
Abstract
Islets transplantation, as a treatment of type 1 diabetes, faces challenges, including the loss of islets in the process of isolation and pre-transplantation due to cellular stresses-induced apoptosis. Accordingly, the optimization of culture plays a decisive role in the transplantation success. In this study, we evaluated the effect of nobiletin on the cultured human islets. Isolated human islets were treated by different concentrations of nobiletin and cultured for 24 and 72 hours. Then, the islets viability, apoptosis, insulin and C-peptide secretion, and apoptosis markers were evaluated. Also, the production of reactive oxygen species (ROS), hypoxia inducible factor 1 alpha (HIF-1α), and its target genes in the islets were examined. Our findings showed that the islets were encountered with hypoxia and oxidative stress after isolation and during culture. These insults induced apoptosis and reduced viability during culture period. Moreover, the secretion of insulin and C-peptide decreased. Nobiletin treatments significantly improved the islets survival through reduction of HIF-1α and ROS production and suppression of apoptosis, along with increased islets function. Islet protective effect of nobiletin might be related to its anti-oxidant, anti-apoptotic and insulinotropic properties. Hence, in order to achieve viable and functional islets for clinical transplantation, the application of nobiletin during pre-transplantation period is useful.
Similar content being viewed by others
Introduction
Diabetes is a rising global health problem. It is estimated that more than 400 million people are afflicted with diabetes worldwide1. Type 1 diabetes involves about 5% to 10% of diabetes patients and is followed by a degenerative autoimmune reaction in β-cells of pancreatic Langerhans, leading to destruction of insulin production. Hence, type 1 diabetes patients need daily insulin injection to regulate the glycometabolic system, but few patients suffer from brittle diabetes and do not have an appropriate quality of life. On the other hand, insulin therapy does not prevent the complications of diabetes like retinopathy, nephropathy, and cardiovascular diseases especially in these group of patients2,3. Therefore, finding alternative approaches for treatment of type 1 diabetes patients is essential. Pancreatic islet transplantation could be an alternative and promising option for type 1 diabetes patient’s treatment. The first report of the islet transplantation in humans is related to the 1980s, which was further developed in the later years and has gained successful outcomes4. Nevertheless, there are several limitations in extensive application of islet transplantation.
Native islets typically possess a dense capillary network which prepares oxygen and nutrients in the islets, and β-cells consume large amounts of oxygen during insulin secretion. Accordingly, oxygen dependent β-cells make them very sensitive to hypoxic and oxidative conditions5. The major regulator of cellular stress responses is hypoxia inducible factor 1 alpha (HIF-1α). During hypoxia, HIF-1α activates a series of genes involved in cell adaptation pathways such as survival, angiogenesis, metabolism, and apoptosis6. Moreover, overproduction of reactive oxygen species (ROS), known as oxidative stress marker, produces under-hypoxic condition7,8,9. ROS and HIF-1α are also correlated together in stressful conditions, and ROS helps HIF-1α stabilization10.
It has been revealed that following ischemic injury of the pancreas due to disconnection from the whole body and during the isolation process, the islets encounter multiple stresses including oxidative stress and hypoxia, which continues during the pre-transplant culture period. These factors seem to be one of the main mediators in the activation of apoptosis, which can affect the islet transplantation outcome because of the low number of viable and functional islets11,12,13.
Since most transplant centers incubate the islets for 24 to 72 hours in culture media in order to control the quality of islets and preparation of recipients14,15, one of the critical steps in achieving a successful transplant is optimizing pre-transplant culture condition. A notable strategy to ameliorate the viable and functional islets is the use of cytoprotective agents in the culture period. Nobiletin (5, 6, 7, 8, 3′, 4′-hexamethoxyflavone) is a polymethoxylated flavonoid found in the citrus fruits and is used in traditional Chinese medicine. Nobiletin is considered by many researchers because of its widespread pharmacological properties such as anti-apoptotic, anti-inflammatory, anti-tumor, anti-oxidant, and anti-diabetes16,17. Moreover, several studies have represented that nobiletin has protective effects on the nervous system18, heart19,20, and kidney21. Findings have suggested that nobiletin protects the cells from apoptosis through activation of cyclic AMP-responsive element binding protein (CREB) and PI3/Akt pathways. These pathways promoted their target molecules including brain-derived neurotrophic factor (BDNF) and B-cell lymphoma 2 (Bcl-2), and inhibited Bcl-2-associated X protein (Bax) and caspase-316,21,22,23,24. It has also been reported that nobiletin reduces the oxidative stress-induced apoptosis via decreasing ROS and increasing the anti-oxidant enzyme activities21,25,26.
Therfore, it is logical to consider strategy to reduce stresses-induced cell death and increase the viability and functionalty of the isolated islets to maintain them in the culture preiod. In the present study, we have studied the effect of nobiletin on human pancreatic islets.
Results
Nobiletin ameliorated islet survival in culture
The evaluation of islets showed that the percentage of viable islets was nearly 100% in the treatment and control groups at 24 hours, while the viability rate was reduced to fewer than 50% in the control group during 72 hours. Treatment with nobiletin increased the viable islets, which was significant in 10 µM concentration (Fig. 1). Thus, the percentage of recovery was about 50–60%.

