
Abstract
Freshwater and marine fish have been important components of human diets for millennia. The Great Lakes of North America, their tributaries and smaller regional freshwater bodies are important Native American fisheries. The ethnohistorical record, zooarchaeological remains, and isotopic values on human bone and tooth collagen indicate the importance of fish in fourteenth- through seventeenth-century ancestral Wendat diets in southern Ontario, which is bordered by three of the Great Lakes. Maize (Zea mays ssp. mays) was the primary grain of Native American agricultural systems in the centuries prior to and following sustained European presence. Here we report new Bayesian dietary mixing models using previously published δ13C and δ15N values on ancestral Wendat bone and tooth collagen and tooth enamel. The results confirm previous estimates from δ13C values that ancestral Wendat diets included high proportions of maize but indicate much higher proportions of fish than has previously been recognized. The results also suggest that terrestrial animals contributed less to ancestral Wendat diets than is typically interpreted based on zooarchaeological records.
Similar content being viewed by others

Introduction
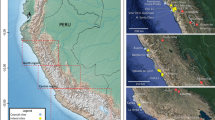
Various lines of evidence indicate freshwater and marine fish have been important components of human diets for millennia1,2,3. Fish biomarkers extracted from pottery fabric and encrusted food residues have shown that fish were cooked in the early pottery in several areas of the world4,5,6,7. Fish skeletal elements and scales are routinely recovered from prehistoric archaeological sites worldwide when fine-scale recovery is implemented8,9,10,11. Isotopic analyses of human bone frequently indicate fish consumption12,13. Ethnohistoric and ethnographic accounts attest to the dietary importance of fish in the recent past14,15. These lines of evidence indicate that the Great Lakes of North America, their tributaries, and surrounding smaller freshwater bodies were important Native American fisheries16. Southern Ontario, Canada, borders Lake Ontario, Lake Erie, Lake Huron and its Georgian Bay, and the St. Lawrence River (Fig. 1), and fish were important dietary components of Native Americans occupying this area17.

Locations of archaeological sites with samples used in the analyses. Figure 1 does not contain copyrighted material. The map was produced in ArcGIS v 10.6 at the New York State Museum, Albany, NY by compiling GIS data (shapefiles) obtained from publicly available data from Statistics Canada, the U.S. Census, and the United States Geological Survey.
Maize (Zea mays ssp. mays) was the primary grain of Native American agricultural systems in eastern North America in the centuries prior to and after European arrivals18. Phytoliths and starch recovered from directly AMS-dated charred cooking residues adhering to the interiors of pottery sherds indicate that maize was introduced into the lower Great Lakes region, presumably including southern Ontario, by ca. cal. 300 BC19,20,21. Macrobotanical remains in the form of charred maize kernels and cob fragments become evident in the southern Ontario archaeological record by ca. cal. AD 50022 and become ubiquitous by ca. cal. AD 900–100018.
Southern Ontario was home to the historical Iroquoian Wendat (Huron) Confederacy, which began to form during the sixteenth century AD23. Over the course of the fourteenth through the beginning of the seventeenth centuries AD, ancestral Wendat communities moved northward from the north shore of Lake Ontario, and eventually converged on an area between the south shore of Georgian Bay to the north and Lake Simcoe to the south (Fig. 1)23. During these centuries, communities coalesced into large villages and towns and formed several nations, which in turn formed the Wendat Confederacy24. Multiple lines of evidence indicate that these communities relied substantially on maize for food, and that maize-based agriculture facilitated the coalescence of communities and the convergence of Wendat communities into a geographically restricted area25.
Maize is a plant that uses the C4 photosynthetic pathway with archaeological δ13C values in northeastern North America ranging from −15.1 to −7.4‰, while most native terrestrial plants and agricultural crops in the region utilize the C3 photosynthetic pathway with archaeological δ13C values in northeastern North America ranging from −28.6 to −23.3‰26,27,28,29,30. As a result, consumers of maize and their consumers have δ13C values that are higher than consumers of predominantly C3 photosynthetic pathway plants and their consumers. δ13C analyses of human bone collagen and apatite suggest that maize consumption began to increase gradually in southern Ontario by ca. cal. AD 50027. Using δ13C values on human bone and teeth, Pfeiffer et al.28,29 suggest that by the cal. fourteenth to fifteenth centuries AD, maize was a dominant source of calories for ancestral Wendat individuals, remained so in the sixteenth century AD, and increased in the seventeenth century AD. Using a linear mixing model with δ13C values they estimate that maize contributed >50% of ancestral Wendat diets during those centuries, and perhaps as much as 65%28. However, they suggest that elevated δ13C values may also reflect the consumption of freshwater fish because fish bone from ancestral Wendat archaeological sites have δ13C values that range mostly between −21‰ and −16‰28. Pfeiffer et al.28 report a mean dog collagen δ13C value of −11.1 ± 0.9‰ and suggest its consumption may have affected human values. Less attention has been paid to stable nitrogen isotopes (δ15N) in terms of ancestral Wendat diets, given that the primary concern of isotopic studies in southern Ontario has been to trace maize consumption histories and has thus focused on δ13C values.
