
Abstract
Depredation of wild turkey (Meleagris gallopavo) nests is a leading cause of reduced recruitment for the recovering and iconic game species. Invasive wild pigs (Sus scrofa) are known to depredate nests, and have been expanding throughout the distributed range of wild turkeys in North America. We sought to gain better insight on the magnitude of wild pigs depredating wild turkey nests. We constructed simulated wild turkey nests throughout the home ranges of 20 GPS-collared wild pigs to evaluate nest depredation relative to three periods within the nesting season (i.e., early, peak, and late) and two nest densities (moderate = 12.5-25 nests/km2, high = 25-50 nests/km2) in south-central Texas, USA during March–June 2016. Overall, the estimated probability of nest depredation by wild pigs was 0.3, equivalent to native species of nest predators in the study area (e.g., gray fox [Urocyon cinereoargenteus], raccoon [Procyon lotor], and coyote [Canis latrans]). Female wild pigs exhibited a constant rate of depredation regardless of nesting period or density of nests. However, male wild pigs increased their rate of depredation in areas with higher nest densities. Management efforts should remove wild pigs to reduce nest failure in wild turkey populations especially where recruitment is low.
Similar content being viewed by others


Introduction
Generalist invasive species negatively impact native species by both consuming and competing with the native species for resources1,2. In particular, the negative impact of invasive wild pigs (Sus scrofa); also termed feral swine, feral hogs, or wild boars3; on native ecosystems has become an increasing concern as populations of wild pigs continue to expand4. Wild pigs were first introduced to North America by European colonists in the early 1500s5. Over the past 3 decades wild pigs have rapidly expanded their range from 18 to 35 of the 50 United States6, because of their generalist nature and continued translocation by humans7,8,9,10. In their expanded range, wild pigs have the potential to triple their population every 5 years in the absence of control efforts10, leading to detrimental effects on native species4. In particular, wild pigs alter habitat and compete with and prey upon native species11,12,13,14.
Wild pigs have been implicated as nest predators for ground-nesting birds and reptiles15,16,17,18. Nest failure is the most substantial limitation on population growth of ground-nesting birds, and has been predominantly attributed to nest depredation19,20,21. The impacts of specific nest predators, such as wild pigs, on the reproductive success of a ground-nesting bird can depend on predator behavior. Reports suggest that wild pigs depredate nests opportunistically22,23, or contrarily seek high concentrations of nests15, although additional reports are variable4,18,24. The inconsistency in findings demonstrate that the mechanisms of nest depredation by wild pigs are not well understood.
Wild turkeys (Meleagris gallopavo) are an iconic game species in the United States that were brought to near extinction from overexploitation during the 19th century25,26. Intensive recovery efforts have since ensued, and in 2003 the economic impact of the spring wild turkey season was valued at $1.8 billion USD nationally27. With the expansion of wild pigs throughout the US, many populations of wild turkey are facing a new challenge from nest depredation e.g.18,24. Wild turkeys exhibit spatially-aggregated nesting behavior by selection of specific habitat structure28. This aggregation makes wild turkey nests particularly vulnerable to predators that actively seek nests in habitat where nesting is most likely to occur29,30. For this reason, it has been suggested that removing wild pigs may increase wild turkey recruitment31.
Simulated nests have been widely used to assess different aspects of nest depredation. However, this practice has been criticized for potential to have different depredation rates and attract different predators than natural nests24,32,33. While traditional transect methods for simulated nest studies may result in inflated depredation rates, simulated nest studies which place nests randomly within turkey nesting habitat have shown depredation rates similar to natural turkey nests34. Additionally, simulated nest studies allow for assessment of the influence of the density of nests on nest depredation35.
We used simulated wild turkey nests to evaluate the magnitude of nest predation by wild pigs relative to native nest predators throughout the nesting season in 2016. We also simulated differing densities of nests to evaluate for changes in the frequency of nest predation. Our first objective was compare the frequency of nest consumption by wild pigs to that of native nest predators. Secondly, we assessed the role of seasonality and density of nests in the depredation of nests by wild pigs. Finally, we evaluated differences in consumption of nests by sex and age of wild pigs. Our results will inform efforts to mitigate the impacts of wild pigs on wild turkeys.
