
Abstract
In our most recent study, we found that in Tanzania infection with Treponema pallidum (TP) subsp. pertenue (TPE) is present in four different monkey species. In order to gain information on the diversity and epidemiological spread of the infection in Tanzanian nonhuman primates (NHP), we identified two suitable candidate genes for multi-locus sequence typing (MLST). We demonstrate the functionality of the MLST system in invasively and non-invasively collected samples. While we were not able to demonstrate frequent interspecies transmission of TPE in Tanzanian monkeys, our results show a clustering of TPE strains according to geography and not host species, which is suggestive for rare transmission events between different NHP species. In addition to the geographic stability, we describe the relative temporal stability of the strains infecting NHPs and identified multi-strain infection. Differences between TPE strains of NHP and human origin are highlighted. Our results show that antibiotic resistance does not occur in Tanzanian TPE strains of NHP origin.
Similar content being viewed by others

Introduction
Nonhuman primates (NHPs) in Africa are naturally infected with Treponema pallidum subsp. pertenue (TPE)1, the bacterium causing human yaws. In our most recent study, we found that in Tanzania infection with TPE is present in four different NHP species (olive baboon (Papio anubis), yellow baboon (Papio cynocephalus), vervet monkey (Chlorocebus pygerythrus), and blue monkey (Cercopithecus mitis))2. Moreover, we showed that infection is geographically widespread within Tanzania. Although we confirmed infection by serology and PCR, the data were insufficient to describe the epidemiology of the disease. Further insights into the inter- and intraspecies spread of the TPE bacterium in Tanzanian NHPs will contribute to our understanding of transmission pathways and pathogen maintenance, which are crucial elements for the identification of a functional disease reservoir3. The chance that NHPs infected with TPE are a potential source for human infection has been discussed for tropical Africa4. However, naturally occurring transmission from NHPs to humans and vice versa has not been confirmed by current data, although phylogenetic analyses of whole genome sequences from TPEs of human and NHP origin suggest a rapid initial radiation of the ancestor of TPE across the different primate taxa, including humans1.
Molecular typing is used to accurately distinguish between different strains of T. pallidum (TP) for epidemiological and surveillance analysis. The method has been extensively applied to the syphilis–causing bacterium (subsp. pallidum, TPA)5 where it is used to describe its spatial, e.g.6,7,8,9, and temporal, e.g.10, subtype composition. Two recent studies suggested new typing systems for human yaws11,12. It was unclear though, whether the existing molecular typing systems for TPA or TPE of human origin can be applied to TP strains originating from NHPs.
In the current study, we identified suitable candidate genes for multi-locus sequence typing (MLST) in TP samples of NHP origin and investigated strain diversity of the NHP infecting strains in Tanzania. We hypothesized that interspecies transmission in NHPs is ongoing. Moreover, we show that our typing system can be applied to samples from other regions of Africa and to analyze TP in non-invasively collected fecal samples.
Materials and Methods
Ethical statement
No animals were handled specifically for this study. The ethical statement for the Tanzanian NHP samples has been published elsewhere2,13. Lesion swabs from Ethiopian grivet monkeys (Chlorocebus aethiops) were taken as part of a research investigation conducted by AA under the Ethiopian Wildlife Conservation Authority reference number 15ET-0000-BS-01. Noninvasively collected fecal samples of western lowland gorillas (Gorilla gorilla gorilla) from the southern part of Odzala-Kokoua National Park (OKNP) originate from a collaboration signed under a MoU between the Foundation Odzala-Kokoua, the German Primate Center, and the Institute of Vertebrate Biology, Czech Academy of Sciences in November 2017.
Design of the multi-locus sequence typing system
In order to identify most suitable candidate genes for MLST in TPE strains of NHP origin, we used 23 available complete and draft genome sequences of TPE from both human and NHPs from Africa and the Pacific regions (Table S1). Several criteria were applied to obtain most suitable gene loci for TPE MLST. First, we identified the most variable genes with accumulated single nucleotide variants (SNVs) in short DNA fragments (genes containing the highest SNVs frequency per kbp) and, at the same time, with potential ability to distinguish all strains used for this analysis (containing 22 and more variable sites; Table S1). We identified six candidate genes (Table 1) and compared the resolution power of phylogenetic trees based on genome-wide data and phylogenetic trees based on sequences of individual genes.