Consumers of animal flesh have higher δ15N values than do plants and primary consumers of plants. Omnivores with higher proportions of terrestrial plants in their diets will have lower δ15N than omnivores with higher proportions of animal flesh in their diets. Of note is that despite the dominance of maize in the diet, δ15N values in Pfeiffer et al.’s28 ancestral Wendat samples are highly elevated. Because maize, like other terrestrial plants, is assumed to have δ15N values < 3.0‰, Pfeiffer et al.28 suggest diets that included large amounts of terrestrial resources should produce δ15N values ≤ 10‰. Given that δ15N values of ancestral Wendat individuals are generally >10‰, Pfeiffer et al., like others27,30, suggest that ancestral Wendat diets included high-trophic-level freshwater fish.
There is ample evidence from the archaeological and ethnohistoric records that freshwater fish was an important contributor to ancestral and historical Wendat diets15,17,31. Based on the ethnohistorical record, Heidenrich15 estimated that fish accounted for 9% of seventeenth century AD daily Wendat diets. Fish bone generally comprises large proportions of faunal assemblages on seventeenth-century and earlier ancestral Wendat sites on which fine-scale recovery was used, and this evidence suggests varied fishing strategies based on time of year and settlement location32,33. While ancestral Wendat δ15N values suggest fish were important components of diets, no quantitative assessments have been made based on those values28,31,34. On the basis of δ15N values Pfeiffer et al.28 suggest that proportions of fish in diets were highest during the fourteenth and seventeenth centuries AD, when ancestral Wendat communities were closest to the north shore of Lake Ontario and the south shore of Georgian Bay, respectively. They suggest dietary fish proportions decreased during the fifteenth and sixteenth centuries AD when communities were located at greater distances from these two fisheries. Hawkins et al.17 suggest their analysis of a large dataset of fish remains supports Pfeiffer et al.’s conclusion, and the contribution of fish to diets was particularly low during the sixteenth century AD before recovering to substantial contributions during the seventeenth century AD.
Based on differences in δ13C and δ15N values in bone collagen and dentine collagen samples, Pfeiffer et al.28 suggest that there were differences between adult and child diets, respectively. Specifically, they suggest maize comprised larger proportions of child than adult diets, while animal protein comprised smaller proportions of child than adult diets.
Potentially complicating interpretations of δ15N values is the role of white-tailed deer (Odocoileus virginianus) in subsistence34. While deer is sometimes considered to have been an important component of ancestral Wendat diets28, deer bone frequently does not comprise high percentages of faunal remain assemblages35. Seventeenth-century AD ethnohistoric records indicate that deer was not a major component of Wendat diets—deer were scarce in the area occupied by the Wendat at that time15. To the contrary, good deer habitat occurs to the south where ancestral Wendat communities lived prior to consolidation south of Georgian Bay in the seventeenth century AD35.
Here we apply a Bayesian dietary mixing model, MixSIAR36, to calculate the proportions of food sources in ancestral Wendat diets. MixSIAR allows multiple isotopic tracers (e.g., δ13C and δ15N) and uses a Markov Chain Monte Carlo simulation to model the probability of proportions of food sources in a consumer’s diet based on the isotopic values of the food sources (e.g., maize, fish) and the consumer (human). As opposed to identifying point estimates of the proportions of certain foods in diets (e.g., maize) from the analysis of single isotopic tracers, Bayesian mixing models, including MixSIAR used here, incorporate the uncertainty in the isotopic values of the food sources, the consumer, and the trophic enrichment factor (TEF) between the food sources and the consumer in the model to ultimately produce a probability of proportions of food sources in a consumer’s diet. An assumption of these models is that the isotopic tracer values of the food sources are representative of the diets of the sampled humans.
