
Abstract
The early evolution of lepidosaurs is marked by an extremely scarce fossil record during the Triassic. Importantly, most Triassic lepidosaur specimens are represented by disarticulated individuals from high energy accretion deposits in Laurasia, thus greatly hampering our understanding of the initial stages of lepidosaur evolution. Here, we describe the fragmentary remains of an associated skull and mandible of Clevosaurus hadroprodon sp. nov., a new taxon of sphenodontian lepidosaur from the Late Triassic (Carnian; 237–228 Mya) of Brazil. Referral to Sphenodontia is supported by the combined presence of a marginal dentition ankylosed to the apex of the dentary, maxilla, and premaxilla; the presence of ‘secondary bone’ at the bases of the marginal dentition; and a ventrally directed mental process at the symphysis of the dentary. Our phylogenetic analyses recover Clevosaurus hadroprodon as a clevosaurid, either in a polytomy with the Late Triassic to Early Jurassic Clevosaurus and Brachyrhinodon (under Bayesian inference), or nested among different species of Clevosaurus (under maximum parsimony). Clevosaurus hadroprodon represents the oldest known sphenodontian from Gondwana, and its clevosaurid relationships indicates that these sphenodontians achieved a widespread biogeographic distribution much earlier than previously thought.
Similar content being viewed by others
Introduction
The Triassic period marks one of the great transitions in the history of life after the mass-extinction at the end of the Permian. Most notably, this was a period of diversification of several major groups of extant terrestrial and marine vertebrates1, including lepidosaurian (Squamata + Rhynchocephalia) reptiles2. Today, Lepidosauria is the largest and the most diverse group of non-avian reptiles with over 10,000 known species3. Most of these are lizards and snakes with rhynchocephalians limited to two recognized sphenodontian species (Sphenodon punctatus and S. guntheri) in New Zealand4. However, during the early Mesozoic, rhynchocephalians (particularly sphenodontians), were the most diverse and widely distributed members of Lepidosauria5.
In the last decade, the discovery of several important fossils has greatly increased our knowledge of the anatomy, systematics, distribution, and early evolution of sphenodontians. These new findings include presumably stem rhynchocephalians of the Late Triassic, such as Gephyrosaurus evansae6 and Penegephyrosaurus curtiscoppi7 from the United Kingdom, and Deltadectes elvetica7 from Switzerland. These three taxa, though not being well-understood phylogenetically, have been tentatively positioned as stem rhynchocephalians, sharing morphological features with Gephyrosaurus and closely related taxa6,7.
By far, the most well-known early sphenodontian clade is Clevosauridae, which currently includes several species allocated to three genera: Brachyrhinodon, Clevosaurus, and Polysphenodon from the Late Triassic to Early Jurassic of Britain8,9,10,11,12, Germany9,13,14, Africa15,16, China17, North America18, and South America19. Although support for this clade has not been universal9,20,21,22, the genus Clevosaurus is currently considered the most specious (six species) and widely distributed (Asia, Europe, North America, South America, Africa) of the clevosaurid taxa17,18,23,24,25.
The new sphenodontian taxon described herein is the oldest unequivocal record of Rhynchocephalia (Sphenodontia) from South America, as well as in Gondwana. It provides the first evidence of a greater diversity of early sphenodontians than previously understood for southern Pangaea and provides new information on the earliest stages of the acquisition of the acrodont dentition in sphenodontians in combination with unique dental specializations.
Results
Systematic Palaeontology
Lepidosauria Dumeril and Bibron, 1839 sensu Evans, 1984
Rhynchocephalia Günther, 1867 sensu Gauthier et al., 1988
Sphenodontia Williston, 1925 sensu Benton, 1985
Referral to Sphenodontia is supported by the combined presence of a marginal dentition ankylosed to the apex of the dentary, maxilla, and premaxilla; the presence of “secondary bone” (sensu Harrison26,27, Fraser28 and Jones22) at the bases of the marginal dentition; a ventrally directed mental process (sensu Apesteguía29,30) at the symphysis of the dentary; presence of a very shallow Meckelian canal.
Clevosauridae Bonaparte and Sues, 2006 (sensu this study)
New definition
Clevosauridae was originally defined by Bonaparte and Sues19 as the least inclusive clade containing the last common ancestor shared between Brachyrhinodon, Polysphenodon, and Clevosaurus plus all of its descendants. Due to the uncertain position of Polysphenodon (i.e. if the genus is or is not closely related to the sister group Brachyrhinodon + Clevosaurus) in other topologies9,20,21,25, here we adopt the interpretation of Clevosauridae as a monophyletic group using the node–based definition of the clade containing the last common ancestor shared between Brachyrhinodon and Clevosaurus plus all of its descendants.
Clevosaurus hadroprodon sp. nov.
ZooBank
urn:lsid:zoobank.org:act:971AD911-D5FB-460B-91AF-25B3D973873E.
Etymology
Species epithet comes from a combination of ‘hadroprodon’ (from the Greek “hadros”-meaning large) and “protos”, meaning first (Greek), and “odous”, meaning tooth (Greek); in reference to the “big first tooth”, i.e. the large tusk-like tooth of the premaxilla and the anteriormost/first dentary tooth position.
Holotype
MMACR PV-027-T, an incomplete right premaxilla, an incomplete right maxilla and right lower jaw in occlusion, and left lower jaw located below these elements.
Referred material
MMACR PV-028-T, an incomplete left lower jaw; and MMACR PV-029-T, fragment of dentary with three isolated teeth.
Locality and horizon
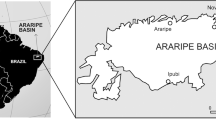
Linha Bernardino locality, Candelária municipality, Rio Grande do Sul State, Southern Brazil (Fig. 1); Santa Maria Formation (Santa Maria Supersequence, Candelária Sequence), Rosário do Sul Group, Paraná Basin; Carnian, Late Triassic31,32 (Fig. 1a–c). Clevosaurus hadroprodon was recovered from immediately beneath the layers that contained the cranial and postcranial remains of the cynodonts Exaeretodon sp. and Trucidocynodon sp. and a distal portion of femur that closely resembles the early dinosaur cf. Pampadromaeus (Fig. 1b). The presence of these fossils referred to Exaeretodon, Trucidocynodon and a Pampadromaeus-like form places this locality within the Hyperodapedon AZ at the base of the Candelária Sequence. Recent high-precision U-Pb zircon geochronology data recovered a weighted mean 206Pb/238U date of 233.23 ± 0.73 Ma for typical Hyperodapedon AZ sites33, which can be biostratigraphically correlated to the sedimentary layers containing C. hadroprodon. As such we conservatively consider the age for the fossil material to be Carnian (Late Triassic).

Location and stratigraphy interpretation. Geological map (a) of Paraná Basin in South America (upper left) and showing the Santa Maria Supersequence in Rio Grande do Sul state (southern Brazil) (detail). The Sanga do Cabral and Santa Maria Supersequences represent the geographical extent of Triassic strata. The star marks the location of the municipality of Candelária where the specimen here described comes from (Modified from Horn et al.30). Summarized stratigraphic scheme of the Linha Bernardino outcrop (b), type locality of Clevosaurus hadroprodon sp. nov. Panoramic view of the Linha Bernardino type locality (c).
