epiphysis (spongy) fragment129.
Data availability
All necessary permits were obtained from the Departament de Cultura of the Generalitat de Catalunya and from the local authorities for the excavation work at the Cova del Gegant under the direction of M.S. and J.D. for the study described here, which complied with all relevant regulations.
References
Rodríguez, A. & Delibes, M. Current range and status of the Iberian lynx Felis pardina Temminck, 1824 in Spain. Biol. Conserv. 61, 189–196 (1992).
Delibes, M. Feeding ecology of the Spanish lynx in the Coto Doñana. Acta Theriol. 25, 309–324 (1980).
Pérez-Ripoll, M. La consommation humaine des lapins pendant le Palólithique dans la région de Valencia (Espagne) et l’étude des niveaux gravétiens de La Cova de les Cendres (Alicante). in Actes des XXIV rencontres internationales d’Archéologie et d’Histoire d’Antibes. (eds. Brugal, J. P. & Desse, J.) 191–206 (Éditions APDCA, 2004).
Pérez-Ripoll, M. Marcas de carnicería, fracturas intencionadas y mordeduras de carnívoros en huesos prehistóricos del Mediterráneo español. (Diputación de Alicante, 1992).
Hockett, B. S. & Haws, J. A. Taphonomic and methodological perspectives of leporid hunting during the Upper Paleolithic of the Western Mediterranean Basin. J. Archaeol. Method. Th. 9, 269–302 (2002).
Aura Tortosa, J. E., Villaverde, V., Pérez Ripoll, M., Martínez Valle, R. & Calatayud, P. G. Big game and small prey: Paleolithic and Epipaleolithic economy from Valencia (Spain). J. Archaeol. Method. Th. 9, 215–268 (2002).
Fa, J. E., Stewart, J. R., Lloveras, L. & Vargas, J. M. Rabbits and hominin survival in Iberia. J. Hum. Evol. 64, 233–241 (2013).
Morin, E. et al. New evidence of broader diets for archaic Homo populations in the northwestern Mediterranean. Sci Adv 5, eaav9106 (2019).
Valverde, J. A. An ecological sketch of the Coto Doñana. (British Birds, 1958).
Delibes, M. & Hiraldo, F. The rabbit as a prey in the Iberian Mediterranean ecosystem in Proceedings of the World Lagomorph Conference (eds. Myers, K. & MacInnes, C. D.) 614–622 (University of Guelph, 1981).
Valverde, J. A. Estructura de una comunidad mediterránea de vertebrados terrestres. (Consejo Superior de Investigaciones Científicas, 1967).
Gibb, J. A. Factors affecting population density in the wild rabbit, Oryctolagus cuniculus (L.), and their relevance to small mammals in Evolutionary Ecology (eds. Stonehouse, B. & Perrins, C.) 33–46 (Macmillan Education UK, 1977).
Parer, I. & Milkovits, G. Recolonisation by rabbits (Oryctolagus cuniculus) after warren ripping or warren fumigation. Rangeland J. 16, 51 (1994).
Gea-Izquierdo, G., Muñoz-Igualada, J. & San Miguel-Ayanz, A. Rabbit warren distribution in relation to pasture communities in Mediterranean habitats: consequences for management of rabbit populations. Wildl. Res. 32, 723–731 (2006).
Pelletier, M. et al. Identifying fossil rabbit warrens: Insights from a taphonomical analysis of a modern warren. J. Archaeol. Sci. Rep. 10, 331–344 (2016).
Lloveras, L., Cosso, A., Solé, J., Claramunt-López, B. & Nadal, J. Taphonomic signature of golden eagles (Aquila chrysaetos) on bone prey remains. Hist. Biol. 30, 835–854 (2018).
Lloveras, L., Moreno-Garcia, M. & Nadal, J. Taphonomic study of leporid remains accumulated by the Spanish Imperial Eagle (Aquila adalberti). Geobios Mem. Spec. 41, 91–100 (2008).