We use δ13C and δ15N values in human dentine and bone collagen and tooth enamel from Pfeiffer et al.’s28,29,34 ancestral Wendat research, recently obtained isotope values for maize from ancestral Wendat sites, and a large database of terrestrial animal (n = 404) and freshwater fish (n = 170) bone collagen isotope data from southern Ontario archaeological sites, which we compiled from several sources28,37,38,39,40, to model fourteenth through seventeenth century AD ancestral Wendat diets. We specifically test four hypotheses generated by the results of Pfeiffer et al.’s28 interpretations of their stable isotope analyses and Hawkins et al.’s17 interpretation of ancestral Wendat fish consumption based on faunal remains:
H1: The proportions of maize in diets was consistent from the fourteenth through sixteenth centuries AD and increased in the seventeenth century AD.
H2: Diets incorporated proportionately more fish during the fourteenth and seventeenth centuries AD than during the intervening centuries, with fish being particularly minor dietary components in the sixteenth century AD diets.
H3: Fish exploitation was largely concentrated on high trophic-level species.
H4: Children, as reflected in dentine collagen and tooth enamel isotope values, had higher proportions of maize in their diets than did adults as reflected in bone collagen isotope values.
This article provides results of the first application of multi-tracer Bayesian dietary modeling on a large series of Native American isotopic values from eastern North America. The combined modeling of δ13C and δ15N values results in the first quantitative estimates of fish proportions in ancestral Wendat diets based on archaeological isotopic data.
Results
All model output and code are presented in the Supplementary Information File. Results of the three-source (i.e., maize, fish, and terrestrial species) MixSIAR models by century are summarized in Fig. 2 and Table 1. Isotopes are differentially incorporated into tissues such that the isotopic values in collagen (bone or dentine) derive mainly from ingested protein, while values in tooth enamel derive from the whole diet41,42,43,44. This routing of isotopes may influence interpretations of diet in populations if they are based solely on one tissue type. Additionally, these tissues may reflect different periods in life depending on when the tissue forms. Tooth enamel and dentine collagen form during tooth development and do not readily remodel, so the δ13C and δ15N values are reflective of intake at younger ages. Bone collagen, however, does remodel and, in general, the isotopic values reflect intake of an older individual. By comparing values from different tissues, such as tooth enamel and bone collagen, a more comprehensive understanding of dietary source proportions is derivable.

Mean values of three-source models by century for (a) dentine collagen, (b) bone collagen, (c) enamel-dentine collagen, (d) enamel-bone collagen. Red circles = fish, orange triangles = maize, green squares = terrestrial animals. Mean values of three fish categories in the five-source models by century for (e) dentine collagen and (f) bone collagen. Green circles = high δ15N fish, light blue circles = medium δ15N fish, and dark blue circles = low δ15N fish. Error bars are 1 standard deviation.
Mean values for the models evince similar trends, that is, the proportion of maize consumed increases while the proportion of fish consumed decreases over time. The dentine collagen-based model exhibits an increase in maize proportions through time, a steady decrease in fish proportions, and a slight increase in terrestrial animal proportions in the seventeenth century AD. The bone collagen model exhibits an increase in maize proportions from the fourteenth to fifteenth century AD followed by a plateau, an overall decrease in fish proportions, but with a plateau in the fifteenth and sixteenth centuries AD, and an increase in terrestrial animal proportions but with a plateau in the fifteenth and sixteenth centuries AD. Overall the dentine collagen model mean values exhibit higher maize and lower terrestrial animal proportions than the means of the bone collagen model. As would be expected these results are substantially the same when the results of individual site models are placed in relative chronological order (Supplementary File S1.4.3).
In both tooth enamel models (Fig. 2c,d), maize proportions increase with a plateau during the fifteenth and sixteenth centuries, while fish proportions decrease with a plateau in the fifteenth and sixteenth centuries. The tooth enamel carbonate models calculate higher mean proportions of maize and lower proportions of fish in Wendat diets, over the four centuries when comparing the collagen (bone and dentine) to tooth enamel model results (Fig. 2). This implies that the collagen models slightly under represent the proportion of maize in the diet. These results indicate we can reject H1—the dentine model exhibits a consistent increase in maize, the bone collagen model does not show an increase in the seventeenth century AD, and the tooth enamel models show an overall increase in maize with a plateau between the fifteenth and sixteenth centuries AD.
The means of all models evince decreases in fish proportions through time. None of the models indicate an increase in fish proportions in the seventeenth century AD contrary to Pfeiffer et al.’s28 interpretation of δ15N values and Hawkins et al.’s17 interpretation of fish remains. As a result, H2 can be rejected. Results for fish proportion means from the five-source models are summarized in Fig. 2 and Table 2. We cannot reject H3 given that high δ15N fish have the highest dietary proportion estimates for fish throughout the sequence in both models.
