
Abstract
In lowland areas of Malaysia, Plasmodium knowlesi infection is associated with land use change and high proportions of the vector Anopheles balabacensis. We conducted a 15-month study in two Malaysian villages to determine the effect of habitat on vector populations in understudied high-altitude, high-incidence districts. Anopheles mosquitoes were sampled in human settlements, plantations and forest edges, and screened for Plasmodium species by PCR. We report the first An. donaldi positive for P. knowlesi. This potential vector was associated with habitat fragmentation measured as disturbed forest edge:area ratio, while An. balabacensis was not, indicating fragmented land use could favour An. donaldi. Anopheline species richness and diversity decreased from forest edge, to plantation, to human settlement. Greater numbers of An. balabacensis and An. donaldi were found in forest edges compared to human settlements, suggesting exposure to vectors and associated zoonoses may be greater for people entering this habitat.
Similar content being viewed by others

Introduction
In South East Asia, the long-tailed macaque (Macaca fascicularis) is the main reservoir of at least five simian malarias, namely, Plasmodium coatneyi, P. inui, P. fieldi, P. cynomolgi and P. knowlesi1,2. Of these, P. knowlesi now routinely infects humans. This zoonotic malaria is particularly problematic in Sabah, Malaysian Borneo, where it is currently the prevalent cause of clinical malaria3,4. Another of these species, P. cynomolgi, previously only shown experimentally to infect humans, has been reported in naturally acquired human cases, first as a single case in peninsular Malaysia5 and, more recently, in five cases in Kapit district of Sarawak, Malaysian Borneo6. Thus, as previously predicted7, simian malaria parasites are now contributing to clinical malaria in humans, derailing regional efforts to eradicate the disease. This is of particular public health importance in Malaysia, which aims to eradicate malaria by 2020.
In 2016, the World Health Organization Malaria Policy Advisory Committee reported that P. knowlesi was the predominant malaria species in Malaysia, comprising 69% of cases8. In Sabah state, this species was responsible for the majority of malaria cases, with 815 and 996 cases reported in 2012 and 2013, respectively9. The most recent study showed that the proportion of P. knowlesi among indigenous malaria cases has continued to increase, rising from 80% in 2015 to 88% in 2016, and then again in 2017 to 98% of all malaria admissions in the state10.
Recent findings have demonstrated a clear link between land use change and P. knowlesi incidence in this region, which strongly supports the idea that an epidemiological change is taking place11. A high P. knowlesi incidence was recorded in Sabah, where land use changes have occurred, such as opening of forest areas for commercial plantations and logging, with the proportion of P. knowlesi of all malaria cases in Sabah for 2014-2018 being 0.66, 0.71, 0.69, 0.88 and 0.89 respectively (2584/3925, 1640/2323, 1600/2318, 3614/4114 and 4131/4630 respectively)12. The large-scale land use changes observed in the region are likely to impact the ecology, behavior and transmission potential of Anopheles mosquitoes as vectors of zoonotic malaria13. Yet data on the degree of contact between vectors and human hosts and how this differs according to land use type in the context of rapid anthropogenic environmental change are not well understood14. Understanding current human infection risk from simian malaria species and anticipating possible future zoonotic spillover of vector-borne disease calls for a more comprehensive examination of local anopheline population ecology, as well as surveillance of their Plasmodium infection status.
We hypothesized that vector composition would change across an ecotone of increasing forest disturbance. We therefore set out to characterize the abundance, diversity and transmission potential of Anopheles species, especially known Plasmodium vectors, biting humans in three habitat types representative of predominant land uses in rural Sabah, namely human settlements, plantations and forest edges. To do this, we carried out a comparative study focusing on Anopheles mosquitoes in two high altitude areas in understudied regions of Sabah that nonetheless experience a high burden of human infection with P. knowlesi (Supplementary Table S1) and are undergoing environmental change. Our results are discussed with reference to known Anopheles species compositions in areas with well-documented vector profiles and the wider challenge of predicting changes in vector populations that may arise from deforestation and agricultural development.
Methods
Study sites
The study was carried out in Ranau and Keningau (Table 1), two inland districts situated at higher altitudes than Kudat where most previous studies on P. knowlesi in Sabah have been conducted. Both areas reported human cases in the year prior to data collection15.