With this approach, we propose a new MLST scheme for TPE strains of NHP origin, based on sequencing of two variable loci (TP0548 and TP0488). The typing scheme is able to reveal 70% of whole genome resolution. Further details on the identification of most variable genes, resolution power, and the selection of most suitable typing loci are provided in the Technical Appendix.
Samples included into the study and DNA extraction
Our study used TP positive DNA samples from 85 NHPs of six different species and three African countries (Tables 2 and S3). The samples originated from previously published2,13 and ongoing research investigations. The different methods of DNA extraction are presented in the Technical Appendix.
DNA target enrichment
Before MLST, DNA extracted from fecal samples was enriched for bacterial DNA using the Looxter Enrichment Kit (Analytik Jena, Jena, Germany) following the manufacturer’s protocol.
Polymerase chain reactions
Multi-Locus Sequence Typing system
TP0548: Amplification of a fragment of the TP0548 gene was achieved using a nested PCR. The two-step PCR amplified a 1,065-bp long fragment of the target gene. Amplification and sequencing primers were used as reported elsewhere14. Briefly, the 50-µl reaction volume comprised 25 µl of the 2x Universe buffer (Universe High Fidelity Hot Start DNA Polymerase Kit, Biotool, Munich, Germany), 17 µl RNAase free water, 2 µl of each 10 µM primer, 1 µl DNA polymerase (1 U/µl), 1 µl of the dNTP mix (10 mM each), and 2 µl template DNA, independent of DNA concentration. Amplification was performed in a SensoQuest Labcycler using the following thermocycler conditions: pre-denaturation at 95 °C for 3 min, followed by 40 and 30 cycles, respectively, each with 95 °C for 15 sec, 48 °C for 15 sec, and 72 °C for 30 sec. Each of the PCR runs ended with a post-extension step at 72 °C for 5 min.
TP0488: The PCR amplified a 837 bp-long fragment of the TP0488 gene using 5′-CCC TGC GCA CCA AGC TC-3′ and 5′-ACA CAG GCC CCA TAA ACT-3′ primers. Briefly, the 51-µl reaction volume comprised 45 µl Platinum PCR Super Mix High Fidelity (ThermoFisher Scientific, Munich, Germany), 2 µl of each 10 µmol/L primer, and 2 µl template DNA, independent of DNA concentration. Amplification was performed in a SensoQuest Labcycler using the following thermocycler conditions: pre-denaturation at 94 °C for 2 min, followed by 80 cycles each with 94 °C for 15 sec, 59 °C for 15 sec, and 68 °C for 1 min.
Additional gene targets
TP0619: Although the TP0619 locus is not part of the newly designed typing system, it was amplified to further discriminate between TPA and Treponema pallidum subsp. endemicum (TEN) strains. We performed the PCR as described previously2. PCR conditions included a pre-denaturation at 95 °C for 3 min, followed by 40 cycles each with 95 °C for 15 sec, 55 °C for 15 sec, and 72 °C for 30 sec. The PCR run ended with a post-extension step at 72 °C for 5 min.
Gel electrophoresis, purification, and DNA sequencing
All PCR products were run on 1% agarose gels to check for PCR performance and correct amplicon size. Products of the appropriate size were gel extracted and purified with the Qiagen Gel Extraction Kit (Qiagen, Hilden, Germany) and subsequently Sanger sequenced using the Microsynth SeqLab Laboratory service (Microsynth, Göttingen, Germany).