Diagnosis
Small sphenodontian rhynchocephalian differing from all other known sphenodontians in possessing the following combination of features: (1) a large, blunt, tusk-like tooth in both premaxilla and first tooth position of dentary; (2) in having an angled, but nearly vertical mandibular symphysis and relatively deep dentary; (3) absence of an edentulous gap between the tusk-like tooth and the remaining dentition; (4) absence of a posterodorsal process of the premaxilla; (5) in lacking well-developed medial-posteromedial expansion of the posterior dentition; (6) and lacking flanges on the teeth, and (7) lacking labially expanded teeth. Feature 4 makes this new taxon different from many sphenodontians, including other species of Clevosaurus, Rebbanasaurus, Godavariasaurus, Priosphenodon, Sphenotitan, Paminzisaurus and Ankylosphenodon. Features 5 and 6 make this taxon different from all other described species of Clevosaurus [Clevosaurus hudsoni, C. bairdi, C. latidens, C. sectumsemper, C. cambrica, and Clevosaurus sp. (China)]
Nomenclatural Acts
The electronic version of this article as a PDF represents published work according to the International Commission on Zoological Nomenclature (ICZN), and thus the new name contained in the electronic version is regarded as published. This published work and the Nomenclatural Acts it contains have been registered in ZooBank with the LSID urn:lsid:zoobank.org:pub:02B4B0AF-B553-410B-94B1-76D7AE277FAE for this publication and urn:lsid:zoobank.org:act:971AD911-D5FB-460B-91AF-25B3D973873E for the species. The online version of this work is archived and available from the digital repositories PubMed Central and CLOCKSS.
Description and taxonomic comparisons
Premaxilla
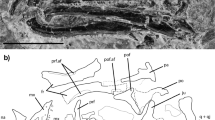
The right premaxilla of the holotype MMACR PV-027-T (Fig. 2a,b) appears to be complete, but is not articulated with the maxilla and is exposed in lingual/posterior view. It bears a single, massive, tusk-like tooth and has a narrow, long (almost 3 mm), mediolaterally compressed, straight ascending nasal process with a slight (approximately 20°) medial deflection with respect to the long axis of the tooth. Because the anterior portion is embedded in the matrix the exact anteroposterior depth of this process is unknown, but it was clearly greater than the mediolateral width. There is no evidence of a palatal portion, incisive process, expanded maxillary process, or a second ascending maxillary process. It appears that the inferior portion of the premaxilla consists only of the bone necessary for attachment of the single tooth.

Type and referred specimens of Clevosaurus hadroprodon sp. nov. Photograph (a) and outline (b) of holotype partial skull (MMACR PV-027-T). Photograph (c) and outline (d) of referred partial left dentary (MMACR PV-028-T). Photograph (e) and outline (f) of posterior view of section in box in c showing cross sectional view of tooth attachement. Abbreviations: Lt Den, left dentary; Lt Mx, left maxilla; MeG, Meckel’s groove; Pmx, right premaxilla; Pmx Fac, premaxillary facet in maxilla; Rt Den, right dentary; Rt Mx, right maxilla; SB, secondary bone; D.V.Cr., dentary ventral crest; Sym, symphyseal region of dentary.
The nasal process of the premaxilla in Clevosaurus hadroprodon differs from the one in Diphydontosaurus34, Gephyrosaurus35, and Planocephalosaurus36 by being vertically-oriented rather than curved. Additionally, the premaxillae of the latter three taxa are more squamate-like than sphenodontian-like in possessing several small teeth along a laterally expanded alveolar portion of the bone. Additionally, C. hadroprodon does not show the wide palatal shelf on the medial portion of the premaxilla as seen in Gephyrosaurus35 and Diphydontosaurus34. A nearly vertical nasal process of the premaxilla is also present in Rebbanasaurus, Godavariasaurus21, and other species of Clevosaurus28. In Rebbanasaurus and Godavariasaurus the process is shorter than that of C. hadroprodon. In most Clevosaurus species the nasal processes tend to be long, with the condition in C. brasiliensis19 being the most similar to C. hadroprodon in relative length. However, differing from C. hadroprodon, C. brasiliensis also possesses a posterodorsal process of the premaxilla19,21,28, which is typical of most other known sphenodontians (e.g., Homoeosaurus, Palaeopleurosaurus, Piocronus, Pleurosaurus, Priosphenodon) and is also present in many archosauromorphs as well as the lepidosauromorphan Kuehneosaurus22. The absence of this process evidenced that only maxilla could have formed the posterior margin of the external naris. This feature is also seen in the early rhynchocephalians, Gephyrosaurus35 and Diphydontosaurus34, respectively, as well as in Sphenodon.
A single premaxillary tooth is also present in the Clevosaurus bairdi18, C. brasiliensis25 as well as in Kallimodon (pers. obs. TRS) and Sphenodon. The single premaxillary tooth of C. hadroprodon is massive and cylindrical making it more characteristically tusk-like. The longitudinal groove on the premaxillary tooth is likely a shallow facet on the distal surface (terminology following Smith and Dodson37) of the tooth to accommodate the tip of the corresponding tusk-like tooth in the first dentary tooth position.
Maxilla
The right maxilla of the holotype (MMACR PV-027-T) is exposed in lateral view (Fig. 2a,b), preserving the portion directly associated with the tooth row, but is otherwise badly crushed and missing the articular portions of the anterior and posterior margins. The preserved portion is mostly the robust, smooth “secondary bone” (see below) that forms at the bases of the teeth which is typical of many sphenodontians. A small portion of the facial process is exposed posteriorly and appears to form a curved posterior process rising above the terminal tooth. This may be the jugal process but is too poorly preserved to be certain. The left maxilla is embedded in the matrix deep to the other elements, but the anteriormost portion is visible in medial view inferior to the premaxilla. Anterior to the two preserved teeth (rotated slightly posteriorly due to breakage of the jaw) is what appears to be a short “diastema”. Anterior to this is a deeper notch, presumably the articulation facet for the premaxilla. Although poorly preserved, the anterior margin of the facial process appears similar to that of most Clevosaurus species in being tall, vertical, and ascending from above the anterior margin of the premaxillary facet. The maxillary dentition is similar to that of the dentary (see below).
