
Abstract
Low catch limits for forage species are often considered to be precautionary measures that can help conserve marine predators. Difficulties measuring the impacts of fisheries removals on dependent predators maintain this perspective, but consideration of the spatio-temporal scales over which forage species, their predators, and fisheries interact can aid assessment of whether low catch limits are as precautionary as presumed. Antarctic krill are targeted by the largest fishery in the Southern Ocean and are key forage for numerous predators. Current krill removals are considered precautionary and have not been previously observed to affect krill-dependent predators, like penguins. Using a hierarchical model and 30+ years of monitoring data, we show that expected penguin performance was reduced when local harvest rates of krill were ≥0.1, and this effect was similar in magnitude to that of poor environmental conditions. With continued climate warming and high local harvest rates, future observations of penguin performance are predicted to be below the long-term mean with a probability of 0.77. Catch limits that are considered precautionary for forage species simply because the limit is a small proportion of the species’ standing biomass may not be precautionary for their predators.
Similar content being viewed by others


Introduction
To conserve large fishes, seabirds, and marine mammals, many stakeholders advocate precautionary management of fisheries that target forage species (e.g., krill, anchovies, and sardines). One strategy to conserve predators is to reserve some proportion of their prey1, perhaps by establishing a low catch limit for the fisheries that target the forage populations or stocks2. However, fishing activities may concentrate where target species are profitably caught, potentially increasing local harvest rates above intended levels3. If management fails to prevent concentrated fishing where dependent predators forage, these predators may be impacted despite a low overall catch limit. From an ecosystem perspective, the level of precaution implied by a low catch limit may be better-assessed relative to the time and space scales over which forage species, their predators, and fisheries interact.
Assessing whether catch limits are precautionary from an ecosystem perspective is challenging because the impacts of forage-fish fisheries on predators are difficult to measure4,5,6. This difficulty arises because predators respond to many drivers, including environmental conditions and food-web interactions that are modulated by competition and responses to the availability of alternative prey. Reducing uncertainty to draw unambiguous inference about fisheries impacts on predators requires data that disentangle the effects of fishing from those of the environment and match the temporal and spatial scales of predator life histories, predator-prey interactions, and fishery catches. Data of this nature may not be available at the broad scale of the forage stock, but may be so on smaller scales. Experimental approaches to estimate the effects of fishing are possible7, but such experiments are rare and can be controversial8. An alternative to experimentation is to leverage long-term observational data that capture natural and anthropogenic variations in the focal system. Such long-term studies in locations where the foraging ranges of predators overlap in time and space with locally intense fishing operations are also rare, but the observations provided by these studies may include contrasts that are sufficient to improve inference about the impacts of forage-fish fisheries on dependent predators.
The Antarctic marine ecosystem provides a useful case study for assessing whether a catch limit established at a regional scale is precautionary given locally high harvest rates. Around the Antarctic Peninsula (AP), Antarctic krill (Euphausia superba) are the target of the largest fishery in the Southern Ocean9 and a key forage species for fishes, seabirds, and marine mammals10. The current catch limit for krill in the entire southwest Atlantic sector of the Southern Ocean is 620,000 tons3. This catch limit is spatially divided across four statistical subareas (48.1, 48.2, 48.3 and 48.4; defined by the Commission for the Conservation of Antarctic Marine Living Resources) to reduce the risks of negative impacts on krill-dependent predators in the region. The catch limit in Subarea 48.1, which surrounds the AP, is 155,000 tons, representing <1% of the estimated standing stock of krill (60.3 Mt) in the four subareas3. The catch limit in Subarea 48.1 is achieved regularly, and catches are more concentrated in space and time than ever before9. Adélie (Pygoscelis adeliae), chinstrap (P. antarcticus), and gentoo (P. papua) penguins that breed around the AP feed on a mixed assemblage of krill, fish, and other invertebrates11, but krill are the predominant prey of these seabirds12,13. In recent decades, the standing biomass of krill near the AP has varied by two orders of magnitude due to variations in the physical and biological environment14,15,16,17. The production of penguin populations around the AP is assumed to be linked to the availability of krill18, but previous attempts to relate penguin performance with changes in krill biomass have either been unsuccessful18,19 or based on broad generalizations inferred from trends in penguin recruitment and abundance20,21. The low regional catch limit for krill and the lack of a quantitative relationship between krill biomass and penguin performance around the AP have been used to support arguments that current management of the krill fishery is precautionary3. We suggest, however, that the concentration of krill catches, in time and space, acts to locally increase the vulnerability of penguins to the indirect impacts of fishing despite the low regional catch limit.