Lloveras, L., Moreno-García, M. & Nadal, J. Taphonomic analysis of leporid remains obtained from modern Iberian lynx (Lynx pardinus) scats. J. Archaeol. Sci. 35, 1–13 (2008).
Lloveras, L., Moreno-García, M. & Nadal, J. The eagle owl (Bubo bubo) as a leporid remains accumulator: taphonomic analysis of modern rabbit remains recovered from nests of this predator. Int. J. Osteoarchaeol. 19, 573–592 (2009).
Rodríguez-Hidalgo, A. et al. Feeding behaviour and taphonomic characterization of non-ingested rabbit remains produced by the Iberian lynx (Lynx pardinus). J. Archaeol. Sci. 40, 3031–3045 (2013).
Lloveras, L., Thomas, R., Lourenço, R., Caro, J. & Dias, A. Understanding the taphonomic signature of Bonelli’s Eagle (Aquila fasciata). J. Archaeol. Sci. 49, 455–471 (2014).
Lloveras, L. et al. The role of the Egyptian Vulture (Neophron percnopterus) as a bone accumulator in cliff rock shelters: an analysis of modern bone nest assemblages from North-eastern Iberia. J. Archaeol. Sci. 44, 76–90 (2014).
Lloveras, L., Moreno-García, M. & Nadal, J. Assessing the variability in taphonomic studies of modern leporid remains from Eagle Owl (Bubo bubo) nest assemblages: the importance of age of prey. J. Archaeol. Sci. 39, 3754–3764 (2012).
Lloveras, L., Moreno-García, M. & Nadal, J. Feeding the foxes: an experimental study to assess their taphonomic signature on leporid remains. Int. J. Osteoarchaeol. 22, 577–590 (2012).
Lloveras, L., Thomas, R., Cosso, A., Pinyol, C. & Nadal, J. When wildcats feed on rabbits: an experimental study to understand the taphonomic signature of European wildcats (Felis silvestris silvestris). Archaeol. Anthropol. Sci. 10, 449–464 (2018).
Cochard, D. Etude taphonomique des léporidés d´une tanière de renard actuelle: apport d´un référentiel à la reconnaissance des accumulations anthropiques. Revue de Paléobiologie, Genève 23, 659–673 (2004).
Hockett, B. S. Corroded, thinned and polished bones created by Golden eagles (Aquila chrysaetos): Taphonomic iimplications for archaeological interpretations. J. Archaeol. Sci. 23, 587–591 (1996).
Hockett, B. S. Comparison of leporid bones in raptor pellets, raptor nests, and archaeological sites in the Great Basin. N. Am. Archaeol. 16(3), 223–238 (1995).
Rodríguez-Hidalgo, A., Saladié, P., Marín, J. & Canals, A. Expansion of the referential framework for the rabbit fossil accumulations generated by Iberian lynx. Palaeogeogr. Palaeocl. 418, 1–11 (2015).
Mallye, J.-B., Cochard, D. & Laroulandie, V. Accumulations osseuses en périphérie de terriers de petits carnivores: les stigmates de prédation et de fréquentation. Ann. Paléontol. 94, 187–208 (2008).
Rodríguez-Hidalgo, A., Saladié, P. & Marín, J. Caracterización de la señal tafonómica del lince ibérico a través de tres conjuntos experimentales in Interaccions entre felins i humans: homenatge a Innocenci Sarrión Montañana (eds. Sanchis, A. & Pascual Benito, J. L.) 135–159 (Museu de Prehistòria de València, 2017).
Sanchis Serra, A. Los restos de Oryctolugus cuniculus en la tafocenosis de Bubo bubo y Vulpes vulpes y su aplicación a la caracterización del registro faunístico arqueológico. Saguntum 32, 31–50 (2000).