23S ribosomal RNA gene restriction enzyme analysis
Additional analyses of the 23 S ribosomal RNA gene was conducted to identify the two point-mutations including A2058G and A2059G that encode for macrolide resistance14,15,16. We note here that this locus is not part of the MLST, but it provides an essential information on macrolide resistance in NHP infecting TP strains. Methods were performed according to the procedures described by Lukehart et al.17 with minor modifications. Amplification was done using a semi-nested PCR. The primers of the first PCR were S-primer 5′-GTA CCG CAA ACC GAC ACA G-3′ and AS-primer 5′-GCG CGA ACA CCT CTT TTT AC-3′ using an annealing temperature of 62 °C and 35 cycles. All other PCR conditions were identical with those for TP0619. The second PCR was performed with the S-primer from the first PCR and AS-primer 5′-AGT CAA ACC GCC CAC CTA C-3′. Amplification was conducted with an annealing temperature of 63 °C and 30 cycles. Again, all other PCR conditions were the same as for TP0619. PCR products were run on 1% agarose gels to check for the expected 628 bp-sized PCR product. Subsequently, each sample was digested overnight with restriction enzyme Mbo II (R0148, BioLabs New England, Beverly, MA, USA) at 37 °C and in a second reaction with Bsa I (R0535, BioLabs New England, Beverly, MA, USA) at 50 °C. Interpretation of results was performed as published previously17. TPA strains Street 14 (known mutation at A2058G, digested with Mbo II) and UW330B (known mutation at A2059G, digested with Bsa I) were included as positive controls.
Data analysis
Sequence data were analyzed, edited, and aligned using Geneious v11.1.4 (Biomatters Ltd., Auckland, New Zealand). We compared sequences with respective orthologs available in GenBank using BLAST search (http://blast.ncbi.nlm.nih.gov/Blast.cgi) with megablast or blastn tools. The Free Phylogenetic Network Software (www.fluxus-eneineering.com) was used to create a median-joining network as described elsewhere18. The respective concatenated sequence alignment (TP0548 and TP0488) was used in combination with the assigned traits ‘NHP species’ and ‘sample location trait’. Maximum-parsimony (MP) trees, with gaps coded as fifth character, were constructed with SeaView 4.5.619. Maximum-likelihood (ML) trees were calculated in IQ-TREE 1.6.120 with the respective best-fit models. To obtain node support, all trees were constructed with 1,000 bootstrap replicates.
Results
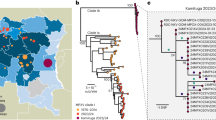
PCR results obtained from 85 NHP samples, of which 79 originated from Tanzanian monkeys (olive baboon, yellow baboon, vervet monkey, and blue monkey), two from Ethiopian grivet monkeys (Chlorocebus aethiops), and four from western lowland gorillas (Gorilla gorilla gorilla) from the Republic of the Congo, are summarized in Table S3. In general, PCR performances of the two gene targets used for the strain typing were equivalent with 78 (TP0548) and 67 (TP0488) resulting sequence data. This created a total of 59 concatenated sequences (the two gene targets combined) that were used for further analysis. We excluded the grivet monkey and the gorilla sample from the network analysis (Fig. 1) in order to maintain the highest possible resolution for the Tanzanian NHP samples where a high number of samples came from one geographic location. However, we used the complete concatenated sequence alignment, including the data obtained from the grivet monkey and gorilla samples, to construct ML (Fig. 2) and MP (Fig. S6) trees. For the latter, gaps were coded as fifth character. NHP strains included in this study clustered with human yaws-causing strains and were clearly separated from the TEN strain Bosnia A as well as the human syphilis-causing TPA strains (Figs 2 and S7). Within the TPE clade, bootstrap support was mostly weak (<80%) with some exceptions. One of these notable exceptions was the separation of the grivet monkey infecting strain from Ethiopia and the strain that was generated from a gorilla fecal sample. Sequences of both strains (14AWM2051017 and 3DZAKM13280917) differed in only one nucleotide position and always clustered together (Figs 2 and S6). Overall, we found a geographic clustering of TPE strains, instead of clustering by host species. None of the samples from the Tanzanian NHPs (n = 72/76; four samples did not generate a PCR product) has been tested positive for the mutations in the 23 S ribosomal RNA genes that code for macrolide resistance. The grivet monkey and the gorilla samples were not tested for microbial resistance.