We investigated the effects of krill fishing on penguins near the AP consistent with best-practices5. Briefly, we compiled time-series data on 20 indices of penguin performance (e.g., foraging-trip duration, post-hatch breeding success, relative cohort strength, fledging mass) at two field sites in the South Shetland Islands20 and on krill biomass22 in the Bransfield Strait and the northern strata of an established survey grid23 (hereafter the Drake Passage stratum). We used the Oceanic Niño Index (ONI) and the Southern Annular Mode (SAM) as proxy indices of environmental conditions that respectively affect penguins24 and krill25. We used recent tracking data26 to match, in time and space, the penguin-performance indices with the estimates of local (stratum-specific) krill biomass and local harvest rates (stratum-specific krill catch divided by stratum-specific krill biomass). We fitted a hierarchical Bayesian model to the integrated data set, first imputing missing estimates of local krill biomass based on its relationship to the sign of the SAM during summer and then estimating the effects of the ONI, local krill biomass, and local harvest rate on penguin performance. Our integrated data characterize a highly variable ecosystem within which penguin performance has responded to fishing while some of our study populations have declined as others have increased (Fig. 1, with panels a and c respectively adapted from26 and including data from20; see Methods for further detail).

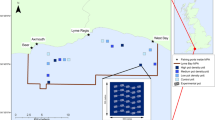
Study area, standardized indices, and trends in penguin abundance. (a) Study area, noting Cape Shirreff (blue dot), Copacabana (red dot), and Subarea 48.1 (dashed line); the thick black line separates the Bransfield Strait and Drake Passage strata, and, within these strata, areas of penguin and fishery overlap (adapted from26 and made available under the Creative Commons CC0 public domain dedication) are respectively colored light red and light blue. (b) Standardized indices of penguin performance during summer and winter at Cape Shirreff (blue) and Copacabana (red), acoustic survey (solid black circles) and imputed (open black circles) estimates of local krill biomass, and the Oceanic Niño Index (thin black line) and Southern Annular Mode (thick black line). (c) Population trends for Adélie (circles), chinstrap (diamonds) and gentoo (squares) penguins (colors as in a) (data prior to 2005 replotted from20). Adapted with permission from Springer Nature: Springer-Verlag Oecologia Divergent responses of Pygoscelis penguins reveal a common environmental driver, J.T. Hinke, K. Salwicka, S.G. Trivelpiece, G.M. Watters, and W.Z. Trivelpiece, © Springer-Verlag 2007.
Results and Discussion
Variations in local krill biomass (LKB), environmental conditions, and local harvest rate (LHR) correlated with penguin performance, but variation in LKB alone had the smallest effect. To aid interpretation of our results, we defined the “best case” as conditions with ONI ≤ −0.5 °C, LKB ≤ 1 Mt, and LHR ≤ 0.01. Although it seems counterintuitive that the best case includes low LKB, some indices of penguin performance decrease when penguins forage on small krill20, and krill biomass is generally greatest when large cohorts of small krill recruit to the adult population16. Relative to the best case, a marginal increase in LKB (to a level >1 Mt) had the smallest effect on penguin performance (Fig. 2) and had the lowest probability (0.7) that expected performance was reduced from the best case (Table 1). The probability that a marginal increase in LKB reduced expected penguin performance below the long-term mean performance was ≤0.04 (Table 1). The insensitivity of penguin performance to variation in LKB corroborates previous failures to parameterize a functional response3,18,19 and seems consistent with view that krill biomass in the AP is generally sufficient to support penguin production27.

Expected performance of penguins attributable to the marginal effects of the Oceanic Niño Index (ONI), local krill biomass (LKB) and local harvest rate (LHR) relative to the “best case”. The best case is ONI ≤ −0.5 °C; LKB ≤ 1 Mt; and LHR ≤ 0.01. The “worst case” is −0.5 °C < ONI < 0.5 °C; LKB > 1 Mt; and LHR ≥ 0.1. The median, interquartile range, and range of the posterior expectations are indicated by each boxplot. Reference lines respectively indicate the median expected performance in the best case (dashed line) and the long-term mean performance.
Warm temperatures (ONI > −0.5 °C) and high LHR (≥0.1) decreased penguin performance, and the effects of these two factors were similar (Fig. 2). The probabilities that the marginal effects of intermediate ONI (−0.5 °C < ONI < 0.5 °C) and high LHR caused expected performance to be less than that of the best case were ≥0.93 (Table 1). The probabilities that these marginal effects caused expected penguin performance to be less than the long-term mean were ≥0.64 (Table 1). In contrast to the best case, we defined the “worst case” as conditions with intermediate ONI, high LKB, and high LHR. Relative to the best case, the worst case caused expected penguin performance to be below the long-term mean with a probability of 0.99 (Table 1). We have little doubt that our study populations responded to both the environment and fishing.