Sanchis Serra, A. & Pascual Benito, J. L. Análisis de las acumulaciones óseas de una guarida de pequeños mamíferos carnívoros (Sitjar Baix, Onda, Castellón): implicaciones arqueológicas. Archaeofauna 20, 47–71 (2011).
Lloveras, L., Moreno-García, M., Nadal, J. & Zilhao, J. Who brought in the rabbits? Taphonomical analysis of Mousterian and Solutrean leporid accumulations from Gruta do Caldeirao (Tomar, Portugal). J. Archaeol. Sci. 38, 2434–2449 (2012).
Rufà, A., Blasco, R., Rivals, F. & Rosell, J. Leporids as a potential resource for predators (hominins, mammalian carnivores, raptors): An example of mixed contribution from level III of Teixoneres Cave (MIS 3, Barcelona, Spain). C. R. Palevol 13, 665–680 (2014).
Lloveras, L. & Moreno García, M. & Soler i Masferrer, N. The application of actualistic studies to assess the taphonomic origin of Musterian rabbit accumulations from Arbreda Cave (North-East Iberia). Archaeofauna 19, 99–119 (2010).
Rosado-Méndez, N. Y., Cebrià, A., Mestres, J., Nadal, J. & Lloveras, L. Biostratinomic marks on rabbit remains and Epipalaeolithic hunter-gatherer behaviour in NE Iberia. The case of Cova de la Guineu (Font Rubí, Barcelona). J. Archaeol. Sci. Rep. 21, 872–883 (2018).
Rodríguez-Hidalgo, A. J., Saladié, P. & Canals, A. Following the white rabbit: A case of a small game procurement site in the Upper Palaeolithic (Sala de las Chimeneas, Maltravieso Cave, Spain). Int. J. Osteoarchaeol. 23, 34–54 (2013).
Brugal, J.-P., Díez Fernández-Lomana, J. C., Huguet, R., Michel, P. & Rosell, J. Karstic cavities, natural bone accumulations and discrete human activities in the European Palaeolithic: some case studies in Paleolithic zooarchaeology in practice (eds. Haws, J. A., Hockett, B. S. & Brugal, J.-P.) 1–12 (BAR international series, 2006).
Hockett, B. S. Taphonomy of a carnivore-accumulated rabbit bone assemblage from Picareiro Cave, central Portugal. J. Iberian Archaeol. 1, 225–230 (1999).
Daura, J. et al. A Neandertal mandible from the Cova del Gegant (Sitges, Barcelona, Spain). J. Hum. Evol. 49, 56–70 (2005).
Quam, R. et al. The Neandertals of northeastern Iberia: new remains from the Cova del Gegant (Sitges, Barcelona). J. Hum. Evol. 81, 13–28 (2015).
Daura, J. et al. Stratigraphic context and direct dating of the Neandertal mandible from Cova del Gegant (Sitges, Barcelona). J. Hum. Evol. 59, 109–122 (2010).
Sanz, M., Daura, J., Égüez, N. & Cabanes, D. On the track of anthropogenic activity in carnivore dens: Altered combustion structures in Cova del Gegant (NE Iberian Peninsula). Quatern. Int. 437, 102–114 (2017).
Pavao, B. & Stahl, P. W. Structural density assays of leporid skeletal elements with implications for taphonomic, actualistic and archaeological research. J. Archaeol. Sci. 26, 53–66 (1999).
Domínguez-Rodrigo, M. et al. Use and abuse of cut mark analyses: The Rorschach effect. J. Archaeol. Sci. 86, 14–23 (2017).
Rodríguez-Hidalgo, A. et al. The Châtelperronian Neanderthals of Cova Foradada (Calafell, Spain) used imperial eagle phalanges for symbolic purposes. Sci Adv 5, eaax1984 (2019).
Green, J. S. & Flinders, J. T. Diameter and pH comparisons of coyote and red fox scats. J. Wildl. Manage. 45, 765 (1981).