Median-joining network using 1,773 bp – long concatemer of TP0488 and TP0548 loci from 57 Tanzanian NHPs samples. The number of mutations, when >1, is given close to branches. Inferred allelic variants (median vectors) are shown as small black connecting circles. If contiguous, indels were considered as a single event only. The number of individuals, when >1, is shown inside the circles and are dependent on circle size. Species trait is given in the geometric form: circle = Papio anubis (n = 46); squares = Papio cynocephalus (n = 5); triangles = Chlorocebus pygerythrus (n = 5); hexagon = Cercopithecus mitis (n = 1). Sample location trait is given by the color code: blue = UG (n = 3); orange = TN (n = 1); brown = Ruaha National Park (n = 4); red = Lake Manyara National Park (n = 34); grey = Serengeti National Park (n = 7); green = Ngorongoro Conservation Area (n = 5); violet = Gombe National Park (n = 3).

Rooted ML tree based on the concatenated sequences used for MLST (TP0548 and TP0488). The tree is based on 1,773 nts and 1,000 bootstrap replicates. Bootstrap values from 80–100% are highlighted as light blue circles of respective size. NHP species and/or GenBank accession numbers of published strains are provided in parentheses following the name of the strain. In all cases were the species is not mentioned, sequences are from TP of human origin. Blue = subsp. pallidum, green = subsp. pertenue, red = subsp. endemicum. The pathogen causing rabbit syphilis, Treponema paraluisleporidarum ecovar Cuniculus strain Cuniculi A, is used as an outgroup. The bar refers to substitutions per site.
Sequences obtained from the TP0619 gene were identical across all NHP species and sampling sites, including the grivet monkey and the gorilla samples. A representative sequence obtained from a vervet monkey sampled at Katavi National Park (4KNF2121016) was published previously under the GenBank accession number MF7541222.
In general, all NHP-derived sequences of the TP0548 locus including the grivet monkey and the gorilla sequence, contained one section of the sequence where most of the nucleotide variation is found (Fig. S1). This distinguishes them not only from human syphilis- (TPA) and bejel-causing (TEN) strains but also from TPE strains of human origin, where there are three and two variable regions, respectively. Sequences of the TP0548 and TP0488 loci showed comprehensive variability within and across the different sampling locations as well as between the different NHP species in Tanzania. Corresponding ML and MP trees each constructed for TP0548 (Figs S2 and S3) and TP0488 (Figs S4 and S5) were similar in topology. We note here that sample size was low for some of the species (e.g., blue monkey) and that we included here only the Tanzanian samples to match the network analysis (Fig. 1).
Since the largest number of samples was taken at Lake Manyara National Park (LMNP) and samples were collected in 2007, 2015, and 2017, respectively, we were able to plot the temporal strain composition for this specific sampling location. Figure 3 illustrates the temporal strain composition for all samples that originate from olive baboons. Using a sequence alignment and base-by-base comparison, we found that in 2007 there were six strains, with a dominating strain “E” (e.g., 4LMF8160815). Samples from olive baboons at LMNP taken eight years later reconfirmed the existence of this strain. The temporal stability of strains was further supported by strain “P” (e.g., 10NCM8170516) that was collected from two olive baboons at the Ngorongoro Conservation Area in 2015 and one olive baboon in 2016 (Table S4).

Temporal strain composition for samples collected from olive baboons at Lake Manyara National Park. The different colors indicate different genotypes. Sampling of olive baboons at LMNP was conducted in 2007 and 2015. The strain “E” (light green; e.g., 4LMF8160815) was the dominating strain in 2007 and was still present in baboons sampled eight years later (2015).