Recent increases in the spatial and temporal concentration of krill catches9,28 provide a plausible mechanism by which the fishery has affected penguins. During the development of the krill fishery, catches contracted from a circumpolar distribution to local hotspots near the AP, the South Orkney Islands, and South Georgia (Fig. 3a, adapted from29). In our Bransfield Strait and Drake Passage strata, annual catches taken prior to 2010 averaged about 34,000 t/yr30, and LHRs ranged from 0 to 0.01. From 2010 through 2016, catches averaged about 121,000 t/yr30, and season-specific LHRs ranged from 0 to 0.32, with rates ≥0.1 occurring 20% of the time in the Bransfield Strait and 10% of the time in the Drake Passage (Supplementary Data S12). For reference, the target harvest rate used to establish precautionary catch limits for the Antarctic krill fishery is 0.0933. All instances when the LHR exceeded 0.1 occurred during winter. Winter catches in the AP region have increased over time and now exceed the total catch taken during summer (Fig. 3b).

Mechanisms by which krill fishing has impacted penguins. (a) Catches (t) contracted from a circumpolar distribution of low catches in the 1980s to higher catches concentrated in the southwest Atlantic sector of the Southern Ocean since 2010. Adapted, with permission from the Secretariat of the Commission for the Conservation of Antarctic Marine Living Resources, from29. (b) Catches, by decade, taken from Statistical Subarea 48.1 (see Fig. 1) during summer (S) and winter (W). (c) Area of overlap (adapted from26 and made available under the Creative Commons CC0 public domain dedication) between foraging predators and the fishery krill fishery (shaded polygons) with surface water movements from drifters (red arrows). Large arrows indicate stronger flows.
Increased catches taken at smaller spatial scales during winter have increased the frequency of poor penguin performance and could reduce future recruitment. If the future is represented by our worst case (Fig. 2), the posterior predictive distributions from our model indicate that any performance index, including recruitment, would be less than its long-term mean with a probability of 0.77 (Table 1). An increased probability of poor recruitment raises the risk of population declines, as has been shown for Adélie penguins31. Although penguins may compensate so that their performance is not reduced after the birds are “challenged”, e.g.32, the population-level consequences of reductions in performance indices other than recruitment (e.g., longer foraging trips, lower adult mass at lay) are not always clear4,5. Given the consequences of poor recruitment and the uncertain impacts from other reductions in performance, the fine-scale concentration of catches taken under the umbrella of a regional catch limit established for an entire year does not seem to be as precautionary as presumed.
Climate change has implications for whether management of the krill fishery will be precautionary in the future. It is reasonable to expect that warmer conditions will become more frequent in the AP33, increasing the probability of negative impacts on penguins. Even as climate changes in the AP, we expect there will be “good” years in which penguin performance is above average. However, because fishery effects on penguin performance are of similar magnitude to those of poor environmental conditions (Fig. 2), performance during good years might nevertheless be reduced if management of the krill fishery unintentionally facilitates high LHRs. If the current management strategy for krill is maintained over the long term, future years like our worst case scenario seem more likely given the propensity of the krill fishery to target hotspots, the regional scale of the current catch limit, and the expectation of continued warming. As noted previously, such conditions are predicted to cause expected penguin performance to fall below its long-term mean with a probability of 0.77 (Table 1).
The recovery of cetacean populations in the Southern Ocean also has implications for whether management of the krill fishery will be precautionary. In the southwest Atlantic sector of the Southern Ocean, humpback whale populations have increased rapidly in the last 30 years, and recent abundance estimates suggest these populations have recovered to levels approximating their pre-exploitation abundances34. On one hand, such recoveries of large cetaceans in the Southern Ocean may reduce the “krill surplus”35 that followed industrial whaling. Alternatively, increased krill consumption by whales may fertilize surface waters and increase the productivity of the ecosystem36,37. Cetaceans forage in areas where our study penguins overlap with the krill fishery38, and the effects of increasing cetacean populations on the structure and function of the ecosystem may affect penguin performance. Determining whether future management of the krill fishery is precautionary will benefit from consideration of such ecosystem perspectives by effectively improving estimates of LKB, and thus LHR.
At least two caveats might bias the LHRs computed here. First, our estimates of LHR might be biased high if the catches were taken from areas in which the throughput of krill was sufficient to replenish local standing stocks. However, penguins foraging from our monitoring sites overlap with the krill fishery in areas where surface currents are relatively weak (Fig. 3c, with panel c adapted from26), and habitat models suggest that chinstrap penguins throughout the AP and South Orkney Islands forage in slow moving water39. Second, the ongoing recovery of whale populations in the Southern Ocean has the potential to impact penguin populations21. If consumption by baleen whales had indeed reduced the standing stocks of krill in areas where our study penguins overlapped with the krill fishery, the LHRs calculated here might be biased low. However, even in this case, the estimated pattern of decreased penguin performance with increased LHR would remain.
Variations in the environment (here indexed by the ONI) and LKB may affect penguin performance in myriad, complex ways. For example, precipitation has been negatively correlated with both chick survival and fledgling mass40,41. Also, when the standing stock of krill is large, small krill are proportionally more abundant22, and, when small krill are more abundant, chinstrap penguins take longer foraging trips20. The literature provides many examples of links between environmental conditions, the characteristics of the prey field, and penguin performance. Our emphasis here has not been to further elucidate or validate such links. Rather we attempt to place the indirect effects of krill fishing in context with these other drivers of penguin performance, which offers a new perspective.