Methods
Study area
Our study area was located in south-central Texas (Bexar County), USA (Latitude 29.622 N, Longitude 98.572 W). Specifically, we worked on Camp Bullis Military Reserve, an 11,000 ha military training base that is bordered by both urban and rural ecosystems. Topography on Camp Bullis is typical of the Edwards Plateau: rolling hills with limestone outcrops and rocky soils36. Vegetation is an oak woodland and grassland matrix37,38. Streams and pools are available seasonally during periods of high precipitation. Camp Bullis contains populations of both wild pigs and Rio Grande wild turkey (M. gallopavo intermedia), as well as potential nest predators including raccoons (Procyon lotor), gray foxes (Urocyon cinereoargenteus), coyotes (Canis latrans), bobcats (Lynx rufus), opossums (Didelphis virginiana), striped skunks (Mephitis mephitis), armadillos (Dasypus novemcinctus), corvids, and snakes23,28. In 2006, a suspected release or immigration from surrounding properties of wild pigs occurred on Camp Bullis, resulting in abundant wild pigs throughout the study area (M. L. Cooksey, U.S. Army Environmental Command, personal communication). In 2017, the density of wild pigs was reported to average 3.5 wild pigs per km2 39.
Capture and monitoring of wild pigs
Methods for this study were similarly reported for a simultaneous study23. We captured wild pigs using corral and box traps baited with whole-kernel corn between 15 January and 15 March 2016. We immobilized adult wild pigs (estimated to be >45 kg) using an intramuscular injection of 3.3 mg/kg Telazol® (Zoetis, Parsippany, New Jersey, USA) and 1.5 mg/kg xylazine hydrochloride (Wildlife Pharmaceuticals, Inc., Windsor, Colorado, USA40). We applied uniquely identifiable ear tags (Allflex® A Cattle Tags, Allflex USA Inc., Dallas, Texas, USA) and Global Positioning System (GPS) satellite-transmitting collars (VERTEX PLUS-2 Collar, VECTRONIC Aerospace GmbH, Berlin, Germany) to the immobilized wild pigs. After handling was complete, we intramuscularly injected 0.2 mg/kg yohimbine hydrochloride (Wildlife Pharmaceuticals, Inc., Windsor, Colorado, USA) as a reversal agent40. To ensure spatial coverage of GPS-collared wild pigs across Camp Bullis, we marked ≤2 wild pigs from any social group captured, and we moved traps regularly into locations where wild pigs had not yet been marked. All capture and handling procedures were approved by the Texas A&M University-Kingsville’s Institutional Animal Care and Use Committee (2015-08-20), and all methods were performed in accordance with guidelines outlined in that protocol.
We programmed the GPS collars to record and store locations at 15-minute intervals throughout the study period. Every sixth location was transmitted via Iridium satellite (Iridium Communications, Inc. McLean, Virginia). A drop-off mechanism on each collar was programmed to automatically disconnect the collars on 15 August 2016, after which the full dataset was retrieved from onboard storage. Overall, we attached GPS collars to 35 wild pigs (17 males and 18 females); seven used inaccessible areas (i.e., adjacent private lands), four collars slipped off, one collar failed, and one collared wild pig was killed by an adjacent landowner during the study. Movements of two collared male wild pigs were expansive, covering most of the study area, and could not be targeted for simulated nest deployment. Therefore, we ultimately considered 20 wild pigs (13 males and 7 females) as our primary study animals.
We calculated real-time home ranges for each wild pig every 14 days using transmitted locations from the previous 28 days to inform our placement of simulated nests. Specifically, we calculated the 95% minimum convex polygon home ranges using package adehabitatHR41 in Program R (v3.3.1, www.r-project.org, The R Foundation, Vienna, Austria42;). We used minimum convex polygons because this method conservatively estimates a maximum extent of space-use for each animal43, ensuring our placement of simulated nests would target the entirety of areas used by collared wild pigs.
Nesting habitat suitability model
We built a habitat suitability model to guide our placement of nests within the home ranges of wild pigs using published metrics of nest site selection in wild turkeys44,45. Specifically, we used ERDAS Imagine® (Hexagon Geospatial, Norcross, Georgia) to classify aerial imagery from the 2014 National Agriculture Imagery Program with 1 m resolution (USDA-FSA-APFO Aerial Photography Field Office, Salt Lake City, Utah) from 4 images into woody and herbaceous cover. We conducted an accuracy assessment using 1,200 ground control points and determined the overall accuracy was 89%. We imported classified images into FRAGSTATS v4.246; and used moving windows analyses with a 100-m radius for woody cover on the following metrics: percent land cover, edge density, patch density, and largest patch index. We used ArcGIS (v10.3.1, Environmental Systems Research Institute, Inc., Redlands, CA, USA) to combine the resulting landscape metrics layers and reclassify them into nesting and non-nesting habitat. A location was considered suitable for nesting if the surrounding 100 m had woody cover of 10 to 70%, edge density of >75 m/ha, patch density of >50 patches/100 ha, and a largest patch index of <50%44,45.
