
Abstract
Traumatic brain injury (TBI) is one of the most common injuries to military personnel, a population often exposed to stressful stimuli and emotional trauma. Changes in sensory processing after TBI might contribute to TBI-post traumatic stress disorder (PTSD) comorbidity. Combining an animal model of TBI with an animal model of emotional trauma, we reveal an interaction between auditory sensitivity after TBI and fear conditioning where 75 dB white noise alone evokes a phonophobia-like phenotype and when paired with footshocks, fear is robustly enhanced. TBI reduced neuronal activity in the hippocampus but increased activity in the ipsilateral lateral amygdala (LA) when exposed to white noise. The white noise effect in LA was driven by increased activity in neurons projecting from ipsilateral auditory thalamus (medial geniculate nucleus). These data suggest that altered sensory processing within subcortical sensory-emotional circuitry after TBI results in neutral stimuli adopting aversive properties with a corresponding impact on facilitating trauma memories and may contribute to TBI-PTSD comorbidity.
Similar content being viewed by others
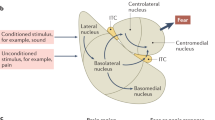
Introduction
Traumatic brain injury (TBI) is one of the most common injuries to military personnel, a population often exposed to stressful stimuli and emotional trauma. TBI typically impairs learning and memory for neutral events1, but may enhance traumatic fear memories2. Furthermore, long-term consequences of TBI include increased risk for neurological and psychiatric disorders3,4,5,6,7. Increasing numbers of military service members that have been exposed to both emotional and physical trauma have promoted interest in understanding the comorbidity between TBI and post-traumatic stress disorder (PTSD), as mild TBI is a significant predictor of PTSD following deployment8. It remains a clinical challenge to know which precipitating event causes PTSD in TBI, the brain injury itself or the psychological stressor(s) surrounding the TBI. Understanding how and whether traumatic memories are encoded differently after TBI will aid in our efforts in recognizing underlying mechanisms and developing effective treatments for this highly prevalent and complex comorbidity. Here we combined widely used preclinical models to identify potential causal links between TBI and PTSD.
Pavlovian fear conditioning is widely used to study fear learning and memory, where an innocuous stimulus (conditional stimulus, CS), such as an auditory cue is paired with an aversive stimulus (unconditional stimulus, US), such as a footshock, which promotes natural defensive responses. Following association between CS and US, both the context and CS alone will elicit a learned conditional response (CR), freezing, indicative of fear. The basolateral amygdala complex is a key locus of plasticity for the formation of fear memories9. The lateral subdivision (lateral amygdala, LA), which receives direct cortical and thalamic auditory input, is known as the auditory interface of the amygdala and is required for auditory fear conditioning10. While the amygdala has been relatively understudied in TBI research, recent data from our lab and others have begun to find TBI-induced changes in amygdala structure and function towards enhanced excitatory processes and increased plasticity11,12,13,14. Specifically, we reported that 48 h after fluid percussion injury (FPI), an experimental model of diffuse concussive-like TBI, rats displayed enhanced contextual fear learning when footshocks were signaled with a white noise CS, but not when unsignaled11. These enhanced fear memories were associated with increased N-methyl-D-aspartate (NMDA) receptor expression in the BLA11. The hippocampus also plays a key role in the formation of contextual fear memories15,16,17, has significant implications in PTSD18,19,20, and is highly susceptible to dysfunction following TBI21,22,23. Thus the neural circuits necessary for encoding adaptive auditory fear memories are vulnerable to disruption after TBI and may lead to increases in fear learning and expression24.
Sensory sensitivity or sensory processing issues including photophobia (light sensitivity), phonophobia (sound sensitivity), hyperacusis (sensitivity to certain sound frequencies), and allodynia (tactile sensitivity) are among the primary physical symptoms of diffuse clinical TBI or concussion25,26,27, and are often categorized and treated separately from neuropsychiatric sequelae28. Experimental evidence suggests that somatosensory input is enhanced after injury29,30. Interactions between aberrant sensory processing and amygdala vulnerability in the TBI brain may impact stress reactions and underlie potential mechanisms of comorbid TBI/PTSD. By separating the physical from emotional trauma, our design provides the opportunity to determine whether effects such as sensory sensitivity from the TBI itself impact function in networks required for fear memory encoding. We hypothesized that FPI enhances contextual fear to auditory fear conditioning as a result of injury-induced altered auditory processing. In this study, we examined the stimulus-specific enhancement of contextual fear learning after FPI and changes in projection-specific functional plasticity in auditory fear circuitry and the hippocampus.
Results
After FPI, white noise, but not pure tone, evokes robust defensive behavior and enhances contextual fear when used in fear conditioning
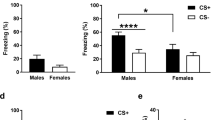
We sought to determine the effects of FPI on auditory and contextual fear memory with white noise used as a CS paired with shock. Outlined in Fig. 1A, two days following either FPI or sham surgery, rats were pre-exposed to white noise CS cues (30 sec/75 dB) and the following day received 10 white noise-shock pairings. Two cohorts of animals were used and data were combined in this experiment for replicability (sham, n = 16; FPI, n = 19). When white noise was presented alone, FPI groups showed increased freezing during cue presentations compared to shams (F(1, 33) = 21.055, p < 0.001; Fig. 1B), and this response was most robust during the post-noise intervals between trials (F(1, 33) = 24.417, p < 0.001; Fig. 1C). The next day, both groups were fear conditioned with 10 white noise-footshock pairings in the same context. Although at low levels (sham, 1.4 ± 2.1%; FPI 6.9 ± 10.4%), FPI groups displayed significantly more freezing during the baseline period prior to the first CS-US trial compared to sham (t(29) = 2.081, p = 0.046; BL: Fig. 1D)., indicating pre-existing unconditional responding from white noise influenced context freezing prior to paired presentations with footshocks. Successful acquisition was indicated by increased freezing over conditioning trials (F(9, 216) = 22.265, p < 0.001; Fig. 1D), however group differences during conditioning did not reach statistical significance (F(1, 29) = 2.513, p = 0.124), Fig. 2D. Over the next three days, both groups were tested for contextual fear extinction in the absence of discrete stimuli, both groups decreased freezing to context across days of testing (F(2, 66) = 113.535, p < 0.001). However, FPI groups exhibited robustly enhanced fear to the training context compared to shams (F(1, 33) = 12.637, p = 0.001), Fig. 1E, consistent with our previous finding11. When tested for white noise CS fear in a new context after levels of fear were decreased by context extinction, there was no effect of injury (F(1, 33) = 0.290, p = 0.594), however there was a significant decrease in freezing across the cue trials (F(6, 198) = 12.298, p < 0.001, Fig. 1F).