Dentary
The anterior portions of both the left (medial view) and right (lateral view) dentaries are present in MMACR PV-027-T (Fig. 2a,b) and the lateral view of the anterior portion of the left dentary is preserved in MMACR PV-028-T (Fig. 2c,d). All three of the dentary specimens are crushed making it difficult to evaluate the anatomy inferior and posterior to the more robust anterior alveolar portions. None of the lower jaw specimens preserve discernable postdentary elements. Based on the preserved portion (nearly 10 mm) of the tooth row in the left dentary of the holotype, this element is relatively long and deep. The best-preserved portion of both dentaries is the massive subdental bone associated with the anterior portion of the tooth row. On the holotype dentaries this subdental bone is exposed in both lateral and medial views. The lateral side is convex, and bears a distinct, ventrally directed, mental process (particularly apparent on MMACR PV-028-T, Fig. 2c,d). There appear to be several small mental foramina near the symphysis (holotype MMACR PV-027-T, Fig. 2a,b), but the surface is heavily pitted, and it is not possible to confidently distinguish the pittings from possible foramina. The medial surface is more vertical with a tall, shallow longitudinal groove extending from the base of the anteriormost tooth to approximately the eighth tooth position. The anteriormost end of this groove appears to extend to the symphysis, but it is not clear if this is not an artifact of the damage to the symphysial portion of the element. More posteriorly the subdental bone flattens, appears blade-like, and bears an articulation facet presumably for the squamous contact with the articular and possibly the prearticular. Inferior to the subdental bone is the deep Meckel’s groove, which is widely open at the symphysis, narrows posteriorly as a short isthmus, and then expands again as it continues posteriorly. The inferior and posterior portions of the dentary are not sufficiently well preserved to further evaluate the Meckel’s groove. The symphyseal portion of the dentary was formed from a relatively thin, ventrally oriented tab that projected below the inferior margin of the dentary.
In comparison with other sphenodontians, the dentary of Clevosaurus hadroprodon is unique in its overall structure. It is apparently similar in its substantial depth but with a much longer tooth row than those of adult C. brasiliensis (see Romo de Vivar and Soares38 for a review of the ontogeny of C. brasiliensis mandibles). Many sphenodontians with relatively long tooth rows such as living Sphenodon and such fossil taxa as Sphenocondor, Sphenovipera, and Tingitana29,39,40 also have mandibles that are long and gracile. Most crown-group sphenodontians are characterized by having dentaries that are relatively short and deep.
The shallow groove dorsal to the Meckelian canal near the symphysis is similar to that described for Cynosphenodon41, but differs in Clevosaurus hadroprodon in being longer and possessing a narrow, rather than deep, ridge of bone separating it from the Meckelian canal. If the narrow extension of this groove beneath the base of the tusk-like dentary tooth towards the symphysis is not an artifact of preservation, then, as with Cynosphenodon41, it may indicate that the holotype specimen of C. hadroprodon is a juvenile (probably between the T3-T4 stages of Robinson42).
The Meckelian canal of Clevosaurus hadroprodon is most similar to that found in C. brasiliensis (Hsiou et al.25) and Sphenotitan leyesi43 in being widely open with a constriction just posterior to the symphysis. A dentary (NHMUK R6102) referred to Clevosaurus hudsoni also has a widely open Meckelian canal, but lacks a conspicuous isthmus posterior to the symphysis, but another dentary (UMZC T1307) for the same taxon does exhibit this morphology suggesting it may be variable (TRS, pers. obs.). The groove dorsal to the Meckelian canal in C. hadroprodon is more pronounced than the narrow groove found in one of the dentaries of C. hudsoni (UMZC T1307), but in the case of C. hadroprodon. the dorsal margin of the groove is bounded by bone rather than by the ventral edge of “secondary bone”.
Dentition
The holotype has heterodont dentition in the upper and lower marginal tooth rows characterized by the distinct dental regions developed in mesiodistal (anteroposterior) sequence (Fig. 2a–d). This includes one anteriormost successional tooth, represented by the single, large, tusk-like tooth on the dentary (or ‘caniniform’ successional tooth, see Apesteguía et al.29), and an equivalent tusk-like tooth in the premaxilla. Clevosaurus hadroprodon has an alternating tooth series distal to the first tooth position in the dentary represented by at least three teeth. There could be additional dentary teeth in this series, but these are not observable in both dentaries due to overlapping of the jaw elements. The anterior portion of the maxilla also has an alternating tooth series, represented by the first six preserved maxillary teeth in the right maxilla. Finally, the alternating series in both jaws is succeeded distally by an additional tooth series, comprised of six preserved teeth in the right maxilla. The number of teeth in the additional series on the dentaries is unknown due to poor preservation of the posterior dentary region.
The single successional tooth of the premaxilla is massive, conical, blunt (tip may be worn), and vertically oriented. The base is firmly ankylosed to the premaxilla and there is no evidence of “secondary bone” (sensu Harrison26,27; Jones22; AKA “secondary dentine” of Fraser28; since the histological characteristics of this tissue have not yet been demonstrated we have adopted the more inclusive term ‘secondary bone’ but use quotes to denote uncertainty of its specific structure, but see Jones22 for a review of alternative terminology).
The tusk-like tooth in the dentary occupies the first tooth position above the symphysis. This tooth, similar to that of the premaxilla, is mesiodistally massive, conical in shape, and apicobasally procumbent (approximately 30°), and is attached to the tip of the dentary slightly below the attachment region of the remaining dentary teeth.
The remaining teeth of the dentaries and the maxillae are similar in morphology. The longest of the dentary tooth rows preserves approximately 19 tooth positions and the most complete maxillary tooth row has 12 tooth positions, though both are broken posteriorly suggesting that several teeth are missing in both the upper and lower marginal tooth rows. Apart from the tusk-like teeth in the first dentary tooth position and in the premaxilla, all the preserved tooth crowns are similar in being labiolingually compressed, generally triangular, and with distinct mesiodistal carinae, but lacking the labiolingual flanges common to other sphenodontians. The teeth also differ from each other in relative size along the tooth row with shorter, narrower (mesiodistally) teeth anteriorly in the dentary and maxilla and taller, wider teeth posteriorly, but with smaller teeth interspersed among these larger teeth. The exact pattern cannot be determined due to damage along each of the tooth rows. There is no evidence in any of the specimens of active tooth replacement.
In the dentaries there is a thin ridge of “secondary bone” that obscures the tooth-jaw contact in both labial and lingual views with the labial being more prominent. All tooth bases of the holotype are fused and ankylosed to the apex of the jaws and lack any discernable evidence of active tooth replacement. There is no clear evidence of this tissue at the bases of the maxillary teeth, but only the labial surface is clearly visible. The “secondary bone” lacks any evidence of wear facets.
Comparative osteology of Clevosaurus hadroprodon to other rhynchocephalians
Clevosaurus hadroprodon can be classified within the Rhynchocephalia, and more specifically within Sphenodontia, based on several characters of jaw morphology: both anterior and posterior tooth series apically placed and ankylosed to the jaw bone labial margin; presence of ‘secondary bone’ deposition along the tooth-jaw contact; a dorsoventrally deep and vertically oriented symphyseal margin of the dentary; deep ventral crest of the dentary medial wall; and the presence of an anterior canine tooth that is apically placed and fused to the dentary. Although some of these features also occur within acrodontan squamates (which generally have a remarkable degree of convergence with sphenodontians), the combination of features in C. hadroprodon is only seen in sphenodontians (see Supplementary Data 1).
Clevosaurus hadroprodon shares with most rhynchocephalians the presence of acrodont tooth implantation in association with ‘secondary bone’. This does differ from the early rhynchocephalians Gephyrosaurus and Diphydontosaurus in that these two taxa lack ‘secondary bone’ and are characterized by pleurodont attachment of polyphyodont teeth along the entire tooth row (Gephyrosaurus), or, the combination of anteriorly pleurodont and posteriorly acrodont attachment (Diphydontosaurus)21,28,34,35,44. Clevosaurus hadroprodon is thus more similar to later evolving forms by having a fully acrodont tooth implantation, and most of them also develop the ‘secondary bone’ in mature individuals at the tooth-jaw contact following the dental line22,28,29,44,45. However, Clevosaurus hadroprodon has a less developed layer of ‘secondary bone’. Whether this reduced development of ‘secondary bone’ is related to taxonomy or ontogeny is unknown (absence is considered a juvenile feature46).