In areas with reduced throughput, fishing may reduce the availably of krill to penguins via absolute reductions in krill biomass (exploitative competition) or from changing the structure of krill swarms or their distribution in the water column (interference competition). Distinguishing these alternatives is difficult in general5 and not possible here. However, concentrated krill fishing in hotspots is associated with sequential reductions in catch per unit fishing effort followed by vessel movements to new areas28. Furthermore, though high LHRs caused reductions in penguin performance (Fig. 2), these occasions were not coincident with biomasses that were also low enough to reduce penguin performance. In combination, our results thus suggest that interference competition has occurred around the AP, whereby krill fishing was sufficient to change the structure or distribution of krill swarms such that both penguins and the fishery itself were impacted. This hypothesis is testable. In our opinion, it is now feasible, using modern acoustic equipment, to estimate what might have been caught without actually bringing krill on deck. Thus, an experiment could be designed in which an acoustic survey is first conducted to estimate LKB; “normal” fishing activities are then conducted, except with open nets so that krill can escape, until more than 10% of the local biomass might have been caught; and penguin performance is simultaneously monitored. It might be useful to compare results from this treatment to those from a reference area in which fishing was conducted with nets that are closed.
Our results demonstrate that it is generally inadvisable to presume the catch limit for a forage species is precautionary simply because the limit is a small proportion of the species’ standing biomass at a regional scale. The dynamic nature of forage species and the fisheries that target them1,4,42,43 means that catch limits applied at coarse spatio-temporal scales can result in unintentionally high LHRs that impact predators. Our integrated, long-term monitoring data demonstrate that this has occurred in the Antarctic krill fishery because the implementation of a presumed precautionary approach did not manage the behavior of the fishery to find and exploit hotspots. When the time and space scales of catch limits are coarser than the scales of predator-prey interactions, it is difficult to achieve ecosystem objectives that include predator conservation6. Impacts on predators can likely be mitigated by spatial and temporal allocation schemes that broadly distribute catches to account for predator-prey-fishery interactions44,45. Failing this, a precautionary approach to conserve predators foraging in fishing hotspots would be to decrease regional catch limits below the levels that are considered precautionary from a single-species perspective.
Methods
Strata
We defined two geographical strata to spatially match season-specific penguin foraging locations with estimates of LKB and LHR. These strata were based on areas where foraging penguins and krill fishing overlap26 and on the availability of krill biomass estimates from research-vessel surveys22,23. We defined the strata as the Drake Passage, combining the western and Elephant Island survey strata of the U.S. AMLR Program22, and the Bransfield Strait (Fig. 1a, adapted from26).
Predictors
We quantified variation in LKB using long-term acoustic survey data22,23 from the AP collected following standard protocols46,47. We computed the density (g/m2) of krill for each nautical mile of survey effort, computed the average density within each stratum, and multiplied those averages by the area of each stratum. Summer surveys were conducted in January and February; winter surveys were conducted in August and September. When more than one survey was conducted in a single season (e.g., January and February of the same year), we averaged the biomass estimates from those surveys. Summer surveys cover the period 1996-2011 (Fig. 1b). Winter surveys are available for 2012, 2014, and 2015 (Fig. 1b).
We computed LHRs by matching catches with temporally and spatially coincident estimates of LKB. Catch data were provided by the Secretariat for the Commission for the Conservation of Antarctic Marine Living Resources. We summed these data by season and stratum and computed LHR as local catch/LKB.
We used two climate indices to characterize seasonal variation in environmental conditions (Fig. 1b). We averaged monthly values of the Southern Annular Mode (SAM48) for winter (April-September) and summer (October-March) indices. We also computed seasonal averages of the Oceanic Niño Index (ONI)49. We assigned summer averages of the SAM and ONI to the second calendar year of the split year in each austral summer (e.g., the average value for October 2004 - March 2005 was assigned to 2005).
We categorized all four predictors. When the SAM is in a positive (negative) phase, westerly winds are stronger and shifted towards the pole50, with a resulting increase (decrease) in air temperature51 and precipitation52 on the western side of the AP. We thus categorized the SAM according to its sign. The ONI defines La Niña and El Niño events when sea-surface temperature anomalies near the equator are respectively ≤−0.5 °C and ≥0.5 °C. Such events interact with the SAM to affect primary production and krill near the Antarctic Peninsula15, so we binned the ONI into three categories (ONI ≤ −0.5 °C; −0.5 °C < ONI < 0.5 °C; and ONI ≥ 0.5 °C) using these two temperature thresholds. Estimates of LKB are imprecise22 and may be biased by a variety of factors, including the throughput of krill through our strata53. Nevertheless, the surveys distinguish periods of relative krill scarcity and abundance. Acoustic estimates of LKB ranged in magnitude from 104 to 107 t. In contrast, krill catches taken within our strata were reported by fishing vessels, treated as known, and ranged in magnitude from ≤103 to 105 t. Given differences in the uncertainties and magnitudes of local biomasses and catches, we categorized estimates of LKB using a threshold of 1 × 106 t (1 Mt) and of LHR using thresholds of 0.01 and 0.1. The threshold for LKB evenly splits the observed orders of magnitude in krill biomass, and that for LHR provides three categories of fishing that reflect low harvest rates, harvest rates up to that used to set catch limits for krill (0.093), and fishing above that level.
