
Abstract
The endocardium is the endothelial component of the vertebrate heart and plays a key role in heart development. Where, when, and how the endocardium segregates during embryogenesis have remained largely unknown, however. We now show that Nkx2-5+ cardiac progenitor cells (CPCs) that express the Sry-type HMG box gene Sox17 from embryonic day (E) 7.5 to E8.5 specifically differentiate into the endocardium in mouse embryos. Although Sox17 is not essential or sufficient for endocardium fate, it can bias the fate of CPCs toward the endocardium. On the other hand, Sox17 expression in the endocardium is required for heart development. Deletion of Sox17 specifically in the mesoderm markedly impaired endocardium development with regard to cell proliferation and behavior. The proliferation of cardiomyocytes, ventricular trabeculation, and myocardium thickening were also impaired in a non-cell-autonomous manner in the Sox17 mutant, likely as a consequence of down-regulation of NOTCH signaling. An unknown signal, regulated by Sox17 and required for nurturing of the myocardium, is responsible for the reduction in NOTCH-related genes in the mutant embryos. Our results thus provide insight into differentiation of the endocardium and its role in heart development.
Similar content being viewed by others
Introduction
Heart development is one of the earliest events of vertebrate organogenesis. Cardiac progenitor cells (CPCs) give rise to the myocardium, endocardium, epicardium, smooth muscle, fibroblasts, and endothelium of coronary vessels in the mammalian heart. In the mouse embryo, CPCs originate between embryonic day (E) 6.25 and E7.5 from nascent mesoderm cells in the primitive streak that express the basic helix-loop-helix (bHLH) transcription factor gene Mesp1. These mesoderm cells are a part of the lateral plate mesoderm (LPM) and populate the heart field as CPCs at the most anterior region of the embryo, and they begin to express CPC markers at the early allantoic bud (EB) stage (E7.5). In addition to well-validated CPC marker genes encoding transcription factors—including Nkx2-5, Isl1, and Tbx51,2,3—analysis of alternative markers has provided the basis for a model of the cell lineage tree and the mechanics of cardiomyocyte differentiation4,5. Among the cell types constituting the heart, the development of endocardial cells has remained largely uncharacterized.
The endocardium is the innermost layer and endothelial component of the heart. During heart development, the endocardium provides a source of cells for the valves, the membranous portion of the ventricular septum, the endothelium of coronary vessels, and cardiac fibroblasts. It plays a key role in ventricular trabeculation, myocardial compaction, as well as valve and coronary vessel formation. In the mouse embryo, endocardial cells are first recognized morphologically as a proendocardium layer between the myocardium and definitive endoderm layers at the one- to two-somite stage, when expression of the transcription factor gene Nfatc1, an early and unique marker of the endocardium, is initiated6,7,8.
In mice, Mesp1-expressing nascent mesoderm cells in the primitive streak have been shown to include two types of cell that give rise to the endocardium: unipotent endocardium precursors, and bipotent cardiac progenitors that contribute to cardiomyocytes and the endocardium9. No molecular marker has been identified to distinguish these two cell types from each other, but both likely become Nkx2-5-positive CPCs1,10. Downstream of Mesp1, the ETS-related transcription factor gene Etv2 (also known as Er71 or Etsrp71) plays a role in endocardium development at the top of the endothelial genetic cascade11. Disruption of Etv2 was found to result in loss of the endocardium12,13, suggesting that the endocardium originates from Etv2+ mesoderm. It remains unclear, however, whether Etv2 determines endocardium fate or only confers competence for endocardium differentiation. Downstream of Etv2, a network of transcription factors—including ETS, SOX, GATA, and RBPJκ—regulates endocardium differentiation. These factors likely activate expression of Kdr/Flk1 and Dll4 in the endocardium14,15, with these genes being implicated in vascular endothelial growth factor (VEGF) and NOTCH signaling essential for endocardium development16,17. However, the network of transcription factors responsible for the induction of endocardium fate remains largely unidentified.
Among three Sry-type HMG box F (SOXF) transcription factor genes—Sox7, Sox17, and Sox18—expression of Sox17 was long regarded as specific for the endoderm including the visceral and definitive endoderm18. However, Sox17 was subsequently shown to be essential for vascular development and definitive haematopoiesis19,20. Lineage tracing for Sox17-expressing cells revealed that they contribute not only to the endoderm but also to the mesoderm that gives rise to the endocardium21. Conditional knockout of Sox17 in the endothelial lineage of mice resulted in the loss of vascular rearrangement in the embryo and yolk sac as well as in that of definitive haematopoiesis19,20. With regard to cardiac development, differentiation of mouse embryonic stem cells into cardiomyocytes in vitro was suppressed in a non-cell-autonomous manner by Sox17 ablation, although the identity of Sox17-expressing cells that are required for cardiomyogenesis has been unclear22.
Here we show that Sox17 is expressed in a subset of CPCs during the early phase of mouse cardiac development. Single-cell gene expression profiling revealed that ~20% to 30% of Nkx2-5+ CPCs are positive for Sox17 expression. SOX17+ mesoderm cells were found in the region corresponding to the heart field in E7.5 mouse embryos. This Sox17 expression is transient, persisting up to E8.5 at the latest. Tracing of Sox17+ cells showed that the endocardium originates from Sox17+ mesoderm cells. Gain-of-function and mesoderm-specific loss-of-function analyses for Sox17 in mouse embryos revealed that Sox17 is required for heart development, although it is not essential or sufficient for endocardial fate. Single-cell gene expression profiling for the mesoderm-specific Sox17 mutant showed that disruption of Sox17 resulted in misregulation of the transcriptome in endocardial and myocardial cells in a cell-autonomous and non-cell-autonomous manner, respectively. Our findings thus provide insight into development of the endocardium and its relation to heart morphogenesis.