Among sphenodontians, Clevosaurus hadroprodon is additionally unusual in its tooth morphology. The teeth are labiolingually compressed, lacking the development of labiolingual features of the marginal dentition common to most other sphenodontians (e.g. a posterolingual crest in Clevosaurus species, Kallimodon, Homeosaurus, Pleurosaurus, and Palaeopleurosaurus—TRS, pers. obs.; broadly expanded posterior teeth with anteriorly directed concavity teeth in opisthodontians). The absence of labiolingual dental features also occurs in the heterodont teeth of Whitakersaurus from the Late Triassic of New Mexico, USA47, and Rebbanasaurus and Godavarisaurus, from the Middle Jurassic–Early Cretaceous of the Kota Formation, India21. Among the latter, Clevosaurus hadroprodon also shares with Rebbanasaurus an extremely deep symphysial margin of the dentary, although differing from the latter in lacking striated tooth crowns.
The presence of a single tooth on the premaxilla of Clevosaurus hadroprodon is shared with the sphenodontians Priosphenodon and Sphenodon as well as the clevosaurs C. brasiliensis19,25, C. bairdi18 and specimens currently attributed to Clevosaurus sp. from China17,22, differently from the pattern observed in other clevosaurs, such as C. hudsoni, C. convalis and C. sectumsemper that possess 2–4 premaxillary teeth11,28,48. In the stem rhynchocephalians Gephyrosaurus and Diphydontosaurus there are typically five or more small pleurodont teeth on the premaxilla34,35, and Planocephalosaurus possesses four subpleurodont premaxillary teeth (“semi-pleurodont”28,34).
The anterior tusk-like tooth in the premaxilla of Clevosaurus hadroprodon is most similar to C. brasiliensis in being tusk-like (e.g., Hsiou et al.25; TRS pers. obs.), and not chisel-shaped as in Sphenodon, Priosphenodon, and Vadasaurus (composed of three fused premaxillary teeth49). Additionally, an enlarged successional tooth (“canine-tooth”) in the dentary has also been described for Cynosphenodon and Sphenovipera23,40, but these occupy tooth positions posterior to the first tooth. The most similar condition to the enlarged, tusk-like, anteriormost tooth of the dentary in C. hadroprodon is possibly the enlarged, albeit of a much smaller relative size, anteriormost dentary tooth of Sphenocondor. However, in Sphenocondor the symphysial portion of the dentary is poorly preserved making it difficult to confirm the exact dimensions of this tooth27. A single, large tooth in the first dentary tooth position is also present in Cynosphenodon huizachalensis23,41. However, in C. huizachalensis this tooth is conical and preceded by an edentulous gap in adults and in juveniles is preceded by at least three “anterior hatchling teeth” and one “first generation successional tooth”41. According to Apesteguía et al.29 the precise homology between the ‘caniniform’ tooth and other successional teeth remains uncertain in fossil taxa, and in this way, we consider that the tusk-like, anteriormost tooth of the dentary in C. hadroprodon as a successional tooth that is larger than subsequently successional teeth29.
As noted above, although important similarities exist between Clevosaurus hadroprodon and Mexican and Indian forms (Cynosphenodon and Rebbanasaurus), the combination of features in C. hadroprodon is unique among known sphenodontians and other clevosaurs, thus supporting its designation as a new species of Clevosaurus.
Discussion
The labiolingually compressed marginal dentition of Clevosaurus hadroprodon is generally similar to the overall dimensions of the “cut and slice” sphenodontian tooth form22 including the marginal teeth of Clevosaurus spp. and the sphenodontine sphenodontians50. However, Clevosaurus hadroprodon differs from these taxa (and nearly all other sphenodontians) in the lack of obvious wear facets on the teeth indicating that the feeding strategy of this new taxon likely did not utilize the intense oral food-processing common to most sphenodontians. The sharply pointed dentition is more similar to that of Agama51 and juvenile Uromastyx52—though these acrodontan lizards also show heavy wear facets in the teeth, particularly in older individuals—and suggests a similar arthropod-based diet. It is possible that the specimens of C. hadroprodon represent juveniles that had not yet been able to achieve significant wear in the marginal dentition, but the presence of “secondary bone” indicates that these specimens had undergone some appreciable ontogenetic development (sensu Duffin46). The presence of a single, large tooth in the premaxilla and the dentary of C. hadroprodon is similar to the one observed in Agama, but in C. hadroprodon these teeth are tusk-like (straight and blunt) rather than sharply pointed and recurved. It is possible that such teeth could be used to subdue a prey item, but it is also possible that they served to aid in non-feeding behaviors such as mate competition or defense. The lack of palatal elements precludes assessment of this dentition and associated implications on feeding strategy, mechanism, or diet.
The teeth of Clevosaurus hadroprodon represent the oldest occurrence of the typical fully acrodont dentition of sphenodontians anywhere in the world, thus providing insights on the early stages of the development of the acrodont dentition in sphenodontians. In sum, the dental morphology in C. hadroprodon demonstrates that the presence of a fully acrodont dentition, reduction in premaxillary tooth count and development of large successional teeth on the premaxillae, typical of most sphenodontians, had evolved by the Late Carnian. Conversely, the simple, triangular shape of the marginal teeth in C. hadroprodon suggests that accessory crests—a feature common to the dentition of most sphenodontians—possibly evolved after the acquisition of the fully acrodont dentition in sphenodontians. But, as stated above, this simple tooth form may be an ontogenetic feature and additional, more complete specimens are needed to verify either of these possibilities.
Phylogenetic analysis recovers Clevosaurus hadroprodon within Clevosauridae. In a parsimony analysis (Fig. 3a) C. hadroprodon resolves within the genus Clevosaurus, whereas in an analysis utilizing bayesian optimality criterion (Fig. 3b), C. hadroprodon is recovered in a polytomy comprised of Brachyrhinodon and the Clevosaurus species. The erection of a new species is justified, although it lacks significant support given: (1) the overall morphological differences between the new species compared to Clevosaurus spp. (see comparative osteology section); and (2) the result of the bayesian inference, which does not recover the genus Clevosaurus as monophyletic. The poor resolution among the clevosaurids and related taxa in the bayesian topology, in addition to the position of Polysphenodon in the parsimony analysis, demonstrates the need for careful and detailed anatomical and systematic revision of Clevosauridae to clarify the interrelationships within these sphenodontians (see also Hsiou et al.25).

Phylogenetic analyses evidencing the position of Clevosaurus hadroprodon sp. nov. within Rhynchocephalia using two distinct optimality criteria. (a) Resultant topology of the strict consensus of eight most parsimonious tree (249 steps each) (CI = 0.43; RI = 0.69); (b) Majority-rule consensus tree derived from the Bayesian analysis. Nodes values represent posterior probabilities. (Note that Clevosauridae sensu Bonaparte and Sues, was not recovered as a monophyletic group in both analyses and the genus Clevosaurus was not recovered as monophyletic in the Bayesian topology). Abbreviations: “Cle”: Clevosauridae; Hom: Homeosauridae; Pleu: Pleurosauridae; Opi: Ophistodontia.