Placement, construction, and monitoring of simulated nests
We divided the 20 collared wild pigs into nest density treatments (9 = moderate, and 11 = high) based on non-overlapping space-use so that the density treatments did not overlap. We generated random points using ArcGIS to select the locations of nests within each home range. We randomly placed simulated nests throughout areas identified as nesting habitat within the home ranges of collared wild pigs. We modified selection of the nest sites on the ground to be ≤50 m from each generated random point and best represent true nesting habitat by selecting for small patches of woody cover with dense understory44. We randomly assigned two treatments for density of nests to the home ranges of individual wild pigs, including: 1) moderate density with 25 nests/km2 nesting habitat, and 2) high density with 50 nests/km2 nesting habitat). We based our moderate density treatment on an estimated density of turkeys in oak savanna habitat in south Texas of 13 birds/km2 with an estimated sex ratio of 45% female47,48, and an estimate of 97% initiation of nests by hens49. We corrected the estimated nests/km2 by the proportion of the study area that was designated as nesting habitat according to our habitat suitability model. We doubled the moderate density value to determine our high density treatment and to evaluate the response of wild pigs to extremely high concentrations of wild turkey nests.
We utilized random block design to mimic natural nesting chronology of wild turkeys and assess the role of timing within nesting period while maintaining our experimental density treatments. Specifically, we split the nesting season into three 28-day nesting periods including: (1) early period from 28 March to 24 April, (2) peak period from 25 April to 22 May, and (3) late period from 23 May to 20 June. Therefore, the simulated nesting density for the moderate density treatment varied from: early = 12.5 nests/km2, peak = 25 nests/km2, and late = 12.5 nests/km2. Whereas, the high density treatment varied from: early = 25 nests/km2, peak = 50 nests/km2, and late = 25 nests/km2. Overall, we constructed 384 simulated turkey nests between 28 March and 20 June 2016 within the home ranges of collared wild pigs, including 137 nests in the moderate and 247 in the high density treatments (Fig. 1).

Treatment areas and subsequent distributions of nests within the study area in Bexar County, Texas, USA during the early, peak, and late nesting periods for (a) the high density, and (b) the moderate density treatment areas. Treatment areas were delineated by combining overlapping home ranges (95% minimum convex polygon) of wild pigs to create contiguous areas for each nesting period.
We constructed simulated nests using unwashed domestic chicken (Gallus domesticus) eggs, which are similar in both size and shape to wild turkey eggs50. We placed 10 eggs in a shallow nest bowl lined with leaves. We made no attempt to obstruct the nest with leaf litter. To minimize the addition of human scent, we wore clean rubber soled boots and latex gloves at all nest sites and while handling equipment. We monitored nests for evidence of depredation every seven days for 28 days. If a nest survived 28 days, we removed the nest and placed a new nest at a new random location to maintain density. Similarly, if a nest was depredated, we placed a new nest in a new random location within the same home range to maintain consistent nest density treatments within the home ranges throughout the study.
We randomly selected and monitored a subset of 50 nests using motion detecting cameras (Reconyx PC900 Hyperfire™ Professional Covert IR Cameras, RECONYX, Inc., Holmen, Wisconsin) placed approximately 25 cm above the nest and 2 m away to identify the predators of nests. For nests with >1 species of predator, we identified the primary predator based on which species spent the most time depredating. We also recorded the number of encounters for each species of predator, which we defined as any time a predator approached within ≤1 m of the nest bowl. Finally, we determined the sex and age class (adult or juvenile) of each observed wild pig by examining their secondary sex characteristics, where adults had enlarged, protruding testes or nipples suggesting sexual maturity. We did not identify sex of juvenile wild pigs. We characterized foraging by wild pigs by sex and age class (adult or juvenile) observed participating in depredation.
Data analysis
We used binomial generalized linear models with logit links (glm) in Program R to evaluate whether our study methods seemed to influence our results. First, there are conflicting reports that cameras on nests may repel predators51,52, or possibly alert predators to the location of monitored nests and inflate predation rates24. Therefore, we analyzed the effect of camera monitoring on the probability of depredation to determine if the sample of nests monitored with cameras was representative of those without cameras. Second, we also recognized that eggs may have become more odorous over time, which may increase the rate of depredations. Therefore, we analyzed whether the number of days a nest had been active influenced the probability it was depredated by a wild pig.
We also compared the proportions of nests depredated by each species of predator using Fisher’s exact test. Then, we used binomial glms to evaluate for influences from the nesting period and density of nests, and the interaction of period × density, on the probability of depredation by each species of predator. We also predicted the probability that a nest would be depredated for each nesting period and nest density treatment for all species of nest predators pooled, and for each species of nest predator individually. Additionally, we predicted the probability a nest would be depredated by male and female wild pigs.