Results
Sox17 expression in CPCs
We previously performed single-cell gene expression profiling of mouse embryonic CPCs from the EB to early head fold (EHF) stage23,24. We validated cell types based on the expression of marker genes including Sox2 for the epiblast or neural ectoderm, Sox17 for the endoderm or arterial endothelial cells18,25, Cryptic (also known as Cfc1) for LPM, and Nkx2-5 or Tbx5 for CPCs1,3 as described previously23. These data revealed that Sox17 was expressed in 21.9% of CPCs (Cryptic+, Nkx2-5+ and/or Tbx5+) at the EB stage (Fig. 1A). We also confirmed that ~20% to 30% of CPCs continued to express Sox17 up to the early somite stage (Fig. 1A), suggesting that Sox17 plays a role in a subset of CPCs.

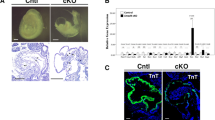
Sox17+ CPCs specific for the endocardium differentiation. (A) Proportion of Sox17-expressing CPCs (Cryptic+, Nkx2-5+ and/or Tbx5+ cells) in mouse embryos at E7.5 (early allantoic bud, late allantoic bud, and early head fold stages) and E8.5 (early somite stage). N values indicate the number of cells examined. The data are derived from our previous study23. (B) Schematic representation of a mouse embryo at the five-somite stage (E8.25) as a left lateral view. The magenta line shows the sectional plane along the anterior (A)-posterior (P) axis in (C). (C–E) Immunofluorescence micrographs for EYFP (green), TNNT (red in C,D) and PECAM1 (red in E). Nuclei (blue) were stained with 4′,6-diamidino-2-phenylindole (DAPI). The boxed region in C is shown at higher magnification in D. The section shown in E is adjacent to that in D. Fg, foregut; En, endoderm. Scale bar, 100 µm.
We next examined the distribution of SOX17-expressing CPCs in mouse embryos by immunofluorescence analysis (Fig. S1). At the EHF stage, SOX17+ CPCs were marked simultaneously by the CPC marker NKX2-5 in the most anterior portion of the embryo corresponding to the heart field (Fig. S1). To distinguish LPM cells from the endoderm, we studied Mesp1Cre/+/Rosa26EYFP/+ mouse embryos, in which Mesp1-expressing mesoderm cells are labeled with enhanced yellow fluorescent protein (EYFP)26,27. Whereas SOX17+ cells were rarely detected among the EYFP+ LPM cells in embryos at the EB and late allantoic bud stages, SOX17+/EYFP+ cells were readily apparent at the EHF stage (Fig. S2). As the endoderm layer thickened and the shape of the foregut pocket became more obvious from the late head fold to zero-somite stage (E8.0), SOX17+ LPM cells became localized more exclusively to a region near the dorsal side of the foregut pocket fold (Fig. S3A). From the three-somite stage (E8.25), the number of SOX17+ LPM cells decreased concomitantly with the decline in expression of SOX17 in the anterior definitive endoderm (Fig. S3B). Only a few SOX17+ LPM cells were apparent at the sinus venosus. These results thus showed that SOX17 is expressed in CPCs during early embryogenesis.
Contribution of Sox17 + CPCs to the endocardium
We next characterized Nkx2-5+/Sox17+ CPCs by polymerase chain reaction (PCR) analysis of marker gene expression with single-cell cDNA. Expression of the endothelial marker genes Dll4 and Pecam1 was highly correlated with that of Sox17 in Nkx2-5+ CPCs at the early somite stage (Fig. S4A), whereas the expression of other marker genes, such as Actn2 (cardiomyocytes) or Acta2 (smooth muscle cells), was not (Fig. S5A)28,29,30,31. These results suggested that Sox17-expressing cells contribute to endothelial-like cells, a conclusion also supported by the preferential expression of Tal1, Kdr, Etv2 and Notch1, marker genes for arterial endothelial precursor cells, in Nkx2-5+/Sox17+ CPCs at the early somite stage (>75% for Tal1, Etv2 and Notch1) as well as in Nkx2-5+ CPCs at the EB stage (100% and 72.7% for Kdr and Etv2, and for Notch1, respectively) (Fig. S4B,C)12,13,32,33. An endocardial marker gene, Nfatc1, also showed a similar expression pattern to Tal1 in Nkx2-5+/Sox17+ CPCs (Fig. S4B). Given that the endothelial component originating from Nkx2-5-expressing CPCs becomes the endocardium, it is likely that Sox17-expressing CPCs give rise to the endocardium34. Of note, other SoxF genes, Sox7 and Sox18, also showed highly correlated with that of Sox17 in Nkx2-5+ CPCs at the early somite stage (>75% for both of Sox7 and Sox18) (Fig. S5B), showing the redundant expression patterns among SoxF genes in the endocardium lineage.
To examine further whether endocardium cells are indeed derived from Sox17-expressing mesoderm cells, we traced the lineage of Sox17-expressing cells in Sox17Sox17-2A-iCre/+/Rosa26EYFP/+ embryos (Fig. 1B–E)21, in which Sox17-expressing cells are labeled with EYFP. We found that the progeny of Sox17-expressing cells were all the PECAM1+ endocardium cells that were enclosed by the troponin T (TNNT)+ myocardium layer at the five-somite stage (E8.5), consistent with the results of a previous study21. Sox17+ CPCs are thus the specific precursors of the endocardium.