Discussion
Frequent transmission of TPE strains across the different NHP species would likely result in a higher number of shared and identical sequences in different species at the same geographic site. Although we found no identical strain sequences in different primate species at one site (Table S4), we still observed a geographic clustering of (closely related) TPE strain sequences and not a clustering according to NHP host species (Fig. 1). This suggests that interspecies transmission occurs, albeit rarely. That we found no identical strain sequences in different primate species could be due to the small sample size for some of the investigated NHP species, but generally argues against frequent transmission. This gains further support by our finding that the strains of NHP origin are relatively stable over time as we show for LMNP (Fig. 3, Table S4). This is consistent with what we know from human infecting TPA strains21,22,23. The 2007 dominating strain “E” (e.g., 4LMF8160815), which was isolated from olive baboons was still present in the infected baboon population in 2015. Although we do not see a strict geographic pattern when the two target genes were analyzed individually (Figs S2–S5), when examining the concatenated alignment (median-joining network (Fig. 1) and the ML tree (Fig. 2)), geographic strain clustering can be observed. This is another indicator for the relative temporal and geographic stability of the strains that infect NHPs. The recently estimated low mutation rate in human TPE strains22 coincides with the genetic stability observed among NHP TPE strains analyzed in this study. Yet, feasible interspecies transmission routes for TP exist3 and have been discussed for flies24 and were proven for sexual intercourse between different NHP species based on host genetic data25.
Despite the differences in the number of variable sites at the TP0548 locus (Fig. S1), we see a close association of NHP and human infecting TPE strains. This was expected, since the genome of TPE LMNP-1 strain (GenBank accession number CP021113), which was obtained from an olive baboon at LMNP in 2007, was found to be closely related to human yaws causing strains similar to all other TPE strains of NHP origin1.
The absence of antibiotic resistance to azithromycin in all tested TP strains from NHPs in Tanzania is a positive sign and is probably related to the absence of treatment of infected monkeys. Currently only Gombe National Park has a history of treating infected baboons with antimicrobials26. In human yaws, it has been shown that after a single treatment round with antibiotic macrolides, resistance emerges27 even though the de novo emergence of such mutations is lower than 10−3 per treated patient28. The risk of emerging antimicrobial resistance is of major concern for human infection and would also draw major implications for the conservation of endangered NHP species such as gorillas (Gorilla gorilla). Similar to human yaws elimination27, responsible treatment of infected NHP populations requires resistance monitoring and possible ring-fencing with effective alternative antimicrobials.
The identification of two different strains obtained from independent PCRs in three different NHPs (TP0548: 11LMF8190815 (P. anubis; 15-bp indel); TP0488: 2UGF8101016 (P. cynocephalus; one SNV) and 24SNM5151115 (P. anubis; three SNVs); Table S4) supports the concept of absence of cross immunity between different TP strains29 and boosts evidence for recombination events found in the TP bacterium30,31,32 under natural conditions.
The initial CDC typing system for TPA made use of the number of 60-bp repeats found in the acidic repeat protein (arp (TP0433)) gene in combination with differences found in the tpr subfamily II genes (tprE (TP0313), tprG (TP0317), and tprJ (TP0621)9. A subsequently introduced enhanced typing system included a portion of the TP0548 locus33. While the enhanced typing system has also been used for the typing of human TPE strains12, it did not overcome the difficulties associated with amplification of 60-bp repeats of the arp gene or the uncertainties associated with amplification of three different tpr-subfamily II genes in one single assay, followed by subsequent restriction enzyme analysis11. A recently published alternative method for MLST included the widely used TP0548 locus but also two additional loci located in the TP0136 and TP0326 genes11. We took these loci into account, but in our analysis, it became evident that the TP0136 locus in the Tanzanian TP strains of NHP origin was highly conserved. A representative sequence is accessible under GenBank accession number CP021113.1 (nt158,275-159,195). Whether this is a characteristic of NHP infecting strains or a spatial property of strains originating from Tanzania is unknown. It underlines, however, that a globally applied strain typing system for TPE requires a comprehensive database of high-quality genomes obtained from larger numbers of clinical samples from yaws endemic areas.