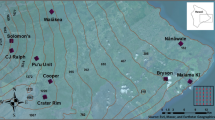
Ranau district is in the west coast division of Sabah, about 108 km east of Kota Kinabalau. The northern part of Ranau is bounded by the Crocker Range and the Pinousok summit. Sampling was undertaken in Kampung Paus, which is approximately 80 km from Ranau town (Fig. 1). Keningau district is situated in a valley bordered by the Crocker Range to the west and the Trus Madi Range to the east and south. Sampling here was performed at Kampung Keritan Ulu, approximately 32 km from Keningau town. Both villages have and are surrounded by abundant and varied potential breeding sites for Anopheles species, including shaded and sunlit forest ground pools, irrigated rice fields, rock pools adjacent to rivers, stumps in bamboo stands, springs, water-filled animal hoof prints and ephemeral puddles.

Location of study sites in Ranau and Keningau. 1 = Kampung Paus, 2 = Kampung Keritan Ulu.
Collection of Anopheles using human landing catches
At both villages, mosquitoes were sampled in three habitat types: human settlements, plantations (including oil palm, rubber and fruit orchards) and forest edges, with two sample points for each habitat type in each village, totaling 12 sample locations. All sample points were at least 50 m from the next closest sampling point and positioned outside away from buildings and paths, except for at human settlements, where collections were made within 5 m of a dwelling. Monthly mosquito collections started in August 2015 and finished in November 2016. Collections were made for two successive nights at all sampling points in one village simultaneously, and then at the second village the following week, every month for 15 months data, except in October 2016 when data could not be collected, giving a total of 360 discrete catches over 60 collection nights.
Mosquitoes were sampled using the human landing catch (HLC) method, as this is still the most efficient method tested locally16, with only Anopheles species retained for further analysis. In HLCs, a volunteer collected mosquitoes by exposing their lower legs (from knee downwards) and collecting each mosquito that landed on them in a plastic specimen tube (2 cm diameter × 6 cm) with a piece of moist tissue at the bottom. Each HLC was performed by two persons who collected mosquitoes for six consecutive hours, from 18:00 to 00:00 hrs, thus covering the peak biting time of most Anopheles species17, with collectors rotated between each station. The HLC collectors were regularly and randomly monitored by a supervisor. The morning following collection, Anopheles mosquitoes were mounted and placed individually in a labeled Eppendorf tube, including hour of collection to record biting profile over the course of the collection period. Samples were then taken to the laboratory at Universiti Malaysia Sabah campus in Kota Kinabalu for further processing.
The HLC done by the volunteers were performed in accordance with relevant guidelines and regulations as approved by the Ethics committee.
Identification of Anopheles species
Anopheles specimens were identified morphologically to species within 24 hours using published identification keys. For the Leucosphyrus group, the key of Sallum et al.18 was used and for other groups the keys developed by Rattanarithikul et al.19 were used. The identified specimens were kept individually in clean 1.5 mL microfuge tubes and stored at −20 °C until used for molecular analysis.
Total DNA extraction and PCR of malaria parasites
Each Anopheles specimen was placed inside a clean 1.5 mL microfuge tube and the tissue was homogenized using a homogenizer. The total DNA was extracted from each specimen following the method of Phillips and Simon20 and stored at −30 °C until PCR analysis. Detection of malaria parasites was performed using the nested PCR Plasmodium genus-specific method described by Singh et al.21. When a sample was found positive for malaria parasites, a second nested PCR was performed to determine the Plasmodium species using species-specific primers in singleplex PCR1,5,21,22. Primers of nine species of Plasmodium were used in this study: P. coatneyi, P. inui, P. fieldi, P. cynomolgi, P. knowlesi, P. falciparum, P. vivax, P. malariae and P. ovale. All these species have been recorded in Malaysia, although P. ovale is an imported species.
Both first and second PCRs were performed with 25 μl final volume. The reaction components were prepared by mixing 5.0 μl of 5X PCR buffer, 0.5 μl of dNTPs (10 mM) mixture, 3.0 μl of 25 mM MgCl2, 1.0 μl each of 10 μM forward and reverse primers, 0.3 μl of Taq DNA polymerase (5 U/μl), 2.0 μl of DNA template and sterile dH2O up to 25 μl final volume. After the first PCR was completed, 2.0 μl of the first PCR product was used as a template in the second PCR. The PCR conditions used were: an initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing for 1 min, and extension at 72 °C for 1 min, and a final extension at 72 °C for 5 min. The annealing temperature was set based on the optimum temperature of the primers (Supplementary Table S2).
