
Abstract
Human milk delivers an array of bioactive components that safeguard infant growth and development and maintain healthy gut microbiota. Milk fat globule membrane (MFGM) is a biologically functional fraction of milk increasingly linked to beneficial outcomes in infants through protection from pathogens, modulation of the immune system and improved neurodevelopment. In the present study, we characterized the fecal microbiome and metabolome of infants fed a bovine MFGM supplemented experimental formula (EF) and compared to infants fed standard formula (SF) and a breast-fed reference group. The impact of MFGM on the fecal microbiome was moderate; however, the fecal metabolome of EF-fed infants showed a significant reduction of several metabolites including lactate, succinate, amino acids and their derivatives from that of infants fed SF. Introduction of weaning food with either human milk or infant formula reduces the distinct characteristics of breast-fed- or formula-fed- like infant fecal microbiome and metabolome profiles. Our findings support the hypothesis that higher levels of protein in infant formula and the lack of human milk oligosaccharides promote a shift toward amino acid fermentation in the gut. MFGM may play a role in shaping gut microbial activity and function.
Similar content being viewed by others

Introduction
Breastfeeding influences the development of the gut microbiota according to the degree of exclusivity1,2. This is at least partly due to the presence of antimicrobial proteins such as secretory immunoglobulin A, lactoferrin and lysozyme in human milk as well as oligosaccharides that function to selectively enhance colonization of specific groups of gut microbes including Bifidobacterium spp.3,4,5. Through selection of specific microbes, microbial richness and diversity are ultimately reduced2 as they influence the growth of other bacteria6,7, which results in modulation of the luminal pH and metabolic profile8,9.
Feeding infant formula has been shown to alter the fecal microbiota from that of breast-fed (BF) infants, induce a different serum metabolic profile than in BF infants10,11,12,13, and has been associated with increased risk of childhood obesity14,15. The difference in serum metabolic profiles may in part be due to differences in the intestinal microbiota, but are also likely due to the substantially higher protein concentration of infant formula as compared to human milk. Specifically, formula-fed (FF) infants have been shown to have higher levels of circulating amino acids, amino acid derivatives and urea than BF infants11,12.
There is a substantial quantity of nitrogenous compounds from dietary origin that escape digestion, and these compounds are retained in the lumen of the terminal ileum16 to provide substrates for microbial fermentation. Amino acids produced by intestinal microbiota are then absorbed and contribute to the host nitrogen pool17. Circulating lysine and threonine, that we previously found to be higher in FF infants13, are considered metabolites with a gut microbial origin18,19,20, as modification of intestinal microbiota via prebiotic supplementation has been shown to reduce free amino acids in cecum, colon, serum and liver of adult mice21, and to slightly lower the blood urea level of infants22. These studies support the importance of exploring gut microbiota as contributing factors to infant metabolism.
The milk fat globule membrane (MFGM) has historically been discarded with the milk fat in the manufacturing of bovine milk based infant formula. However, protective activity of MFGM against pathogens and viral infections has been shown23,24. Indeed, we found that providing MFGM to young infants in formula reduces the incidence of acute otitis media25, and to older infants as a supplement in weaning food reduces the prevalence of diarrhea26 and decreases circulating IL-227 as well as trimethylamine-N-oxide27, an oxidized microbial byproduct. These studies led to the hypothesis that MFGM may act as a substrate that selectively promotes growth of healthy infant gut microbiota, and that supplementing an MFGM concentrate may alter gut microbial composition and by-products to a profile that is more similar to an exclusively BF reference group.