Based on the preceding, it is evident that child and adult diets were different in terms of maize and terrestrial animal proportions, but not for fish. As a result, H4 cannot be rejected–child and adult diets did differ in the respective samples.
Discussion
Previous δ13C analyses of human bone and teeth and the ethnohistorical record suggest maize was a major component of ancestral Wendat diets, perhaps comprising upwards of 65% of the diet. Bayesian analysis shows that maize dietary proportions increased in importance from the fourteenth to seventeenth centuries AD, at least in adults. As evidenced by the dentine collagen model, maize appears to also have been an increasingly important component of child diets through the sequence. The bone- and dentine-collagen models suggest that about 50% of the protein derived from maize, and the tooth enamel models imply mean maize proportions over 60% throughout most of the analyzed sequence supporting the assessment that maize at times was the major component of ancestral Wendat diets. One estimate of fish consumption of 9% is based on seventeenth-century ethnohistorical records15. The Bayesian three-source models indicate fish proportions decreased through time, but the mean calculations for proportion both the dentine collagen and enamel models of fish in the diet was always above 9% across the fourteenth through seventeenth centuries. In adult diets decreased dietary fish proportion was replaced by terrestrial animals. The three-source model using enamel δ13C follow the same trends for child diets with increasing maize and decreasing fish proportions. The decreased proportions of fish in child diets was replaced primarily by increased maize, combined with terrestrial animals in the seventeenth century AD in both the dentine collagen and enamel models. Previous δ15N analyses suggest high-trophic-level fish, that typically have high δ15N values, were an important component of ancestral Wendat diets. The five-source collagen models show that the high δ15N fish group, which includes salmonids, is the dominant fish source in all bone and dentine collagen model iterations.
These results suggest that maize and fish were regular components of ancestral Wendat diets throughout the sequence. Both constituted large proportions of the diet contributing to the formation of dentine and bone collagen as well as tooth enamel. Other foods, such as terrestrial animals contributed substantially less. Deer flesh apparently was not the primary source of animal protein, rather, fish was the primary source, probably due to an easier access to fish compared to deer. Large fish like salmons can be passively trapped from spring to fall with wooden hoop nets while deer were captured using fence and pens and needed a significant number of experienced people to build the structures and drive the deer into them15. The increased proportion of terrestrial animals in the seventeenth-century AD bone collagen model may reflect deer consumption, which would be at odds with poor deer habitat in the region south of Georgian Bay. In earlier centuries, with villages located in favorable deer habitat, the mean dietary proportion of terrestrial animals contributing to collagen formation was <10%, while during the fourteenth century, i.e. when villages were more concentrated close to Lake Ontario and closer larger fish sources, the mean value is ~2%. This latter value may also be explained if deer harvesting was seasonal as recorded in the ethnohistoric record and deer flesh was limited to immediate consumption or associated feasts. Similarly, the ethnohistoric record indicates dog flesh, which has high δ13C, was generally eaten ceremonially and at feasts rather than frequently15. This occasional consumption would not influence human isotopic values.
As with previous isotopic analyses of human tissue in southern Ontario27,28,29,30, our analysis is limited by small sample sizes. However, the present analysis contributes to our understandings of ancestral Wendat diets by allowing us to assign quantitative estimates of fish and terrestrial animal dietary proportions for the individuals represented in the available samples. As in other areas of the world, while changes occurred over the centuries considered here, freshwater fish was a critical component of these individuals’ diets, while terrestrial protein was of substantially less importance. These results should inform future inferences about ancestral Wendat diets based on zooarchaeological analyses.