Landscape classification and quantification
To determine the landscape characteristics of the study areas, very high-resolution satellite images were acquired from Bing Maps and Google Earth for areas covering the collection points in Kampung Paus (Ranau) and Kampung Keritan Ulu (Keningau). Imagery dates closest to the start of data collection were July 2014 for Ranau and April 2015 for Keningau. Polygons representing areas of uniform land cover type were manually digitized from the imagery by an experienced spatial information scientist (AC) using ArcGIS Pro v.10.4.1 software and class defined to emphasize variation in structure (Fig. 2a,b). Forest cover, denoted as disturbed forest, was observed to have varying degrees of human disturbance activity throughout both regions and, thus all forest cover is not considered primary forest. Disturbed shrublands were identified as areas of shorter and less dense vegetation than areas of disturbed forests. However, these areas could represent forest recovery post-disturbance. Human settlements were identified by the presence for buildings and cleared area surround these buildings not apparently in use for agriculture. Plantations were identified as having tree cover that is of apparent uniform tree types and lower canopy variation than disturbed forest, palm trees being the most apparent tree type upon visual interpretation. Plantation areas where verified from ground-based observation. Plantation density designations of low, medium or high were based on the density of canopy cover; low representing less than 30%, medium 30%–60%, and high >60%. These density designations also denote an assumed structural variation, which high plantations having the largest trees and medium and low designations with underdeveloped trees. Agriculture was apparent in satellite imagery and rice fields added from ground-based observation. Additional classes of Road, Pond, and River were clearly identifiable from visual interpretation.

Land cover classifications in Ranau and Keningau. (a) Kampung Paus in Ranau district. (b) Kampung Keritan Ulu in Keningau Distrcit. Red markers indicate mosquito collection points at human settlements (triangles), plantations (squares) and forest edges (circles).
Locations of mosquito collection points, recorded in the field by GPS, were added to the landcover maps and overlaid with a 500 m buffer, which was clipped from the map to define the habitat local to each collection point. Within each buffer, the cumulative area of disturbed forest, plantation and human settlement was calculated to determine percent landcover. Additionally, perimeter length of disturbed forest was determined to derive estimates of the forest edge:area ratio, indicating local forest fragmentation, where more fragmented forest (smaller, more isolated patches) have a larger forest edge:area ratio23.
Mosquito diversity and statistical analysis
Analyses were performed using the R programming language for statistical analysis (version 3.2.2)24. Species diversity was investigated using four indices for disturbed forest, plantation and human settlement, based on all Anopheles collected. These were total species richness (S) and rarefied species richness Sr, which accounts for the increased likelihood of sampling rare species in larger sample sizes and is the species richness normalized to the smallest sample size from an ecotype, Simpson’s Diversity Index (1 − D), Shannon-Weiner Diversity Index (H), and evenness (E). Diversity calculations were derived from R package “vegan”25.
The variation in Anopheles species abundance between different habitats and villages was analysed by negative binomial Generalized Linear Mixed Models (GLMMs). Habitat was set as a fixed effect and sampling night as a random effect. Models incorporating zero-inflation parameters were tested, but this was found to have a non-significant effect on model fit and was not included as a parameter in the model. Preliminary GLMM analysis showed that the negative binomial distributions gave better than, or similar, fits to the Poisson distribution. Tukey contrasts (α = 0.05) were used to compare differences in species abundance between habitats and villages.
To assess the effect of landcover on mosquito abundance, data were transformed to log(catch + 1) and analysed using a linear model, with habitat type (forest edge, plantation, human settlement) and forest edge:area ratio within a 500 m buffer of the collection point set as factors.
Ethical clearance
This project was approved by the Ethics Committee of Universiti Malaysia Sabah (JKEtika 1/16(3)). All volunteers who carried out mosquito collections gave informed consent and were offered antimalarial prophylaxis throughout the study period.
Results
Composition of Anopheles species
A total of 1,071 Anopheles mosquitoes were collected, representing fourteen different species (Table 2). The total numbers of species caught in each village were similar (10 in Keningau and 11 in Ranau), although more individuals were caught at Ranau (620 versus 451 at Keningau). The five most abundant species in both villages were the same: An. balabacensis, An. barbumbrosus, An. donaldi, An. maculatus and An. tessellatus. However, the ranking of species abundance was different: in Keningau, An. maculatus (26.2%) was dominant, but it constituted only 2.4% of the total catch in Ranau. Conversely, in Ranau, An. donaldi was the dominant species (40.5%), whereas it accounted for 23% of Anopheles caught in Keningau. In both villages, the proportion of An. balabacensis was similar (23-25%) and more mosquitoes (45-49%) were caught in the forest edge than the other habitats.
Species diversity
Anopheline species richness decreased from sites at forest edges, to plantations, to settlements, and even when adjusted to the lowest sample size (228 individuals in human settlements), rarefied richness was found to be lowest in human settlements (Table 3). Both Simpson’s and Shannon-Weiner diversity indices were highest at forest edges and lowest in human settlements (Table 3).