Response – penguin performance
We used monitoring parameters based on observations collected during 1982-2016 at two field camps in the South Shetland Islands to quantify variations in penguin performance. Unless otherwise noted, methods of data collection have been described previously20,54. All methods were performed in accordance with relevant guidelines and regulations, including field methods that were reviewed and approved by the University of California San Diego Institutional Animal Care and Use Committee (ID: S05480) and authorized under U.S. Antarctic Conservation Act permits (ID: ACA 2017-012). Some parameters reflect winter conditions (mean clutch-initiation date, mean female and male masses at lay, mean egg density, and relative cohort strength; Fig. 1b). Other parameters reflect conditions during the summer breeding season (fledgling mass, foraging-trip duration, and post-hatch breeding success; Fig. 1b). In all but one case, these parameters integrate over days to months. Relative cohort strength integrates over a period ≥1 year, but we assumed that most of its variation is attributable to survival during the first few months of independence20. We transformed mean clutch-initiation dates and foraging-trip durations so that larger values would indicate better performance. The former parameter was transformed to days prior to 31 December (earlier clutch initiations indicate better performance); the latter was transformed to hours <60 hours (shorter trips indicate better performance). Relative cohort strength (proportion of banded penguins resighted in their breeding colonies) and post-hatch breeding success (proportion of chicks crèched) were logit-transformed. Mean egg density was calculated from the total mass and volume of eggs from 2-egg clutches. Egg data were collected during the first week after clutch completion by weighing and measuring the maximum length and width of each egg from 50 nests per species. Egg volume was estimated from its empirical relationship55 to egg length and width. We standardized the penguin performance indices specific to each combination of monitoring parameter (transformed or otherwise), species, and site to have zero mean and unit variance (Fig. 1b).
We matched penguin-performance indices to estimates of LKB and LHR in the Bransfield Strait or Drake Passage strata using results from multi-year tracking studies that identify when and where penguins foraged26. We matched the winter and summer performance indices for Adélie and gentoo penguins breeding at Copacabana (Fig. 1a, adapted from26) with LKB and LHR in the Bransfield Strait. We matched winter indices for chinstrap penguins breeding at Cape Shirreff (Fig. 1a, adapted from26) with LKB and LHR in the Drake Passage, while those for gentoo penguins breeding at the same site were matched with predictors in the Bransfield Strait. After matching, we pooled performance indices within 18 bins defined by all combinations of the categorized predictors.
Model
We fitted an analysis of variance model with two components. The first component imputes missing estimates of LKB because krill surveys were not conducted every year. We modeled LKB as a function of stratum and the sign of the SAM during summer.
\({K}_{ij}\) is the expected value of \(\mathrm{ln}\,LKB\) in stratum \(i\) given sign \(j\) of the SAM during summer, and \({\phi }^{2}\) is the variance of \(\mathrm{ln}\,LKB\). We truncated this likelihood with a lower limit equal to the catch taken from stratum \(i\) (so \(LK{B}_{ij}\) would not be less than \({{\rm{catch}}}_{i}\)) and an upper limit equal to 100 Mt (so \(LK{B}_{ij}\) would be less than twice the estimate of krill biomass used to manage the krill fishery46). We used results from the acoustic surveys to specify prior distributions for the imputation model.
\({\bar{k}}_{ij}\) is the mean \(\mathrm{ln}\,LKB\) computed from survey observations in stratum \(i\) given sign \(j\) of the SAM, and \({s}_{ij}\) is the standard deviation of these log-biomass estimates. We predicted missing estimates of LKB \((LK{B}_{ij}^{\ast })\) from Eq. (1) using the sign of the SAM for summers when acoustic surveys were not conducted. Missing estimates of LHR, denoted as \(LH{R}^{\ast }\), were estimated from \(\,LK{B}_{ij}^{\ast }\).
We did not impute missing values for winter.
In the second component of our model, we treated all indices of penguin performance as exchangeable observations, and modeled performance (\(P\)) as a function of categorized ONI (\(o\)), LKB (\(b\)), and LHR (\(h\)).
\(P\) is the expected performance, and \({\sigma }^{2}\) is the residual variance in performance. \(\alpha \) quantifies mean performance across the set of predictor categories, and the parameters \({\beta }_{1},{\beta }_{2},\,{\beta }_{3},{\beta }_{4},\,{\rm{and}}\,{\beta }_{5}\,\)quantify the degree to which each predictor causes expected performance to deviate from this mean. We specified prior distributions that limit inference to the study populations and the predictor bins considered here.