We used a binomial hypothesis test to assess the hypothesis that wild pigs will consume a nest once they discover it (i.e., observed on camera at the nest). Specifically, we compared the number of times wild pigs depredated a nest to the number of times wild pigs approached a nest ≤ 1 m but did not depredate the nest. We compared the ratio of foraging events to the number of times a nest was discovered among predator species using Fisher’s exact test. We used the level of α = 0.05 to represent statistical and biological influences for all tests.
Results
We observed that 300 of 384 simulated nests (78.1%) were partially or wholly depredated. Nests with cameras did not have a different depredation rate (75.0%) than those without (78.3%; F1, 382 = 0.285, P = 0.594). We also found no evidence that increasing odors from the eggs increased predation rates through time. The estimated odds that a nest would be depredated by wild pigs decreased by 6.8% for every day a nest was active (F1, 1378 = 10.19, P = 0.001).
From the 52 nests monitored with cameras, we identified 6 wildlife species that depredated nests, including wild pigs, gray foxes, raccoons, coyotes, striped skunks, and a nine-banded armadillo. Wild pigs depredated 15 of the 52 (29%) nests. Gray foxes (21%), raccoons (19%), and coyotes (17%) were also frequent nest predators (Table 1). In all cases, the primary nest predators were also the first nest predators to visit the nests. We also observed nest depredation by an unidentified snake species during manual nest checks but did not observe via camera. We only recorded 1 observation of depredation by an armadillo, therefore we excluded armadillos from further analysis. In seven instances of nest depredations we could not identify the predator.
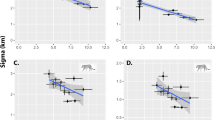
We found that no species of nest predator depredated a higher proportion of nests than another species (two-sided P = 0.079). Similarly, the 95% CIs on the predicted probabilities of depredation overlapped for wild pigs, coyotes, gray foxes, and raccoons (Fig. 2). The interaction of period × density was not significant for any species of nest predator, therefore we excluded the interaction from all models. For all nests, the probability of nest depredation varied by period with the highest probability during peak nesting (F2, 381 = 29.88, P < 0.001; Table 2), but did not vary by density treatment (F1, 382 = 1.437, P = 0.231). The probability that a nest was depredated by a wild pig did not differ by nesting period (F2, 49 = 1.788, P = 0.178; Fig. 3) or by density treatment (F1, 50 = 1.911, P = 0.173; Fig. 4).

Predicted probability that a nest will be depredated by each species of nest predator in both primary and secondary depredation events with 95% confidence intervals for the 2016 wild turkey nesting period in Bexar County, Texas.

Predicted probability that a nest will be depredated by each species of nest predator during each nesting period with 95% confidence intervals for the 2016 wild turkey nesting period in Bexar County, Texas.

Predicted probability that a nest will be depredated by each predator species for each density treatment with 95% confidence intervals for the 2016 nesting period in Bexar County, Texas. For each species, an asterisk indicates a significant (P < 0.05) difference between nest densities.
Wild pigs depredated nests during 58.2% of observed encounters with nests. This was not significantly different from the proportion depredated at discovery by raccoons, gray foxes, or coyotes (two tailed P = 0.071). Probability of depredation given that a nest had been discovered did not vary by sex (F2, 149 = 0.068, P = 0.935) or age class (F2, 149 = 0.302, P = 0.739) of wild pigs. The probability that a nest would be depredated by a male wild pig was substantially greater in the areas with a high versus moderate densities of nests (F1, 50 = 5.288, P = 0.026), but did not differ among nesting periods (F2, 49 = 1.633, P = 0.206; Table 2). The probability that a nest would be depredated by a female wild pig did not differ by density or nesting period (F2, 49 = 0.252, P = 0.778; F1, 50 = 0.159, P = 0.692; Table 2).
Discussion
Our finding that nearly 80% of simulated wild turkey nests being depredated is concerning for wild turkey populations in areas with dense populations of wild pigs. Wild pigs were responsible for depredating as many nests as the most frequently observed native nest-predator, depredating nearly 30% of nests. These results are contrary to previous reports that wild pigs are minor nest predators relative to native species e.g.28,53, which leads us to re-pose the question of whether depredation by wild pigs is additive to that observed by native predators. In the absence of wild pigs, depredation rates of non-artificial wild turkey eggs ranged widely from 18–65%54,55,56, but were always lower than the 80% we observed with artificial eggs and a dense population of wild pigs present. For other ground nesting species (e.g., sea turtles), the addition of wild pigs with native predators did appear to produce additive nest predations57,58. Wild pigs were also frequently the first species to visit the nests in our study, which could likely lead to nest abandonment even if not all eggs were destroyed. Therefore, we suggest that wild pigs may be causing additive nest failures for wild turkeys and more research is needed to test this hypothesis.