Variation in catch with habitat
Landcover classifications for the two study villages are shown in Fig. 2. In Kampung Paus in Ranau (Fig. 2a), within the 500 m buffer surrounding collection sites, disturbed forest accounted for average landcover of 7% around human settlement sites, 12% around plantation sites and 22% around forest edge sites. The average percentage of disturbed forest was generally higher in Kampung Keritan Ulu in Keningau (Fig. 2b), with 30%, 37% and 67% around human settlement, plantation and forest edge sites, respectively. Plantation landcover varied less between the two villages. The buffers around human settlement, plantation and forest edge sites contained, respectively, 29%, 26% and 17% plantation in Ranau, and 31%, 20% and 11% plantation in Keningau. The percentage area attributed to human settlement was relatively small in both villages and across all sampling sites; in Ranau this ranged from 12% around forest edge sites, to 13% around human settlement and 14% around plantation, while in Keningau human settlement collection sites had an average of 7% of this landcover type, plantation only 2%, while forest edge sites had no land classified as human settlements in the surrounding 500 m.
Subsequent analysis focuses on the four most abundant species of medical importance identified in Table 2. The mean nightly catch of An. balabacensis, An. donaldi, An. barbumbrosus and An. maculatus in both villages decreased from forest edge, to plantation, to human settlement (Fig. 3). An. maculatus were present in very low numbers in Ranau and trends could not be observed.

Variation in mosquito catch per person per hour (mean ± SE) according to habitat type for the four most common Anopheles species in Ranau and Keningau. HU = human settlement, PL = plantation, FE = forest edge.
Habitat type significantly affected predicted mean catch per night for the four medically important species detected (Table 4). In both study locations, An. balabacensis and An. donaldi were caught more frequently at forest edges than around human settlements. In Keningau, An. maculatus was also caught in higher numbers at forest edges, as was An. barbumbrosus in Ranau. However, the mean numbers of An. barbumbrosus in Keningau and An. maculatus in Ranau were not significantly different between the three habitats.
The relationship between catch and forest fragmentation was analyzed for An. balabacensis and An. donaldi, as these were the only two species found to be positive for P. knowlesi (see ‘Infection rate of Anopheles species’). Across all habitats, significantly more An. donaldi were caught at collection points with a greater forest edge:area ratio in the surrounding disturbed forest (Fig. 4A; F = 22.2; p < 0.001). This was not the case for An. balabacensis, where there was no significant difference in catch with respect to disturbed forest edge:area ratio in the 500 m buffer surrounding each collection point, irrespective of the habitat type (Fig. 4B; F = 1.7; p = 0.18).

Total catch of Anopheles species (mean ± se) in three habitats in relation to the edge-area ratio of surrounding disturbed forest. Mean based on total sampling for 15 months at twelve collection points. Forest edge:area ratio calculated for a 500 m buffer surrounding each collection point. (A) Anopheles donaldi. (B) Anopheles balabacensis.
Temporal variation in catch
Mosquito biting rates varied across the sampling period, showing seasonal peaks for most species, which were caught in greater numbers between November 2015 and January 2016, June and July 2016, and September and November 2016 (Fig. 5A,B). The number of An. balabacensis has a discernible peak in Ranau and Keningau, although the peak occurred in November 2016 in Ranau, but in December 2015 in Keningau. In Ranau, the trends for the other three species are not clear, whereas in Keningau, these species showed bimodal peaks in abundance between December 2015 and January 2016, and again between June and July 2016.

Number of mosquito bites/ person/ night (mean ± SE) across fifteen sampling months in Ranau and Keningau. HU = human settlement, PL = plantation, FE = forest edge. Sampling could not be done in October 2016. Note x-axis scales differ. (A) Kampung Paus in Ranau district. (B) Kampung Keritan Ulu in Keningau district.
Biting time of An. balabacensis peaked at 7-8 p.m. in both Ranau and Keningau and decreased later, although An. donaldi and An. maculatus appeared to bite more at 6-7 p.m. (Fig. 6A,B). In general, the numbers of mosquitoes biting decreased later into the night. Peak biting time in settlements lagged those observed in the other two habitats.

Biting times of Anopheles species (mean bites per person per hour ± SE) in three habitats of Keningau from 6 pm to midnight. HU = human settlement, PL = plantation, FE = forest edge. Mean based on total sampling for 15 months. Note x-axis scales differ. (A) Kampung Paus in Ranau district. (B) Kampung Keritan Ulu in Keningau district.