Islet viability evaluation was done by Fluorescein diacetate and propidium iodide staining in different concentrations of nobiletin. (A) Green cells were alive and red cells were dead. Scale bars = 100 and 200 μm. (B) The percentage of live cells was also calculated and shown as charts. The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. The results were represented as mean ± SD. **p < 0.01.
Nobiletin reduced islets apoptosis in culture
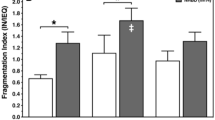
Apoptotic cells were detected by TUNEL assay based on DNA fragmentation within the islets. Reduction of apoptotic cells was revealed in the presence of nobiletin in different groups during 24 and 72 hours culture. However, this decrement was significant only in 10 μM nobiletin in comparison with the control group at 72 hours (Fig. 2).

Islet apoptosis evaluation with TUNEL assay in different concentrations of nobiletin. (A) The TUNEL positive cells are visible as green stained nuclei. Scale bar = 100 μm. (B) The percentage of TUNEL positive cells was also calculated and shown as graphs. The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. Results were represented as mean ± SD. *p < 0.05, and ***p < 0.001.
Nobiletin changed the anti-apoptotic and pro-apoptotic molecules expression in the cultured islets
To confirm the effect of nobiletin on the islets survival, we investigated Bcl-2, Bax, and active Caspase-3 protein expression during 24 and 72 hours by immunocytochemistry and the calculated H-scores were shown as histograms (Figs 3 and 4A–C). Bcl-2 as an anti-apoptotic molecule, Bax as pro-apoptotic molecule, and active caspase-3 as an enzyme in the final stages of apoptosis were measured. Nobiletin treatment in a dose-dependent manner increased Bcl-2 protein, while it decreased Bax and active caspase-3 protein during culture periods (Fig. 3). The expression of Bax protein significantly reduced in 10 µM concentration (Fig. 4A) and the Bcl-2 level significantly increased in 5 µM and 10 µM concentrations at 72 hours treatment (Fig. 4B). Active Caspase-3 had no significant changes in 24 hours compared to the control group; however, during 72 hours culture the active Caspase-3 was increased in the control group and nobiletin treatment dose-dependently reduced the active caspase-3 level, indicating the inhibition of apoptosis. Reduction of Caspase-3 was significant in 5 µM and 10 µM nobiletin concentrations (Fig. 4C).

Bax, Bcl-2, and active Caspase-3 immunocytochemistry in the islets in different concentrations of nobiletin (Yellow arrows show positive protein expression). Nobiletin decreased Bax, active Caspase-3 protein level, and increased Bcl-2 protein level, especially at 72 hours. The experiment was done in triplicate from four donors. Scale bar = 10 µm and 20 µm.

Pro-apoptotic and anti-apoptotic protein and gene expression. (A–C) Bax, Bcl-2, and active Caspase-3 protein in the islets in different concentrations of nobiletin that were calculated with H-score method, based on the percentage and intensity of the brown stained area in the islets. (D–F) Bax, Bcl-2, and Caspase-3 mRNA levels in different concentrations of nobiletin. The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. Results were represented as mean ± SD. *p < 0.05, **p < 0.01, and ***p < 0.001.
The gene expression evaluation of Bcl-2, Bax, and Caspase-3 indicated an up-regulated Bcl-2 mRNA and a down-regulation of Bax and caspase-3 mRNA during 24 and 72 hours culture in nobiletin treatment groups compared to the control group. The reduction of Bax was statistically significant in 5 µM and 10 µM, and enhancement of Bcl-2 was significant in 10 µM concentration (Fig. 4D,E). These results showed that nobiletin could rescue the islets from apoptosis and increase their viability.
Nobiletin increased C-peptide and insulin secretion in the cultured islets
Glucose stimulation C-peptide and insulin indexes showed no significant change in the control and nobiletin treatment groups at 24 hours. However, nobiletin treatment enhanced the C-peptide and insulin indexes compared to the control group during 72 hours incubation. This increase was significant in 10 µM concentration. The mentioned result indicated that nobiletin could improve the islet function in the culture. Furthermore, the evaluation of insulin mRNA showed that nobiletin increased the mRNA level of insulin. This increase correlated positively with up-regulation of insulin secretion. Increase of insulin mRNA level was significant in 5 µM and 10 µM of nobiletin (Fig. 5).

Glucose stimulation C-peptide and insulin indexes and insulin gene expression. (A) C-peptide and (B) insulin stimulation indexes and (C) insulin mRNA expression in different concentrations of nobiletin. The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. Results were represented as mean ± SD. *p < 0.05 and **p < 0.01.