We identified several gene loci as suitable candidates to be used in TPE strains of human and NHP origin. The two gene targets that we selected for MLST (TP0548 and TP0488) originated from a number of suitable candidates (Table 2) and selection was based on best PCR performance in the clinical NHP samples that were included in this study. We neither aimed for the design of a TPE typing system that can be used in a clinical environment for human infection, nor did we anticipate a typing system that is suitable for a global approach. Amplification of relatively long sequence parts (e.g., TP0548 enhanced typing system determines 84 bp33 vs. 1,065 bp determined in this study) was therefore not considered an issue. Both loci that were used for the typing, TP0548 and TP0488, are reported to show signs of recombination in human syphilis causing TPA strains30,31,32. This, however, was not considered a limitation since our MLST typing system was designed to describe strain variability within a given population of NHPs and is not used to describe a detailed geophylogeny of TP. While the TP0619 sequence, which codes for a protein family of Domains of Unknown Function (DUF)2715 and which appear to be restricted to TP, is not part of our typing system, it is useful to support the difference of the TP strains of NHP origin analyzed in this study from human syphilis-causing TPA strains. It is currently unclear if wild NHPs are also naturally infected with TPA strains.
Invasive sampling of NHPs in the wild is generally associated with challenges in terms of ethics and logistics. As a consequence, it is easier to obtain a greater number of non-invasively collected fecal samples than samples that originate from invasively sampled individuals. Moreover, prospective epidemiological studies would benefit from sampling regimes that allow the screening and subsequent typing of TP strains in non-invasively collected samples from a greater geographic area. For this reason, we tested our newly established MLST system in noninvasively collected fecal samples from gorillas at Odzala-Kokoua National Park in the Republic of the Congo, a place where gorillas with ulcerative skin lesions have been frequently sighted34 (Fig. S7). We were able to successfully strain type the gorilla samples and could show that the strain from which we obtained all three target sequences, clusters with a strain isolated from a grivet monkey in Ethiopia. The reason for the close association of the strain of the grivet monkey and the gorilla origin is currently unclear and answering this question requires intensified sampling in the respective geographic areas and in-between to obtain more sequence data.
Conclusion
The high number of TP infected NHPs in Africa, the different species that have been confirmed as hosts2,4, as well as the recently documented close genetic and functional similarity of NHP and human infecting TPE1 requires epidemiological data for a better understanding on how the infection is maintained in primate populations and whether or not it is transmitted to humans. Our study provides an important contribution to answer the question on interspecies transmission in primate infecting TP, although further sampling is needed to increase confidence in the results. However, with our data we were able to show that interspecies transmission in Tanzanian monkeys is likely although rare. As humans are primates, the most important question to answer in future studies is, whether or not TPE strains of NHP origin transmit to humans and/or vice versa. The interspecies transmission of TPE in nonhuman primates, is not necessarily predictive for spillovers to humans.
Data Availability
GenBank accession numbers for the sequences generated in this study can be found in Table S5.
References
Knauf, S. et al. Nonhuman primates across sub-Saharan Africa are infected with the yaws bacterium Treponema pallidum subsp. pertenue. Emerg Microbes Infect 7, 157 (2018).
Chuma, I. S. et al. Widespread Treponema pallidum infection in nonhuman primates, Tanzania. Emerg Infect Dis 24, 1002–1009 (2018).
Hallmaier-Wacker, L. K., Munster, V. J. & Knauf, S. Disease reservoirs: from conceptual frameworks to applicable criteria. Emerg Microbes Infect 6, e79 (2017).
Knauf, S., Liu, H. & Harper, K. N. Treponemal infection in nonhuman primates as possible reservoir for human yaws. Emerg Infect Dis 19, 2058–60 (2013).
Peng, R. et al. Molecular typing of Treponema pallidum: a systematic review and meta-analysis. PLoS Negl Trop Dis 5, e1273 (2011).
Salado-Rasmussen, K. et al. Molecular typing of Treponema pallidum in Denmark: A nationwide study of syphilis. Acta Derm Venereol 96, 202–6 (2016).
Grange, P. et al. Molecular subtyping of Treponema pallidum in Paris, France. Sex Transm Dis 40, 641–4 (2013).