Methods
For the Bayesian models, we used the MixSIAR GUI v. 3.1 in R v. 3.5.245. Stable carbon (δ13C) and nitrogen (δ15N) isotope values from southern Ontario archaeological sources were used as the isotopic tracers. Data for animal bone recovered primarily from southern Ontario archaeological sites dating from AD 900–1700 (source) (Supplementary Information File S.2) were compiled from published sources28,37,38,39,40. Nine of the assayed fish bones are from the St. Lawrence Valley in Quebec and six are nineteenth-century sites in southern Ontario38. Data for humans (consumers) were obtained from Pfeiffer et al.28 (Supplementary Information File S3). To utilize the human tooth enamel data in the Bayesian models that employ collagen stable isotope data for sources and trophic enrichment factors, we converted the tooth enamel δ13C values using a Δ13Cenamel-collagen value of −5.7‰ (Supplementary Information File S1.2). (Zhu and Sealy, 2018). For the δ15N values, we used the dentine and bone collagen δ15N of the same teeth. Maize δ13C and δ15N values were obtained from ancestral Wendat site samples, reported here for the first time (Table S.4.1) and are the first published δ15N values on maize in the region. Three-source and five-source models were run and presented on data from each site separately, then run and presented combining data from sites into their respective centuries as in Pfeiffer et al.28. Since Hawkins et al. (2019) found no zooarchaeological evidence of Atlantic salmon (Salmo salar) in 17th c. Wendat localities, we ran additional models of the 17th century data in MixSIAR removing samples of this species from the fish source (3 source model) and the high δ15N fish source (5 source model). Results of these additional models showed extremely limited changes in the modelled proportions of sources consumed (Supplementary Information File S1.4.1.1, S1.4.6.1) The three-source model groups included maize (n = 14), all fish (n = 170), and all terrestrial prey (n = 404). The five-source-model groups had the all fish group split into three different groups based on statistically significant differences in δ15N values among fish taxa. The statistically significant differences in δ15N values also capture known ecological differences in the analyzed fish species with the high δ15N fish encompassing offshore fish species, while the medium and low δ15N fish groups contain nearshore fish species. Therefore, the five-source model groups included, maize (n = 14), all terrestrial prey (n = 404), high δ15N fish (n = 104), medium δ15N fish (n = 43), and low δ15N fish (n = 23) (see Supplementary Information File S1.1). Terrestrial prey included the following species: bear (Ursus americanus, n = 50), beaver (Castor canadensis, n = 3), deer (Odocoileus virginianus, n = 191), fox (Vulpes vulpes, n = 6), groundhog (Marmota monax, n = 18), muskrat (Ondatra zibithecus, n = 3), porcupine (Erethizon dorsatum, n = 3), rabbit/hare (Leporidae, n = 8), raccoon (Procyon lotor, n = 47), squirrel (Sciurus carolinensis, n = 14), and turkey (Meleagris gallopavo, n = 61). While obviously not capturing all potential variation in isotope values for the various taxa and isotopic groupings, the source value dataset is large enough to avoid modeling issues arising from small source sample sizes46,47 in the isotopic groups used here. Additionally, we found no statistically significant differences between salmon data from Ontario Ancestral Wendat sites and the remaining salmon samples implying baseline values from different watersheds were similar for the fish sources in the models (Supplementary Information File S.1.1). Because the maize δ15N values were higher than expected we carried out a series of experiments to determine if charring affects maize isotopic values and adjusted the maize δ15N values used in the Bayesian modeling accordingly (Supplementary Information File S.4). Sources were input as means and standard deviations. Collagen to collagen source (food) to consumer (human) trophic enrichment factors (TEF) used in the collagen models were +1.1‰ ± 0.2‰ for δ13C and +3.8‰ ± 1.1‰ δ15N48,49. The maize (source) to consumer (human) TEF for collagen was +5.0‰ ± 0.1‰ for δ13C, and +3.0‰ ± 0.1‰ for δ15N43. The Bayesian mixing models met the criteria of the Gelman-Rubin and Geweke diagnostics. Results are reported as means and standard deviations as well as percent credible interval range (posterior probabilities) from 0.025 to 0.975.
Data availability
All data generated or analyzed during this study are included in this published article and its Supplementary Information File.
References
Boethius, A. & Ahlström, T. Fish and resilience among Early Holocene foragers of southern Scandinavia: A fusion of stable isotopes and zooarchaeology through Bayesian mixing modelling. J. Archaeol. Sci. 93, 196–210, https://doi.org/10.1016/j.jas.2018.02.018 (2018).
Cristiani, E. et al. Dental calculus and isotopes provide direct evidence of fish and plant consumption in Mesolithic Mediterranean. Sci. Rep. 8, 8147, https://doi.org/10.1038/s41598-018-26045-9 (2018).
Lopes, M. S. et al. The path towards endangered species: prehistoric fisheries in southeastern Brazil. PLOS ONE 11, e0154476, https://doi.org/10.1371/journal.pone.0154476 (2016).
Craig, O. E. et al. Ancient lipids reveal continuity in culinary practices across the transition to agriculture in northern Europe. Proc. Natl. Acad. Sci. USA 108, 17910–17915, https://doi.org/10.1073/pnas.1107202108 (2011).
Craig, O. E. et al. Earliest evidence for the use of pottery. Nature 496, 351–354, https://doi.org/10.1038/nature12109 (2013).