Sox17 expression biases CPCs to express an endothelial gene program
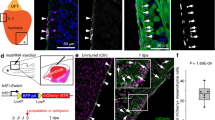
The specificity of Sox17 expression for the endothelial cell lineage suggested that Sox17 plays a role in endocardial differentiation of CPCs. We therefore next examined whether Sox17 expression is sufficient to establish endocardium cell fate in CPCs by forced expression of Sox17 in Nkx2-5-expressing CPCs with the use of a bacterial artificial chromosome (BAC)-based Nkx2-5Sox17-IRES-LacZ transgene (Tg) (Fig. 2A). Expression of the Tg was confirmed to mimic endogenous Nkx2-5 expression in CPCs (Fig. 2B–E). The most severe defect of the Tg+ embryos at E9.5 was anomalous looping and structure of the heart tube (Fig. 2F–J), but no obvious morphological abnormalities were apparent before this stage. Inside the heart of both severely (Fig. 2K–M) and moderately (Fig. S6A,B) affected embryos, ventricular trabeculation was impaired. Apoptosis was frequently observed among MYH (a sarcomeric myosin heavy chain protein)-expressing cardiomyocytes in Tg+ heart tube (Fig. S6C–E), whereas the proliferation of NKX2-5+ cells inside the heart of Tg+ embryos was not significantly affected (Fig. S6F–H). The overall gain-of-function phenotype thus suggested that ectopic and excess Sox17 expression is toxic. Of note, cardiomyocytes expressing PECAM1 were apparent in some Tg+ embryos with a severe phenotype, a phenomenon never observed in wild-type (WT) embryos (Fig. 2K–M), suggesting that Sox17 expression biases the fate of CPCs toward an endothelial-like phenotype but is not sufficient to induce an endocardial cell fate in these cells. This conclusion is consistent with the observation that expression of Etv2 was not detected in the Tg-expressing (LacZ-positive) area of Tg+ embryos (Fig. 2C,E). On the other hand, Tg+ embryos with a mild to moderate phenotype manifested pronounced aggregation of Isolectin B4 (an endothelial marker)-positive endocardium cells rather than a monolayer (Fig. S6B). The intimate interaction between the endocardium and myocardium was also missing. These findings suggested that Sox17 expression in the endocardium must be maintained at an appropriate level for proper regulation of cell behavior.

Gain of function of Sox17 in Nkx2-5-expressing CPCs. (A) Construction of the BAC-based Nkx2-5Sox17-IRES-LacZ Tg. (B–E) Double whole-mount in situ hybridization for Etv2 (blue) and for either Nkx2-5 (brown in B,D) or LacZ (brown in C,E) in E8.25 Tg(−) or Tg(+) embryos, respectively. (F–J) The heart of a Tg(−) embryo (F) and of Tg(+) embryos with severe (G,H), moderate (I), or mild (J) phenotypes at E9.5. Dashed magenta lines show the outline of the heart tube. (K–M) Immunofluorescence micrographs for the sarcomere myosin heavy chain MYH1 (red) and PECAM1 (green) in the left ventricle of the heart of E9.5 Tg(−) (K) and Tg(+) (L) embryos. (M) The boxed region in L is shown at higher magnification. Blue, DAPI; Arrowheads, cardiomyocytes expressing PECAM1; Scale bars, 100 µm.
Cardiac defects induced by Sox17 deletion in mesoderm
To elucidate the physiological function of Sox17 in endocardium development, we conducted conditional depletion of Sox17. Since it cannot be excluded that Sox17 expression in CPCs starts earlier than Nkx2-5, we used the mice with Mesp1Cre/+ and floxed Sox17 (Sox17fl/fl)19 alleles to knockout Sox17 in all cardiac cell lineages. Although multiple anomalies were apparent in Mesp1Cre/+/Sox17fl/fl embryos at E9.5, no obvious morphological abnormalities were detected before this stage. Growth retardation and ballooning of the pericardial sac were observed after approximately the 20-somite stage, suggestive of a severe defect in the peripheral circulation that likely resulted from embryonic heart failure (Fig. 3A,B). Since the defect of the primitive haematopoiesis is possible to cause cardiac malformation, we also confirmed the yolk sac development in the mutants (Fig. S7). The primitive haematopoiesis seemed not affected in the mutant at E9.5, consistent with the previous observation that a haematopoiesis defect was only found after E11.5 in Sox17 mutant19. However, remodeling of the yolk sac vessels was also impaired in the mutant embryos. In WT embryos, the network of capillary vessels that connect to the blood islands, where primitive haematopoiesis gives rise to the production of primary erythrocytes, undergoes remodeling to form the larger vessel network of the yolk sac at E9.5 (Fig. S7). In contrast, such vascular rearrangement did not occur in the mutant. These results thus indicated that Sox17 is essential for vascular development in addition to the heart development.

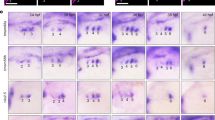
Cardiac defects associated with mesoderm-specific loss of function for Sox17 in mouse embryos. (A,B) Mesp1+/+/Sox17fl/fl (WT) (A) and Mesp1Cre/+/Sox17fl/fl (Sox17 KO) (B) embryos at E9.5. Scale bar, 1 mm. (C,D) Immunofluorescence micrographs in the heart of WT (C) and Sox17 KO (D) embryos at E9.75. The boxed regions in the upper panels are shown at higher magnification in the lower panels. Red, PECAM1; Green, TNNT; Blue, DAPI; LA, left atrium; RA, right atrium; LV, left ventricle; RV, right ventricle. Scale bar, 100 µm. (E–G) Reverse transcription and real-time PCR analysis of the relative expression levels for the NOTCH signaling-related genes Dll4 (E), Notch1 (F), and Nrg1 (G) in the heart of WT and Sox17 KO embryos at E8.5 (left panels) and E9.5 (right panels). Nrg1 expression at E8.5 was below the threshold for detection (N.A., not amplified). Only significant P values (Student’s t test) are indicated. Means ± SD.