The Carnian age of Clevosaurus hadroprodon makes it the oldest known unequivocal sphenodontian from southern Pangaea (Gondwana). Apart from this new record, the former oldest records are the Brazilian Clevosaurus brasiliensis19,24,25,38 (Norian age; 225.42 ± 0.37 Ma, representing the maximum age of the Riograndia AZ, at the top of the Candelária Sequence33) and the early opisthodontid sphenodontian Sphenotitan leyesi43 from the late Norian–Rhaetian of the Quebrada del Barro Formation in Argentina53,54 (see Supplementary Information). Clevosaurus hadroprodon also represents an early diversification of specialized sphenodontians that occurred within the Gondwanan region of Pangaea prior to the Late Triassic.
A major component of this early diversification of sphendontians is driven by the origins of clevosaurs, within which Clevosaurus hadroprodon figures among the oldest known representatives (Fig. 4), along with Brachyrhinodon and Polysphenodon from the late Carnian–Norian55 of Britain (see also Supplementary Fig. 1 for maximum clade credibility tree and Supplementary Data 2).

Time–scaled strict consensus topology of Rhynchocephalian relationships (derived from the parsimony analysis). Thickened lines represent the range of age of each taxon according the first appearance date (FAD) and last appearance date (LAD) of the fossil record (see supplementary data 3).
The occurrence of Clevosauridae in the late Carnian is notable in that it is approximately 10 my younger than the oldest known rhynchocephalian, cf. Diphydontosaurus (“Vellberg jaws” specimens) from the Ladinian (Middle Triassic) of Germany5 and only 18 million years younger than the estimated divergence between Gephyrosaurus and all other rhynchocephalians2. Therefore, clevosaurs comprised the earliest evolving major group of lepidosaurs (with six species that eventually dispersed throughout five different continents), and seem to have developed a widespread geographical distribution very early in their evolutionary history. By the end of the Carnian, clevosaurs were present in at least two distantly located regions of Pangaea that today constitute parts of the United Kingdom and Brazil18,22,25. By the Early Jurassic clevosaurs had further dispersed across the fragmenting remains of Pangaea into North America18, Asia17, and South Africa16. As such, clevosaurs represent the earliest group of lepidosauromorphs to achieve such cosmopolitan distribution.
The dentition of Clevosaurus hadroprodon represents the oldest occurrence of the typical fully acrodont dentition of sphenodontians anywhere in the world, thus providing insights on the early stages of the development of the acrodont dentition in sphenodontians. For instance, the morphology of Clevosaurus hadroprodon suggests that accessory crests that are commonly observed on the dentition of most sphenodontians developed at a later stage in sphenodontian evolution, after the acquisition of the fully acrodont dentition, but not to the exclusion of other dental specializations such as the tusk-like teeth present in this taxon. It further suggests that the reduction in the number of premaxillary teeth and development of large successional teeth on the premaxillae, also typical of most sphenodontians, occurred by the Late Carnian, along with the development of a fully acrodont dentition.
Clevosaurus hadroprodon (Fig. 5), along with Clevosaurus brasiliensis, indicate that some of the oldest known sphenodontians evolved in South America, and along with Indian taxa, such as Godavariasaurus and Rebbanasaurus, indicate that some of the earliest diverging members of the Sphenodontia occurred in geographically distant parts of southern Pangaea, during the initial breakup of Gondwana. This illustrates the importance of the role of the Gondwanan lepidosaur fauna in our growing understanding of the earliest stages of sphenodontian evolution and global biogeographic distribution.

Life reconstruction of Clevosaurus hadroprodon. Reconstruction created by Jorge Blanco.
Methods
Fossil material
Sphenodontian specimens described here are stored at the Museu Municipal Aristides Carlos Rodrigues (MMACR-PV-T, Paleovertebrates-Triassic Collection), Candelária municipality, Rio Grande do Sul State, Southern Brazil. All specimens are incomplete and poorly preserved. The holotype (MMACR PV-027-T) corresponds to an incomplete right premaxilla, an incomplete right maxilla and right lower jaw in occlusion aspect, and left lower jaw located below to these elements. Additional specimens include an incomplete left lower jaw (MMACR PV-028-T) and a fragment of dentary with three isolated teeth (MMACR PV-029-T). Several early and late Mesozoic sphenodontians were used in the comparative morphology, mostly in firsthand examination (by ASH and TRS).
Type horizon and locality
Clevosaurus hadroprodon was collected in the Linha Bernardino outcrop, located in the outskirts of the Candelária municipality (UTM 22J, 324483 E/6717888N, datum WGS 84), Rio Grande do Sul State, Southern Brazil (Fig. 1a–c). The Triassic exposure occurs in a ravine of about six meters depth composed of fine to medium-grained sandstones, intercalated with mudstone layers and conglomerates (Fig. 1b). Carbonate concretions occur along most of the section, and the fossils tend to occur in association with the concretions. The specimens referred to Clevosaurus hadroprodon were originally embedded in concretions, abundant at the lowermost conglomeratic level of the outcrop. Other specimens, representing other taxa, were collected in the fine sandstone levels, also associated with concretions. They include partial lower jaws and some postcranial elements (e.g., humerus) referred to the traversodontid cynodont Exaeretodon riograndensis, partial skull referred to the probainognathian cynodont Trucidocynodon sp., and fragments of long bones of a dinosaur specimen, among other indeterminate materials. A fragment of a femur resembles the early dinosaur Pampadromaeus barberenai56,57; although further preparation and comparisons are needed to clarify its taxonomy. The uppermost portion of the exposure consists of pinkish non-fossiliferous massive sandstone (Fig. 1b). The presence of specimens referred to Exaeretodon, Trucidocynodon and a Pampadromaeus-like form allows correlation of this outcrop as part of the Hyperodapedon AZ, at the base of the Candelária Sequence. The fossil samples from this outcrop are relatively scarce in comparison to other Triassic localities in the state of Rio Grande do Sul and a precise estimation of its relative age is only tentative. Exaeretodon, Trucidocynodon and a Pampadromaeus-like form have also been recovered from the Janner Site, in the municipality of Agudo. At this locality these taxa are part of a more diverse fauna including probainognathian (e.g. Trucidocynodon riograndensis) and traversodontid (Exaeretodon riograndensis) cynodonts58,59, dinosaurs and rhynchosaurs. Based on its faunal content and its relationships with other outcrops from southern Brazil, the Linha Bernardino site can be also correlated to the Ischigualasto Formation (Ischigualasto-Villa Unión Basin, Argentina60,61). A recent analysis provided a high-precision U-Pb zircon geochronology to some sections of south Brazil Triassic exposures, including a typical Hyperodapedon AZ locality (i.e. Cerro da Alemoa)33. The recovered weighted mean 206Pb/238U date of 233.23 ± 0.73 Ma is slightly older than the datum for the Ischigualasto Formation33. Nonetheless, the combined data suggest that the age of the faunal association of the Linha Bernardino site is more likely Late Carnian (MCL, pers. comm.).