Heron, C. et al. Cooking fish and drinking milk? Patterns in pottery use in the southeastern Baltic, 3300–2400 cal BC. J. Archaeol. Sci. 63, 33–43, https://doi.org/10.1016/j.jas.2015.08.002 (2015).
Taché, K. & Craig, O. E. Cooperative harvesting of aquatic resources and the beginning of pottery production in north-eastern North America. Antiquity 89, 177–190, https://doi.org/10.15184/aqy.2014.36 (2015).
Harris, T. E., Lilley, K. A. & Walter, R. K. The varying role of vertebrae in Pacific fishbone analysis: comparing tropical versus temperate midden assemblages. Int. J. Osteoarchaeol. 27, 1038–1047, https://doi.org/10.1002/oa.2628 (2017).
Jones, T. L., Gobalet, K. W. & Codding, B. F. The archaeology of fish and fishing on the central coast of California: the case for an under-exploited resource. J. Anthropol. Archaeol. 41, 88–108, https://doi.org/10.1016/j.jaa.2015.11.002 (2016).
McKechnie, I. & Moss, M. L. Meta-analysis in zooarchaeology expands perspectives on Indigenous fisheries of the Northwest Coast of North America. J. Archaeol. Sci. Rep. 8, 470–485, https://doi.org/10.1016/j.jasrep.2016.04.006 (2016).
Schmölcke, U. et al. Neolithic fish remains from the freshwater shell midden Riņņukalns in northern Latvia. Environ. Archaeol. 21, 325–333, https://doi.org/10.1179/1749631415Y.0000000011 (2016).
Richards, M. P., Karavanić, I., Pettitt, P. & Miracle, P. Isotope and faunal evidence for high levels of freshwater fish consumption by Late Glacial humans at the Late Upper Palaeolithic site of Šandalja II, Istria, Croatia. J. Archaeol. Sci. 61, 204–212, https://doi.org/10.1016/j.jas.2015.06.008 (2015).
Salazar-García, D. C., Fontanals-Coll, M., Goude, G. & Subirà, M. E. To “seafood” or not to “seafood”? An isotopic perspective on dietary preferences at the Mesolithic-Neolithic transition in the Western Mediterranean. Quat. Int. 470, 497–510, https://doi.org/10.1016/j.quaint.2017.12.039 (2018).
Lovis, W. A. & Hart, J. P. Fishing for dog food: ethnographic and ethnohistoric insights on the freshwater reservoir in northeastern North America. Radiocarbon 57, 557–570, https://doi.org/10.7183/0002-7316.78.3.536 (2015).
Heidenreich, C. Huronia: A History and Geography of the Huron Indians, 1600–1650. (McClelland & Stewart 1971).
Cleland, C. E. The inland shore fishery of the northern Great Lakes: its development and importance in prehistory. Amer. Antiq. 47, 761–784, https://doi.org/10.2307/280281 (1982).
Hawkins, A. L., Needs-Howarth, S., Orchard, T. J. & Guiry, E. J. Beyond the local fishing hole: a preliminary study of pan-regional fishing in southern Ontario (ca. 1000 CE to 1750 CE). J. Archaeol. Sci. Rep. 24, 856–868, https://doi.org/10.1016/j.jasrep.2019.03.007 (2019).
Hart, J. P. & Lovis, W. A. Reevaluating what we know about the histories of maize in northeastern North America: a review of current evidence. J. Archaeol. Res. 21, 175–216, https://doi.org/10.1007/s10814-012-9062-9 (2013).
Gates St-Pierre, C. & Thompson, R. G. Phytolith evidence for the early presence of maize in southern Quebec. Amer. Antiq. 80, 408–415, https://doi.org/10.7183/0002-7316.80.2.408 (2015).
Hart, J. P., Brumbach, H. J. & Lusteck, R. Extending the phytolith evidence for early maize (Zea mays ssp. mays) and squash (Cucurbita sp.) in central New York. Amer. Antiq. 72, 563–583, https://doi.org/10.2307/40035861 (2007).
Albert, R. K., Kooiman, S. M., Clark, C. A. & Lovis, W. A. Earliest microbotanical evidence for maize in the northern Lake Michigan basin. Amer. Antiq. 83, 345–355, https://doi.org/10.1017/aaq.2018.10 (2018).
Crawford, G. W., Smith, D. G. & Bowyer, V. E. Dating the entry of corn (Zea mays) into the lower Great Lakes region. Amer. Antiq. 62, 112–119, https://doi.org/10.2307/282382 (1997).