Examination of the heart tube revealed anomalous looping in Mesp1Cre/+/Sox17fl/fl embryos at E9.5 (Fig. S8A–F). In WT embryos, the borders of the outflow tract, ventricle, and atrium were identified by folds with a sharp angle. However, the fold angles were obtuse in the mutant. The endothelial-like phenotype of the endocardium was not impaired in the heart of mutant embryos at E9.75, as revealed by the expression of PECAM1 (Fig. 3C,D), indicating that Sox17 is not essential for endocardium fate in CPCs. However, the number of endocardium cells appeared to be reduced in the mutant. Whereas the endocardium was in intimate contact with the myocardium layer in WT embryos, such “touchdown” sites35,36 were far fewer in the mutant (asterisks in Fig. S9A–C). The size of cardiomyocytes was also reduced in the mutant embryos. Importantly, myocardial trabeculation was severely affected and the number of ventricular trabeculae composed of more than three cardiomyocytes was significantly reduced in the mutant (Fig. S9D). Together, these observations indicated that the physiological function of the endocardium was so anomalous in the Sox17 mutant that maturation of the myocardium was affected in a non-cell-autonomous manner.
We next examined the expression levels of NOTCH signaling-related genes in the developing heart, given that a Dll4-Notch1-Nrg1 pathway plays an important role in the differentiating endocardium as well as in ventricular trabeculation37. The expression level of Dll4 did not differ significantly between WT and Mesp1Cre/+/Sox17fl/fl embryos (Fig. 3E). At E8.5, the abundance of Notch1 mRNA (Fig. 3F) and NOTCH1 protein (Fig. S8G,H) was unchanged in the mutant. However, at E9.5, the expression of Notch1 and Nrg1 (a downstream target of the NOTCH signal) in the heart was down-regulated significantly in the mutant embryos (Fig. 3F,G), suggesting that SOX17 does not directly induce NOTCH signaling but instead maintains it in the embryonic heart. Further, the expression levels of Hey1 and Hey2, which regulate the cell proliferation of cardiomyocytes downstream of NOTCH signaling38,39, tended to be reduced in the mutant heart at E9.5 (Fig. S9E,F). The down-regulation of NOTCH signaling apparent at E9.5 is consistent with the previous finding that such signaling is required for trabeculation40.
Transcriptome changes in differentiating cardiac cells induced by Sox17 deletion
The heart anomalies of mesoderm-specific Sox17 mutant embryos were not apparent until E9.5, even though SOX17 is expected to function from E7.5 to E8.5 (Figs 1; S1–S3). We therefore examined whether the loss of function for Sox17 affects the endocardium from the earliest phase of its development, before the morphological abnormality is evident, with the use of single-cell microarray-based expression profiling of endocardium cells at E8.5 (nine-somite stage). Marked differences in gene expression profiles were detected between WT and Mesp1Cre/+/Sox17fl/fl cells. The normalized signal intensities of 114 and 171 probe sets (2.71% of total probe sets called “present”) were increased and decreased, respectively, by a factor of at least 5 in endocardium cells of the mutant relative to those of WT embryos (Fig. 4A; Table S1; SI, Gene Lists 1, 2). The gene expression profile of cardiomyocytes of E8.5 mutant embryos also differed from that of control cardiomyocytes, with the normalized signal intensities of 84 and 274 probe sets (4.78% of total probe sets called “present”) being increased and decreased, respectively (Fig. 4B; Table S1; SI, Gene Lists 3, 4), supporting the notion that Sox17 expression in the endocardium contributes to regulation of the genetic program for cardiomyocyte differentiation from the early stage of heart development. Of note, mitochondria related genes (Acsl4, Mcat, Micu2, Mrpl4, Mrpl9, Mrpl4, Mrpl13, Mrpl30, Mrpl36, Mrpl38, Mrps24, Ndufa4, Ndufa10, Tomm34, Oxa1l, Slc25a13), sarcomere-related genes (Hrc, Speg, Tpm4) and cardiac development-related genes (Cited2, Gata4, Irx4, Ldb3, Pfkm, Tln1) were significantly down-regulated in the mutant cardiomyocytes, suggesting that cardiomyocyte maturation was impaired (Gene List 4).

Altered gene expression profiles in the endocardium and myocardial cells of mesoderm-specific Sox17 mutant embryos. (A,B) Hierarchical clustering and heat map for 285 (A) and 358 (B) probe sets for differentially expressed genes in the endocardium and myocardial cells, respectively, of WT and Sox17 KO embryos at E8.5. (C,D) Immunofluorescence micrographs for pHH3 (green) and PECAM1 (red) in the heart of WT (C) and KO (D) embryos at E9.5. Blue is DAPI. Arrowheads indicate pHH3+ endocardium cells. LV, left ventricle; Scale bars, 50 µm. (E) Proportion of pHH3+ cells among PECAM1+ cells in the heart of WT and Sox17 KO embryos from E8.5 to E9.5. Means ± SD. Only P values of ≤0.05 (Student’s t test) are indicated.
Pathway analysis for the lists of genes whose expression was affected in mutant embryos revealed that molecules related to “cellular growth and proliferation,” “cellular development,” and “cell cycle” were significantly enriched for both endocardium cells and cardiomyocytes (Tables S2, S3). Examination of cell proliferation in the heart from E8.5 to E9.5 revealed that the ratio of KI67+ (mitotic) endocardium cells or cardiomyocytes did not differ significantly between WT and mutant embryos at E8.5 or E9.0 but was significantly reduced in the mutant at E9.5 (Fig. S10A,B). Similarly, the ratio of phosphorylated histone H3 (pHH3)-positive (M-phase) cells in the endocardium of mutant embryos was significantly reduced at E9.0 (Fig. 4C–E). A significant reduction in the ratio of cardiomyocytes in M phase was apparent in the mutant at E9.5 (Fig. S10C–E), consistent with the notion that an initial anomalous molecular event in the endocardium subsequently affects the myocardium. Given that a reduction in Notch1 expression in the mutant heart was not apparent until E9.5 (Fig. 3F), a primary event caused by the lack of Sox17 expression in the endocardium might secondarily induce down-regulation of Notch1 expression and thereby affect cell proliferation and trabeculation in the myocardium. Together, these results suggest that Sox17 expression in the endocardium precursor cells is essential for regulation of endocardium development in a cell-autonomous manner and for that of myocardium development in a non-cell-autonomous manner.