Nobiletin changed HIF-1α gene and protein expression in the cultured islets
In order to evaluate the hypoxic stress in the cultured islet, we detected the expression of HIF-1α as the main hypoxic marker by immunocytochemistry. It was found that the HIF-1α was expressed in the cultured islets. There was no difference in HIF-1α protein level between the control and nobiletin treated groups at 24 hours, whereas nobiletin reduced HIF-1α protein in the islets during 72 hours culture. Reduction of HIF-1α was significant at 5 µM and 10 µM concentrations. The gene expression evaluation of HIF-1α did not apparently change in any groups in the culture periods, suggesting that HIF-1α regulation in islets may be regulated in post-transcriptional changes at the level of protein stability under hypoxia6,27,28 (Fig. 6).

HIF-1 α protein and gene expression. (A) HIF-1α protein expression in the different concentrations of nobiletin. The chart shows the H-score rate based on the percentage and intensity of the brown stained area in the islets (Yellow arrows show positive protein expression). (B) HIF-1α gene expression in different concentrations of nobiletin. The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. Results were represented as mean ± SD. *p < 0.05. Scale bar = 10 µm.
Nobiletin decreased p53 protein expression in the cultured islets
To demonstrate that hypoxic stress induces apoptosis, p53 protein expression was checked as a downstream molecule of HIF-1α. p53 was just expressed during 72 hours culture and nobiletin suppressed it in 5 µM and 10 µM concentrations (Fig. 7).

p53 protein expression (A) p53 protein expression by immunocytochemistry in the islets in different concentrations of nobiletin. Scale bar = 10 µm. (B) The chart shows the H-score rate based on the percentage and intensity of the brown stained area islets in different groups (Yellow arrows show positive protein expression). The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. Results were represented as mean ± SD. *p < 0.05 and **p < 0.01.
Nobiletin reduced ROS production in the cultured islets
Evaluation of ROS production in the islets revealed that nobiletin dose-dependently decreased it during 24 and 72 hours incubation. Diminution of ROS in the presence of nobiletin was statistically significant at 5 µM and 10 µM (Fig. 8).

ROS measurement in the islets in different concentrations of nobiletin. The chart shows ROS formation based on DCF fluorescent intensity. The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. Result were expressed as mean ± SD. *p < 0.05, **p < 0.01, and ***p < 0.001.
Nobiletin enhanced the VEGF secretion and gene expression in the cultured islets
After confirming the hypoxic stress introduced into the cultured islets, we measured VEGF as a downstream target gene of HIF-1α that was expressed in mild hypoxic condition to adopt the cells with low oxygen. Our results showed that VEGF increased in mRNA and protein secretion level in the islets probably following the HIF-1α expression. However, there was a reduction in the control group at 72 hours; nobiletin treatment dose-dependently enhanced the amount of mRNA and protein secretion level of VEGF. The increase was significant in 10 µM concentration. There was, however, no apparent change between the control and nobiletin treatment groups during 24 hours incubation (Fig. 9).

VEGF secretion and gene expression. (A) VEGF secretion and (B) VEGF mRNA expression in the islets in different concentrations of nobiletin. The experiment was done in triplicate from four donors and data were analyzed by Kruskal-Wallis and Post-hoc tests. Results were represented as mean ± SD. *p < 0.05 and **p < 0.01.
Discussion
Protection of the isolated islets and improvement of their function are necessary for successful transplantation. The widespread loss of islets during isolation and following that in the culture period is due to several insults including hypoxia and oxidative stress that can induce apoptosis. Therefore, the study of apoptotic induced mechanisms and strategies to block or diminish this process is essential for successful islet transplantation29.
In this study, we used nobiletin as a pharmacological approach to mitigate human pancreatic islet apoptosis and improve the viability and functionality of them in the culture. It has been reported that nobiletin exerts anti-apoptotic effects through suppression of intrinsic apoptotic pathway in acute kidney injury and neurodegenerative diseases21,22. Our result showed that nobiletin at 10 µM concentration significantly improved the islet survival. Furthermore, during 72 hours, nobiletin inhibited the Bax and caspase-3 and increased the Bcl-2 expression at both mRNA and protein levels. These findings indicated that the dead of islets through apoptosis reduced by nobiletin treatment.
Our observations were in agreement with other studies on nobiletin. It was shown that nobiletin improved the cisplatin-induced kidney injury through decreasing Bax and Caspase-3 and increasing Bcl-2 expression21. Similarly, it was reported that nobiletin protected against ischemic cerebral injury22 and improved isoflurane-induced cognitive impairment23 in rats via up-regulation of Bcl-2 expression and activation of Akt,CREB and BDNF pathways. Since the islets show some of the characteristics of neuronal phenotype due to having an endodermal lineage30, the suggestive protection mechanism of nobiletin may be through Akt/CREB pathway that should be investigated on the islets.