Werdelin, L. The evolution of lynxes. Ann. Zool. Fennici 18, 37–71 (1981).
Rafuse, D. J. et al. Análisis comparativo de los patrones de modificaciones óseas de dos carnívoros sudamericanos: el gato montés (Leopardus geoffroyi) y el zorro pampeano (Lycalopex gymnocercus). Aportes para la identificación de la acción de pequeños carnívoros en el registro arqueológico. Magallania 42, 165–184 (2014).
Rodríguez-Hidalgo, A., Saladié, P., Marín, J. & Canals, A. Bird-bone modifications by Iberian lynx: A taphonomic analysis of non-ingested red-legged partridge remains. Quatern. Int. 421, 228–238 (2016).
Massigoge, A. et al. Estudio comparativo de las marcas de dientes producidas por dos pequeños carnívoros sudamericanos. Rev. Chil. Antropol. 0, 42–49 (2014).
Hockett, B. S. Toward distinguishing human and raptor patterning on leporid bones. Am. Antiq. 56, 667–679 (1991).
Schmitt, D. N. The taphonomy of Golden Eagle prey accumulations at Great Basin roosts. J. Ethnobiol. 15, 237–256 (1995).
Bochenski, Z. M., Tomek, T., Tornberg, R. & Wertz, K. Distinguishing nonhuman predation on birds: pattern of damage done by the white-tailed eagle Haliaetus albicilla, with comments on the punctures made by the golden eagle Aquila chrysaetos. J. Archaeol. Sci. 36, 122–129 (2009).
Rosado-Méndez, N. Y., Lloveras, L., Daura, J., Nadal, J. & Sanz, M. Predator agents and leporid accumulations: The case of terrasses de la riera dels Canyars (Gavà, Barcelona, Spain). J. Archaeol. Method. Th. 22, 980–1005 (2015).
Lloveras, L., Moreno-García, M., Nadal, J. & Thomas, R. Blind test evaluation of accuracy in the identification and quantification of digestion corrosion damage on leporid bones. Quat. Int. 330, 150–155 (2014).
Egeland, A., Egeland, C. P. & Bunn, H. Taphonomic analysis of a modern Spotted Hyaena (Crocuta crocuta) den from Nairobi, Kenya. J. Taph. 6, 275–299 (2008).
Pokines, T. J. & Kerbis-Peterhans, J. C. Spotted hyaena (Crocuta crocuta) den use and taphonomy in the Masai Mara National Reserve, Kenya. J. Archaeol. Sci. 34, 1914–1931 (2007).
Skinner, J. D., Henschel, J. R. & Jaarsveld, A. S. Bone-collecting habits of spotted hyaenas Crocuta crocuta in the Kruger National Park. S. Afr. J. Zool. 21, 303–308 (1986).
Mwebi, O. & Brugal, J. Comparative taphonomical studies of sympatric hyaenids (Crocuta crocuta and Hyaena hyaena) bone assemblages, insights from modern dens in Kenya. Quaternaire 29, 13–20, https://doi.org/10.4000/quaternaire.8563 (2018).
Arriaza, M. C., Domínguez-Rodrigo, M., Yravedra, J. & Baquedano, E. Lions as bone accumulators? Paleontological and ecological implications of a modern bone assemblage from Olduvai Gorge. PLoS One 11, e0153797 (2016).
Brain, C. K. The hunters of the hunted? An introduction to African cave taphonomy. (The University of Chicago Press, 1981).
Gil-Sánchez, J. M., Ballesteros-Duperón, E. & Bueno-Segura, J. F. Feeding ecology of the Iberian lynx Lynx pardinus in eastern Sierra Morena (Southern Spain). Acta Theriologica 51, 85–90 (2006).
Diedrich, C. G. & Žák, K. Prey deposits and den sites of the Upper Pleistocene hyena Crocuta crocuta spelaea (Goldfuss, 1823) in horizontal and vertical caves of the Bohemian Karst (Czech Republic). Bulletin of Geosciences 81, 237–276 (2006).