Infection rate of Anopheles species
A total of 1,069 individuals (99.8% of total catch) were assayed for Plasmodium infection using PCR. These were 253 An. balabacensis, 355 An. donaldi, 133 An. maculatus, 199 An. barbumbrosus, 103 An. tesellatus and 26 of the other rarer species (Supplementary Table S3). Positive results are presented in Table 5. Malaria parasites were only detected in An. balabacensis and An. donaldi. One An. balabacensis from Ranau was found to be positive with P. knowlesi. However, in Keningau, two An. donaldi tested positive for P. knowlesi parasites. Other simian malaria parasites detected were P. cynomolgi (in An. balabacensis and An. donaldi) and P. inui (in An. balabacensis only), with one unidentified Plasmodium species. Six out of seven positive specimens were collected from the forest edge habitat; no positive specimens were found in human settlements.
Discussion
Overall, the species composition of Anopheles was found to be similar between Kampung Paus in Ranau and Kampung Keritan Ulu in Keningau, even though these villages are located approximately 97 km from each other. Both localities share the same major species, namely An. balabacensis, An. donaldi, An. maculatus and An. barbumbrosus, and there is no single predominant species. This contrasts with previous results from Kudat district (Fig. 1), where the confirmed P. knowlesi vector An. balabacensis dominates, typically constituting 90-96% of Anopheles found in HLCs (Supplementary Table S4)17. Kudat is a low-lying area to the north of Sabah and sampling sites in this district tend to be at low elevations relative to Ranau and Keningau, typically around 19-180 m above sea level. Fornace et al.11 found associations between lower elevations and P. knowlesi incidence; in the present study Ranau and Keningau are both high elevation districts with a high number of P. knowlesi cases, but with a low proportion of An. balabacensis relative to other species in mosquito collections (Supplementary Table S1). Although we did not test this empirically, this could suggest that, alongside elevation, other factors may be locally important in mediating disease transmission.
The forest edge had the highest abundance, richness and diversity of mosquito species, while the human settlement the lowest. Old-growth and secondary forest, which occur around both study villages, offer a range of environmental niches suitable for diverse Anopheles species; herein refugia, suitable oviposition sites and other non-human hosts for many Anopheles species are plentiful26,27. It is possible that some of the mosquitoes caught in the settlements may be “vagrant” species flying in from the surrounding forests, as indicated by the apparent time lag of an hour or so in peak biting time for key species in human settlements compared to forest edge and plantations and this warrants spatially explicit investigation. Oil palm plantations are monocultures with minimal flora diversity28, although when established, they may provide anopheline breeding sites from wheel ruts and footpath depressions filled with water and potential refugia within the palms’ structure and their associated epiphytes29 which may explain the intermediate abundance of vectors between the three habitats. However, previous literature suggests this is unlikely, as no Anopheles were found resting in plantations in oil palm and rubber plantations in Kudat30 and malaria vector abundance was found to decrease in areas cultivated for oil palm31. More mosquitoes were trapped during the period between December and February, and May and July, which coincides with Sabah’s two main rainy seasons. The rain would provide more breeding grounds in the form of temporary rain pools, road puddles, puddles in rocks, animal hoof prints in the forests, plantations and at human settlements32,33. It has been shown in Côte d’Ivoire that monthly malaria incidence was positively associated with the quantity of rain one and two months before34. Moderate precipitation tends to increase potential breeding grounds although too much rainfall can flush away breeding habitats temporarily.
Prior to the current study, the only mosquito species found to be infected with P. knowlesi were members of the Leucosphyrus group35. The locally incriminated vector in Sabah is An. balabacensis, which falls within the Leucosphyrus complex (part of the Leucosphyrus group). In our study, we collected two An. donaldi, a member of the Barbirostris group, both of which were confirmed positive for P. knowlesi via PCR. Morphological vector species identification was independently verified by three experienced vector biologists, and PCR for Plasmodium detection was repeated five times. This finding is not sufficient to incriminate An. donaldi as a vector of P. knowlesi to humans, as we used whole mosquito samples for parasite detection, thus the positive samples could be carriers of P. knowlesi, rather than infectious vectors. As An. donaldi is a confirmed vector of human malaria parasites and positive specimens were collected via human landing catches, it does raise questions about the potential role of An. donaldi in zoonotic transmission. If subsequently incriminated through positive detection of parasites in the head or salivary glands, this would have implications for public health attempts to manage P. knowlesi in areas where An. balabacensis and An. donaldi coexist, as their bionomics differ and may require different intervention strategies, particularly as An. donaldi will enter dwellings to feed, highlighting the importance of maintaining bed net coverage.