Williamson, R. F. The archaeological history of the Wendat to A.D. 1651: an overview. Ont. Archeol. 94, 3–64, https://www.ontarioarchaeology.org/resources/Publications/OA%20No%2094%20Journal_2014.pdf#page9 (2014).
Birch, J. Coalescent communities: settlement aggregation and social integration in Iroquoian Ontario. Amer. Antiq. 77, 646–670, https://doi.org/10.7183/0002-7316.77.4.646 (2012).
Birch, J. & Williamson, W. F. The Mantle Site: An Archaeological History of an Ancestral Wendat Community (Altamira 2013).
Hart, J. P., Lovis, W. A., Schulenberg, J. K. & Urquhart, G. R. Paleodietary implications from stable carbon isotope analysis of experimental cooking residues. J. Archaeol. Sci. 34, 804–813, https://doi.org/10.1016/j.jas.2006.08.006 (2007).
Harrison, R. G. & Katzenberg, M. A. Paleodiet studies using stable carbon isotopes from bone apatite and collagen: examples from Southern Ontario and San Nicolas Island, California. J. Anthropol. Archaeol. 22, 227–244, https://doi.org/10.1016/S0278-4165(03)00037-0 (2003).
Pfeiffer, S., Sealy, J. C., Williamson, R. F., Needs-Howarth, S. & Lesage, L. Maize, fish, and deer: investigating dietary staples among ancestral Huron-Wendat villages, as documented from tooth samples. Amer. Antiq. 81, 515–532, https://doi.org/10.1017/S0002731600003978 (2016).
Pfeiffer, S., Williamson, R. F., Sealy, J. C., Smith, D. G. & Snow, M. H. Stable dietary isotopes and mtDNA from Woodland period southern Ontario people: results from a tooth sampling protocol. J. Archaeol. Sci. 42, 334–345, https://doi.org/10.1016/j.jas.2013.11.008 (2014).
Schwarcz, H. P., Melbye, J., Katzenberg, M. A. & Knyf, M. Stable isotopes in human skeletons of southern Ontario: reconstructing palaeodiet. J. Archaeol. Sci. 12, 187–206, https://doi.org/10.1016/0305-4403(85)90020-2 (1985).
Hawkins, A. L., Malleau, K. & Elliott, D. A consideration of participants in Huron-Wendat subsistence strategies across the pre-contact and early colonization periods. J. Archaeol. Sci. Rep. 20, 873–880, https://doi.org/10.1016/j.jasrep.2017.10.024 (2018).
Needs-Howarth, S. & Thomas, S. C. Seasonal variation in fishing strategies at two Iroquoian village sites near Lake Simcoe, Ontario. Environ. Archaeol. 3, 109–120, https://doi.org/10.1179/env.1998.3.1.109 (1998).
Needs-Howarth, S. J. Native Fishing in the Great Lakes: A Multidisciplinary Approach to Zooarchaeological Remains from Precontact Iroquoian Villages Near Lake Simcoe, Ontario (University of Groningen 1999), http://hdl.handle.net/11370/f1686b3f-156f-4361-a3af-47f2d9ec623a.
Pfeiffer, S., Sealy, J. C., Williamson, R. F., Forrest, C. & Lesage, L. Patterns of weaning among ancestral Huron-Wendat communities, determined from nitrogen isotopes. Amer. Antiq. 82, 244–261, https://doi.org/10.1017/aaq.2016.36 (2017).
Needs-Howarth, S. & Hawkins, A. L. Tending and drive hunting: A density-mediated attrition model can explain age profiles of white-tailed deer at Iroquoian village sites. Environ. Archaeol. 22, 175–188, https://doi.org/10.1080/14614103.2016.1154238 (2017).
Stock, B. C. et al. Analyzing mixing systems using a new generation of Bayesian tracer mixing models. PeerJ 6, e5096, https://doi.org/10.7717/peerj.5096 (2018).
Booth, L. A Stable Isotope Analysis of Faunal Remains from Special Deposits on Ontario Iroquoian Tradition Sites (The University of Western Ontario 2014), https://ir.lib.uwo.ca/etd/2644.
Guiry, E. J. et al. Lake Ontario salmon (Salmo salar) were not migratory: A long-standing historical debate solved through stable isotope analysis. Sci. Rep. 6, 36249, https://doi.org/10.1038/srep36249 (2016).
Morris, Z. H. Reconstructing Subsistence Practices of Southwestern Ontario Late Woodland Peoples (A.D. 900-1600) Using Stable Isotopic Analyses of Faunal Material (The University of Western Ontario 2015). https://ir.lib.uwo.ca/etd/2921.