Sauqué, V., Rabal-Garcés, R., Sola-Almagro, C. & Cuenca-Becós, G. Bone accumulation by leopards in the late Pleistocene in the Moncayo Massif (Zaragoza, NE Spain). PLoS One 9, e92144 (2014).
Aubry, T., Fontugne, M. & Moura, M.-H. Les occupations de la grotte de Buraca Grande depuis le Paléolithique supérieur et les apports de la séquence holocène à l’étude de la transition Mésolithique/Néolithique au Portugal. Bull. Soc. Préhist. Fr. 94, 182–190 (1997).
Sanchis, A. et al. Pleistocene leopards in the Iberian Peninsula: New evidence from palaeontological and archaeological contexts in the Mediterranean region. Quaternary Sci. Rev. 124, 175–208 (2015).
Lyman, R. L. Vertebrate Taphonomy. (Cambrigde University Press, 1994).
Fernández, N. & Palomares, F. The selection of breeding dens by the endangered Iberian Lynx (Lynx pardinus): implications for its conservation. Biol. Conserv. 94, 51–61 (2000).
Boutros, D. et al. Characterisation of Eurasian lynx Lynx lynx den sites and kitten survival. Wildlife Biol. 13, 417–429 (2007).
Fernández, N., Palomares, F. & Delibes, M. The use of breeding dens and kitten development in the Iberian lynx (Lynx pardinus). J. Zool. 258, 1–5 (2002).
Daura, J. et al. Terrasses de la Riera dels Canyars (Gavà, Barcelona): the landscape of Heinrich Stadial 4 north of the ‘Ebro frontier’ and implications for modern human dispersal into Iberia. Quat. Sci. Rev. 60, 26–48 (2013).
Sanz, M. & Daura, J. Taphonomic analysis of an ungulate-dominated accumulation at the Pleistocene Cova del Rinoceront site near Barcelona, Spain (northeastern Iberian Peninsula). Palaeogeogr. Palaeocl. 498, 24–38 (2018).
Sanz, M. & Daura, J. Carnivore involvement in bone assemblages based on taphonomic and zooarchaeological analyses of Cova del Coll Verdaguer site (Barcelona, Iberian Peninsula). Hist. Biol. 30, 807–820 (2018).
Laroulandie, V. Alpine chough Pyrrhocorax graculus from Pleistocene sites between Pyrenees and Alps: Natural versus cultural assemblages in Birds in Archaeology: Proceedings of the 6th Working Meeting (BWG) of ICAZ (eds. Prummel, W., Brinkhuizen, D. & Zeiler, J.), 219–232 (Barkhuis, 2010).
Laroulandie, V. Anthropogenic versus non-anthropogenic bird bone assemblages: new criteria for their distinction in Biosphere to lithosphere: new studies in vertebrate taphonomy (ed. O'Connor, T.) 25–30 (Oxbow Books, 2005).
Laroulandie, V. Taphonomie et archéozoologie des oiseaux en grotte: applications aux sites Paléolithiques du Bois-Ragot (Vienne), de Combe Saunière (Dordogne) et de La Vache (Ariège). (Université Sciences et Technologies-Bordeaux I, 2000).
Álvarez, M. C. et al. Bone modification and destruction patterns of leporid carcasses by Geoffroy’s cat (Leopardus geoffroyi): An experimental study. Quat. Int. 278, 71–80 (2012).
Arriaza, M. C. et al. Lagomorph predation represented in a middle Palaeolithic level of the Navalmaíllo Rock Shelter site (Pinilla del Valle, Spain), as inferred via a new use of classical taphonomic criteria. Quater. Int. 436, 294–306 (2017).