White noise promotes defensive behavior after fluid percussion injury and contributes to enhanced contextual fear learning when paired with footshocks. (A) Experimental design. (B) During noise pre-exposure with presentations of white noise cues in the absence of footshocks, FPI (fluid percussion injury) groups froze significantly more than shams and (C) to an even greater degree at the offset and during the intervals between trials. (D) When white noise was paired with footshocks, both groups increased freezing across trials indicating learning, however the FPI group was not different than sham. (E) Freezing in the conditioning context was robustly increased in the FPI groups across three days of testing. Both groups decreased freezing across days, indicating contextual fear extinction. (F) When tested in a novel context there were no group differences in freezing to the white noise cue. ***p < 0.001 vs. Sham; data are represented as mean ± SEM.

Lateral fluid percussion injury reduces tone fear memory. (A) Experimental design. (B,C) FPI (fluid percussion injury) rats displayed slightly elevated levels of freezing behavior during pre-exposure to pure tone (2800 Hz/75 dB) trials (B) and during inter stimuli intervals (C). (D) FPI had no effect on baseline freezing prior to the first tone-shock conditioning trial. Although both groups learned, FPI had no effect on freezing across acquisition trials when pure tones were paired with mild footshocks. (E) While both groups decreased freezing across context extinction sessions, FPI had no effect on fear to the conditioning context. (F) FPI rats froze less during tone CS trials when presented in a novel context. *p < 0.05 vs. Sham. Data are represented as mean ± SEM.
Phonophobia and hyperacusis are common and often persistent physical symptoms after TBI or concussion28,31,32. Therefore, we hypothesized that injury-induced altered sensory processing led to the phonophobia-like phenotype and enhanced contextual fear also found by Reger et al.11. Because white noise is comprised of a mixture of all frequencies it may differentially elicit phonophobic and/or hyperacoustic reactions. To test for stimulus specificity of the injury effect, we tested a new cohort of FPI and sham rats under the same protocol using low frequency pure tones at the same intensity (30 sec/2800 Hz/75 dB). During pre-exposure, FPI rats froze some during tone trials (F(1, 14) = 8.693, p = 0.011; Fig. 2B), and between trials (F(1, 14) = 7.767, p = 0.015; Fig. 2C) compared to sham, with an overall significant increase across trials (F(6, 84) = 2.242, p = 0.047). However, the magnitude of the effect was far greater to white noise, and in particular following the offset of the stimulus during the post-noise interval. When analyzed together by a two way mixed factors ANOVA, we saw a significant injury x stimulus interaction for pre-exposure inter-trial-intervals (ITI; F(1, 47) = 4.666, p = 0.036); FPI-tone vs. FPI-white noise post hoc comparison (F(1, 23) = 4.272, p = 0.05), Supplemental Fig. S2), indicating a greater photophobic-like response to white noise above tones. When tested the next day, baseline freezing did not differ between injury conditions after pre-exposure to pure tones as it did in the first experiment following white noise (t(14) = 1.183, p = 0.256; BL: Fig. 2D). During conditioning, both groups increased freezing across acquisition trials throughout the session (F9, 126) = 50.22, p < 0.001), however both groups reached ceiling after the first trial, Fig. 2D. Over the next three days, both groups were returned to the same context to measure contextual fear and extinction to the conditioning context. While both groups decreased freezing across context extinction days (F(2, 42) = 26.9, p < 0.001), there was no effect of injury on freezing to the context (Fig. 2E). When both groups were placed in a novel context and presented with the trained tone CS, FPI rats displayed significantly less freezing during the tone test (F(1, 98) = 5.453, p = 0.0216; Fig. 2F), indicating a deficit in recall of the trained cue, which is in contrast to our previous finding11. These data provide novel evidence that after diffuse TBI, white noise alone evokes freezing defensive behavior. This in turn leads to enhanced contextual fear when used as a CS, consistent with our initial findings with white noise11, while tone fear memory may be impaired.
White noise exposure causes increased amygdala, but reduced hippocampal activity following FPI
Our behavioral data suggest that FPI makes white noise in and of itself aversive, or “US-like.” We then asked how the injured brain responds differently to white noise using activity dependent Arc immunolabeling within important targets including the lateral amygdala (LA) and dorsal dentate gyrus (DG) of the hippocampus. The LA receives direct sensory information from auditory cortical and thalamic areas, which is required for the formation of auditory fear10. The dorsal DG is highly vulnerable to insult after TBI33 and required for contextual fear memory formation34. Two days following FPI or sham surgery, animals were placed in a novel context and exposed to either 7 white noise alone trials, similar to the pre-exposure sessions in earlier experiments (75 dB), or were quiet controls and brains were processed for Arc immunohistochemistry IHC (n = 5–6/group). Arc is rapidly expressed in projection neurons as a result of a single behavioral experience35, and is necessary for associative fear memories in the amygdala36,37. Interestingly, two way between subjects multivariate ANOVA revealed a significant injury effect (Wilks’ Lambda (F(4, 16) = 21.14, p < 0.001), and an injury x noise condition multivariate interaction (Wilks’ Lambda (F(4, 16) = 5.242, p = 0.007) for Arc positive cells across ipsi- and contralateral lateral amygdala (LA) and dorsal dentate gyrus (DG). Between subjects MANOVA effects showed a significant injury x noise condition effect within the ipsilateral LA (F(1, 19) = 13.465, p = 0.002), where posthoc analyses showed that only noise-exposed FPI rats had robust increased Arc expression relative to all other groups, (p < 0.05; Fig. 3A). Between subjects MANOVA revealed that in the DG of the hippocampus there was also a significant injury x noise condition interaction. Post hoc analyses for sham controls showed that regardless of hemisphere, there was a significant effect of noise condition (Wilks’ Lambda, F(4, 7) = 13.052, p = 0.002), where noise-exposed shams had more Arc induction than quiet controls, Fig. 3C. This was not the case in FPI animals, where regardless of noise condition, there was significantly less Arc induction in the ipsilateral hemisphere (F(1, 19) = 73.961, p < 0.001), which is in direct contrast to effects observed in the LA, Fig. 3C. This pattern of findings suggests dysregulated activity within the limbic regions that are important for processing contextual fear in the presence of white noise stimuli that in the absence of FPI are not aversive or noxious.