To provide a mechanistic explanation regarding the influence of MFGM on gut microbiota, we evaluated the fecal microbiome and metabolome of a subgroup of 90 infants who participated in a prospective, double-blind, randomized controlled study28,29, where FF infants were randomly assigned to receive a standard infant formula (SF) or a bovine MFGM isolate-supplemented, low-energy, low-protein experimental formula (EF) from ~2 until 6 months of age. An exclusively BF reference group was included in this study. The influence of feeding on the composition and metabolic activity of intestinal microbiota was evaluated through secondary analyses of feces collected from these infants at baseline (~2 months), 4, 6 and 12 months of age via 16S rRNA amplicon sequencing and quantitative metabolomic profiling.
Results
Characteristics of study population and data exclusion
Out of a total of 240 infants who participated in a clinical trial concerning the outcomes of feeding a formula supplemented with a bovine MFGM isolate, which initially consisted of BF, n = 80; SF-fed, n = 80, and EF-fed, n = 80 (ClinicalTrials.gov identifier: NCT00624689), following exclusions28,29 a subset of 90 infants (15 males and 15 females from each treatment group) were randomly selected for fecal microbiome and metabolome analysis at 2, 4, 6, and 12 months. 96.2%, 96.5% and 82.1% of infants in the BF, SF, and EF group respectively were born vaginally. Inclusion criteria for the original study were: exclusively formula-fed at <2 months of age or intention to exclusively breastfeed until 6 months of age; gestational age at birth: 37–42 weeks; birth weight 2.5–4.5 kg and absence of chronic illness. After data generation, a few samples were further excluded, including infants who stopped consumption of study formula (n = 2), infants who had no record of a food diary (n = 1), infants from the BF group that were heavily mixed-fed (n = 3), infants from the FF group who consumed another formula (n = 4), and infants who had a record of antibiotics use (n = 3). For fecal metabolome data, samples that contained urea were suspected of being contaminated by urine since normal fecal samples do not contain urea due to bacterial urease activity. Therefore, fecal samples containing urea were excluded from analysis. The number of samples from each intervention and time point are summarized in SI Tables 1, 2.
Study formula intake and complementary food consumption
Our previous work on this cohort showed that infants consuming the low-energy, low-protein EF consumed a higher volume, yielding the same energy intake as infants consuming SF28. In this randomized subgroup, the intake of formula was only slightly higher in the EF group (effect sizes |δ| at each month were negligible or small, SI Fig. 1a). Although EF contains less energy than SF (SI Table 3), there was no significant difference in energy intake from consumption of these two formulas (Kruskal-Wallis’ H test, effect sizes |δ| at each month were negligible or small, SI Fig. 1c). Starting from 4 months, a variety of complementary foods were introduced (SI Fig. 1b) as suggested by the Swedish National Food Agency recommendation30. With increasing energy from complementary food, energy from study formula decreased between 4 and 6 months of age (SI Fig. 1c). By 6 months, fewer than 30 infants in this subgroup were still exclusively BF or fed study formula and had no exposure to complementary food (SI Fig. 1b,d). To account for the influence of complementary food introduction in our analysis, samples collected at 4 and 6 months were divided into two subgroups depending on complementary food intake and defined as: with (>10% daily energy from) or without (<10% daily energy from) complementary food.
Gut microbiome
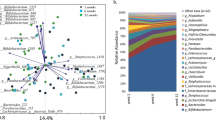
To explore the effect of early diet on intestinal microbiota, 16 s rRNA gene sequencing of infant feces was performed. Analysis of core microbes of all infants revealed a high prevalence of Bifidobacterium, Streptococcus, [Ruminococcus] (from Lachnospiraceae family), Veillonella, Enterococcus and Bacteroides from 2 to 6 months of age. Over time, as more weaning food was introduced, the relative abundance of Blautia in the stool increased (Fig. 1, SI Fig. 2). By 12 months of age, Bifidobacterium, Streptococcus and Bacteroides remained highly prevalent. With increases in microbial richness, additional microbes become highly prevalent in the gut (Fig. 1).