Aubry, T. et al. Modalités d’occupations au Paléolithique supérieur dans la grotte de Buraca Escura (Redinha, Pombal, Portugal). Revista Portuguesa de Arqueología 4, 19–46 (2001).
Boscaini, A., Madurell-Malapeira, J., Llenas, M. & Martínez-Navarro, B. The origin of the critically endangered Iberian lynx: Speciation, diet and adaptive changes. Quaternary Sci. Rev. 123, 247–253 (2015).
Garcia-Perea, R., Gisbert, J. & Palacios, F. Review of the biometrical and morphological features of the skull of the Iberian lynx, Lynx pardina (Temminck, 1824). Säugetierkundliche Mitteilungen 32, 249–259 (1985).
Johnson, W. E. et al. Phylogenetic and phylogeographic analysis of Iberian lynx populations. J. Hered. 95, 19–28 (2004).
Ghezzo, E., Boscaini, A., Madurell-Malapeira, J. & Rook, L. Lynx remains from the Pleistocene of Valdemino cave (Savona, Northwestern Italy), and the oldest occurrence of Lynx spelaeus (Carnivora, Felidae). Rend. Lincei Sci. Fis. Nat. 26, 87–95 (2015).
Yravedra, J. Aprovechamiento cárnico del lince (Lynx pardina) durante el Pleistoceno superior en el interior de la Península Ibérica. Munibe 57, 303–311 (2005).
Arribas, O. Fauna y paisaje de los Pirineos en la Era Glaciar. (Lynx Editions, 2004).
Brugal, J.-P. Carnivores pléistocènes (Hyénidés, Canidés, Félidés) dans les grottes du Portugal in 1 a Reunión de científicos sobre cubiles de hiena (y otros grandes carnívoros) en los yacimientos arqueológicos de la Península Ibérica (eds. Baquedano, E. & Rosell, J.) 91–106 (Museo Arqueológico Regional de la Comunidad de Madrid, 2010).
Rodríguez-Varela, R. et al. Ancient DNA evidence of Iberian lynx palaeoendemism. Quat. Sci. Rev. 112, 172–180 (2015).
Reitz, E. J. & Wing, E. S. Zooarchaeology. (Cambridge University Press, 1999).
Lyman, R. L. Quantitative Paleozoology. (Cambridge University Press, 2008).
Andrews, P. Owls, caves and fossils. (British Museum, 1990).
Dodson, P. & Wexlar, D. Taphonomic investigations of owl pellets. Paleobiology 5, 275–284 (1979).
Levine, M. A. The use of crown height measurements and eruption-wear-sequences to age horse teeth in Ageing and sexing animal bones from archaeological sites (eds. Wilson, B., Grigson, C. & Payne, S.) 223–250 (Archaeopress, 1982).
Mariezkurrena, K. Contribución al conocimiento del desarrollo de la dentición y el esqueleto postcraneal de Cervus elaphus. Munibe 35, 149–202 (1983).
Hillson, S. Teeth. (Cambridge University Press, 1986).
Barone. Atlas de mammiferes domestiques. (Vigot Frères, 1966).
Smuts, G. L., Anderson, J. L. & Austin, J. C. Age determination of the African lion (Panthera leo). J. Zool. 185, 115–146 (1978).
Garcia-Perea, R. Patterns of postnatal development in skulls of lynxes, genus Lynx (Mammalia: Carnivora). J. Morphol. 229, 241–254 (1996).
Hockett, B. S. & Bicho, N. The Rabbits of Picareiro Cave: Small Mammal hunting during the Late Upper Palaeolithic in the Portuguese Estremadura. J. Archaeol. Sci. 27, 715–723 (2000).
Callou, C. De la garenne au clapier. Histoire et archéologie du lapin européen. (Muséum National d’Histoire Naturelle, 2003).