It may also be the case that variations in response to environmental change vary both between and within species. For example, in Sarawak, in the four years immediately after conversion of a forest area to plantations of oil palm, An. donaldi showed a significant decline in abundance of 64%31, whereas work in the Kinabatangan area of Sabah reported exophagic An. donaldi had replaced An. balabacensis as the dominant vector of P. falciparum following deforestation and the implementation of malaria vector control activities36. In our study, although we did not observe a predominance of An. donaldi, we found a greater abundance of An. donaldi at collection points where the surrounding forest patches had a high degree of fragmentation, so the availability of edge habitats may be favourable to An. donaldi. With respect to An. balabacensis, we did not find evidence that forest fragmentation affected abundance. A recent study showed An. balabacensis preferred disturbed, logged forest over primary forest37. Together these results imply a degree of resilience to degraded forests, irrespective of edge availability or size of patch, although the mechanism underlying this would require further exploration. Precise landscape characteristics, and anthropogenic changes in the biotic and abiotic composition of habitats, may therefore be critical in determining how suitable the overall forest-matrix environment is to the success of these anopheline species.
We collected only one An. balabacensis that returned a positive result for P. knowlesi by PCR. Previous studies also showed that the number of An. balabacensis infected with P. knowlesi is extremely low, for example as low as 0.14% (2/1425)38 and often zero (out of 64139; out of 1,61640). The infected mosquitoes were mostly collected in the early night as reported previously38,39. It is very likely that most An. balabacensis are infected in the forest where the reservoir hosts abound. Host-seeking An. balabacensis are attracted to both humans and macaques16 and this specific trait in host-seeking behavior, coupled with increased potential for interaction between reservoir, vector and human host at forest edges35, may increase the likelihood of human clinical infection with P. knowlesi. Further research is required to establish whether An. donaldi, which we found to be carrying P. knowlesi parasites, plays an active role in P. knowlesi transmission. Other potential vectors in the study area include An. maculatus, a known vector of human malaria in peninsular Malaysia41. We also found specimens of both An. balabacensis and An. donaldi to be positive for P. cynomolgi, a Plasmodium species that was recently recorded in the first naturally-acquired human infection5; An. donaldi has also been shown to be a vector of an unidentified Plasmodium species36. Thus, public health surveillance should be alert to the presence of vectors carrying a range of malaria parasites and the possibility that P. cynomolgi may become a zoonosis causing routine human infection.
The frontline vector control methods practiced in Malaysia under the malaria elimination programme for human-specific malaria, i.e. the application of insecticides in houses either through use of long-lasting insecticide treated nets or indoor residual spraying with K-Othrine, appear to be less effective against P. knowlesi as its confirmed vector bites outdoors where these interventions are not effective. If An. donaldi is a vector, presumably it would continue to be controlled by these standard interventions, although we note in the current study this species was readily caught biting outdoors. Plasmodium knowlesi infection risk therefore appears to be linked to evening exposure to vectors in peri-domestic or agricultural settings40. Given the limited impact of current vector control on zoonotic vectors, control of P. knowlesi in endemic areas requires more information on the vector bionomics within a range of habitats that reflect current land use patterns alongside data on human movement patterns. Data on differences in malaria vector abundance and ecology are critical for understanding the potential to reduce zoonotic transmission both locally within the current P. knowlesi epicentre in Sabah, and other South East Asian regions where this disease is present and other parasitic vector-borne diseases may be emerging. Future research might be directed towards elucidating the potential role of An. donaldi in P. knowlesi transmission and the mechanisms underlying the paradox of sustained transmission in areas where vectors such as An. balabacensis have a low infection rate by P. knowlesi and are relatively small in population size.
Data Availability
All primary data is presented within the manuscript. Data sets can be made available after a written expression of request for data with no apparent competing interest and in compliance with the project’s data sharing agreements.
Change history
20 November 2019
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Lee, K. S. et al. Plasmodium knowlesi: reservoir hosts and tracking the emergence in humans and macaques. PLoS Pathog., 2011. 7.
Lee, K.-S. & Vythilingam, I. Plasmodium knowlesi: Emergent Human Malaria in Southeast Asia, in Parasites and their vectors: A special focus on Southeast Asia, Y. A. L. Lim & Vythilingam, I. Editors, Springer Vienna: Vienna. p. 5–31 (2013).
Barber, B. E. et al. Plasmodium knowlesi Malaria in Children. Emerg Infect Dis 17(5), 814–820 (2011).
William, T. et al. Increasing incidence of Plasmodium knowlesi malaria following control of P. falciparum and P. vivax Malaria in Sabah, Malaysia. PLoS Negl Trop Dis 7(1), e2026 (2013).