In the next step, we surveyed the effect of nobiletin on the β-cell function. It was found that nobiletin increased the insulin at both mRNA and protein secretion levels and C-peptide secretion at 72 hours. The anti-apoptotic and insulinotopic features of nobiletin on the human isolated islets are similar to glucagon-like peptide-1and prolactin31,32 where both these drugs reduced the number of apoptotic islets via inhibition of active Caspase-8, -9, and -3 and up-regulation of Bcl-2 in the cultured rat islets and improved the function of islets through increase in the insulin secretion31,32. In a recent report, nobiletin (10 µM) also inhibited β-cell apoptosis through activation of cyclic adenosine monophosphate (cAMP) pathway and downstream target protein kinase A (PKA) in INS-1D β-cell line and enhanced insulin secretion through another downstream target of cAMP called exchange proteins directly activated by cAMP (EPACs)24. Of note to this study, nobiletin may act via cAMP pathway in the isolated islets.
Islets are faced with hypoxia during the isolation and pre-transplant culture period because of the disconnection from the vascular network13. During short-term hypoxia, HIF-1α induces the expression of genes needed for metabolic adaptation including VEGF, thereby moving toward cell survival; however, in long-term hypoxia, over-aggregation of HIF-1α induces the expression of pro-apoptotic gene Bcl-2 interacting protein 3(BNIP3) or stabilizes the tumor suppressor gene p53, and moves toward apoptosis6,7,13,33.
Our observation showed that nobiletin (10 µM (significantly decreased the level of HIF-1α protein, scavenged ROS production, and suppressed p53 at 72 hours. The amount of VEGF mRNA and secretion also significantly increased in the presence of nobiletin. In the hypoxic response system, the VEGF expression is upregulated not only by HIF-1-mediated increase in gene transcription, but also via increased stability of VEGF mRNA, and increased VEGF protein export29,34,35. In our study, the HIF-1α was decreased by nobiletin at 72 hours; however, the increase in VEGF protein was remained. With this in mind, there could be other pathways involved in nobiletin induced increase in VEGF besides HIF-1α. In the current study, it was confirmed that the islets after isolation encountered hypoxia and oxidative stress that continued in the culture period. The stabilization of HIF-1α protein following hypoxia induced the expression of VEGF mRNA that could improve the islets survival during 24 hours culture because VEGF is an important growth factor for revascularization of the islets, especially after transplantation29. Our observations were similar to the effect of liraglutide on the isolated rat islets where this drug promoted the islet survival via enhancement of VEGF expression by activation of mammalian target of rapamycin (mTOR) pathway36, suggesting perhaps nobiletin was acting through this important pathway in angiogenesis.
At 72 hours culture, intracellular hypoxic in the islets led to chronic aggregation of HIF-1 α and stabilization of p53 that shifted the cells toward mitochondrial apoptosis pathway. It has been reported that the aggregation of HIF-1α protein is considered as an indicator of hypoxia that correlated with islet apoptosis6,29,37,38. According to our study, it seems that the duration of hypoxia and amount of HIF-1α protein affects the islet survival and death that would be confirmed with HIF-1α knockdown during different culture times.
Hypoxia also induces oxidative stress in the cells through overproduction of ROS8,10. Previous studies have shown that overproduction of ROS in the isolated islet can lead to β-cell death, due to weak intrinsic anti-oxidant defense system7. Moreover, overproduction of ROS influences cells through two pathways; one of them is the stabilization of HIF-1α and p53 and another one is disruption of the mitochondrial membrane and cytochrome C release with subsequent caspase 8, 9 and 3 activation39. It has been revealed that nobiletin plays an important role in apoptosis inhibition by restoring anti-oxidant activity and reducing the production of ROS in acute kidney injury and CNS ischemia21,22,23. Thus, nobiletin might reduce the islet apoptosis through its anti-oxidant properties. The anti-oxidant feature of nobiletin on the islets was similar to exendin-4 and glutathione ethyl ester where both these drugs decreased the islet apoptosis through inhibition of ROS overproduction7,40.
The clinical significance of this finding is that nobiletin was able to maintain the survival of islets during 72 hours. Indeed, nobiletin helped the islets to overcome the prolonged hypoxia and oxidative stresses in the culture and in this way reduced the islets death by more than 50% and increased the insulin production and secretion. The effect of nobiletin on the isolated human islets was mediated via anti-apoptotic, insulinotropic, and anti-oxidant properties (Fig. 10). However, it would be important to investigate the capacity of the treated islets with nobiletin in diabetic animal models. Nowadays, most transplant centers keep the islets in the culture media for 24–72 hours before transplantation that allows additional time to control the quality of the islets, islet transportation to other centers, patient preparation, coordination of transplantation team, and preoperative immunosuppressive drug administration14,15,41. Accordingly, nobiletin is suggested as an appropriate pharmacological substance for protection of the islets in the pre-transplant culture period that will result in a large number of islets for transplantation, thereby improving the transplant outcomes.

Schematic effect of nobiletin on human isolated islets in the culture. Nobiletin decreased hypoxia markers and oxidative stress induced apoptosis and improved islets survival and function.
Conclusions
Developing protective strategies in order to overcome stress-induced apoptosis in the isolated islets would be helpful and provide higher numbers of viable and functional islets in the pre-transplant culture period, ultimately leading to successful transplantation. Nobiletin, as a cytoprotective agent, can provide an opportunity to improve the survival and function of the human pancreatic islets through its anti-apoptotic, anti-oxidant, and insulinotropic properties.