Cochard, D. & Brugal, J. Importance des fonctions de sites dans les accumulations paléolethiques de léporidés. in Petits animaux et sociétés humaines. Du complément alimentaire aux ressources utilitaires. XXIV recontres internatonales d’arqueologie et d’historie d’Antibes (eds. Brugal, J. & Desse, J.) 283–296 (2004).
Jones, E. L. Prey choice, mass collecting, and the wild European rabbit (Oryctolagus cuniculus). J. Anthropol. Archaeol. 25, 275–289 (2006).
Stiner, M. C. The use of mortality patterns in archaeological studies of hominid predatory adaptations. J. Anthropol. Archaeol. 9, 305–351 (1990).
Ericson, P. G. P. Interpretations of archaeological bird remains: A taphonomic approach. J. Archaeol. Sci. 14, 65–75 (1987).
Bocheński, Z. M. Owls, Diurnal Raptors and Humans: signatures on avian bones in Biosphere to Lithosphere: new studies in vertebrate taphonomy (ed. O’Connor, T.) 31–45 (Oxbow Books, 2005).
Grayson, D. K. & Delpech, F. Specialized Early Upper Palaeolithic hunters in southwestern France? J. Archaeol.Sci. 29, 1439–1449 (2002).
Bocheński, Z. M., Boev, Z., Mitev, I. & Tomek, T. Patterns of bird bone fragmentation in pellets of the Tawny Owl (Strix aluco) and the Eagle Owl (Bubo bubo) and their taphonomical implications. Acta Zool. Cracov. 36, 313–328 (1993).
Villa, P. & Mahieu, E. Breakage patterns of human long bones. J. Hum. Evol. 21, 27–48 (1991).
Bunn, H. T. Comparative analysis of modern bone assemblages from a San hunter-gatherer camp in the Kalahari desert, Botswana, and from a spotted hyena den near Nairobi, Kenia in Animals and Archaeology: 1.- Hunters and Their Prey (eds. Clutton-Brock, J. & Grigson, C.) 143–148 (British Archaeological Reports. International Series 163, 1983).
Capaldo, S. D. & Blumenschine, R. J. A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on bovid long bones. Am. Antiquity 59, 724–748 (1994).
Egeland, C. P. Zooarchaeological and taphonomic perspectives on hominid and carnivore interactions at Olduvai Gorge, Tanzania. (Indiana University, 2007).
Haynes, G. Evidence of carnivore gnawing on Pleistocene and recent mammalian bones. Paleobiology 6, 341–351 (1980).
Haynes, G. A guide for differentiating mammalian carnivore taxa responsible for gnaw damage to herbivore limb bones. Paleobiology 9, 164–172 (1983a).
Binford, L. R. Bones. Ancient men and modern myths. (Ac. Press, 1981).
Sanchis, A., Morales, J. V. & Pérez Ripoll, M. Creación de un referente experimental para el estudio de las alteraciones causadas por dientes humanos sobre huesos de conejo in La investigación experimental aplicada a la arqueología. Actas del segundo congreso internacional de arqueología experimental (eds. Morgado, A., Baena, J. & García, D.) 343–349 (2011).
Andrés, M., Gidna, A. O., Yravedra, J. & Domínguez-Rodrigo, M. A study of dimensional differences of tooth marks (pits and scores) on bones modified by small and large carnivores. Archaeol. Anthropol. Sci. 4, 209–219 (2012).
Potts, R. & Shipman, P. Cutmarks made by stone tools on bones from Olduvai Gorge, Tanzania. Nature 291, 577–580 (1981).
Shipman, P. & Rose, J. Evidence of butchery and hominid activities at Torralba and Ambrona: An evaluation using microscopic techniques. J. Archaeol. Sci. 10, 465–474 (1983).
Domínguez-Rodrigo, M., de Juana, S., Galán, A. B. & Rodríguez, M. A new protocol to differentiate trampling marks from butchery cut marks. J. Archaeol. Sci. 36, 2643–2654 (2009).