Wild turkey nests had substantially higher probabilities of predation by male wild pigs in areas with higher nest densities. This could be indicative of males showing an adaptive response to the pulsed resources of nests, similar to nest depredation behaviors reported in other studies e.g.15,18,57. Wild pigs are known to exploit seasonal resources, such as ground nests, as those resources become available59,60. However, male wild pigs also moved farther distances and had larger home ranges than females in this study area61,62, suggesting that males could have discovered more nests randomly during their increased movements.
Female wild pigs did not appear to increase predation pressure on nests seasonally or in response to increased nest density. Our attempt to mimic natural wild turkey nesting behaviors, by dynamically adjusting nest densities throughout nesting periods, may have reduced our ability to detect responses to either variable. However, our results corroborate similar findings that wild pigs did not actively seek nests regardless of nest density or period23, therefore we do not expect this was the case. More research on the behaviors of wild pigs is needed to conclude whether males exhibited an adaptive response to wild turkey nests, and females did not.
Our results paired with previous studies establish that wild pigs fulfill three detrimental roles for wild turkeys (i.e., consume nests, compete for resources, and alter habitat), and have the potential to reduce turkey populations11,13,63,64. Wild turkeys are not the only ground-nesting species susceptible to nest depredation by wild pigs e.g.15,18,57, particularly as the range of wild pigs continues to expand throughout the US and worldwide. For example, Snow, et al.9 predicts that wild pigs have a high propensity to invade portions of the nesting range of a number of sensitive species including piping plovers (Charadrius melodus), greater sage-grouse (Centrocercus urophasianus), and northern pintails (Anas acuta)65,66,67. Additionally, long-term studies of various North American turtle species indicate that additive mortality of as little as 3% may result in population decline, suggesting turtles may be highly sensitive to depredation by wild pigs68,69. Halting the range expansion of wild pigs may be essential to the conservation of a variety of ground-nesting species.
While coyotes and gray foxes appeared to respond to our density treatments, this relationship is complicated by predator interactions between coyotes and gray foxes which can result in avoidance by gray foxes70. To our knowledge, there is no evidence that wild pigs avoid interactions with other nest predators, thus we did not expect that predator interactions impacted the observed probability a nest would be depredated by wild pigs. Contrarily, if other predators practiced avoidance in response to wild pigs, this could lower the overall depredation rate for wild turkey nests. Avoidance of wild pigs by other nest predators could also introduce bias in nest studies that only monitor nests in areas where wild pigs are known to be active. However, as wild pigs are ubiquitous throughout our study site, we believe our results accurately depict the potential impact of wild pigs on wild turkey nest success where wild pig density is high.
It is important to note that this study utilized simulated turkey nests. While simulated nest studies have shown depredation rates similar to natural turkey nests34, it is difficult to draw concrete conclusions regarding predation rates in the absence of the hen32. The presence of hens has increased predations at nests because the hen can be prey itself54, resulting in nest failure. In addition, the use of surrogate eggs could have led to eggs rotting over time and becoming more detectable, although we found no evidence that this increased detection by wild pigs. Contrarily, we found that probabilities of depredation decreased through time. It is unclear why this relationship existed, but we suspect it was related to some nests being placed in locations that were randomly less detectable by nest predators, despite our best attempts to place all nests equally within turkey nesting habitat. The relationship was likely not related to diminishing human disturbance through time because each nest was revisited once per week. It was also not related to changes in densities of available nests because we replaced any depredated nests with new nests, therefore keeping densities consistent. Ultimately, the use of simulated nests allowed us to evaluate the influence of treatment of nesting densities and periods, which would not have been feasible using natural nests e.g.35.
Conclusions
Invasion of wild pigs into new regions may result in increased nest mortalities for wild turkeys, thus curtailing the expansion of wild pigs is highly important for protecting wild turkeys and other ground-nesting species. Efforts to protect wild turkeys by controlling wild pigs should occur immediately prior to nesting period. Determining whether nest depredation by wild pigs is additive relative to native nest predators represents an important line of future research. Further investigation of potential behavioral responses of male wild pigs to increased availability of nests are also needed to understand the impact of wild pigs on reproductive success of wild turkeys.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Vitense, K., Wirsing, A. J., Tyson, R. C. & Anderson, J. J. Theoretical impacts of habitat loss and generalist predation on predator–prey cycles. Ecological Modelling 327, 85–94 (2016).
Mooney, H. A. & Cleland, E. E. The evolutionary impact of invasive species. Proceedings of the National Academy of Sciences 98, 5446–5451 (2001).
Keiter, D. A., Mayer, J. J. & Beasley, J. C. What is in a “common” name? A call for consistent terminology for nonnative Sus scrofa. Wildlife Society Bulletin 40, 384–387 (2016).