Arc protein induction in response to white noise after FPI. (A) Lateral amygdala, (LA) White noise exposure in FPI (fluid percussion injury) rats caused robust Arc induction within the ipsilateral LA relative to contralateral and all other groups, ***p < 0.001 vs. contra and all other groups. (C) Dorsal dentate gyrus, (DG) FPI caused a significant overall reduction in Arc protein in response to novel context exploration alone or in the presence of white noise exposure in the ipsilateral DG (*p < 0.05). White noise exposure led to greater Arc induction within the dorsal DG in uninjured sham controls (##p < 0.01), but not FPI groups. Data are represented as mean ± SEM. (B,D) Photomicrographs are representative images of Arc immunohistochemistry (IHC) within the ipsilateral LA (B) and DG (D) in groups exposed to white noise.
Increased activity in ipsilateral thalamo-amygdala projecting neurons during white noise exposure after lateral FPI
The LA receives direct sensory input from cortical and thalamic regions that drive plasticity and are known to be required for auditory fear learning38. In order to determine which amygdala inputs may be disrupted following FPI that drive the increased activity in the LA, we measured Arc expression in MGN-LA and Te3-LA projecting neurons during white noise exposure 48 h after lateral FPI in rats that had previously received bilateral LA retrograde tracer infusions with CTB (Fig. 4A). Replicating earlier experiments, FPI rats froze significantly greater in the context than shams between white noise trials (75 dB; t(17) = 3.711, p = 0.0017). Data included in CTB + Arc analyses were only from subjects with accurate LA CTB placement verified by expression in infusion site and by ipsilateral LA afferent retrograde CTB expression (sham ipsi, n = 9; sham contra, n = 8; FPI ipsi, n = 5; FPI contra, n = 8). MANOVA revealed a significant effect of injury for LA projecting cells in the MGN that were positive for Arc (F(2, 7) = 9.275, p = 0.011; Fig. 4B), and between group effects supported increased MGN-LA Arc activity in both ipsi (F(1, 8) = 9.509, p = 0.015) and contra (F(1, 8) = 9.175, p = 0.016) indicating that there was a significant increase in activity within MGN-LA cells in FPI compared to sham; Fig. 4B). There was no significant multivariate effect for Arc for the secondary auditory cortex (Te3)-LA (Fig. 4C). This effect was corroborated with a different IEG also involved in synaptic plasticity, c-Fos (see Supplemental Fig. S4), indicating that white noise after FPI specifically increased MGN-LA activity that may drive increased amygdala plasticity and underlie corresponding sensitivity to white noise and elicited defensive behavior.

Increased activity in ipsilateral thalamo-amygdala projecting neurons during white noise exposure after lateral FPI. (A) Experimental design. (B) Increased Arc activity in MGN-LA (medial geniculate nucleus-lateral amygdala) projection neurons (*p < 0.05), but not Te3-LA (secondary auditory cortex-lateral amygdala; C) during white noise exposure after FPI. *p < 0.05; Data are represented as mean ± SEM. (B) Representative Arc activity (red) in retrolabeled CTB (cholera toxin subunit B) in LA afferents (green) in ipsilateral MGN. Scale bar 200 µm.
Discussion
Combining well established models of brain trauma and emotional trauma, this study investigated the novel role of auditory sensitivity on enhanced learned fear following diffuse TBI. Our data showed that lateral FPI robustly increased contextual fear when white noise was used as a cue paired with footshocks (US), but not with pure tone cues. We also found that in the absence of footshocks, 75 dB white noise elicited phonophobia-like defensive behavior (freezing) following FPI. Furthermore, we found increased induction of Arc protein within the ipsilateral lateral amygdala (LA) only in white noise-exposed FPI groups relative to sham and quiet FPI controls, indicating that the white noise alone initiates greater amygdala activity after brain injury. We then determined that increased activity in thalamo-amygdala projection neurons were more active during white noise on the injured hemisphere. We also observed a reduction in Arc activity within the dorsal dentate gyrus (DG) of the hippocampus in injured animals during white noise exposure, indicating dysregulated network function within brain regions required for the formation of contextual and auditory fear memories. Collectively, our data indicate that lateral FPI led to sensitivity to white noise that significantly facilitated the formation of contextual fear memories and amygdala activity during the early phase following injury.
White noise is aversive after FPI
Contextual fear memories are most robust when the US (a footshock) is unsignaled or unpredictable39,40,41. When the shock is preceded by an auditory stimulus as a CS, the CS is predictive of the shock and therefore more strongly conditioned than the context in which the experience occurred, although there is still a degree of conditioning to the context itself. The contextual memory is known to require both the hippocampus and amygdala42,43. Our current and previous data support that the context is more strongly conditioned after FPI when white noise is used as a CS11. This white noise-specific effect is also supported by our behavioral data indicating that FPI had no effect on contextual fear during baseline after pure tone pre-exposure or following tone-shock fear conditioning. Furthermore, our previous study showed that FPI did not affect contextual fear memory when footshocks were unsignaled11. At higher decibels, white noise functions as a US44,45,46,47. Emerging evidence suggests differential plasticity and defensive behavior patterns in response to white noise and pure tones in auditory fear conditioning48,49, and may even recruit additional circuitry50. Given that white noise is a mixture of all frequencies it may differentially elicit phonophobic and/or hyperacoustic reactions following FPI. Perhaps at a lower intensity and/or within frequency ranges captured, white noise may be processed as aversive after TBI. We also observed increased Arc induction in the ipsilateral LA in the FPI group that received noise exposure. Importantly, LA neurons receive US-evoked excitatory inputs from both thalamic and cortical areas51,52. Auditory processing changes have also been observed in human concussion and may serve as a potential biomarker by way of evoked frequency-following response26. Among clinical risk factors for persistent post-concussion symptoms in a large pediatric cohort, only noise sensitivity, headache, and fatigue were included in the final predictive model27,see also53. Human brain imaging studies show elevated amygdala activity in response to aversive auditory stimuli54,55. The sensory processing difference in the FPI group may support an additive effect across cumulative exposure to both aversive white noise and footshocks that lead to overall enhanced conditional fear to the context and reflects the phonophobia-like phenotype.