Stiner, M. C., Kuhn, S. L., Weiner, S. & Bar-Yosef, O. Differential burning, recrystallization, and fragmentation of archaeological bone. J. Archaeol. Sci. 22, 223–237 (1995).
Behrensmeyer, A. K., Kathleen, D. G. & Yanagi, G. T. Trampling as a cause of bone surface damage and pseudocutmarks. Nature 319, 768–771 (1986).
Fiorillo, A. R. An experimental study of trampling: implications for the fossil record in Bone Modification (eds. Bonnichsen, R. & M.H.Sorg) 61–72 (University of Maine Center for the Study of the First Americans, 1989).
Fernández-Jalvo, Y. & Andrews, P. Atlas of Taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification. (Springer, 2016).
Team, R. C. R: Un Lenguaje y Entorno para la Estadística Informática. FR Estadística. Viena, Austria. Obtenido de http://www.rproject.org.
Hammer, Ø. & Others. PAST PAleontological STatistics Version 2.17 Reference Manual. (2012).
Binford, L. R. Faunal remains from Klassies River Mouth. (Ac. Press, 1984).
Jouy-Avantin, F., Debenath, A., Moigne, A.-M. & Moné, H. A Standardized method for the description and the study of coprolites. J. Archaeol. Sci. 30, 367–372 (2003).
Sanz, M., Daura, J., Égüez, N. & Brugal, J.-P. Not only hyenids: A multi-scale analysis of Upper Pleistocene carnivore coprolites in Cova del Coll Verdaguer (NE Iberian Peninsula). Palaeogeogr., Palaeocl. 443, 249–262 (2016).
Larkin, N. R., Alexander, J. & Lewis, M. D. Using experimental studies of recent faecal material to examine hyaena coprolites from the West Runton Freshwater Bed, Norfolk, U.K. J. Archaeol. Sci. 27, 19–31 (2000).
Acknowledgements
This research was supported by the projects “Els canvis climàtics durant el plistocè superior a la costa central catalana i l’impacte en les poblacions neandertals i humans anatòmicament moderns (CLT009/18/00022 Direcció General del Patrimoni Cultural, Generalitat de Catalunya)”, 2017SGR-00011 (Generalitat de Catalunya) and HAR2017-86509 (Spanish Government). A. Rodríguez-Hidalgo, M. Sanz and J. Daura were financed by the Spanish Ministry of Science, Innovation and Universities (A.R-H: IJC-2018-037447-I and M.S.: IJCI-2017-33908, Subprograma Juan de la Cierva. J.D.: RYC-2015-17667, Ramón y Cajal contract). A. Sánchez-Marco is a member of the consolidated research group 2017SGR86, Generalitat de Catalunya. He also acknowledges support from project CGL2016-76431-P (AEI/FEDER, EU) of the State Research Agency and the European Fund for Regional Development. Thanks to the International Research Network (IRN 0871 CNRS-INEE): Taphonomy European Network (TaphEN).
Author information
Authors and Affiliations
Contributions
All authors have contributed to the realization of this work. M.S. and J.D. have directed the excavation work. A.R.-H. and M.S. have analysed the materials and together with J.D. have written the paper. A.S.-M. has taxonomically determined the remains of birds. All authors have read, corrected and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rodríguez-Hidalgo, A., Sanz, M., Daura, J. et al. Taphonomic criteria for identifying Iberian lynx dens in quaternary deposits. Sci Rep 10, 7225 (2020). https://doi.org/10.1038/s41598-020-63908-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-63908-6
This article is cited by
-
The residential occupation of the Gruta da Aroeira (Almonda, Portugal) cave site: shedding light on hunting and subsistence practices in the Middle Pleistocene of western Eurasia
Archaeological and Anthropological Sciences (2024)
-
The characterization of bearded vulture (Gypaetus barbatus) coprolites in the archaeological record
Scientific Reports (2023)
-
Quaternary taphonomy: understanding the past through traces
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