We specified a half-Cauchy prior distribution for the standard deviation of penguin performance and a uniform hyperprior for the scale parameter (\(\omega \)) of the half-Cauchy56,57.
We specified a design matrix with sum-to-zero contrasts that carries uncertainties in \(LK{B}^{\ast }\) and \(LH{R}^{\ast }\) through the model.
We used JAGS58, via R59, to sample from the posterior distributions of the model parameters. After 250,000 adaptive and 500,000 burn-in iterations, we sampled 5,000 points (retaining values from every 25th iteration during a further 125,000 iterations) from the posterior distributions characterized by three Monte Carlo chains initiated at different points in the parameter space. We evaluated our model with a variety of diagnostics. Diagnostics, data files, and the code needed to replicate our analysis are provided as Supplemental Information. Three R packages are needed to run our code: rjags60, coda61, and ggmcmc62.
Data availability
All model code and data used to conduct this study are included in Supplementary Information files of this published article.
References
Pikitch, E. et al. Little fish, big impact: managing a crucial link in ocean food webs. (Lenfest Ocean Program, Washington, D.C. 2012).
Constable, A. J., de la Mare, W. K., Agnew, D. J., Everson, I. & Miller, D. Managing fisheries to conserve the Antarctic marine ecosystem: practical implementation of the Convention on the Conservation of Antarctic Marine Living Resources (CCAMLR). ICES J. Mar. Sci. 57, 778–791 (2000).
Hill, S. L. et al. Is current management of the Antarctic krill fishery in the Atlantic sector of the Southern Ocean precautionary? CCAMLR Sci. 23, 31–51 (2016).
Hilborn, R. et al. When does fishing forage species affect their predators? Fish. Res. 191, 211–221 (2017).
Sydeman, W. J. et al. Best practices for assessing forage fish fisheries-seabird resource competition. Fish. Res. 194, 209–221 (2017).
Hunsicker, M. E. et al. Functional responses and scaling in predator-prey interactions of marine fishes: contemporary issues and emerging concepts. Ecol. Lett. 14, 1288–1299 (2011).
Pichegru, L., Gremillet, D., Crawford, R. J. M. & Ryan, P. G. Marine no-take zone rapidly benefits endangered penguin. Biol. Lett. 6, 498–501 (2010).
Cherry, M. African penguins put researchers in a flap. Nature 514, 283 (2014).
Nicol, S., Foster, J. & Kawaguchi, S. The fishery for Antarctic krill - recent developments. Fish. Fish. 13, 30–40 (2012).
Everson, I. Marine Interactions in Antarctic Ecology, volume 2. (ed. Laws, R. M.) 463–490 (Academic Press 1984).
Polito, M. J., Lynch, H. J., Naveen, R. & Emslie, S. D. Stable isotopes reveal regional heterogeneity in the pre-breeding distribution and diets of sympatrically breeding Pygoscelis spp. penguins. Mar. Ecol. Prog. Ser. 421, 265–277 (2011).
Volkman, N. J., Presler, P. & Trivelpiece, W. Diets of Pygoscelid penguins at King George Island, Antarctica. Condor 82, 373–378 (1980).
Miller, A. K. & Trivelpiece, W. Z. Cycles of Euphausia superba recruitment evident in the diet of Pygoscelid penguins and net trawls in the South Shetland Islands, Antarctica. Polar Biol. 30, 1615–1623 (2007).
Loeb, V. et al. Effects of sea-ice extent and krill or salp dominance on the Antarctic foodweb. Nature 387, 897–900 (1997).
Saba, G. K. et al. Winter and spring controls on the summer food web of the coastal West Antarctic Peninsula. Nat. Commun. 5, 4318, https://doi.org/10.1038/ncomms5318 (2014).
Atkinson, A. et al. Krill (Euphausia superba) distribution contracts southward during rapid regional warming. Nat. Clim. Change 9, 142–147 (2019).
Flores, H. et al. Impact of climate change on Antarctic krill. Mar. Ecol. Prog. Ser. 458, 1–19 (2012).
Scientific Committee for the Conservation of Antarctic Marine Living Resources. Report of the twenty-second meeting of the Scientific Committee. CCAMLR, Hobart; https://www.ccamlr.org/en/meetings/27 (2003).
Hewitt, R. P. et al. Options for allocating the precautionary catch limit of krill among small-scale management units in the Scotia Sea. CCAMLR Sci. 11, 81–97 (2004).
Hinke, J. T., Salwicka, K., Trivelpiece, S. G., Watters, G. M. & Trivelpiece, W. Z. Divergent responses of Pygoscelis penguins reveal a common environmental driver. Oecologia 153, 845–855 (2007).
Trivelpiece, W. Z. et al. Variability in krill biomass links harvesting and climate warming to penguin populations in Antarctica. Proc. Natl. Acad. Sci. USA 108, 7625–7628 (2011).