Discussion
We have here shown that Sox17 is transiently expressed from E7.5 to E8.5 specifically in CPCs undergoing differentiation into the endocardium in mouse embryos. Gain-of-function and loss-of-function analyses revealed that Sox17 is neither necessary nor sufficient for the induction of endocardial cell fate in CPCs. However, its expression in endocardium precursor cells is required for proper heart development and biases CPCs toward an endothelium-like phenotype. Our results suggest that SOX17 renders endocardium cells competent for proliferation and interaction with cardiomyocytes, with such interaction regulating cardiomyocyte growth and maturation (Fig. S10F).
Much of the available information relevant to endocardium differentiation has been derived from studies of the development of haematopoietic stem cells and the arterial endothelium. In mice, Etv2 is essential for establishment of the endothelial and haematopoietic cell lineages in Mesp1+ mesoderm, serving as a downstream target of bone morphogenetic protein (BMP), NOTCH, and WNT signaling pathways20. Null mutation of Etv2 results in embryonic mortality and loss of endothelial cells including the endocardium as well as of blood cells12,13. In addition, Notch1, which plays an important role in development of haemogenic arterial endothelial cells, is also required for development of the endocardium41,42,43. Mesodermal cells expressing Notch1 at E6.5 were found to contribute mostly to the endocardium44. Given that Sox17 appears to function in the haematopoietic lineage and that the endocardium likely possesses haemogenic ability19,45, the endocardium may share a common or similar developmental gene program including Sox17 with the haematopoietic lineage. This notion is further supported by the observation that zebrafish mutants of cloche, which encodes a bHLH-PAS-type transcription factor and is expressed in endothelial and haematopoietic precursors, do not develop an endocardium or haematopoietic stem cells46,47. Of interest, NKX2-5 was shown to transactivate Etv2 in Nkx2-5+ cells that contribute to the endocardium, suggesting that the cardiac program functions upstream of the endothelial-haemogenic progenitor gene program essential for endocardium development48. However, the endothelial-haemogenic program including Etv2 and Tal1 prevents the induction of cardiomyocyte fate in CPCs48,49. Endocardium fate determination might thus be initiated by the cardiac program coupled to the endothelial-haemogenic program among Mesp1-expressing nascent mesoderm cells, with the endothelial-haemogenic program subsequently suppressing the cardiomyocyte program after commitment to the endocardium. Validation of the mechanism underlying fate determination for the endocardium will require further studies of the roles of Mesp1, Sox17, Etv2, Notch1, and other early endothelial-haemogenic factor genes.
Sox17 has been shown to be required for vascular development. Its expression was thus found to promote sprouting angiogenesis during retinal vascularization and tumor angiogenesis20,50. Loss or gain of function of Sox17 in the endothelial lineage of mouse embryos resulted in decreased and increased vascular density, respectively51. SOX17 was suggested to destabilize vascular endothelial cells and thereby to secure the motility of tip cells during angiogenesis by regulating the proliferation, adhesion, and cytoskeletal organization of endothelial cells as well as the extracellular matrix. The phenotypes resulting from loss or gain of function of Sox17 in our study are likely consistent with such a role, given the observed defects in cell shape, cell proliferation, and cell-cell interaction (Figs 2–4, Figs S6–10). Endocardial cells that sprout toward the myocardium via an angiogenesis-like process may thus generate the touchdown sites at which cardiomyocytes are able to initiate their trabeculation35,36. For the endocardium to exert such an action, Sox17 must be expressed at an appropriate level, given the phenotypes associated with its gain or loss of function (Figs 2, 3). On the other hand, a specific role for Sox17 in heart development is difficult to discern because of the functional redundancy of SOXF transcription factor genes. Sox7 is expressed in a subset of the Mesp1+ cell lineage from the earliest phase44, and Sox7 mutant embryos manifest an overall phenotype similar to that of embryos lacking Sox1752. Identification of the direct targets of SOX17 and the other SOXF transcription factors should provide insight into the roles of these proteins in development.
Activated DLL4-NOTCH1 signaling in the endocardium was shown to be essential for ventricular trabeculation, with the consequent up-regulation of Ephrin B2-EphB4-Nrg1 signaling promoting the differentiation of cardiomyocytes36,37. We found that loss of function of Sox17 resulted in defective trabeculation, likely as a result of the significant down-regulation of Notch1 and Nrg1 expression in the heart apparent at E9.5. These findings appear consistent with the previous observation that SOX17 directly activates Notch1 to establish the haemogenic endothelium42. However, expression of Notch1 was not affected in our Sox17 loss-of-function mutant at E8.5, indicating that Sox17 is dispensable for the induction of Notch1 expression at the earliest phase of endocardium development, in contrast to the situation for haematopoietic stem cell development. The reduction in the level of Notch1 expression in our Sox17 mutant at E9.5 might be a secondary impairment due to a primary defect in the endocardium, or Sox17 may be required for maintenance of Notch1 expression. Alternatively, rather than SOX17, one or both of the other SOXF transcription factors (SOX7, SOX18) might directly activate Notch1 in the endocardium lineage. Further studies of the relative roles of SOX17 and the other SOXF proteins should provide insight into the development of the endocardium.