Morris, Z., White, C., Hodgetts, L. & Longstaffe, F. Maize provisioning of Ontario Late Woodland turkeys: isotopic evidence of seasonal, cultural, spatial and temporal variation. J. Archaeol. Sci. Rep. 10, 596–606, https://doi.org/10.1016/j.jasrep.2016.06.017 (2016).
Sealy, J., Armstrong, R. & Schrire, C. Beyond lifetime averages: tracing life histories through isotopic analysis of different calcified tissues from archaeological human skeletons. Antiquity 69, 290–300, https://doi.org/10.1017/S0003598X00064693 (1995).
Clementz, M. T. & Koch, P. L. Differentiating aquatic mammal habitat and foraging ecology with stable isotopes in tooth enamel. Oecol. 129, 461–472, https://doi.org/10.1007/s004420100745 (2001).
Ambrose, S. H., Butler, B. M., Hanson, D. B., Hunter‐Anderson, R. L. & Krueger, H. W. 1997. Stable isotopic analysis of human diet in the Marianas Archipelago, Western Pacific. Am. J. Phys. Anthropol. 104, 343–361, https://10.1002/(SICI)1096-8644(199711)104:3<343::AID-AJPA5>3.0.CO;2-W.
Zhu, M. & Sealy, J. Multi‐tissue stable carbon and nitrogen isotope models for dietary reconstruction: Evaluation using a southern African farming population. Am. J. Phys. Anthropol. 168, 145–153, https://doi.org/10.1002/ajpa.23731 (2019).
Stock, B. C., Semmens, B. X. & MixSIAR, G. U. I. User Manual. Version 3, 1, https://doi.org/10.5281/zenodo.1209993 (2016).
Makarewicz, C. A. & Sealy, J. Dietary reconstruction, mobility, and the analysis of ancient skeletal tissues: expanding the prospects of stable isotope research in archaeology. J. Archaeol. Sci. 56, 146–158 (2015).
Phillips, D. L. et al. Best practices for use of stable isotope mixing models in food-web studies. Can. J. Zool. 92, 823–835, https://doi.org/10.1139/cjz-2014-0127 (2014).
Bocherens, H. et al. Reconstruction of the Gravettian food-web at Předmostí I using multi-isotopic tracking (13C, 15N, 34S) of bone collagen. Quat. Int. 359, 211–228, https://doi.org/10.1016/j.quaint.2014.09.044 (2015).
Ledogar, S. H., Feranec, R. S. & Zuhlke, J. M. Isotopic evidence for broad diet including anadromous fish during the mid-Holocene in northeastern North America. J. Archaeol. Sci. Rep. 19, 505–512, https://doi.org/10.1016/j.jasrep.2018.03.029 (2018).
Acknowledgements
We thank Susan Pfeiffer, University of Toronto, for the ancestral Wendat isotope values and comments on a draft of this article; Louis Lesage, Huron-Wendat Nation, and Judith Sealy, University of Cape Town, for comments on the paper; Carla S. Hadden and Jennifer Birch, University of Georgia for providing the isotope values on maize; and Susan Winchell-Sweeney at the New York State Museum for Fig. 1.
Author information
Authors and Affiliations
Contributions
R.S.F. and J.P.H. designed the study and wrote the manuscript. R.S.F. performed the Bayesian analyses and maize kernel experiments. J.P.H. prepared Fig. 2. R.S.F. compiled the Supplementary Information File. R.S.F. and J.P.H. reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
41598_2019_53076_MOESM1_ESM.docx
Fish and maize: Bayesian mixing models of fourteenth- through seventeenth-century AD ancestral Wendat diets, Ontario, Canada
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Feranec, R.S., Hart, J.P. Fish and maize: Bayesian mixing models of fourteenth- through seventeenth-century AD ancestral Wendat diets, Ontario, Canada. Sci Rep 9, 16658 (2019). https://doi.org/10.1038/s41598-019-53076-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-53076-7
This article is cited by
-
Human and dog Bayesian dietary mixing models using bone collagen stable isotope ratios from ancestral Iroquoian sites in southern Ontario
Scientific Reports (2023)
-
Multiproxy Analysis of Adhered and Absorbed Food Residues Associated with Pottery
Journal of Archaeological Method and Theory (2022)
-
Ancient dog diets on the Pacific Northwest Coast: zooarchaeological and stable isotope modelling evidence from Tseshaht territory and beyond
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