Opposing activity within emotional and cognitive neural substrates after FPI
In response to white noise, we found altered and opposing levels of functional activity within the sensory amygdala and hippocampal subregions between FPI and sham controls. We observed overall reduced Arc activity within the granule cell layer of the dorsal DG after injury. Within the sham groups, white noise resulted in increased dorsal DG Arc induction in white noise-exposed compared to quiet controls. This difference that was absent in the FPI groups perhaps reflects greater elemental learning about the context in the presence of an innocuous auditory cue to the uninjured brain, as it is understood that DG signaling is involved in contextual pattern separation56. Furthermore, Arc is induced in hippocampal and cortical calcium/calmoldulin-dependent protein kinase II (CaMKII) neurons during context exploration57 that promotes functional plasticity in memory consolidation, which is compromised after FPI. Reduced Arc observed in the injured DG is consistent with the literature regarding hippocampal function and plasticity after brain injury58, including reports of impairments in hippocampal LTP59,60, hippocampal-dependent learning and memory61, and reduced synaptic morphology22. Curiously, the FPI group exhibited reduced activity in the dorsal DG but also had enhanced context fear under similar conditions after noise-shock conditioning. While we know that the hippocampus is critical for contextual fear62, it is important to consider that our TBI model does not produce a lesion, but a dysfunctional hippocampal system, which in some cases leads to aberrant enhanced fear responses63. In contrast to the reduced activity in the injured DG, white noise exposure caused increased activity in the ipsilateral sensory amygdala (LA) in the FPI group, which may be driving the enhanced fear learning. Importantly, this effect in the LA was not observed in the quiet FPI group, supporting a stimulus-elicited effect in LA neurons that may reflect the phonophobia-like behavior in this group. Our projection mapping data suggests that this increase is driven by increased activity within ipsilateral auditory thalamus (MGN) neurons projecting to the LA. This effect was specific to the MGN and not secondary auditory cortex (Te3)-LA projections, indicating the specificity of the inputs that drive increased plasticity and corresponding defensive behavior. We also found increased astrocyte reactivity in FPI groups as measured by glial fibrillary acidic protein (GFAP) in bilateral LA, DG, and auditory cortex (Supplemental Fig. S3), and in ipsilateral MGN. Reactive glia as a result of injury may contribute to sensitized plasticity in auditory fear neural circuitry and is a target of future research. Together, our Arc data suggest an injury-induced mismatch between reduced activity in structures vulnerable to TBI that coordinate cognitive processes and context learning amid increased activity in thalamo-amygdala projections, which are known to drive fear and defensive behavior and is implicated in PTSD.
TBI interactions with sensory-emotional network function
Auditory consequences after TBI are common in military populations, and resulting conditions like tinnitus may complicate post traumatic stress and emotional-sensory networks64. Our study is the first to show an interaction between sensory sensitivity and increased amygdala activity underlying defensive behavior after TBI. Consistent with our Arc findings, experimental models of tinnitus and hyperacusis have been linked to lateral amygdala hyperactivity during exposure to auditory stimuli65. Furthermore, sensory sensitivity has also been reported in other modalities using midline FPI, where whisker stimulation causes behavioral morbidity and elevated stress response29. Importantly, our findings may support an underlying mechanistic link with recent clinical studies in human TBI and other clinical populations with sensory symptoms. After TBI, anxiety interactions with sensory disturbances may underlie persistent clinical symptoms. One study assessed 12 different symptoms in physical, cognitive, and emotional categories, and found that noise sensitivity and anxiety were the only two initial TBI symptoms that were significant predictors for persistent post concussive syndrome66. Furthermore, another study showed that patients with chronic TBI showed persistent auditory sensitivity in a subset patients that were comorbid with PTSD67. Additional recent work by Papesh et al. showed generalized dysfunctional sensory gating and failure to habituate to auditory tones following blast exposure68. In other patient populations with sensory sensitivity such as in autism spectrum disorder, greater amygdala activity55 and thalamo-amygdala connectivity corresponds to degree of sensory overresponsitivity severity69. Whether interactions between sensory sensitivity and emotional reactivity and fear have a bidirectional relationship contributing to challenges in recovery after TBI remains to be determined and using animal models can allow for systematic comparison.
Conclusions
We report that auditory sensitivity after diffuse TBI may contribute to enhanced fear learning and amygdala hyperactivity. We observed dysregulated activity between the hippocampus and thalamo-amygdala projections in response to white noise, which corresponded with a phonophobic behavioral phenotype. This led to a stimulus-specific enhancement in fear learning after white noise cued fear conditioning. These data add to the developing body of literature aimed at understanding the complexity of neurobiological interactions between comorbid TBI and PTSD24. Extending to other neural systems, it has also been hypothesized that changes in sensory processing after TBI may also affect cognitive and motor deficits70 and may contribute to increased post traumatic headache and migraine71. Elucidating the mechanisms that are affected in sensory and emotional systems at the circuit, cellular, and molecular levels are critical next steps in understanding how the injured brain perceives and reacts to environmental stimuli differently. In summary, our data provide implications for altered sensory processing after TBI, where otherwise neutral stimuli may adopt aversive properties and impact encoding of traumatic memories.
Methods
Subjects and lateral fluid percussion injury (FPI)
Young adult male Sprague-Dawley rats (Envigo, 250–275 g upon arrival) were pair housed and maintained on a 12 hour light/dark cycle with ad libitum food and water. All rats were handled for 1 min/day 4 days prior to surgery. All procedures were conducted with approval from the University of California Los Angeles Institutional Care and Use Committee and were in compliance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Young adult males were utilized due to epidemiological data supporting that males are at a significantly higher risk for TBI, with the highest male-to-female ratios occurring in young adulthood72. Rats underwent either sham surgery or mild-moderate lateral fluid percussion injury (FPI). Lateral FPI is a general brain movement injury that exposes the entire brain to forces generated by the percussion73,74,75. FPI was induced using a previously published protocol11,76,77,78 typically used in our laboratory. Animals were anesthetized under a 1–2% isoflurane-oxygen mixture. A midline incision was made followed by a left hemisphere 3 mm diameter craniotomy centered 3 mm posterior and 6 mm lateral to bregma. A plastic injury cap was adhered to the skull with silicone gel and dental cement and filled with sterile saline. The animal was removed from anesthesia and the injury cap was attached to the fluid percussion injury device (Virginia Commonwealth University, Richmond, Virginia). Upon toe pinch response, a brief fluid pulse (~20 msec) of saline was administered directly to the dura mater. Apnea and loss of consciousness (LOC; measured by toe pinch response) were measured to determine injury severity (Fig. S1). Rats were then placed back on anesthesia to remove injury cap and suture the scalp. Sham animals received the same surgical procedures except fluid pulse impact. Upon completion of surgery, animals were placed in a heated recovery chamber until normal behavior resumed and returned to the vivarium. Animals were weighed and monitored post operatively for at least a week after surgery or until the end of the experiment. Injury severity as measured by toe pinch withdrawal was within the mild-moderate range, similar across experiments (269.2 sec mean ± 142.33 sec standard deviation) and counterbalanced within experiments comparing noise and no noise exposure after FPI (Supplemental Fig. S1).