Reiss, C. S., Cossio, A. M., Loeb, V. & Demer, D. A. Variations in the biomass of Antarctic krill (Euphausia superba) around the South Shetland Islands, 1996-2006. ICES J. Mar. Sci. 65, 497–508 (2008).
Reiss, C. S. et al. Overwinter habitat selection by Antarctic krill under varying sea-ice conditions: implications for top predators and fishery management. Mar. Ecol. Prog. Ser. 568, 1–16 (2017).
Croxall, J. P., Trathan, P. N. & Murphy, E. J. Environmental change and Antarctic seabird populations. Science 297, 1510–1514 (2002).
Loeb, V. J. & Santora, J. A. Climate variability and spatiotemporal dynamics of five Southern Ocean krill species. Prog. Oceanogr. 134, 93–122 (2015).
Hinke, J. T. et al. Identifying risk: concurrent overlap of the Antarctic krill fishery with krill-dependent predators in the Scotia Sea. PLoS One 12, e0170132, https://doi.org/10.1371/journal.pone.0170132 (2017).
Sailley, S. F. et al. Carbon fluxes and pelagic ecosystem dynamics near two western Antarctic Peninsula Adélie penguin colonies: an inverse model approach. Mar. Ecol. Prog. Ser. 492, 253–272 (2013).
Santa Cruz, F., Ernst, B., Arata, J. A. & Parada, C. Spatial and temporal dynamics of the Antarctic krill fishery in fishing hotspots in the Bransfield Strait and South Shetland Islands. Fish. Res. 208, 157–166 (2018).
Commission for the Conservation of Antarctic Marine Living Resources. Krill Fishery Report 2018. (CCAMLR, Hobart); https://www.ccamlr.org/en/document/publications/krill-fishery-report-2018. (2018).
Commission for the Conservation of Antarctic Marine Living Resources. Statistical Bulletin, vol. 30. (CCAMLR, Hobart); https://www.ccamlr.org/en/publications/statistical-bulletin (2018).
Hinke, J. T., Trivelpiece, S. G. & Trivelpiece, W. Z. Variable vital rates and the risk of population declines in Adélie penguins from the Antarctic Peninsula region. Ecosphere 8, e01666, https://doi.org/10.1002/ecs2.1666 (2017).
Ballard, G. et al. Responding to climate change: Adélie Penguins confront astronomical and ocean boundaries. Ecology 91, 2056–2069 (2010).
Clarke, A. et al. Climate change and the marine ecosystem of the western Antarctic Peninsula. Philos. T. R. Soc. Lon. B. 362, 149–166 (2006).
Zerbini, A. N. et al. Assessing the recovery of an Antarctic predator from historical exploitation. R. Soc. Open Sci. 6, 190368, https://doi.org/10.1098/rsos.190368 (2019).
Laws, R. Seals and whales of the Southern Ocean. Philos. Trans. R. Soc. Lond. 279, 81–96 (1977).
Roman, J. & McCarthy, J. J. The whale pump: marine mammals enhance primary productivity in a coastal basin. PLoS One 5, 10, https://doi.org/10.1371/journal.pone.0013255 (2010).
Nicol, S. et al. Southern Ocean iron fertilization by baleen whales and Antarctic krill. Fish. Fish. 11, 203–209 (2010).
Weinstein, B. G., Double, M., Gales, N., Johnston, D. W. & Friedlander, A. S. Identifying overlap between humpback whale foraging grounds and the Antarctic krill fishery. Biol. Cons. 210, 184–191 (2017).
Trathan, P. N. et al. Managing fishery development in sensitive ecosystems: Identifying penguin habitat use to direct management in Antarctica. Ecosphere 9, e02392, https://doi.org/10.1002/ecs2.2392 (2018).
Chapman, E. W., Hofmann, E. E., Patterson, D. L., Ribic, C. A. & Fraser, W. R. Marine and terrestrial factors affecting Adélie penguin Pygoscelis adeliae chick growth and recruitment off the western Antarctic Peninsula. Mar. Ecol. Prog. Ser. 436, 273–289 (2011).
Cimino, M. A., Fraser, W. R., Patterson-Fraser, D. L., Saba, V. S. & Oliver, M. J. Large-scale climate and local weather drive interannual variability in Adélie penguin chick fledging mass. Mar. Ecol. Prog. Ser. 513, 253–268 (2014).
Atkinson, A. et al. Sardine cycles, krill declines, and locust plagues: revisiting ‘wasp-waist’ food webs. Trends Ecol. Evol. 29, 309–316 (2014).
Cury, P. et al. Small pelagics in upwelling systems: patterns of interaction and structural changes in “wasp-waist” ecosystems. ICES J. Mar. Sci. 57, 603–618 (2000).
Plagányi, E. E. & Butterworth, D. S. The Scotia Sea krill fishery and its possible impacts on dependent predators: modeling localized depletion of prey. Ecol. App. 22, 748–761 (2012).