Materials and Methods
Human islet isolation and in vitro experiment
Human islets were isolated based on the semi-automated protocol of Ricordi et al.42,with a few modifications43. After obtaining the written informed consent for research from dead brain donors’ relatives, four human pancreases (Table 1) were taken in accordance with the institutional ethics committee (IR.TUMS.REC.1394.1306). Briefly, the pancreatic duct was distended with collagenase NB1 and neutral protease (Serva, Germany). After mechanical and enzymatic digestion in a Ricordi chamber, the islets purification was performed in a continuous Biocoll (Biochrom, Germany) gradient in a cell sorter processor called COBE 2991. The counting and purity of the islet were assessed by dithizone staining and the number of islets was expressed in the islet equivalents (IEQ)44. The isolated islets with more than 80% purity were used to study. They were maintained in CMRL 1066 (Gibco, UK) medium containing 1% FBS (Gibco, UK), 1% antibiotic/antimycotic (Sigma, Germany), and 6.25 µg/ml ITS (Sigma, Germany) in 5% CO2 at 37 °C for a night. Then, 700 IEQ/well islets (75 IEQ/cm2) were treated with nobiletin (Sigma, Germany) in three concentrations of 0.5, 5, and 10 µM45,46,47 and incubated in 5% CO2 at 37 °C for 24 and 72 hours. The islets without nobiletin treatment were used as the control group. The experiments were analyzed in triplicate.
Islets viability evaluation
Viability of the islets was evaluated by live/dead fluorescent dyes method based on the combination of 5 mg/ml Fluorescein diacetate by live cells (Sigma, Germany) and 2 mg/ml propidium iodide by dead cells (Sigma, Germany). Images were captured under fluorescence microscope (Olympus, CKX53, Japan). The viability rate was calculated by the percentage of the fluorescein diacetate stained area to the total area.
Islets apoptosis detection
Apoptotic islets were determined by Terminal deoxynucleotidyl transferase–mediated dUTP nick end labeling (TUNEL) assay using Click-iT® Plus TUNEL Assay kit (Life technology, France) according to the manufacturer’s protocol. Nuclear counter staining was done with DAPI (Sigma, Germany). Images were captured under fluorescence microscope. The ratio between the apoptotic cells and nucleuses was used to calculate the percentage of apoptotic islets.
Gene expression evaluation
Total RNA was extracted in each islet groups by RNA-Sol isolation kit (Alphabio, Canada), based on the manufacturer’s protocol. Then, RNA was converted to cDNA with PrimeScript TM RT Reagent Kit (Takara, Japan). Primers were designed by Primer BLAST at NCBI and sequences were: human insulin (F: 5′-CTTCTACACACCCAAGACCC-3′; R: 5′-CTGGTACAGCATTGTTCCAC-3′), human Caspase-3 (F: 5′-ACTCCACAGCACCTGGTTATT-3′; R: 5′-TCTGTTGCCACCTTTCGGTT-3′), human Bax (F: 5′-TTCTGACGGCAACTTCAACT-3′; R: 5′-GGAGGAAGTCCAATGTCCAG-3′), human Bcl-2 (F: 5′-GATGGGATCGTTGCCTTATGC-3′; R: 5′-CAGTCTACTTCCTCTGTGATGTTGT-3′),human HIF-1α (F: 5′-GCAGCAACGACACAGAAACT-3′; R: 5′-TTCAGCGGTGGGTAATGGAG-3′), human vascular endothelial growth factor (VEGF) (F: 5′-CTTCAAGCCATCCTGTGTGC-3′; R: 5′-ATCCGCATAATCTGCATGGTG-3′) and human Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (F: 5′-GCTCATTTCCTGGTATGACAACG-3′; R: 5′-CTCTCTTCCTCTTGTGCTCTTG-3′), as a housekeeping gene. Relative gene expression was evaluated with real time RT-PCR by SYBR® Premix Ex TaqTM II kit (Takara, Japan) on an Applied Biosystems StepOnePlus™ System (ABI, USA). The fold changes were calculated by 2−ΔΔCT for each gene. GAPDH was recruited as the internal control.
Protein expression by immunocytochemistry assay
The islet groups were fixed in 4% paraformaldehyde and embedded in low melting agarose (Sigma, Germany). After dehydration process, the second embedding was performed in paraffin, cut in 5 μm sections, deparaffinised, and surveyed by immunocytochemistry. Immuno-labelling was done with primary antibodies specific to anti-human Bax (dilution 1:50; Abcam, France), anti-human Bcl-2 (dilution 1:250; Abcam, France), anti-human active Caspase-3 (dilution 1:50; Abcam, France), anti-human HIF-1α (dilution 1:50; Medaysis, USA), and anti-human p53 (dilution 1:50; Dako, Canada). All of these primary antibodies were detected by HRP- secondary antibody (dilution 1:200; Abcam, France). Finally, the samples were colored with a chromogen 3,3′- diaminobenzidine (DAB) (Dako, Canada) and counterstained with haematoxylin. The brown stained areas were scored based on the H-score method with following formula: H score = 1 × (% mild staining) + 2 × (% moderate staining) + 3 × (% strong staining)48.