Phylogenetic analyses and time scaled–tree
Taxon scoring changes
Sophineta was deleted from the dataset because only some of the skull and mandible elements can be confidentially linked to a single taxon, with all the referred postcranial material lacking any anatomical connection to the skull and mandible elements (TRS pers. obs.). Since deletion of all postcranial scorings would highly reduce the usefulness of this taxon as an outgroup by re-scoring several cells with missing data, we deleted this taxon altogether from the analysis.
All specimens attributed to Clevosaurus brasiliensis were merged into a single operational taxonomic unit (OTU), since all of them have been subject of a recent revision of this species and confidently attributed to a single taxon25. We agree with the interpretation of lack of diagnostic features to separate C. wangi, C. mcgilli and C. petilus18,22, and thus we follow Sues et al.17 in considering all three Chinese clevosaurs as a single OTU, represented by C. petilus.
Character changes
Characters 62 and 63 were scored with missing data (“?”) for all taxa and were therefore removed from the present dataset. New character 68 added.
Character-state changes
New states added herein: Ch. 47, state 3.
Taxon scoring and re-scoring
Ch. 8. Clevosaurus sp. SAM:?→0; Clevosaurus brasiliensis: 0/1→0 (only specimen with a “deep” maxilla has the posterior end of the maxilla incomplete, with the process extending ventral to the broken jugal).
Ch. 12. Clevosaurus petilus: 0→1; Brachyrhinodon: 0→?.
Ch.19: Clevosaurus brasiliensis: 1→0.
Ch.21: Clevosaurus SAM: 2→? (jugal posterior border seemingly broken); Clevosaurus petilus 1→? (jugal posterior border poorly preserved).
Ch.33. Clevosaurus latidens: 3→? (symphysis not preserved); Sphenotitan: 3→0; Oenosaurus: 2/3→2; Planocephalosaurus: 1→2; Pelecymala: 1→2. Beachyrhinodon 1→? (symphysis poorly preserved); Clevosaurus hudsoni: 1→2; Clevosaurus convalis: 1→0; Clevosaurus bairdi: 1→2; Clevosaurus SAM:?→2; Clevosaurus brasiliensis: 1→2.
Ch. 36: Clevosaurus SAM:?→0; Oenosaurus: 1→?.
Ch. 44: Pleurosaurus goldfussi 2→ 3 (BSPG 1925 I 18 [holotype], BSPG 1978 I 7); Pleurosaurus gingsburi 2/3→3 (MNHN 1983-4-CNJ 67 [holotype], BSPG 1977 XIX 40); Palaeopleurosaurus posidoniae 2→3 (SMN 50721).
Ch.47: Opisthias: 1→2.
Ch. 50. Brachyrhinodon 1→?.
Ch. 51. Brachyrhinodon 1→?.
Ch. 57: Clevosaurus brasiliensis: 0→?.
Ch. 69: Sphenotitan: 2→?.
Ch. 72: Clevosaurus bairdi: 2→3; Clevosaurus hudsoni: 2→1; Clevosaurus brasiliensis: 2→1; Beachyrhinodon 2→?; Clevosaurus convalis: 2→?.
Rogue taxon identification
An initial set of analyses (Analysis 1) was conducted with all taxa initially available in the data set. This analysis resulted in consensus trees with important sectors of the tree highly unresolved (results available in Supplementary Information). Therefore, we conducted a rogue taxon identification analysis using the RogueNaRok algorithm, which can improve tree accuracy and resolution when specific wildcards are the main factor contributing to poorly resolved nodes62. We also utilized the TNT pruning trees algorithm to detect wild card taxa in specific nodes of the consensus tree inferred with maximum parsimony. The results indicate that the removal of Clevosaurus convalis, Sphenovipera and Theretairus would highly improve phylogenetic resolution. A second set of analyses were conducted after removal of those three taxa, which yielded better trees that are reported in the main text.
Phylogenetic analyses
Maximum parsimony analysis was conducted in TNT v. 1.163 using 100 tree replicates obtained by random addition sequence (RAS), and searching for new tree topologies with tree bisection and reconnection (TBR), saving 100 trees per replication. For Analysis 1, a total of 907 most parsimonious trees (MPTs) were obtained with 258 steps each. After removal of the three rogue taxa, the 40 remaining taxa were analyzed in a new set of analyses (Analysis 2). In Analysis 2, we recovered a total of eight MPTs with 249 steps each.
The Bayesian inference analysis was conducted using Mr. Bayes v. 3.2.664 using the Mkv model for morphological data65, and with rate variation across characters sampled from a gamma distribution. Each analysis was performed with two independent runs of 1 × 107 generations each, with eight chains per run and four swaps attempted per swapping generation. The relative burn-in fraction was set to 50% and the chains were sampled every 50 generations. The temperature parameter for the four chains in each independent run was set to 0.02. Convergence of independent runs was assessed through the average standard deviation of split frequencies (ASDSF < 0.01) and potential scale reduction factors (PSRF ≈ 1 for all parameters) calculated at the end of the Bayesian runs. We used Tracer v. 1.666 to determine whether the runs reached stationary phase and to ensure that the effective sample size (ESS) for each parameter was greater than 200.
The strict consensus tree calculated from the eight MPTs from the parsimony Analysis 2 was time–scaled using the package strap67 for the software R68. The range of time for each taxon was delimited consulting the relevant literature (see Supplementary Data 3 and 4). It was employed in the scaling, the Brussate parameter69, in which all branch lengths are shared equally among the tree, in order to avoid zero values (i.e. only retaining positive length for each branch).
Data Availability
The phylogenetic analyses, time scaled–tree and geological unit and time range references of the Rhynchocephalia taxa data of this study are available as Supplementary Information.
Change history
06 May 2021
A Correction to this paper has been published: https://doi.org/10.1038/s41598-021-88869-2
References
Sues, H.-D. & Fraser, N. C. Triassic Life on land: the great transition. 236 (Columbia University Press, 2010).
Simões, T. R. et al. The origin of squamates revealed by a Middle Triassic lizard from the Italian Alps. Nature 557, 706–709 (2018).
Uetz, P & Hosek, J. The Reptile Database. Available at, http://www.reptile-database.org/ (2017).
Jones, M. E. H. & Cree, A. Tuatara. Curr. Biol. 22, 986–987 (2012).
Jones, M. E. et al. Integration of molecules and new fossils supports a Triassic origin for Lepidosauria (lizards, snakes, and tuatara). BMC Evol. Biol. 13, 208 (2013).
Whiteside, D. I. & Duffin, C. J. Late Triassic terrestrial microvertebrates from Charles Moore’s ‘Microlestes’ quarry, Holwell, Somerset, UK. Zool. J. Linn. Soc. 179, 677–705 (2017).
Whiteside, D. I., Duffin, C. J. & Furrer, H. The Late Triassic lepidosaur fauna from Hallau, North-Eastern Switzerland, and a new ‘basal’ rhynchocephalian Deltadectes elvetica gen. et sp. nov. Neues Jahrb. fur Geol. und Palaontologie - Abhandlungen 285, 53–74 (2017).
Swinton, W. E. A new Triassic rhynchocephalian from Gloucestershire. J. Nat. Hist. Ser. 11(4), 591–594 (1939).