Barrios-Garcia, M. N. & Ballari, S. A. Impact of wild boar (Sus scrofa) in its introduced and native range: a review. Biological Invasions 14, 2283–2300 (2012).
Mayer, J. J. & Brisbin, I. L. Wild pigs in the United States: Their History, Comparative Morphology, and Current Status. (University of Georgia Press, 2008).
Corn, J. L. & Jordan, T. R. Development of the national feral swine map, 1982–2016. Wildlife Society Bulletin 41, 758–763 (2017).
Gipson, P. S., Hlavachick, B. & Berger, T. Range expansion by wild hogs across the central United States. Wildlife Society Bulletin 26, 279–286 (1998).
McClure, M. L. et al. Modeling and mapping the probability of occurrence of invasive wild pigs across the contiguous United States. PloS One 10, e0133771 (2015).
Snow, N. P., Jarzyna, M. A. & VerCauteren, K. C. Interpreting and predicting the spread of invasive wild pigs. Journal of Applied Ecology 54, 2022–2032, https://doi.org/10.1111/1365-2664.12866 (2017).
Timmons, J. B. et al. Feral hog population growth, density and harvest in Texas. Texas A&M AgriLife Extension Service Report.(Texas A&M University: College Station, TX.) Report no. SP-472 (2012).
Ditchkoff, S. S. & Mayer, J. J. In Wild pigs: biology, damage, control techniques, and management. Savanna River National Laboratory, Aiken, SC, USA (eds Mayer, J. J. & Brisbin, I. L.) 105–143 (2009).
Lacki, M. J. & Lancia, R. A. Effects of wild pigs on beech growth in Great Smoky Mountains National Park. Journal of Wildlife Management 50, 655–659 (1986).
Singer, F. J., Swank, W. T. & Clebsch, E. E. Effects of wild pig rooting in a deciduous forest. Journal of Wildlife Management 48, 464–473 (1984).
Taylor, R. B. & Hellgren, E. C. Diet of feral hogs in the western South Texas Plains. The Southwestern Naturalist 42, 33–39 (1997).
Challies, C. N. Feral pigs (Sus scrofa) on Auckland Island: status, and effects on vegetation and nesting sea birds. New Zealand Journal of Zoology 2, 479–490 (1975).
Elsey, R. M., Mouton, E. C. & Kinler, N. Effects of feral swine (Sus scrofa) on alligator (Alligator mississippiensis) nests in Louisiana. Southeastern Naturalist 11, 205–218 (2012).
Stegeman, L. C. The European wild boar in the Cherokee national forest, Tennessee. Journal of Mammalogy 19, 279–290 (1938).
Tolleson, D., Rollins, D., Pinchak, W., Ivy, M. & Hierman, A. Impact of feral hogs on ground-nesting gamebirds. A compendium for resource managers (Hanselka, C. W. & Cadenhead, J. F., eds.). Texas Agricultural Extension Service, San Angel, USA, 76–83 (1993).
Sherry, T. W., Wilson, S., Hunter, S. & Holmes, R. T. Impacts of nest predators and weather on reproductive success and population limitation in a long‐distance migratory songbird. Journal of Avian Biology 46, 559–569 (2015).
Thogmartin, W. E. & Schaeffer, B. A. Landscape attributes associated with mortality events of wild turkeys in Arkansas. Wildlife Society Bulletin 28, 865–874 (2000).
Vangilder, L. D. & Kurzejeski, E. W. Population ecology of the eastern wild turkey in northern Missouri. Wildlife Monographs 130, 1–50 (1995).
Wood, G. W. & Barrett, R. H. Status of wild pigs in the United States. Wildlife Society Bulletin 7, 237–246 (1979).
Sanders, H. N., Hewitt, D. G., Perotto-Baldivieso, H. L., VerCauteren, K. C. & Snow, N. P. Opportunistic predation of wild turkey nests by wild pigs. Journal of WIldlife Management 84, 293–300 (2019).
Dreibelbis, J. et al. Vol. 10 235–242 (Proceedings of the National Wild Turkey Symposium, 2011).
Dickson, J. G. The wild turkey: biology and management. (Stackpole Books, 1992).
Tapley, J. L., Kennamer, J. E. & Abernethy, R. K. Status and distribution of the wild turkey in 2004. Proceedings of the Ninth National Wild Turkey Symposium 9, 21–31 (2005).
National Wild Turkey Federation. The 2003 economic contributions of spring turkey hunting. Southwick Associates, Fernandina Beach, Florida, USA (2003).
Dreibelbis, J. Z. et al. Predation of Rio Grande wild turkey nests on the Edwards Plateau, Texas. The Wilson Journal of Ornithology 120, 906–911 (2008).