Pope, V. et al. Molecular subtyping of Treponema pallidum from North and South Carolina. J Clin Microbiol 43, 3743–3746 (2005).
Pillay, A. et al. Molecular subtyping of Treponema pallidum subspecies pallidum. Sex Transm Dis 25, 408–414 (1998).
Grillová, L. et al. Molecular typing of Treponema pallidum in the Czech Republic during 2011 to 2013: increased prevalence of identified genotypes and of isolates with macrolide resistance. J Clin Microbiol 52, 3693–3700 (2014).
Godornes, C., Giacani, L., Barry, A., Mitjà, O. & Lukehart, S. Development of a Multilocus sequence typing (MLST) scheme for Treponema pallidum subsp. pertenue: Application to yaws in Lihir Island, Papua New Guinea. PLoS Neglect Trop Dis 11, e0006113 (2017).
Katz, S. S. et al. Molecular strain typing of the yaws pathogen, Treponema pallidum subspecies pertenue. PLoS ONE 13, e0203632 (2018).
Knauf, S. et al. Treponema infection associated with genital ulceration in wild baboons. Vet Pathol 49, 292–303 (2011).
Matějková, P. et al. Macrolide treatment failure in a case of secondary syphilis: a novel A2059G mutation in the 23S rRNA gene of Treponema pallidum subsp. pallidum. J Med Microbiol 58, 832–836 (2009).
Chen, C. et al. Detection of the A2058G and A2059G 23S rRNA gene point mutations associated with azithromycin resistance in Treponema pallidum by use of a TaqMan real-time multiplex PCR assay. J Clin Microbiol 51, 908–913 (2013).
Stamm, L. & Bergen, H. A point mutation associated with bacterial macrolide resistance is present in both 23S rRNA genes of an erythromycin-resistant Treponema pallidum clinical isolate. Antimicrob Agents and Chemother 44, 806–807 (2000).
Lukehart, S. et al. Macrolide resistance in Treponema pallidum in the United States and Ireland. N Engl J Med 351, 154–158 (2014).
Bandelt, H., Forster, P. & Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol Biol and Evol 16, 37–48 (1999).
Gouy, M., Guindon, S. & Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol Biol Evol 27, 221–224 (2010).
Nguyen, L.-T. T., Schmidt, H. A., Haeseler, A. & von Minh, B. Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol and Evol 32, 268–274 (2015).
Strouhal, M. et al. Complete genome sequences of two strains of Treponema pallidum subsp. pertenue from Ghana, Africa: Identical genome sequences in samples isolated more than 7 years apart. PLoS Negl Trop Dis 11, e0005894 (2017).
Grillová, L. et al. Molecular characterization of Treponema pallidum subsp. pallidum in Switzerland and France with a new multilocus sequence typing scheme. PloS ONE 13, e0200773 (2018).
Grillová, L. et al. Sequencing of Treponema pallidum subsp. pallidum from isolate UZ1974 using anti-treponemal antibodies enrichment: First complete whole genome sequence obtained directly from human clinical material. PLoS ONE 13, e0202619 (2018).
Knauf, S. et al. Isolation of Treponema DNA from necrophagous flies in a natural ecosystem. EBioMed 11, 85–90 (2016).
Zinner, D., Chuma, I. S., Knauf, S. & Roos, C. Inverted intergeneric introgression between critically endangered kipunjis and yellow baboons in two disjunct populations. Biol Lett, 14 (2018).
Collins, D., Sindimwo, A. & Wallis, P. Reproductive disease of olive baboons (Papio anubis) of Gombe National Park: outbreak, time-course, and attempts to limit recurrence. In: Climate change: challenges and opportunities towards sustainable wildlife conservation and development. 236–248 (Tanzania Wildlife Research Institute, 2011).
Mitjà, O. et al. Re-emergence of yaws after single mass azithromycin treatment followed by targeted treatment: a longitudinal study. Lancet 391, 1599–1607 (2018).
Šmajs, D. & Pospíšilová, P. Macrolide resistance in yaws. Lancet 391, 1555–1556 (2018).