Fear conditioning and behavior
All behavioral testing began 48 h after surgery based on our initial findings on amygdala function following FPI11,74. Training and auditory cue testing occurred in two distinct conditioning chambers (context A and context B) that differed in transport, location, odor, lighting, chamber shape, and flooring (Med Associates Inc., Georgia, VT). Percent time freezing to auditory stimuli and context were recorded as measures of auditory cued and contextual fear, respectively. Behavioral testing protocols differed slightly depending on the goals of the experiments, as outlined below.
To examine the stimulus-specificity of FPI enhanced fear, rats were pre-exposed to seven trials of either pure tones (46 sec/2800 Hz/75 dB) or white noise (46 sec/75 dB) CSs one day prior to training. Auditory fear conditioning occurred in the same context (context A) and was based on the strong delay protocol in Reger et al., 2012 (10 CS-US pairings/46 sec CS; 2 sec/0.9 mA footshock US), which produced increased contextual fear conditioning after FPI. Over the following three days, all groups were tested for contextual fear extinction (context A; 20 min/day). The next day, both groups were tested in context B for auditory cue fear memory with 7 CS-alone trials. Experimental designs for behavioral experiments are outlined in Figs 1A and 2A.
Arc immunohistochemistry
To examine the effects of how white noise is processed in the injured brain within auditory fear neurocircuitry, a 2 × 2 design was used where 48 h after surgery, FPI and sham rats were presented with white noise stimuli (7 trials/46 sec/75 dB) or quiet context exposure for the same duration. Ninety minutes later, all animals were euthanized and brains were processed for immediate early gene immunohistochemistry for activity-regulated cytoskeletal-associated protein (Arc) protein. Briefly, 90 min following white noise or chamber exposure, rats were retrieved from home cages and anesthetized with isoflurane until respiration ceased and were then transcardially perfused with 0.1 M phosphate buffered saline (PBS) followed by 4% paraformaldehyde. Brains were collected and postfixed in 4% paraformaldehyde for 2 h and subsequently cryoprotected in 30% sucrose over 2–3 d then flash frozen and stored in −80 °C until sectioning. Frozen sections (40 μm) containing the amygdala and dorsal hippocampus were stored in antifreeze (30% glycerol/30% ethylene glycol) at −20 °C until they were processed for immunohistochemistry. For Arc immunohistochemistry, selected tissue sections were washed in 0.1 M PBS and incubated in quenching solution (30% H202), washed and incubated in blocking solution for 1 h at room temperature (5% normal goat serum, 0.4% Triton X-100, and 1% bovine serum albumin). Sections were then incubated overnight at 4 °C in primary antibody (anti-Arc, 1:1000, Synaptic Systems). The next day, sections were washed in 0.1 M PBS and incubated in biotinylated secondary antibody (goat anti-rabbit, 1:200, Invitrogen) for 1 h at room temperature. After three washes, staining was amplified using VectaStain ABC kit (Vector Laboratories) and developed in DAB peroxidase substrate (Vector Laboratories). Sections were mounted on electrostatic slides (Fisher) and coverslipped with permount. Arc cell counts for lateral amygdala (LA) and dorsal dentate gyrus (DG) were manually scored using NIH ImageJ by trained observers blind to experimental conditions. Averages from each region and each hemisphere within subject were averaged as one value, counted from three sections per animal.
Anatomical tracing and activation analysis
To determine injury-induced changes in activity within LA projecting neurons that are known to be required for auditory fear, we looked at Arc expression in LA projecting neurons during white noise exposure after FPI. Three weeks prior to FPI surgery, rats (n = 20) received bilateral stereotaxic infusions of cholera toxin B (CTB; ThermoFisher; 5 µg/µl) into the lateral amygdala (LA; A/P: −2.7 mm; M/L: + 5.0 mm; D/V: −8.2 mm relative to bregma). Similar to the previous experiments, 48 h after FPI, all rats were exposed to 7 trials of 75 dB white noise and 90 min later rats were perfused. Brains were processed for Arc immunofluorescence histochemistry. Selected tissue sections (40 µm were washed in 0.1 M PBS and incubated in blocking solution for 1 h at room temperature (5% normal goat serum, 0.4% Triton X-100). Sections were then incubated overnight at 4 °C in primary antibody (anti-Arc, 1:1000, Synaptic Systems). The next day, sections were washed in 0.1 M PBS and incubated in secondary antibody (goat anti-rabbit Alexa fluor 594, 1:500, Life Technologies) for 1 h at room temperature. Sections were washed in 0.1 M PBS, mounted on electrostatic slides (Fisher), and coverslipped with Vectashield (Fisher Scientific). Percentage of Arc + cells in LA-projecting cells (CTB + ) was analyzed in regions that provide auditory information to the LA79, the auditory thalamus (medial geniculate nucleus; MGN) and secondary auditory cortex (Te3).
Data analysis
Behavioral data were analyzed using mixed factors analysis of variance (ANOVA) for injury group (sham, FPI) across trials. Arc data were analyzed by a two way between groups multivariate ANOVA for injury group (sham, FPI), and noise condition (no noise, + noise) as independent variables for each brain region and hemisphere separately (LA and dorsal DG for both ipsilateral (ipsi) and contralateral (contra) relative to injury). CTB-Arc data were analyzed as a multivariate ANOVA for injury across brain regions separately (ipsi and contra MGN and Te3). Statistical significance was determined at p-value of 0.05 or less, and when significant interactions were detected, post hoc analyses were performed for simple main effects.
References
Vakil, E. The effect of moderate to severe traumatic brain injury (TBI) on different aspects of memory: a selective review. Journal of Clinical and Experimental Neuropsychology 27, 977–1021, https://doi.org/10.1080/13803390490919245 (2005).
Gil, S., Caspi, Y., Ben-Ari, I. Z., Koren, D. & Klein, E. Does memory of a traumatic event increase the risk for posttraumatic stress disorder in patients with traumatic brain injury? A prospective study. Am. J. Psychiatry 162, 963–969, https://doi.org/10.1176/appi.ajp.162.5.963 (2005).
Rao, V. & Lyketsos, C. Neuropsychiatric sequelae of traumatic brain injury. Psychosomatics 41, 95–103, https://doi.org/10.1176/appi.psy.41.2.95 (2000).
Moore, E. L., Terryberry-Spohr, L. & Hope, D. A. Mild traumatic brain injury and anxiety sequelae: a review of the literature. Brain injury 20, 117–132, https://doi.org/10.1080/02699050500443558 (2006).
Vaishnavi, S., Rao, V. & Fann, J. R. Neuropsychiatric problems after traumatic brain injury: unraveling the silent epidemic. Psychosomatics 50, 198–205, https://doi.org/10.1176/appi.psy.50.3.198 (2009).