Watters, G. M., Hill, S. H., Hinke, J. T., Matthews, J. & Reid, K. Decision-making for ecosystem-based management: evaluating options for a krill fishery with an ecosystem dynamics model. Ecol. App. 23, 710–725 (2013).
Scientific Committee for the Conservation of Antarctic Marine Living Resources. Report of the twenty-ninth meeting of the Scientific Committee. CCAMLR, Hobart; https://www.ccamlr.org/en/meetings/27 (2010).
Calise, L. & Skaret, G. Sensitivity investigation of the SDBWA Antarctic krill target strength model to fatness, material contrasts and orientation. CCAMLR Sci. 18, 97–122 (2011).
U.S. National Weather Service. https://www.cpc.ncep.noaa.gov/products/precip/CWlink/daily_ao_index/aao/monthly.aao.index.b79.current.ascii
U.S. National Weather Service. https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v4.shtml
Thompson, D. W. J. & Wallace, J. M. Annular modes in the extratropical circulation. Part I: month-to-month variability. J. Clim. 13, 1000–1016 (2000).
Marshall, G. J., Orr, A., van Lipzig, N. P. M. & King, J. C. The impact of a changing Southern Hemisphere Annular Mode on Antarctic Peninsula summer temperatures. J. Clim. 19, 5388–5404 (2006).
Marshall, G. J., Thompson, D. W. J. & van den Broeke, M. R. The signature of Southern Hemisphere atmospheric circulation patterns in Antarctic precipitation. Geophys. Res. Lett. 44, 11580–11589 (2017).
Hofmann, E. E. & Murphy, E. J. Advection, krill, and Antarctic marine ecosystems. Antarct. Sci. 16, 487–499 (2004).
Commission for the Conservation of Antarctic Marine Living Resources. CCAMLR ecosystem monitoring program standard methods. (CCAMLR, Hobart, 2014).
Hoyt, D. F. Practical methods of estimating volume and fresh weight of bird eggs. Auk 96, 73–77 (1979).
Gelman, A. Prior distributions for variance parameters in hierarchical models (comment on article by Browne and Draper). Bayesian Anal. 1, 515–534 (2006).
Polson, N. G. & Scott, J. G. On the half-cauchy prior for a global scale parameter. Bayesian Anal. 7, 887–902 (2002).
Plummer, M. JAGS: A Program for Analysis of Bayesian Graphical Models Using Gibbs Sampling. Proceedings of the 3rd International Workshop on Distributed Statistical Computing. March 20-22, Vienna, Austria. ISSN 1609-395×(2003).
R Core Team. R: a language and environment for statistical computing. https://www.R-project.org (2018).
Plummer, M., Stukalov, A. & Denwood, M. Bayesian graphical models using MCMC. http://mcmc-jags.sourceforge.net (2016).
Plummer, M., Best, N., Cowles, K. & Vines, K. CODA: convergence diagnosis and output analysis for MCMC. R News 6, 7–11 (2006).
Fernández-i-Marín, X. ggmcmc: analysis of MCMC samples and Bayesian inference. J. Stat. Softw. 70, 1–20 (2016).
Acknowledgements
This research was conducted as part of the U.S. Antarctic Marine Living Resources Program of the National Oceanic and Atmospheric Administration (NOAA). We thank the numerous field teams that collected data on krill and penguins over the last 3 + decades. We thank A.M. Cossio for synthesis of acoustic survey data, W.Z. and S.G. Trivelpiece for a life-long commitment to the study of penguins in Antarctica, D.J. Krause and M. Mangel for comments on an early version of this manuscript, and G. Cutter for being a naturalist. Borrego Springs, California provided the desert heat required to cook this manuscript. Finally, the Editor and two referees provided extensive suggestions to improve our paper.
Author information
Authors and Affiliations
Contributions
G.M.W. designed the study; G.M.W., J.T.H. and C.S.R. conducted research, analyzed data, and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Watters, G.M., Hinke, J.T. & Reiss, C.S. Long-term observations from Antarctica demonstrate that mismatched scales of fisheries management and predator-prey interaction lead to erroneous conclusions about precaution. Sci Rep 10, 2314 (2020). https://doi.org/10.1038/s41598-020-59223-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-59223-9
This article is cited by
-
Whale recovery and the emerging human-wildlife conflict over Antarctic krill
Nature Communications (2024)
-
CS-PHOC: weekly census counts of Southern Ocean phocids at Cape Shirreff, Livingston Island
Scientific Data (2024)
-
Availability to predators and a size structure of the Antarctic krill Euphausia superba in the 48.1 CCAMLR subarea
Scientific Reports (2024)
-
Using latent behavior analysis to identify key foraging areas for Adélie penguins in a declining colony in West Antarctic Peninsula
Marine Biology (2024)
-
Climate change impacts on Antarctic krill behaviour and population dynamics
Nature Reviews Earth & Environment (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