Fletcher, K., Aebischer, N. J., Baines, D., Foster, R. & Hoodless, A. N. Changes in breeding success and abundance of ground‐nesting moorland birds in relation to the experimental deployment of legal predator control. Journal of Applied Ecology 47, 263–272 (2010).
Schmidt, K. A. & Whelan, C. J. Nest predation on woodland songbirds: When is nest predation density dependent? Oikos 87, 65–74 (1999).
Petty, B. D. et al. Effects of feral hog control on nest fate of eastern wild turkey in the post oak savannah of Texas. Proceedings of the National Wild Turkey Symposium 9, 169–172 (2005).
Mezquida, E. T. & Marone, L. Are results of artificial nest experiments a valid indicator of success of natural nests? The Wilson Bulletin 115, 270–276 (2003).
Wilson, G. R., Brittingham, M. C. & Goodrich, L. J. How well do artificial nests estimate success of real nests? The Condor 100, 357–364 (1998).
Hernandez, F., Henke, S. E., Silvy, N. J. & Rollins, D. Comparison of success between actual northern bobwhite and wild turkey nests and simulated nests. Wildlife Society Bulletin 29, 1212–1218 (2001).
Hewitt, D. G. & Kirkpatrick, R. L. Ground nest predation and ruffed grouse densities in southwest Virginia. The Wilson Bulletin 105, 521–524 (1993).
Gould, F. Texas plants-a checklist and ecological summary. MP-585/Revised. Tex. Agr. Exp. Sta. Texas A&M Univ., College Station (1975).
Hudler, D. B. Modeling paleolandscapes in central Texas. Dissertation, University of Texas, Austin, USA (2000).
Wills, F. H. Historic vegetation of Camp Bullis and Camp Stanley, southeastern Edwards Plateau, Texas. Texas Journal of science 58, 219–230 (2006).
Snow, N. P. et al. Exposure of a Population of Invasive Wild Pigs to Simulated Toxic Bait containing Biomarker: Implications for Population Reduction. Pest Management Science 75, 1140–1149 (2019).
Sweitzer, R. A. et al. Immobilization and physiological parameters associated with chemical restraint of wild pigs with Telazol® and xylazine hydrochloride. Journal of wildlife diseases 33, 198–205 (1997).
Calenge, C. The package adehabitat for the R software: a tool for the analysis of space and habitat use by animals. Ecological modelling 197, 516–519 (2006).
Mohr, C. O. Table of equivalent populations of North American small mammals. The American Midland Naturalist 37, 223–249 (1947).
Walter, W. & Fischer, J. Manual of applied spatial ecology. State College, PA: Pennsylvania State University (2016).
Dreibelbis, J. Z. Nesting ecology of Rio Grande wild turkeys in the Edwards Plateau of Texas, MS Thesis, Texas A&M University, (2008).
Perotto Baldiviezo, H. L. GIS-based multiple scale study of Rio Grande wild turkey habitat in the Edwards Plateau of Texas, PhD Dissertation, Texas A&M University (2005).
McGarigal, K., Cushman, S. A. & Ene, E. FRAGSTATS v4: spatial pattern analysis program for categorical and continuous maps. University of Massachusetts, Amherst, MA, USA (2012).
Beasom, S. L. Turkey productivity in two vegetative communities in south Texas. Journal of Wildlife Management 34, 166–175 (1970).
Collier, B. A. et al. Variation in brood sex ratios of Texas Rio Grande wild turkeys. Journal of Wildlife Management 71, 1793–1799 (2007).
Keegan, T. W. & Crawford, J. A. Renesting by Rio Grande wild turkeys after brood loss. Journal of Wildlife Management 57, 801–804 (1993).
Hernandez, F., Rollins, D. & Cantu, R. Evaluating evidence to identify ground-nest predators in west Texas. Wildlife Society Bulletin 25, 826–831 (1997).
Richardson, T. W., Gardali, T. & Jenkins, S. H. Review and meta-analysis of camera effects on avian nest success. The Journal of Wildlife Management 73, 287–293 (2009).
Conner, L. M., Rutledge, J. C. & Smith, L. L. Effects of mesopredators on nest survival of shrub-nesting songbirds. The Journal of Wildlife Management 74, 73–80 (2010).
Henry, V. G. Predation on dummy nests of ground-nesting birds in the southern Appalachians. Journal of Wildlife Management 33, 169–172 (1969).
Vander Haegen, W. M., Dodge, W. E. & Sayre, M. W. Factors affecting productivity in a northern wild turkey population. Journal of Wildlife Management 52, 127–133 (1988).
Vangilder, L. D., Kurzejeski, E. W., Kimmel-Truitt, V. L. & Lewis, J. B. Reproductive parameters of wild turkey hens in north Missouri. Journal of wildlife management 51, 535–540 (1987).