Glucose-stimulated insulin and C-peptide secretion
After the nobiletin treatment period, the islets were washed with PBS and RPMI 1640 without glucose (Gibco, Germany) containing 0.5% BSA, and 2.8 or 20 mM glucose was then added, and incubated in 5% CO2 at 37 °C for 2 hours. Both supernatants were collected and stored at −80 °C. The secreted insulin and C-peptide49,50,51,52 were measured with ELISA kit (Monobind, USA). In each group, the stimulation index was calculated by dividing the amount of C-peptide and the insulin released from the islets in 20 mM glucose medium into the amount of C-peptide and the insulin released from the islets in 2.8 mM glucose medium in parallel and reported as the results40,43,53.
ROS measurement
Produced ROS was measured by a cell permeable and oxidation sensitive fluorescent dye 2′,7′-dichlorofluorescein diacetate (DCFH-DA) (Sigma, Germany)54 after the treatment period in all groups. The islets were suspended in serum-free RPMI 1640 medium and 10 μM DCFH-DA and incubated in darkness at 37 °C for 30 min. Then, the fluorescence intensity was monitored with a microplate reader (FLUOstar Omega®, BMG Labtech, Germany) with an excitation wavelength at 485 nm and an emission wavelength at 535 nm. The fluorescence intensity was normalized with total protein concentration measured by Bradford reagent at 630 nm and reported as the results54.
VEGF secretion assay
After the culture period, the islet supernatants were collected and stored in at −80 °C. VEGF secretion was detected with human VEGF ELISA kit (Life technology, France) according to the manufacturer’s instruction. The results were expressed as pg/ml.
Statistical analysis
The data were analyzed using SPSS version 24 (SPSS Inc., Chicago, IL), and the results were represented as mean ± S.D. We applied Kruskal-Wallis and Post-hoc tests to compare the groups and between the two groups, respectively. The graphs were drawn by Prism version 6 (Graph Pad Software, San Diego, California). Statistical significance was considered as P < 0.05.
Ethics approval
Ethics approval to conduct the study was obtained from the Shiraz and Tehran University of Medical Sciences Institutional Ethics Committee (IR.TUMS.REC.1394.1306).
Data Availability
All data generated in this study are included in the manuscript.
References
Organization, W. H (2016).
You, W.-P. & Henneberg, M. Type 1 diabetes prevalence increasing globally and regionally: the role of natural selection and life expectancy at birth. BMJ Open Diabetes Research and Care 4, e000161 (2016).
Bruni, A., Gala-Lopez, B., Pepper, A. R., Abualhassan, N. S. & Shapiro, A. J. Islet cell transplantation for the treatment of type 1 diabetes: recent advances and future challenges. Diabetes, metabolic syndrome and obesity: targets and therapy 7, 211 (2014).
Srinivasan, P., Huang, G., Amiel, S. & Heaton, N. Islet cell transplantation. Postgraduate medical journal 83, 224–229 (2007).
Cavallari, G. et al. Mesenchymal stem cells and islet cotransplantation in diabetic rats: improved islet graft revascularization and function by human adipose tissue-derived stem cells preconditioned with natural molecules. Cell transplantation 21, 2771–2781 (2012).
Greijer, A. & Van der Wall, E. The role of hypoxia inducible factor 1 (HIF-1) in hypoxia induced apoptosis. Journal of clinical pathology 57, 1009–1014 (2004).
Padmasekar, M. et al. Exendin-4 protects hypoxic islets from oxidative stress and improves islet transplantation outcome. Endocrinology 154, 1424–1433 (2013).
Pialoux, V. et al. Relationship between oxidative stress and HIF-1α mRNA during sustained hypoxia in humans. Free Radical Biology and Medicine 46, 321–326 (2009).
Guzy, R. D. & Schumacker, P. T. Oxygen sensing by mitochondria at complex III: the paradox of increased reactive oxygen species during hypoxia. Experimental physiology 91, 807–819 (2006).
Guzy, R. D. et al. Mitochondrial complex III is required for hypoxia-induced ROS production and cellular oxygen sensing. Cell metabolism 1, 401–408 (2005).
de Souza, B. M. et al. Effect of co-culture of mesenchymal stem/stromal cells with pancreatic islets on viability and function outcomes: a systematic review and meta-analysis. Islets 9, 30–42 (2017).
Wang, M., Crager, M. & Pugazhenthi, S. Modulation of apoptosis pathways by oxidative stress and autophagy in β cells. Experimental diabetes research 2012 (2012).
Velmurugan, K. et al. Antiapoptotic actions of exendin-4 against hypoxia and cytokines are augmented by CREB. Endocrinology 153, 1116–1128 (2012).
Toso, C. et al. Liraglutide, a long‐acting human glucagon‐like peptide 1 analogue, improves human islet survival in culture. Transplant International 23, 259–265 (2010).
Shapiro, A. J., Pokrywczynska, M. & Ricordi, C. Clinical pancreatic islet transplantation. Nature Reviews Endocrinology 13, 268 (2017).