Methods
Mice
Sox17fl (MGI ID: 3717121)19 and Rosa26EYFP Cre reporter (MGI ID: 2449038)27 mice were obtained from The Jackson Laboratory. Mesp1Cre mice (MGI ID: 2176467) were described previously26. All animal procedures were performed under project licenses (70/7254 and 70/7449) approved by the Home Office according to the Animals (Scientific Procedures) Act 1986 in the U.K., or with approval of the Osaka University Animal Experimentation Committee (license number: 29-039-004) in Japan.
Staging of mouse embryos
Developmental stages of mouse embryos were classified according to morphology as previously described23. The morning of the day of vaginal plug detection was set as E0.5.
BAC transgenesis
The construction of BAC transgene and transgenic mice production were performed as previously described23,53. The BAC Nkx2-5Sox17-IRES-LacZ-BghpA Tg was constructed with a BAC recombination system as shown in Fig. 2A. The BAC clone RP24-324A10 contains 175.0 kb of the mouse Nkx2-5 locus and drives expression of the Sox17-IRES-LacZ cassette (containing full-length mouse Sox17 cDNA) according to the genomic context of Nkx2-5. A Sox17-IRES-LacZ-BghpA cassette was introduced in-frame into the mouse Nkx2-5 gene of BAC clone RP24-324A10. For recombination with the BAC, left-arm XbaI-XhoI and right-arm EcoRI-EcoRV fragments were amplified by PCR independently with the primer sets Nkx2-5-L-arm-F (5′-XbaI site-GTCGACCGTTTAGACTCAGCATAACAG-3′) and Nkx2-5-L-arm-R (5′-XhoI site-CAGGTTTCACAGCGCCAGGTG-3′) as well as Nkx2-5-R-arm-F (5′-EcoRI site-GATAAAAAAGGTAAGGAGAAC-3′) and Nkx2-5-R-arm-R (5′-EcoRV site-GGCAGGGTGGGCTACACAAGG-3′), respectively. The right-arm EcoRI-EcoRV fragment was cloned into pL453 as Frt-Neor-Frt-R arm, and the left-arm XbaI-XhoI and IRES-LacZ XhoI-BamHI fragments were then simultaneously introduced to yield L arm-IRES-LacZ-Frt-Neor-Frt-R arm. A XhoI-BglII fragment of mouse Sox17 cDNA obtained by PCR with the primers 5′-XhoI site-GTCGCCACCATGAGCAGCCCGGATGCGGGA-3′ and 5′-TCTGCGTTGTGCAGATCTGGG-3′ and a BglII-XbaI fragment of the Sox17 cDNA were simultaneously cloned into the plasmid to yield L arm-Sox17-IRES-LacZ-Frt-Neor-FRT-R arm. EL250 cells transformed with the RP24-324A10 BAC clone were subjected to electroporation with the L arm-Sox17-IRES-LacZ-Frt-Neor-FRT-R arm fragment and then to selection with kanamycin. Following removal of the Neor cassette by arabinose treatment (Flp induction), the BAC Tg was prepared and used for microinjection.
RNA isolation, RT, and real-time PCR analysis
Embryonic hearts were dissected in ice-cold phosphate-buffered saline (PBS). Total RNA was isolated with the use of RNeasy Mini Kit (Qiagen) and was subjected to reverse transcription (RT) with the use of Superscript III (ThermoFisher Scientific) and an oligo(dT) primer. The resulting cDNA was subjected to real-time PCR analysis with specific primer sets (Table S4) and with the use of a QuantiTect SYBR Green PCR Kit and Rotorgene (Qiagen). Data were normalized by Gapdh expression.
Histology
Mouse embryos were dissected in ice-cold PBS and fixed with 4% paraformaldehyde in PBS at 4 °C for 2 h. For assay of BrdU incorporation, pregnant mice were injected intraperitoneally with 500 µg of BrdU (Sigma-Aldrich) at 3 h before embryo removal. Fixed embryos were embedded in OCT compound (Sakura Finetek), frozen, sectioned at a thickness of 8 µm, and stained with primary and secondary antibodies (Table S5) as previously described5,23. DNA was counterstained with DAPI (Merck). Immunofluorescence micrographs were acquired with an LSM510 confocal (Zeiss) or BZ8000 fluorescence (Keyence) microscope. Whole mount in situ hybridization was performed as described5,23.
Single-cell microarray analysis
The dissected hearts were treated with trypsin for 3 min at 37 °C to isolate single cells. First-strand cDNA was synthesized for 10 min at 50 °C with Superscript III reverse transcriptase (ThermoFisher Scientific). The cell type for each single-cell cDNA preparation was identified by PCR of marker gene expression with specific primer sets (Table S4). The T3 promoter was added to the 5′ end of each cDNA by PCR with the T3V3 primer (5′-CCAAGCTCGAAATTAACCCTCACTAAAGGGAGAATATCTCGAGGGCGCGCCGGATCC-3′) and V1dT24 primer (5′-ATATGGATCCGGCGCGCCGTCGACTTTTTTTTTTTTTTTTTTTTTTTT-3′). Cells were isolated from the heart ventricle of control (Sox17fl/fl) or Sox17 knockout (Mesp1Cre/+/Sox1fl/fl) embryos at E8.5, and single-cell cDNA was synthesized by RT as previously described with some modifications23. The T3 promoter was added to the 5′ end of each cDNA by PCR. Virtual mRNAs were synthesized from the T3 promoter-cDNA constructs with a MEGAScript T3 Transcription Kit (ThermoFisher Scientific), and they (50 ng) were then labeled with Cy3 with the use of a Low RNA Input QuickAmp (One Color) Labeling Kit before hybridization with a SurePrint G3 Mouse GE v2 8 × 60 K Microarray with the use of a Gene Expression Hybridization Kit (Agilent). The processed data were analyzed statistically with Genespring GX software (Agilent). Expression levels of <50 were set to 50, per chip normalization was based on the 50th percentile, and per gene normalization was based on the median. Gene subtraction was performed as shown in Table S1. Gene lists were analyzed with Ingenuity Pathway Analysis software (Qiagen Bioinformatics). The microarray data obtained for this study are deposited in the Gene Expression Omnibus database (GEO, http://www.ncbi.nlm.nih.gov/geo) under the accession number GSE125323.