Rosenthal, M., Christensen, B. K. & Ross, T. P. Depression following traumatic brain injury. Arch. Phys. Med. Rehabil. 79, 90–103 (1998).
Fleminger, S. Long-term psychiatric disorders after traumatic brain injury. Eur. J. Anaesthesiol. 42, 123–130, https://doi.org/10.1017/S0265021507003250 (2008).
Yurgil, K. A. et al. Association between traumatic brain injury and risk of posttraumatic stress disorder in active-duty Marines. JAMA Psychiatry 71, 149–157, https://doi.org/10.1001/jamapsychiatry.2013.3080 (2014).
Fanselow, M. S. & LeDoux, J. E. Why we think plasticity underlying Pavlovian fear conditioning occurs in the basolateral amygdala. Neuron 23, 229–232 (1999).
LeDoux, J., Cicchetti, P., Xagoraris, A. & Romanski, L. M. The lateral amygdaloid nucleus: sensory interface of the amygdala in fear conditioning. J. Neurosci. 10, 1062–1069 (1990).
Reger, M. L. et al. Concussive brain injury enhances fear learning and excitatory processes in the amygdala. Biological psychiatry 71, 335–343, https://doi.org/10.1016/j.biopsych.2011.11.007 (2012).
Almeida-Suhett, C. P. et al. Reduced GABAergic inhibition in the basolateral amygdala and the development of anxiety-like behaviors after mild traumatic brain injury. PLoS One 9, e102627, https://doi.org/10.1371/journal.pone.0102627 (2014).
Zuckerman, A. et al. Controlled Low-Pressure Blast-Wave Exposure Causes Distinct Behavioral and Morphological Responses Modelling Mild Traumatic Brain Injury, Post-Traumatic Stress Disorder, and Comorbid Mild Traumatic Brain Injury-Post-Traumatic Stress Disorder. Journal of neurotrauma, https://doi.org/10.1089/neu.2015.4310 (2016).
Hoffman, A. N. et al. Early and Persistent Dendritic Hypertrophy in the Basolateral Amygdala following Experimental Diffuse Traumatic Brain Injury. Journal of neurotrauma 34, 213–219, https://doi.org/10.1089/neu.2015.4339 (2017).
Anagnostaras, S. G., Gale, G. D. & Fanselow, M. S. Hippocampus and contextual fear conditioning: recent controversies and advances. Hippocampus 11, 8–17, 10.1002/1098-1063(2001)11:1<8::AID-HIPO1015>3.0.CO;2–7 (2001).
Fanselow, M. S. Contextual fear, gestalt memories, and the hippocampus. Behav. Brain Res. 110, 73–81 (2000).
Wiltgen, B. J., Sanders, M. J., Anagnostaras, S. G., Sage, J. R. & Fanselow, M. S. Context fear learning in the absence of the hippocampus. J. Neurosci. 26, 5484–5491, https://doi.org/10.1523/JNEUROSCI.2685-05.2006 (2006).
Smith, M. E. Bilateral hippocampal volume reduction in adults with post-traumatic stress disorder: a meta-analysis of structural MRI studies. Hippocampus 15, 798–807, https://doi.org/10.1002/hipo.20102 (2005).
Shin, L. M., Rauch, S. L. & Pitman, R. K. Amygdala, medial prefrontal cortex, and hippocampal function in PTSD. Annals of the New York Academy of Sciences 1071, 67–79, https://doi.org/10.1196/annals.1364.007 (2006).
Pitman, R. K. et al. Biological studies of post-traumatic stress disorder. Nat. Rev. Neurosci. 13, 769–787, https://doi.org/10.1038/nrn3339 (2012).
Vespa, P. M. et al. Nonconvulsive seizures after traumatic brain injury are associated with hippocampal atrophy. Neurology 75, 792–798, https://doi.org/10.1212/WNL.0b013e3181f07334 (2010).
Gao, X., Deng, P., Xu, Z. C. & Chen, J. Moderate traumatic brain injury causes acute dendritic and synaptic degeneration in the hippocampal dentate gyrus. PLoS One 6, e24566, https://doi.org/10.1371/journal.pone.0024566 (2011).
Paterno, R., Folweiler, K. A. & Cohen, A. S. Pathophysiology and Treatment of Memory Dysfunction After Traumatic Brain Injury. Curr. Neurol. Neurosci. Rep. 17, 52, https://doi.org/10.1007/s11910-017-0762-x (2017).
Hoffman, A. N. & Taylor, A. N. Stress reactivity after traumatic brain injury: implications for comorbid post-traumatic stress disorder. Behav. Pharmacol., https://doi.org/10.1097/FBP.0000000000000461 (2019).
Hoffer, M. E. Mild traumatic brain injury: neurosensory effects. Current Opinion in Neurology 28, 74–77, https://doi.org/10.1097/WCO.0000000000000164 (2015).
Kraus, N. et al. Auditory biological marker of concussion in children. Scientific reports 6, 39009, https://doi.org/10.1038/srep39009 (2016).
Zemek, R. et al. Clinical Risk Score for Persistent Postconcussion Symptoms Among Children With Acute Concussion in the ED. JAMA 315, 1014–1025, https://doi.org/10.1001/jama.2016.1203 (2016).
Kontos, A. P. et al. A revised factor structure for the post-concussion symptom scale: baseline and postconcussion factors. American Journal of Sports Medicine 40, 2375–2384, https://doi.org/10.1177/0363546512455400 (2012).
McNamara, K. C., Lisembee, A. M. & Lifshitz, J. The whisker nuisance task identifies a late-onset, persistent sensory sensitivity in diffuse brain-injured rats. Journal of neurotrauma 27, 695–706, https://doi.org/10.1089/neu.2009.1237 (2010).
Thomas, T. C., Hinzman, J. M., Gerhardt, G. A. & Lifshitz, J. Hypersensitive glutamate signaling correlates with the development of late-onset behavioral morbidity in diffuse brain-injured circuitry. Journal of neurotrauma 29, 187–200, https://doi.org/10.1089/neu.2011.2091 (2012).
Ceranic, B. J., Prasher, D. K., Raglan, E. & Luxon, L. M. Tinnitus after head injury: evidence from otoacoustic emissions. Journal of Neurology, Neurosurgery, and Psychiatry 65, 523–529 (1998).
Waddell, P. A. & Gronwall, D. M. Sensitivity to light and sound following minor head injury. Acta Neurologica Scandinavica 69, 270–276 (1984).