Ta, T. H. et al. First case of a naturally acquired human infection with Plasmodium cynomolgi. Malar J., 2014. 13.
Law, Y. H. Rare human outbreak of monkey malaria detected in Malaysia. Nature news. (2018).
Coatney, G. R. Simian malarias in man: facts, implications, and predictions. Am J Trop Med Hyg 17(2), 147–55 (1968).
World Health Organization Malaria Policy Advisory Committee, Outcomes from the evidence review group on Plasmodium knowlesi. (2017).
William, T. et al. Changing epidemiology of malaria in Sabah, Malaysia: increasing incidence of Plasmodium knowlesi. Malar J 13, 390 (2014).
Cooper, D.J. et al. Plasmodium knowlesi Malaria in Sabah, Malaysia, 2015–2017: Ongoing Increase in Incidence Despite Near-elimination of the Human-only Plasmodium Species. Clinical Infectious Diseases, (2019).
Fornace, K. M. et al. Association between Landscape Factors and Spatial Patterns of Plasmodium knowlesi Infections in Sabah, Malaysia. Emerg Infect Dis 22(2), 201–8 (2016).
Ministry of Health Malaysia, Knowlesi Malaria: Malaysia’s Experience in Vector Control (2019).
Yasuoka, J. & Levins, R. Impact of deforestation and agricultural development on anopheline ecology and malaria epidemiology. Am J Trop Med Hyg 76(3), 450–60 (2007).
Imai, N. et al. Transmission and Control of Plasmodium knowlesi: A Mathematical Modelling Study. PLOS Neglected Tropical Diseases 8(7), e2978 (2014).
Pahrol, M., Noraishah, M. S. & Nasir, R. A. Spatial Distribution of Malaria Incidence in Sabah from 2012 to 2016. Geoinformatics & Geostatistics: An Overview, 6 (2018).
Hawkes, F. et al. Evaluation of electric nets as means to sample mosquito vectors host-seeking on humans and primates. Parasit Vectors 10(1), 338 (2017).
Wong, M. L. et al. Seasonal and Spatial Dynamics of the Primary Vector of Plasmodium knowlesi within a Major Transmission Focus in Sabah, Malaysia. PLoS Negl Trop Dis 9(10), e0004135 (2015).
Sallum, M. A. M. et al. Revision of the Leucosphyrus group of Anopheles (Cellia) (Diptera, Culicidae). Rev. Bras. entomol. 49(1), 1–152 (2005).
Rattanarithikul, R. et al. Illustrated keys to the mosquitoes of Thailand. IV. Anopheles. Southeast Asian J Trop Med Public Health 37(Suppl 2), 1–128 (2006).
Phillips, A. J. & Simon, C. Simple, Efficient, and Nondestructive DNA Extraction Protocol for Arthropods. Annals of the Entomological Society of America 88(3), 281–283 (1995).
Singh, B. et al. A genus and species specific nested polymerase chain reaction malaria detection assay for epidemiologic studies. Am J Trop Med Hyg. 60 (1999).
Imwong, M. et al. Spurious amplification of a Plasmodium vivax small-subunit RNA gene by use of primers currently used to detect P. knowlesi. J Clin Microbiol 47(12), 4173–5 (2009).
Helzer, C. J. & Jelinski, D. E. The Relative Importance of Patch Area and Perimeter–Area Ratio To Grassland Breeding Birds. Ecological Applications 9(4), 1448–1458 (1999).
R Core Team and R: A language and environment for statistical computing (2013).
Oksanen, J. et al. vegan: community ecology package. R package version 2, 3–5 (2016).
Rejmánková, E. et al. Ecology of Larval Habitats., in Anopheles mosquitoes-New insights into malaria vectors., S. Manguin, Editor 2013, Rijeka: Intech Open. p. 397–446.
Sánchez-Ribas, J. et al. New classification of natural breeding habitats for Neotropical anophelines in the Yanomami Indian Reserve, Amazon Region, Brazil and a new larval sampling methodology. Memorias Do Instituto Oswaldo Cruz 110(6), 760–770 (2015).
Fitzherbert, E. B. et al. How will oil palm expansion affect biodiversity? Trends in Ecology and Evolution 23, 538–545 (2008).
Prescott, G. W., Edwards, D. P. & Foster, W. A. Retaining biodiversity in intensive farmland: epiphyte removal in oil palm plantations does not affect yield. Ecology and evolution 5(10), 1944–1954 (2015).
Brown, R. et al. Evaluation of resting traps to examine the behaviour and ecology of mosquito vectors in an area of rapidly changing land use in Sabah, Malaysian Borneo. Parasites & Vectors 11(1), 346 (2018).