References
Lints, T. J. et al. Nkx-2.5: a novel murine homeobox gene expressed in early heart progenitor cells and their myogenic descendants. Development 431, 419–431 (1993).
Cai, C. et al. Isl1 Identifies a Cardiac Progenitor Population that Proliferates Prior to Differentiation and Contributes a Majority of Cells to the Heart. Dev. Cell 5, 877–889 (2003).
Bruneau, B. G. et al. A Murine Model of Holt-Oram Syndrome Defines Roles of the T-Box Transcription Factor Tbx5 in Cardiogenesis and Disease. Cell 106, 709–721 (2001).
Liang, X. et al. HCN4 Dynamically Marks the First Heart Field and Conduction System Precursors. Circ. Res. 113, 399–407 (2013).
Ishida, H. et al. GFRA2 Identifies Cardiac Progenitors and Mediates Cardiomyocyte Differentiation in a RET-Independent Signaling Pathway Article GFRA2 Identifies Cardiac Progenitors and Mediates Cardiomyocyte Differentiation in a RET-Independent Signaling Pathway. Cell Rep. 16, 1026–1038 (2016).
Kaufman, M. & Navaratnam, V. Early differentiation of the heart in mouse embryos. J. Anat. 133, 235–246 (1981).
Deruiter, M. C. et al. The development of the myocardium and endocardium in mouse embryos Fusion of two heart tubes? Anat. Embryol. (Berl). 185, 461–473 (1992).
Misfeldt, A. M. et al. Endocardial cells are a distinct endothelial lineage derived from Flk1 + multipotent cardiovascular progenitors. Dev. Biol. 333, 78–89 (2009).
Lescroart, F. et al. Early lineage restriction in temporally distinct populations of Mesp1 progenitors during mammalian heart development. Nat. Cell Biol. 16, 829–840 (2014).
Zamir, L. et al. Nkx2. 5 marks angioblasts that contribute to hemogenic endothelium of the endocardium and dorsal aorta. eLlFE 6, 1–31 (2017).
Bondue, A. et al. Mesp1 acts as a master regulator of multipotent cardiovascular progenitor specification. Cell Stem Cell 3, 69–84 (2008).
Lee, D. et al. Article ER71 Acts Downstream of BMP, Notch, and Wnt Signaling in Blood and Vessel Progenitor Specification. Cell Stem Cell 2, 497–507 (2008).
Ferdous, A. et al. Nkx2-5 transactivates the ETS-re;ated protein 71 gene and Specifies an Endothelial/Endocardial Fate in the Developing Embryo. Proc. Natl. Acad. Sci. 106, 814–819 (2009).
Becker, P. W. et al. An intronic Flk1 enhancer directs arterial-specific expression via RBPJ-mediated venous repression. Arterioscler. Thromb. Vasc. Biol. 36, 1209–1219 (2016).
Wythe, J. D. et al. ETS factors regulate Vegf-dependent arterial specification. Dev. Cell 26, 45–58 (2013).
Shalaby, F. et al. Failure of blood-island formation and vasculogenesis in Dlk-1-deficient mice. Nature 376, 62–66 (1995).
Gale, N. W. et al. Haploinsufficiency of delta-like 4 ligand results in embryonic lethality due to majour deects in arterial and vascular development. Proc. Natl. Acad. Sci. 101, 15949–15954 (2004).
Kanai-azuma, M. et al. Depletion of definitive gut endoderm in Sox17-null mutant mice. Development 129, 2367–2379 (2002).
Kim, I., Saunders, T. L. & Morrison, S. J. Sox17 Dependence Distinguishes the Transcriptional Regulation of Fetal from Adult Hematopoietic Stem Cells. Cell 130, 470–483 (2007).
Yang, H. et al. Sox17 promotes tumor angiogenesis and destabilizes tumor vessels in mice. J. Clin. Invest. 123, 418–431 (2013).
Engert, S., Liao, W. P., Burtscher, I. & Lickert, H. Sox17-2A-iCre: A knock-in mouse line expressing Cre recombinase in endoderm and vascular endothelial cells. Genesis 47, 603–610 (2009).
Liu, Y. et al. Sox17 is essential for the specification of cardiac mesoderm in embryonic stem cells. Proc. Natl. Acad. Sci. 104, 3859–3864 (2007).
Kokkinopoulos, I. et al. Single-cell expression profiling reveals a dynamic state of cardiac precursor cells in the early mouse embryo. PLoS One 10, 1–25 (2015).
Downs, K. M. & Davies, T. Staging of gastrulating mouse embryos by morphological landmarks in the dissecting microscope. Development 118, 1255–66 (1993).
Liao, W. P. et al. Generation of a mouse line expressing Sox17-driven Cre recombinase with specific activity in arteries. Genesis 47, 476–483 (2009).
Saga, Y. et al. MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tube. Development 126, 3437–47 (1999).
Srinivas, S. et al. Cre reporter strains produced by targeted insertion of. BMC Dev. Biol. (2001).
Rao, P. K. et al. Isolation and Characterization of the Notch Ligand Delta4. Exper imenta l Cell Res. 260, 379–386 (2000).
Baldwin, H. S. et al. Platelet endothelial cell adhesion molecule-1 (PECAM-1/CD31): alternatively spliced, functionally distinct isoforms expressed during mammalian cardiovascular development. Development 120, 2539–53 (1994).
Perou, C. M. et al. Comparative mapping in the beige-satin region of mouse chromosome 13. Genomics 39, 136–146 (1997).