Turner, T. & Hollander, D. Biology of the treponematoses based on studies carried out at the International Treponematosis Laboratory Center of the Johns Hopkins University under the auspices of the World Health Organization. Monogr Ser World Health Organ 35, 3–266 (1957).
Pětrošová, H. et al. Whole genome sequence of Treponema pallidum ssp. pallidum, strain Mexico A, suggests recombination between yaws and syphilis strains. PLoS Negl Trop Dis 6, e1832 (2012).
Grange, P. et al. Treponema pallidum 11qj subtype may correspond to a Treponema pallidum subsp. endemicum strain. Sex Transm Dis 43, 517–518 (2016).
Mikalová, L. et al. Human Treponema pallidum 11q/j isolate belongs to subsp. endemicum but contains two loci with a sequence in TP0548 and TP0488 similar to subsp. pertenue and subsp. pallidum, respectively. PLoS Negl Trop Dis 11, e0005434 (2017).
Marra, C. et al. Enhanced molecular typing of Treponema pallidum: geographical distribution of strain types and association with neurosyphilis. J Infect Dis 202, 1380–1388 (2010).
Levrero, F., Gatti, S., Gauthier-Hion, A. & Menard, N. Yaws disease in a wild gorilla population and its impact on the reproductive status of males. Am J Phys Anthropol 132, 568–575 (2007).
Brinkman, M. et al. A novel Treponema pallidum antigen, TP0136, is an outer membrane protein that binds human fibronectin. Infect Immun 76, 1848–1857 (2008).
Radolf, J. D. & Kumar, S. The Treponema pallidum outer membrane. Curr Top Microbiol Immunol 415, 1–38 (2018).
Acknowledgements
The study was funded by the German Research Foundation (DFG): KN1097/3-1 and KN1097/4-1 [SK], and RO3055/2-1 [CR]. In addition, it was partly supported by the Grant Agency of the Czech Republic (GA17-25455S; GC18-23521J) to DS. We thank the Government of the United Republic of Tanzania (URT) for permission and logistical support that was needed. Institutions include the Ministry for Education and Vocational Training (MoEVT), the Commission for Science and Technology (COSTECH), Ministry for Natural Resources and Tourism (MNRT), Ministry for of Agriculture, Natural Resources, Livestock and Fisheries (SMZ), Department of Forestry and Non-renewable Natural Resources (SMZ), Tanzania Wildlife Authority (TAWA), Tanzania Wildlife Research Institute (TAWIRI), Tanzania National Parks (TANAPA), Ngorongoro Conservation Area Authority (NCAA), Sokoine University of Agriculture (SUA), Department of Livestock Development (SMZ). We are also grateful for the support received from the Jane Goodall Institute (JGI) and the National Institute for Medical Research (NIMR). Most importantly, our gratitude goes to all people and staff-members working with government and partner institutions in Germany and Tanzania who tirelessly strived to facilitate smooth undertaking of the project at all levels from planning, execution of fieldwork as well as laboratory analyses. Sheila A. Lukehart and Charmie Godornes (University of Washington) are thanked for technical support and scientific advice.
Author information
Authors and Affiliations
Contributions
The study was designed by I.S.C., C.R., D.S. and S.K. Design of the MLST system was done by C.R., L.G., J.O., D.S. and S.K. Laboratory work took place at Sokoine University of Agriculture and the German Primate Center and was performed by I.S.C., C.R., L.H.W., S.L. and S.K. Data were analyzed by I.S.C., C.R., D.S. and S.K. All authors (I.S.C., C.R., A.A., T.B., D.A.C., L.G., L.H.W., R.R.K., J.D.K., S.L., U.M., J.O., K.J.P., A.P., F.A.S., D.S. and S.K.) contributed to the manuscript preparation.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chuma, I.S., Roos, C., Atickem, A. et al. Strain diversity of Treponema pallidum subsp. pertenue suggests rare interspecies transmission in African nonhuman primates. Sci Rep 9, 14243 (2019). https://doi.org/10.1038/s41598-019-50779-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-50779-9
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