Lowenstein, D. H., Thomas, M. J., Smith, D. H. & McIntosh, T. K. Selective vulnerability of dentate hilar neurons following traumatic brain injury: a potential mechanistic link between head trauma and disorders of the hippocampus. J. Neurosci. 12, 4846–4853 (1992).
Bernier, B. E. et al. Dentate Gyrus Contributes to Retrieval as well as Encoding: Evidence from Context Fear Conditioning, Recall, and Extinction. J. Neurosci. 37, 6359–6371, https://doi.org/10.1523/JNEUROSCI.3029-16.2017 (2017).
Miyashita, T., Kubik, S., Haghighi, N., Steward, O. & Guzowski, J. F. Rapid activation of plasticity-associated gene transcription in hippocampal neurons provides a mechanism for encoding of one-trial experience. Jounal of Neuroscience 29, 898–906, https://doi.org/10.1523/JNEUROSCI.4588-08.2009 (2009).
Ploski, J. E. et al. The activity-regulated cytoskeletal-associated protein (Arc/Arg3.1) is required for memory consolidation of pavlovian fear conditioning in the lateral amygdala. J. Neurosci. 28, 12383–12395, https://doi.org/10.1523/JNEUROSCI.1662-08.2008 (2008).
Gouty-Colomer, L. A. et al. Arc expression identifies the lateral amygdala fear memory trace. Molecular psychiatry 21, 364–375, https://doi.org/10.1038/mp.2015.18 (2016).
Romanski, L. M. & LeDoux, J. E. Equipotentiality of thalamo-amygdala and thalamo-cortico-amygdala circuits in auditory fear conditioning. J. Neurosci. 12, 4501–4509 (1992).
Fanselow, M. S. Signaled shock-free periods and preference for signaled shock. Journal of Experimental Psychology: Animal Behavior Processes 6, 65–80 (1980).
Rescorla, R. A. In The psychology of learning and motivation: Advances in research and theory Vol. 6 (ed. Bower, G. H.) (Academic Press, 1972).
Rescorla, R. A. & Wagner, A. R. In Classical conditioning II: current research and theory (eds Black, A. H. & Prokasy, W. F.) (Appleton-Century-Crofts, 1972).
Fanselow, M. S. From contextual fear to a dynamic view of memory systems. Trends in cognitive sciences 14, 7–15, https://doi.org/10.1016/j.tics.2009.10.008 (2010).
Maren, S., Phan, K. L. & Liberzon, I. The contextual brain: implications for fear conditioning, extinction and psychopathology. Nat. Rev. Neurosci. 14, 417–428, https://doi.org/10.1038/nrn3492 (2013).
Borszcz, G. S., Cranney, J. & Leaton, R. N. Influence of long-term sensitization on long-term habituation of the acoustic startle response in rats: central gray lesions, preexposure, and extinction. Journal of Experimental Psychology: Animal Behavior Processes 15, 54–64 (1989).
Helmstetter, F. J. & Bellgowan, P. S. Hypoalgesia in response to sensitization during acute noise stress. Behavioral neuroscience 108, 177–185 (1994).
Bellgowan, P. S. & Helmstetter, F. J. Neural systems for the expression of hypoalgesia during nonassociative fear. Behavioral neuroscience 110, 727–736 (1996).
Leaton, R. N. & Cranney, J. Potentiation of the acoustic startle response by a conditioned stimulus paired with acoustic startle stimulus in rats. Journal of Experimental Psychology: Animal Behavior Processes 16, 279–287 (1990).
Park, S. et al. Sound tuning of amygdala plasticity in auditory fear conditioning. Scientific reports 6, 31069, https://doi.org/10.1038/srep31069 (2016).
Fadok, J. P. et al. A competitive inhibitory circuit for selection of active and passive fear responses. Nature 542, 96–100, https://doi.org/10.1038/nature21047 (2017).
Zhang, G. W. et al. A Non-canonical Reticular-Limbic Central Auditory Pathway via Medial Septum Contributes to Fear Conditioning. Neuron 97, 406–417 e404, https://doi.org/10.1016/j.neuron.2017.12.010 (2018).
Ledoux, J., Ruggiero, D. A., Forest, R., Stornetta, R. & Reis, D. J. Topographic organization of convergent projections to the thalamus from the inferior colliculus and spinal cord in the rat. Journal of Comparative Neurology 264, 123–146, https://doi.org/10.1002/cne.902640110 (1987).
Shi, C. & Davis, M. Pain pathways involved in fear conditioning measured with fear-potentiated startle: lesion studies. J. Neurosci. 19, 420–430 (1999).
Wojcik, S. M. Predicting mild traumatic brain injury patients at risk of persistent symptoms in the Emergency Department. Brain Inj. 28, 422–430, https://doi.org/10.3109/02699052.2014.884241 (2014).
Mirz, F., Gjedde, A., Sodkilde-Jrgensen, H. & Pedersen, C. B. Functional brain imaging of tinnitus-like perception induced by aversive auditory stimuli. Neuroreport 11, 633–637 (2000).
Green, S. A. et al. Overreactive brain responses to sensory stimuli in youth with autism spectrum disorders. J. Am. Acad. Child Adolesc. Psychiatry 52, 1158–1172, https://doi.org/10.1016/j.jaac.2013.08.004 (2013).
McHugh, T. J. et al. Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network. Science 317, 94–99, https://doi.org/10.1126/science.1140263 (2007).
Vazdarjanova, A. et al. Spatial exploration induces ARC, a plasticity-related immediate-early gene, only in calcium/calmodulin-dependent protein kinase II-positive principal excitatory and inhibitory neurons of the rat forebrain. Journal of Comparative Neurology 498, 317–329, https://doi.org/10.1002/cne.21003 (2006).
Temple, M. D., Worley, P. F. & Steward, O. Visualizing changes in circuit activity resulting from denervation and reinnervation using immediate early gene expression. J. Neurosci. 23, 2779–2788 (2003).
Sick, T. J., Perez-Pinzon, M. A. & Feng, Z. Z. Impaired expression of long-term potentiation in hippocampal slices 4 and 48 h following mild fluid-percussion brain injury in vivo. Brain research 785, 287–292 (1998).
D’Ambrosio, R., Maris, D. O., Grady, M. S., Winn, H. R. & Janigro, D. Selective loss of hippocampal long-term potentiation, but not depression, following fluid percussion injury. Brain research 786, 64–79 (1998).
Smith, D. H., Lowenstein, D. H., Gennarelli, T. A. & McIntosh, T. K. Persistent memory dysfunction is associated with bilateral hippocampal damage following experimental brain injury. Neurosci. Lett. 168, 151–154 (1994).