Min, B. et al. Nucleotide sequence of a mouse vascular smooth muscle alpha-actin cDNA screened for vascular smooth muscle (VSM) alpha-actin cDNA probes for use in studies of smooth muscle myogenesis. Nucleic Acids Res. 16, 10374 (1988).
Drake, C. J. & Fleming, P. A. Vasculogenesis in the day 6.5 to 9.5 mouse embryo. 95, 1671–1679 (2000).
Weinmaster, G., Roberts, V. J. & Lemke, G. A homolog of Drosophila Notch expressed during mammalian development. Development 205, 199–205 (1991).
Stanley, E. G. et al. Efficient cre-mediated deletion in cardiac progenitor cells con- ferred by a 3′UTR-ires-Cre allele of the homeobox gene Nkx2-5. Int. J. Dev. Biol. 46, 431–439 (2002).
Jiménez-Amilburu, V. et al. In Vivo Visualization of Cardiomyocyte Apicobasal Polarity Reveals Epithelial to Mesenchymal-like Transition during Cardiac Trabeculation. Cell Rep. 17, 2687–2699 (2016).
del Monte-nieto, G. et al. Control of cardiac jelly dyamics by NOTCH1 and NRG1 defines the building plan for trabeculation. Nature, 557, 439-445 (2018).
Grego-Bessa, J. et al. Notch Signaling Is Essential for Ventricular Chamber Development. Dev. Cell 12, 415–429 (2007).
Fischer, A. et al. The Notch target genes Hey1 and Hey2 are required for embryonic vascular development. 901–911 (2004).
Kokubo, H. et al. Mouse hesr1 and hesr2 genes are redundantly required to mediate Notch signaling in the developing cardiovascular system. 278, 301–309 (2005).
De la Pompa, J. L. & Epstein, J. A. Coordinating Tissue Interactions: Notch Signaling in Cardiac Development and Disease. Dev. Cell 22, 244–254 (2012).
Marcelo, K. L. et al. Hemogenic endothelial cell specification requires c-Kit, notch signaling, and p27-mediated cell-cycle control. Dev. Cell 27, 504–515 (2013).
Clarke, R. L. et al. The expression of Sox17 identifies and regulates haemogenic endothelium. Nat. Cell Biol. 15, 502–510 (2013).
Richard, C. et al. Endothelio-Mesenchymal Interaction Controls runx1 Expression and Modulates the notch Pathway to Initiate Aortic Hematopoiesis. Dev. Cell 24, 600–611 (2013).
Lescroart, F. et al. Defining the earliest step of cardiovascular lineage segregation by single-cell RNA-seq. Science (80-.). 359, 1177–1181 (2018).
Nakano, H. et al. Haemogenic endocardium contributes to transient definitive haematopoiesis. Nat. Commun. 4, 1–10 (2013).
Stainier, D. Y. R. et al. cloche, an early acting zebrafish gene, is required by both the endothelial and hematopoietic lineages. Development 121, 3141–3150 (1995).
Reischauer, S. et al. Cloche is a bHLH-PAS transcription factor that drives haemato-vascular specification. Nature 535, 294–298 (2016).
Rasmussen, T. L. et al. ER71 directs mesodermal fate decisions during embryogenesis. Development 138, 4801–4812 (2011).
Van Handel, B. et al. Scl Represses Cardiomyogenesis in Prospective Hemogenic Endothelium and Endocardium. Cell 150, 590–605 (2012).
Corada, M. et al. Sox17 is indispensable for acquisition and maintenance of arterial identity. Nat. Commun. 4, 1–14 (2013).
Lee, S. H. et al. Notch pathway targets proangiogenic regulator Sox17 to restrict angiogenesis. Circ. Res. 115, 215–226 (2014).
Wat, M. J. et al. Mouse model reveals the role of SOX7 in the development of congenital diaphragmatic hernia associated with recurrent deletions of 8p23.1. Hum. Mol. Genet. 21, 4115–4125 (2012).
Uehara, M. et al. Removal of maternal retinoic acid by embryonic CYP26 is required for correct Nodal expression during early embryonic patterning. Genes Dev. 23, 1689–1698 (2009).
Acknowledgements
This work was supported by a U.K. Medical Research Council New Investigator Research Grant (G0900105), a British Heart Foundation Project Grant (PG/11/102/29213) and a Japan Society for the Promotion of Science Grants-in-Aid for Scientific Research (B) (17H04228) to K.Y.; by a Japan Society for the Promotion of Science Grants-in-Aid for Scientific Research (C) (17K08490) to R.S. We thank Nicholas Greene and Mitsuru Morimoto for technical advice as well as Brigid Hogan, Stefan Hoppler, Pete Scambler, and Paul Riley for helpful discussion.
Author information
Authors and Affiliations
Contributions
K.Y. designed research; K.Y., R.S., K.K., L.R., S.E., M.U., H.I., I.K., Y. Shintani, C.M., J.K., H.L. and K.S. performed research; K.Y., R.S., S.M., Y.K., M.K.-A., P.K., Y. Saga, and Y. Sawa analyzed data.; and K.Y. and R.S. wrote the paper.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Saba, R., Kitajima, K., Rainbow, L. et al. Endocardium differentiation through Sox17 expression in endocardium precursor cells regulates heart development in mice. Sci Rep 9, 11953 (2019). https://doi.org/10.1038/s41598-019-48321-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-48321-y
This article is cited by
-
A SOX17-PDGFB signaling axis regulates aortic root development
Nature Communications (2022)
-
Setd4 controlled quiescent c-Kit+ cells contribute to cardiac neovascularization of capillaries beyond activation
Scientific Reports (2021)
-
In vitro generation of functional murine heart organoids via FGF4 and extracellular matrix
Nature Communications (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