Melville, H. I., Conway, W. C., Morrison, M. L., Comer, C. E. & Hardin, J. B. Artificial nests identify possible nest predators of eastern wild turkeys. Southeastern Naturalist 13, 80–92 (2014).
Engeman, R. M., Addison, D. & Griffin, J. Defending against disparate marine turtle nest predators: nesting success benefits from eradicating invasive feral swine and caging nests from raccoons. Oryx 50, 289–295 (2016).
Engeman, R. M. et al. Dramatic and immediate improvements in insular nesting success for threatened sea turtles and shorebirds following predator management. Journal of Experimental Marine Biology and Ecology 395, 147–152 (2010).
Schley, L. & Roper, T. J. Diet of wild boar Sus scrofa in Western Europe, with particular reference to consumption of agricultural crops. Mammal review 33, 43–56 (2003).
Ballari, S. A. & Barrios-García, M. N. A review of wild boar Sus scrofa diet and factors affecting food selection in native and introduced ranges. Mammal Review 44, 124–134 (2014).
Lavelle, M. J. et al. Evaluation of movement behaviors to inform toxic baiting strategies for invasive wild pigs (Sus scrofa). Pest Management Science 74, 2504–2510 (2018).
Snow, N. P. & VerCauteren, K. C. Movement responses inform effectiveness and consequences of baiting wild pigs for population control. Crop Protection 124, 104835 (2019).
Crome, F. H. & Moore, L. Cassowaries in North-Eastern Queensland-Report of a survey and a review and assessment of their status and conservation and management needs. Wildlife Research 17, 369–385 (1990).
Norman, G. W. & Steffen, D. E. Effects of recruitment, oak mast, and fall-season format on wild turkey harvest rates in Virginia. Wildlife Society Bulletin 31, 553–559 (2003).
Aldridge, C. L. et al. Range-wide patterns of greater sage-grouse persistence. Diversity and Distributions 14, 983–994 (2008).
Ivan, J. S. & Murphy, R. K. What preys on piping plover eggs and chicks? Wildlife Society Bulletin 33, 113–119 (2005).
Rice, M. B., Haukos, D. A., Dubovsky, J. A. & Runge, M. C. Continental survival and recovery rates of northern pintails using band-recovery data. Journal of Wildlife Management 74, 778–787 (2010).
Congdon, J. D., Dunham, A. E. & Sels, R. V. L. Demographics of common snapping turtles (Chelydra serpentina): implications for conservation and management of long-lived organisms. American Zoologist 34, 397–408 (1994).
Doroff, A. M. & Keith, L. B. Demography and ecology of an ornate box turtle (Terrapene ornata) population in south-central Wisconsin. Copeia 1990, 387–399 (1990).
Atwood, T. C., Fry, T. L. & Leland, B. R. Partitioning of anthropogenic watering sites by desert carnivores. Journal of Wildlife Management 75, 1609–1615 (2011).
Acknowledgements
We would like to thank C. Blass, J. Halseth, M. Hewitt, C. Kholer, A. Killam, M. Lavelle, M. Matticola, M. Sveine, and E. VanNatta for assistance with data collection. D. Nolte and M. Bodenchuck were instrumental in early discussions and development of this study. We are grateful for logistical support provided by B. Friesenhahn, S. Carasco, and R. Tabor from Joint Base San Antonio Camp Bullis, Guadalupe River State Park, Texas Wildlife Services, the National Wildlife Research Center, and the Caesar Kleberg Wildlife Research Institute at Texas A&M University-Kingsville. We appreciate manuscript edits from M. Garrick. Eggs for nest construction were donated by Cal-Maine Foods, Inc. Mention of commercial products or companies does not represent an endorsement by the US government. Financial support was provided by the United States Department of Agriculture, Animal and Plant Health Inspection Service, Wildlife Services (16-7100-0363-CA), and the Houston Safari Club.
Author information
Authors and Affiliations
Contributions
H.N.S. and N.P.S. contributed to the study design, data collection, analysis, interpretation, and writing of the manuscript. D.G.H., H.L.P. and K.C.V. contributed to the study design and writing of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sanders, H.N., Hewitt, D.G., Perotto-Baldivieso, H.L. et al. Invasive Wild Pigs as Primary Nest Predators for Wild Turkeys. Sci Rep 10, 2625 (2020). https://doi.org/10.1038/s41598-020-59543-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-59543-w
This article is cited by
-
A review of the impacts of invasive wild pigs on native vertebrates
Mammalian Biology (2022)
-
Anthropogenic threats drive spatio-temporal responses of wildcat on Mt. Etna
European Journal of Wildlife Research (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