Kim, J. J. & Fanselow, M. S. Modality-specific retrograde amnesia of fear. Science 256, 675–677 (1992).
Cushman, J. D. et al. Juvenile neurogenesis makes essential contributions to adult brain structure and plays a sex-dependent role in fear memories. Frontiers in behavioral neuroscience 6, 3, https://doi.org/10.3389/fnbeh.2012.00003 (2012).
Carpenter-Thompson, J. R., Akrofi, K., Schmidt, S. A., Dolcos, F. & Husain, F. T. Alterations of the emotional processing system may underlie preserved rapid reaction time in tinnitus. Brain research 1567, 28–41, https://doi.org/10.1016/j.brainres.2014.04.024 (2014).
Chen, Y. C. et al. Tinnitus and hyperacusis involve hyperactivity and enhanced connectivity in auditory-limbic-arousal-cerebellar network. elife 4, e06576, https://doi.org/10.7554/eLife.06576 (2015).
Dischinger, P. C., Ryb, G. E., Kufera, J. A. & Auman, K. M. Early predictors of postconcussive syndrome in a population of trauma patients with mild traumatic brain injury. Journal of Trauma 66, 289–296, https://doi.org/10.1097/TA.0b013e3181961da2 (2009).
Balba, N. M. et al. Increased Sleep Disturbances and Pain in Veterans With Comorbid Traumatic Brain Injury and Posttraumatic Stress Disorder. J. Clin. Sleep Med. 14, 1865–1878, https://doi.org/10.5664/jcsm.7482 (2018).
Papesh, M. A. et al. Blast Exposure Impairs Sensory Gating: Evidence from Measures of Acoustic Startle and Auditory Event-Related Potentials. Journal of neurotrauma 36, 702–712, https://doi.org/10.1089/neu.2018.5801 (2019).
Green, S. A., Hernandez, L., Bookheimer, S. Y. & Dapretto, M. Reduced modulation of thalamocortical connectivity during exposure to sensory stimuli in ASD. Autism Res. 10, 801–809, https://doi.org/10.1002/aur.1726 (2017).
Carron, S. F., Alwis, D. S. & Rajan, R. Traumatic Brain Injury and Neuronal Functionality Changes in Sensory Cortex. Front. Syst. Neurosci. 10, 47, https://doi.org/10.3389/fnsys.2016.00047 (2016).
Lucas, S., Hoffman, J. M., Bell, K. R., Walker, W. & Dikmen, S. Characterization of headache after traumatic brain injury. Cephalalgia 32, 600–606, https://doi.org/10.1177/0333102412445224 (2012).
Frost, R. B., Farrer, T. J., Primosch, M. & Hedges, D. W. Prevalence of traumatic brain injury in the general adult population: a meta-analysis. Neuroepidemiology 40, 154–159, https://doi.org/10.1159/000343275 (2013).
Thompson, H. J. et al. Lateral fluid percussion brain injury: a 15-year review and evaluation. Journal of neurotrauma 22, 42–75, https://doi.org/10.1089/neu.2005.22.42 (2005).
Hovda, D. A., Yoshino, A., Kawamata, T., Katayama, Y. & Becker, D. P. Diffuse prolonged depression of cerebral oxidative metabolism following concussive brain injury in the rat: a cytochrome oxidase histochemistry study. Brain research 567, 1–10 (1991).
Yoshino, A., Hovda, D. A., Kawamata, T., Katayama, Y. & Becker, D. P. Dynamic changes in local cerebral glucose utilization following cerebral conclusion in rats: evidence of a hyper- and subsequent hypometabolic state. Brain research 561, 106–119 (1991).
Fineman, I., Giza, C. C., Nahed, B. V., Lee, S. M. & Hovda, D. A. Inhibition of neocortical plasticity during development by a moderate concussive brain injury. Journal of neurotrauma 17, 739–749 (2000).
Ip, E. Y., Giza, C. C., Griesbach, G. S. & Hovda, D. A. Effects of enriched environment and fluid percussion injury on dendritic arborization within the cerebral cortex of the developing rat. Journal of neurotrauma 19, 573–585, https://doi.org/10.1089/089771502753754055 (2002).
Osteen, C. L., Giza, C. C. & Hovda, D. A. Injury-induced alterations in N-methyl-D-aspartate receptor subunit composition contribute to prolonged 45calcium accumulation following lateral fluid percussion. Neuroscience 128, 305–322, https://doi.org/10.1016/j.neuroscience.2004.06.034 (2004).
Maren, S. & Quirk, G. J. Neuronal signalling of fear memory. Nat. Rev. Neurosci. 5, 844–852, https://doi.org/10.1038/nrn1535 (2004).
Acknowledgements
This work was supported in part by F32NS098694: AH; R01MH062122: MF; UCLA Depression Grand Challenge Fellowship Fund: MF; Staglin Center for Brain and Behavioral Health: MF; UCLA Brain Injury Research Center; 1PO1NS058489: DH, CG; 1R01NS27544: DH, CG; Centre for NeuroSkills: DH; Joseph Drown Foundation, UCLA Steve Tisch BrainSPORT: CG; Easton Labs for Brain Health. The authors would like to thank Yan Cai, Zach Pennington, Michael Conoscenti, and Sonya Watson for technical assistance.
Author information
Authors and Affiliations
Contributions
A.N.H. designed experiments, performed surgeries, behavioral experiments, tissue processing, imaging, quantification, data analysis, and prepared manuscript; J.L. assisted with behavioral experiments; D.A.H. and C.C.G. assisted with experimental design and manuscript preparation; M.S.F. advised on experimental design, analysis, and manuscript preparation.
Corresponding author
Ethics declarations
Competing Interests
Disclosures unrelated to this project: A.N.H. and C.C.G. have received past research funding from Avanir Pharmaceuticals. M.S.F. is director of research for Neurovation Labs. C.C.G. receives consulting fees from NFL-Neurological Care Program, NHLPA, serves on the advisory panel for the Major League Soccer, NBA, NCAA, US Soccer Federation, California State Athletic Commission, Highmark International, and has received speaker fees from Medical Education Speakers Network and book royalties from Blackwell Publishing: Prioritized Neurological Differential Diagnosis.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hoffman, A.N., Lam, J., Hovda, D.A. et al. Sensory sensitivity as a link between concussive traumatic brain injury and PTSD. Sci Rep 9, 13841 (2019). https://doi.org/10.1038/s41598-019-50312-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-50312-y
This article is cited by
-
Biomarkers and Endophenotypes of Post-traumatic Headaches
Current Pain and Headache Reports (2024)
-
Models for Treating Post-traumatic Headache
Current Pain and Headache Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