Fraser, N. C. & Benton, M. J. The Triassic reptiles Brachyrhinodon and Polysphenodon and the relationships of the sphenodontids. Zool. J. Linn. Soc. 96, 413–445 (1989).
Fraser, N. C. A new sphenodontian from the early Mesozoic of England and North America: implications for correlanting early Mesozoic continental deposits. New Mex. Museum Nat. Hist. Sci. Bull. 135–139 (1993).
Säilä, L. K. A new species of the Sphenodontian reptile Clevosaurus from the lower Jurassic of South Wales. Palaeontology 48, 817–831 (2005).
Keeble, E., Whiteside, D. I. & Benton, M. J. The terrestrial fauna of the Late Triassic Pant-y-ffynnon Quarry fissures, South Wales, UK and a new species of Clevosaurus (Lepidosauria: Rhynchocephalia). Proc. Geol. Assoc. 1–21 (2018).
Jaekel, O. Ein Uebersicht über die Wirbeltiere. Berlin: Borntraeger. 145–146 (1911).
Huene, F. V. Uber Rhynchosaurier und andere Reptilien aus den Gondwana- Ablagerungen Sudamerikas. Geol. und Palaontologische Abhandlungen 17, 1–62 (1929).
Gow, C. E. & Raath, M. A. Fossil vertebrate studies in Rhodesia: sphenodontid remains from the Upper Triassic of Rhodesia. Palaeontol. Africana 20, 121–122 (1977).
Sues, H. D. & Reisz, R. R. First record of the early Mesozoic sphenodontian Clevosaurus (Lepidosauria: Rhynchocephalia) from the Southern Hemisphere. J. Paleontol. 69, 123–126 (1995).
Wu, X.-C. In In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods (eds Fraser, N. C., Sues, H.-D.) 38–69 (Cambridge University Press, 1994).
Sues, A. H., Shubin, N. H. & Olsen, P. E. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the Mccoy Brook Formation (Lower Jurassic) of Nova Scotia, Canada. J. Vertebr. Paleontol. 14, 327–340 (1994).
Bonaparte, J. F. & Sues, H. D. A new species of Clevosaurus (Lepidosauria: Rhynchocephalia) from the Upper Triassic of Rio Grande do Sul, Brazil. Palaeontology 49, 917–923 (2006).
Wilkinson, M. & Benton, M. J. Sphenodontid phylogeny and the problems of multiple trees. Philos. Trans. R. Soc. B Biol. Sci. 1–16 (1996).
Evans, S., Prasad, G. & Manhas, B. Rhynchocephalians (Diapsida: Lepidosauria) from the Jurassic Kota Formation of India. Zool. J. Linn. Soc. 133, 309–334 (2001).
Jones, M. E. H. The Early Jurassic clevosaurs from China (Diapsida: Lepidosauria). New Mex. Museum Nat. Hist. Sci. Bull. 548–562 (2006).
Reynoso, V.-H. A Middle Jurassic Sphenodon-like sphenodontian (Diapsida: Lepidosauria) from Huizachal Canyon, Tamaulipas, Mexico. J. Vertebr. Paleontol. 16, 210–221 (1996).
Arantes, B., Soares, M. & Schultz, C. Clevosaurus brasiliensis (Lepidosauria, Sphenodontia) do Triássico Superior do Rio Grande do Sul: anatomina pós-cranianana e relações filogenéticas. Rev. Bras. Paleontol. 12, 43–54 (2009).
Hsiou, A. S., França, M. A. G. & Ferigolo, J. New data on the Clevosaurus (Sphenodontia: Clevosauridae) from the Upper Triassic of Southern Brazil. PLoS One 10, 1–21 (2015).
Harrison, H. S. Hatteria punctata, its dentitions and its incubation period. Anat. Anz. 20, 145–158 (1901).
Harrison, H. S. The development and succession of teeth in Hatteria punctata. Q J Microsc Sci 44, 161–219 (1901).
Fraser, N. C. The osteology and relationships of Clevosaurus (Reptilia: Sphenodontida). Philos. Trans. R. Soc. B Biol. Sci. 321, 125–178 (1988).
Apesteguía, S., Gómez, R. O. & Rougier, G. W. A basal sphenodontian (Lepidosauria) from the Jurassic of Patagonia: New insights on the phylogeny and biogeography of Gondwanan rhynchocephalians. Zool. J. Linn. Soc. 166, 342–360 (2012).
Apesteguía, S., Gómez, R. O. & Rougier, G. W. The youngest South American rhynchocephalian, a survivor of the K-Pg extinction. Proc. R. Soc. B 281, 1–6 (2014).
Zerfass, H. et al. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: a contribution to Southwestern Gondwana palaeogeography and palaeoclimate. Sediment. Geol. 161, 85–105 (2003).
Horn, B. L. D. et al. A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. J. South Am. Earth Sci. 55, 123–132 (2014).
Langer, M. C., Ramezani, J. & Da Rosa, Á. A. S. U-Pb age constraints on dinosaur rise from south Brazil. Gondwana Res. 57, 133–140 (2018).
Whiteside, D. I. The head of the Rhaetian sphenodontid Diphydontosaurus avonis gen. et sp. nov. and the modernizing of a living fossil. Philos. Trans. R. Soc. B Biol. Sci. 597–597 (1986).
Evans, S. The skull of a new eosuchian reptile from the Lower Jurassic of South Wales. 203–264 (1980).
Fraser, N. C. A new rhynchocephalian from the British Upper Trias. Palaeontology 25, 709–725 (1982).
Smith, J. B. & Dodson, P. A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. J. Vertebr. Paleontol. 23, 1–12 (2003).
Romo de Vivar, P. R. & Bento Soares, M. Dentary morphological variation in Clevosaurus brasiliensis (Rhynchocephalia, Clevosauridae) from the Upper Triassic of Rio Grande do Sul, Brazil. PLoS One 10, e0119307 (2015).
Evans, S. E. & Sigogneau-Russell, D. New sphenodontians (Diapsida: Lepidosauria: Rhynchocephalia) from the Early Cretaceous of North Africa. J. Vertebr. Paleontol. 17, 45–51 (1997).
Reynoso, V.-H. Possible evidence of a venom apparatus in a Middle Jurassic sphenodontian from the Huizachal red beds of Tamaulipas, México. J. Vertebr. Paleontol. 25, 646–654 (2005).
Reynoso, V.-H. Growth patterns and ontogenetic variation of the teeth and jaws of the Middle Jurassic sphenodontian Cynosphenodon huizachalensis (Reptilia: Rhynchocephalia). Can. J. Earth Sci. 40, 609–619 (2003).
Robinson, P. L. In Morphology and Biology of Reptiles (eds Bellairs, A. d’A. & Cox, B.) 43–64 (Linnean Society of London by Academic Press, 1976).
Martinez, R. N. et al. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the Late Triassic of Argentina and the early origin of the herbivore opisthodontians. Proc. R. Soc. B Biol. Sci. 280, 20132057–20132057 (2013).
Evans, S. E., Prasad, G. V. R. & Manhas, B. K. Fossil lizards from the Jurassic Kota Formation of India. J. Vertebr. Paleontol. 22, 299–312 (2002).