Huang, H. et al. The multifunctional effects of nobiletin and its metabolites in vivo and in vitro. Evidence-Based Complementary and Alternative Medicine 2016 (2016).
Lee, Y.-S. et al. Nobiletin improves obesity and insulin resistance in high-fat diet-induced obese mice. The Journal of nutritional biochemistry 24, 156–162 (2013).
Braidy, N. et al. Neuroprotective effects of citrus fruit-derived flavonoids, nobiletin and tangeretin in Alzheimer’s and Parkinson’s disease. CNS & Neurological Disorders-Drug Targets (Formerly Current Drug Targets-CNS & Neurological Disorders) 16, 387–397 (2017).
Zhang, N. et al. Nobiletin, a polymethoxy flavonoid, protects against cardiac hypertrophy induced by pressure-overload via inhibition of napdh oxidases and endoplasmic reticulum stress. Cellular Physiology and Biochemistry 42, 1313–1325 (2017).
Wu, X. et al. Nobiletin attenuates adverse cardiac remodeling after acute myocardial infarction in rats via restoring autophagy flux. Biochemical and biophysical research communications 492, 262–268 (2017).
Malik, S. et al. Nobiletin ameliorates cisplatin-induced acute kidney injury due to its anti-oxidant, anti-inflammatory and anti-apoptotic effects. Experimental and Toxicologic Pathology 67, 427–433 (2015).
Zhang, L. et al. Nobiletin protects against cerebral ischemia via activating the p-Akt, p-CREB, BDNF and Bcl-2 pathway and ameliorating BBB permeability in rat. Brain research bulletin 96, 45–53 (2013).
Bi, J., Zhang, H., Lu, J. & Lei, W. Nobiletin ameliorates isoflurane-induced cognitive impairment via antioxidant, anti-inflammatory and anti-apoptotic effects in aging rats. Molecular medicine reports 14, 5408–5414 (2016).
Takii, M. et al. Insulinotropic and anti-apoptotic effects of nobiletin in INS-1D β-cells. Journal of Functional Foods 30, 8–15 (2017).
Qu, Y. et al. Nobiletin prevents cadmium-induced neuronal apoptosis by inhibiting reactive oxygen species and modulating JNK/ERK1/2 and Akt/mTOR networks in rats. Neurological research 40, 211–220 (2018).
Nakajima, A. et al. Nobiletin, a citrus flavonoid, improves cognitive impairment and reduces soluble Aβ levels in a triple transgenic mouse model of Alzheimer’s disease (3XTg-AD). Behavioural brain research 289, 69–77 (2015).
Uchida, T. et al. Prolonged hypoxia differentially regulates hypoxia-inducible factor (HIF)-1α and HIF-2α expression in lung epithelial cells Implication of Natural Antisense HIF-1α. Journal of Biological Chemistry 279, 14871–14878 (2004).
Kallio, P. J., Pongratz, I., Gradin, K., McGuire, J. & Poellinger, L. Activation of hypoxia-inducible factor 1α: posttranscriptional regulation and conformational change by recruitment of the Arnt transcription factor. Proceedings of the National Academy of Sciences 94, 5667–5672 (1997).
Miao, G. et al. Dynamic Production of Hypoxia‐Inducible Factor‐1α in Early Transplanted Islets. American journal of transplantation 6, 2636–2643 (2006).
Møller, C. J. et al. Differential expression of neural cell adhesion molecule and cadherins in pancreatic islets, glucagonomas, and insulinomas. Molecular Endocrinology 6, 1332–1342 (1992).
Farilla, L. et al. Glucagon-like peptide 1 inhibits cell apoptosis and improves glucose responsiveness of freshly isolated human islets. Endocrinology 144, 5149–5158 (2003).
Terra, L., Garay-Malpartida, M., Wailemann, R., Sogayar, M. & Labriola, L. Recombinant human prolactin promotes human beta cell survival via inhibition of extrinsic and intrinsic apoptosis pathways. Diabetologia 54, 1388–1397 (2011).
MORITZ, W. et al. Apoptosis in hypoxic human pancreatic islets correlates with HIF-1α expression. The FASEB Journal 16, 745–747 (2002).
Stein, I., Neeman, M., Shweiki, D., Itin, A. & Keshet, E. Stabilization of vascular endothelial growth factor mRNA by hypoxia and hypoglycemia and coregulation with other ischemia-induced genes. Molecular and cellular biology 15, 5363–5368 (1995).
Ozawa, K. et al. Expression of the oxygen-regulated protein ORP150 accelerates wound healing by modulating intracellular VEGF transport. The Journal of clinical investigation 108, 41–50 (2001).
Langlois, A. et al. In vitro and in vivo investigation of the angiogenic effects of liraglutide during islet transplantation. PloS one 11, e0147068 (2016).
Ma, Z. et al. Preconditioning with associated blocking of Ca2+ inflow alleviates hypoxia-induced damage to pancreatic β-cells. PloS one 8, e67498 (2013).