Chang, M. S. et al. Changes in abundance and behaviour of vector mosquitoes induced by land use during the development of an oil palm plantation in Sarawak. Trans R Soc Trop Med Hyg 91(4), 382–386 (1997).
Hamza, A. M. & Rayah, E. A. E. A Qualitative Evidence of the Breeding Sites of Anopheles arabiensis Patton (Diptera: Culicidae) in and Around Kassala Town, Eastern Sudan. International journal of insect science 8, 65–70 (2016).
Emidi, B. et al. Effect of physicochemical parameters on Anopheles and Culex mosquito larvae abundance in different breeding sites in a rural setting of Muheza, Tanzania. Parasites & Vectors 10(1), 304 (2017).
M’Bra, R. K. et al. Impact of climate variability on the transmission risk of malaria in northern Côte d’Ivoire. PLOS ONE 13(6), e0182304 (2018).
Davidson, G. et al. The Role of Ecological Linkage Mechanisms in Plasmodium knowlesi Transmission and Spread. EcoHealth (2019).
Vythilingam, I. et al. The impact of development and malaria control activities on its vectors in the Kinabatangan area of Sabah, East Malaysia. Acta Trop 96(1), 24–30 (2005).
Brant, H. L. et al. Vertical stratification of adult mosquitoes (Diptera: Culicidae) within a tropical rainforest in Sabah, Malaysia. Malar J., 15 (2016).
Chua, T.H. et al. Phylogenetic analysis of simian Plasmodium spp. infecting Anopheles balabacensis Baisas in Sabah, Malaysia. PLoS Negl Trop Dis, 11(10), p. e0005991 (2017).
Manin, B. O. et al. Investigating the Contribution of Peri-domestic Transmission to Risk of Zoonotic Malaria Infection in Humans. PLoS Negl Trop Dis 10(10), e0005064 (2016).
Ng, S. H. Distribution of Anopheles mosquitoes and Plasmodium knowlesi transmission in Kudat, Sabah Borneo, in Department of Pathobiology & Medical Diagnostics, Universiti Malaysia Sabah, Kota Kinabalu, Sabah, Malaysia: Sabah (2007).
Reid, J. A. Anopheline mosquitoes of Malaya and Borneo., Staples Printers Limited at their Kettering, Northants: Government of Malaysia. 520 (1968).
Acknowledgements
This research was funded by the Malaysian Ministry of Education Fundamental Research Grant Scheme (FRG0388-SKK-2/2014) ‘Ecological determinants of populations of Anopheles species, vectors of knowlesi malaria in Sabah’ and supported by the Royal Society of Tropical Medicine and Hygiene Small Grant Scheme ‘Bionomics of Anopheles balabacensis, the vector of zoonotic malaria in Sabah, Malaysian Borneo’. We thank Stephen Young for statistics advice, Robert Cheke for proof-reading, and study village communities for their participation.
Author information
Authors and Affiliations
Contributions
F.M.H., B.O.M., T.H., S.D., R.H., J.J. and T.H.C. conceived and deisgned the study; B.O.M., T.H.C. and F.M.H. performed experiments; F.M.H., B.O.M., A.C., S.D. and T.H.C. analysed the data; F.M.H., T.H.C., B.O.M. and A.C. wrote the manuscript; all authors read, commented on and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
41598_2019_49842_MOESM1_ESM.pdf
Vector compositions change across forested to deforested ecotones in emerging areas of zoonotic malaria transmission in Malaysia
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hawkes, F.M., Manin, B.O., Cooper, A. et al. Vector compositions change across forested to deforested ecotones in emerging areas of zoonotic malaria transmission in Malaysia. Sci Rep 9, 13312 (2019). https://doi.org/10.1038/s41598-019-49842-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-49842-2
This article is cited by
-
Bayesian spatio-temporal modelling of environmental, climatic, and socio-economic influences on malaria in Central Vietnam
Malaria Journal (2024)
-
The public health response to a Plasmodium malariae outbreak in Penampang district, Sabah during a COVID-19 movement control order
Malaria Journal (2023)
-
Spatial analyses of Plasmodium knowlesi vectors with reference to control interventions in Malaysia
Parasites & Vectors (2023)
-
Natural vectors of Plasmodium knowlesi and other primate, avian and ungulate malaria parasites in Narathiwat Province, Southern Thailand
Scientific Reports (2023)
-
Effect of pyriproxyfen on development and survival of Anopheles gambiae sensu stricto under forested and deforested areas
The Journal of Basic and Applied Zoology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