Fraser, C. New Triassic sphenodontids from South-West England and review of their classification. Palaeontology 29, 165–186 (1986).
Duffin, C. J. The first sphenodontian remains (Lepidosauromorpha, Reptilia) from the Late Triassic of the Gaume (southern Belgium). Bull. la Société belge Géologie 104, 35–41 (1995).
Heckert, A. B., Lucas, S. G., Rinehart, L. F. & Hunt, A. P. A new genus and species of sphenodontian from the Ghost Ranch Coelophysis Quarry (Upper Triassic: Apachean), Rock Point Formation, New Mexico, USA. Palaeontology 51, 827–845 (2008).
Klein, C. G., Whiteside, D. I., de Lucas, V. S., Viegas, P. A. & Benton, M. J. A distinctive Late Triassic microvertebrate fissure fauna and a new species of Clevosaurus (Lepidosauria: Rhynchocephalia) from Woodleaze Quarry, Gloucestershire, UK. Proc. Geol. Assoc. 126, 402–416 (2015).
Bever, G. S. & Norell, M. A. A new rhynchocephalian (Reptilia: Lepidosauria) from the Late Jurassic of Solnhofen (Germany) and the origin of the marine Pleurosauridae. R. Soc. Open Sci. 4, 1–16 (2017).
Jones, M. E. H. Skull shape and feeding strategy in Sphenodon and other rhynchocephalia (Diapsida: Lepidosauria). J. Morphol. 269, 945–966 (2008).
Cooper, J. S., Poole, D. F. G. & Lawson, R. The dentition of agamid lizards with special reference to tooth replacement. J. Zool. 162, 85–98 (1970).
Cooper, J. S. & Poole, D. F. G. The dentition and dental tissues of the agamid lizard, Uromastyx. J. Zool. 169, 85–100 (1973).
Martínez, R. N. et al. A new Late Triassic vertebrate assemblage from Northwestern Argentina. Ameghiniana 52, 379–390 (2015).
Martínez, R. N. & Apaldetti, C. A Late Norian—Rhaetian Coelophysid Neotheropod (Dinosauria, Saurischia) from the Quebrada Del Barro Formation, Northwestern Argentina. Ameghiniana 54, 488–505 (2017).
Walker, A. D. The age of the Cuttie’s Hillock Sandstone (Permo-Triassic) of the Elgin area. Scottish J. Geol. 9, 177–183 (1973).
Cabreira, S. F. et al. New stem-sauropodomorph (Dinosauria, Saurischia) from the Triassic of Brazil. Naturwissenschaften 98, 1035–1040 (2011).
Müller, R. T., Langer, M. C. & Dias-da-Silva, S. Biostratigraphic significance of a new early sauropodomorph specimen from the Upper Triassic of southern Brazil. Hist. Biol. 29, 187–202 (2017).
Abdala, F., Barberena, M. C. & Dornelles, J. A new species of the traversodontid cynodont Exaeretrodon from the Santa Maria Formation (Middle/Late Triassic) of southern Brazil. J. Vertebr. Paleontol. 22, 313–325 (2002).
Oliveira, T. V., Soares, M. B. & Schultz, C. L. Trucidocynodon riograndensis gen. nov. et sp. nov. (Eucynodontia), a new cynodont from the Brazilian Upper Triassic (Santa Maria formation). Zootaxa 71, 1–71 (2010).
Martinez, R. N. et al. A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea. Science 331, 206–210 (2011).
Martinez, R. N. et al. Vertebrate succession in the Ischigualasto Formation. J. Vertebr. Paleontol. 32, 10–30 (2012).
Aberer, A. J., Krompass, D. & Stamatakis, A. Pruning rogue taxa improves phylogenetic accuracy: an efficient algorithm and webservice. Syst. Biol. 62, 162–166 (2013).
Goloboff, P. A., Farris, J. S. & Nixon, K. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786 (2008).
Ronquist, F. et al. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542 (2012).
Lewis, P. O. A likelihood approach to estimating phylogeny from discrete morphological character data. Syst. Biol. 50, 913–925 (2001).
Rambaut, A., Suchard, M.A., Xie, D. & Drummond, A. J. Tracer v1.6. Available at, http://beast.bio.ed.ac.uk/Tracer (2014).
Bell, M. A. & Lloyd, G. T. Strap: an R package for plotting phylogenies against stratigraphy and assessing their stratigraphic congruence. Palaeontology 58, 379–389 (2015).
R Development Core Team (2011), R: a language and environment for statistical computing. 900051 Available at: http://www.r-project.org/ (2011).
Brusatte, S. L., Benton, M. J., Ruta, M. & Lloyd, G. T. The first 50 Myr of dinosaur evolution: macroevolutionary pattern and morphological disparity. Biol. Lett. 4, 733–736 (2008).
Acknowledgements
We are grateful to the curator Carlos Nunes Rodrigues from the Museu Municipal Aristides Carlos Rodrigues (MMACR, Candelária municipality, Rio Grande do Sul State, Brazil) for loaning of the fossil sphenodontian for this study. This work was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, grant 2011/14080-0 to A.S.H) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, grant 309434/2015-7 to A.S.H and 401833/2010-0 to C.L.S.).
Author information
Authors and Affiliations
Contributions
A.S.H. conceived the project; A.S.H., R.N., T.R.S., S.O. and M.W.C. designed the research, interpreted and wrote the manuscript with input of all other authors; T.R.S. acquired and conducted phylogenetic analyses; R.N. composed the figures of the specimens; S.O. conducted the time-scale tree analysis in R and conducted the final figures of the specimens; F.P., A.G.M., P.R.R.M., M.B.S. and C.L.S. wrote the interpretations of the geology, type locality and age; F.P. and A.L. collected and prepared the sphenodontian materials. A.S.H., R.N., T.R.S., S.O. and A.G.M. contributed to the review of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this Article was revised: The Nomenclatural Acts and the ZooBank registration LSID numbers for this publication urn:lsid:zoobank.org:pub:02B4B0AF-B553-410B-94B1-76D7AE277FAE and the species urn:lsid:zoobank.org:act:971AD911-D5FB-460B-91AF-25B3D973873E were omitted.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hsiou, A.S., Nydam, R.L., Simões, T.R. et al. A New Clevosaurid from the Triassic (Carnian) of Brazil and the Rise of Sphenodontians in Gondwana. Sci Rep 9, 11821 (2019). https://doi.org/10.1038/s41598-019-48297-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-48297-9
This article is cited by
-
A new sphenodontian (Diapsida: Lepidosauria) from the Upper Triassic (Norian) of Germany and its implications for the mode of sphenodontian evolution
BMC Ecology and Evolution (2024)
-
An exceptionally preserved Sphenodon-like sphenodontian reveals deep time conservation of the tuatara skeleton and ontogeny
Communications Biology (2022)
-
A Triassic stem lepidosaur illuminates the origin of lizard-like reptiles
Nature (2021)
-
Sphenodontian phylogeny and the impact of model choice in Bayesian morphological clock estimates of divergence times and evolutionary rates
BMC Biology (2020)
-
Tuatara genome reveals diverse insights into a remarkable reptile
Nature (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