Zheng, X. et al. Acute hypoxia induces apoptosis of pancreatic β-cell by activation of the unfolded protein response and upregulation of CHOP. Cell death & disease 3, e322 (2012).
Redza-Dutordoir, M. & Averill-Bates, D. A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1863, 2977–2992 (2016).
do Amaral, A. S. R. et al. Glutathione ethyl ester supplementation during pancreatic islet isolation improves viability and transplant outcomes in a murine marginal islet mass model. PLoS One 8, e55288 (2013).
Froud, T. et al. Islet transplantation in type 1 diabetes mellitus using cultured islets and steroid‐free immunosuppression: Miami experience. American Journal of Transplantation 5, 2037–2046 (2005).
Ricordi, C., Lacy, P. E. & Scharp, D. W. Automated islet isolation from human pancreas. Diabetes 38, 140–142 (1989).
Kaviani, M. et al. Cytoprotective effects of olesoxime on isolated human pancreatic islets in order to attenuate apoptotic pathway. Biomedicine & Pharmacotherapy 112, 108674 (2019).
Azarpira, N. et al. Human islet cell isolation: the initial step in an islet transplanting program in Shiraz, Southern Iran. Exp Clin Transplant 12, 139–142 (2014).
Kim, H. J., Yonezawa, T., Teruya, T., Woo, J. T. & Cha, B. Y. Nobiletin, a polymethoxy flavonoid, reduced endothelin‐1 plus SCF‐induced pigmentation in human melanocytes. Photochemistry and photobiology 91, 379–386 (2015).
Guney Eskiler, G., Deveci, A. O., Bilir, C. & Kaleli, S. Synergistic Effects of Nobiletin and Sorafenib Combination on Metastatic Prostate Cancer Cells. Nutrition and cancer, 1–14 (2019).
Cui, Y. et al. Anti-neuroinflammatory activity of nobiletin on suppression of microglial activation. Biological and Pharmaceutical Bulletin 33, 1814–1821 (2010).
Smith, J. et al. Quantitative and qualitative characterization of Two PD-L1 clones: SP263 and E1L3N. Diagnostic pathology 11, 44 (2016).
Smirnov, A., Snigur, G. & Voronkova, M. In Apoptosis and medicine (IntechOpen, 2012).
Polonsky, K. et al. C-peptide and insulin secretion. Relationship between peripheral concentrations of C-peptide and insulin and their secretion rates in the dog. The Journal of clinical investigation 74, 1821–1829 (1984).
Landin-Olsson, M., Nilsson, K., Lernmark, Å. & Sundkvist, G. Islet cell antibodies and fasting C-peptide predict insulin requirement at diagnosis of diabetes mellitus. Diabetologia 33, 561–568 (1990).
Tatarkiewicz, K. et al. C-peptide responses after meal challenge in mice transplanted with microencapsulated rat islets. Diabetologia 44, 646–653 (2001).
Labriola, L. et al. Beneficial effects of prolactin and laminin on human pancreatic islet-cell cultures. Molecular and cellular endocrinology 263, 120–133 (2007).
Heidari, R. et al. Protective Effect of Glycine and Tri-Methyl Glycine (Betaine) Against Heavy Metals-Induced Oxidative Stress in Liver-Derived Post-Nuclear Supernatant (PNS). Trends in Pharmaceutical Sciences 4 (2018).
Acknowledgements
The authors are grateful to the Transplant Research Center at Shiraz University of Medical Sciences of Iran for financial supports and to Shiraz Organ Transplant Center for providing the pancreas. This work was supported by Tehran University of Medical Sciences (TUMS), Grant Number 94-04-87-30223, and conducted at the Transplant Research Center at Shiraz University of Medical Sciences. This research was a part of Ph.D. project number 9121436008 of TUMS.
Author information
Authors and Affiliations
Contributions
S.K., M.H.GH. and N.A. designed the study; S.K., M.K. and Z.J. performed experiments; M.H.GH., N.A., A.SH. and N.M. coordinated the study; S.K., M.K., and Z.J. analyzed data; S.K. wrote the manuscript; B.G., A.SH., N.M., S.N., E.M., I.H.A.A., M.H.GH. and N.A. interpreted data and revised the manuscript; M.H.GH. and N.A. did the latest revision of manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Keshtkar, S., Kaviani, M., Jabbarpour, Z. et al. Protective effect of nobiletin on isolated human islets survival and function against hypoxia and oxidative stress-induced apoptosis. Sci Rep 9, 11701 (2019). https://doi.org/10.1038/s41598-019-48262-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-48262-6
This article is cited by
-
Nobiletin is capable of regulating certain anti-cancer pathways in a colon cancer cell line
Naunyn-Schmiedeberg's Archives of Pharmacology (2023)
-
Development of a novel method for measuring tissue oxygen pressure to improve the hypoxic condition in subcutaneous islet transplantation
Scientific Reports (2022)
-
Potential Role of Polyphenolic Flavonoids as Senotherapeutic Agents in Degenerative Diseases and Geroprotection
Pharmaceutical Medicine (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