The core fecal microbiota of infants. The core microbiota is defined as highly prevalent microbes at the genus level present in >80% of samples.
As expected, higher levels of fecal Streptococcus was observed at 2 months of age (SI Fig. 2), which is likely due to bacterial communities arising from the environment, areolar skin and mother’s milk31,32,33. At 4 and 6 months of age when no or low complementary food was introduced, the impact of feeding on the fecal microbial profile was the largest (Fig. 2a), with higher abundance of Bifidobacterium species in BF compared with FF infants (Fig. 2c). BF infants had a more heterogeneous fecal microbiome profile only during the exclusive feeding period (Fig. 2b). By 12 months of age, the fecal microbiome became more homogenous (Fig. 2b) and the BF and FF fecal microbiomes became indistinguishable (Fig. 2a).

Analysis of the community structure of fecal microbiota reveals differences between breast-fed (BF, green) and formula-fed (Experimental Formula, EF, orange; Standard Formula, SF, red) infants at 2, 4 and 6 months of age. (a) Principal Coordinates Analysis (PCoA) of the log transformed weighted Unifrac distance. The centroids of each cluster (centroid of mass) are calculated as the average of PC1 and PC2 for each group. The ellipses were constructed based on multivariate normal distribution at 95% confidence level. The effect size (R2) and significance (p-value) between dietary groups were evaluated using permutational MANOVA via the Adonis test (permutation = 999). (b) Divergence (the spread within the group) was significantly higher in the BF infants during the exclusive feeding period. The measurement of divergence is calculated as 1- the average spearman correlation between samples and the overall group-wise average. The group difference was evaluated using a Kruskal-Wallis H test follow by the post-hoc Dunn test at p < 0.05. (c,d) Significantly differentiating intestinal microbes between the breast-fed (BF, green) and formula-fed (FF, pink) infants. The group differences were evaluated using Analysis of Composition of Microbiomes (ANCOM) followed by FDR correction at p < 0.05.
While fecal Bifidobacteria (Fig. 2c) and other lower abundant microbes (Fig. 2d) were significantly different between BF and FF, the difference between the two FF groups was minor. Some infants who consumed EF had a higher percentage of Akkermansia in the stool, and this was more prevalent when no or low complementary food was introduced (SI Fig. 3a). At 12 months, the number of individuals with Haemophilus was lower in infants who consumed EF compared with those who consumed SF (SI Fig. 3b, ANCOM test followed by FDR correction).
In comparison to BF infants, FF infants showed a slightly higher microbial richness (observed species) at 2 months (p = 0.077, Kruskal Wallis H test), but not statistically different (or close to statistically different) at other time points examined. When applying “Tail” statistics index (a more robust estimate of microbial diversity to account for low-abundance rare species34), no significant difference was detected between the two FF groups or between BF and FF infants, which may be due to the fact that some of the FF-infants were likely breast-fed after delivery for some time before enrolment at ~2 months of age.
Fecal metabolome
To evaluate the impact of early diet on intestinal microbial fermentation capability, corresponding fecal metabolites were examined. In agreement with prior studies35, BF infants in this cohort also had higher stool frequency and softer/looser stool consistency compared to FF infants25. In this subgroup, we observed significantly higher fecal water % and Bristol score in BF infants compared to the FF infants when infants were exclusively breast-fed or formula-fed respectively. However, after introduction of complementary food the water content in the stool of BF infants decreased to a level similar to both formula groups (Fig. 3a). Comparison of the water content in the stool of the formula groups revealed that infants consuming EF had less water in their stool than infants consuming SF (Fig. 3a). Since fecal water % was significantly different between groups, instead of normalizing fecal metabolite concentration by wet or dry fecal weight, values were expressed in µM.

(a) Fecal water % and Bristol score are significantly higher in the breast-fed (BF) infants during the exclusive feeding period. The group difference was evaluated using Kruskal-Wallis H test followed by a post-hoc Dunn test, p < 0.05. Fecal metabolite concentrations are associated with fecal water %. (b) Heatmap of the correlation coefficient (Pearson r > 0.4 or <−0.4, p < 0.05) of fecal metabolite concentrations correspond to fecal water %. Amount of water in infant stool is positively correlated with fecal concentration of (c) lactate, (d) 1,2-propanediol, (e) glucose and (f) galactose and is negatively correlated with (g) isovalerate and (h) isobutyrate. The correlation between fecal water % and metabolite concentration were evaluated using data collected from 2 months (baseline) and 4 and 6 months (without complementary food).
Fecal metabolite concentrations were associated with the amount of water in the stool. With increased water, higher concentrations of sugar monomers (glucose, galactose) and fermentation byproducts from carbohydrate metabolism (lactate, 1,2-propanediol, and succinate) were observed. In contrast, the concentration of microbial by-products from amino acid degradation (isobutyrate, isovalerate, dimethylamine, and phenylacetate) were associated with lower fecal water % (Fig. 3b–h).
Starting from 2 months of age, a profound difference in the fecal metabolome between BF and FF was observed and continued to 4 and 6 months when no or small amounts of complementary food were introduced. The effect of diet on the fecal metabolome was reduced in the group with complementary food consumption and became indistinguishable at 12 months of age (Fig. 4a, Adonis Test). In agreement with the fecal microbiome data (Fig. 2b), BF infants had a more heterogeneous fecal metabolome profile than FF infants, while the difference between the two FF groups was almost indistinguishable through Principal Coordinates Analysis (Fig. 4b).

The fecal metabolome reveals differences between breast-fed (BF, green) and formula-fed (Experimental Formula, EF, orange; Standard Formula, SF, red) infants at 2, and 4 & 6 months of age. (a) Principal Coordinates Analysis (PCoA) of the Euclidean distance matrix of the generalized log transformed fecal metabolite concentration data. The centroids of each cluster (centroid of mass) were calculated as the average PC1 and PC2 of all samples for each group. The ellipses were constructed based on multivariate normal distribution at 95% confidence level. The effect size (R2) and significance (p-value) between dietary groups were evaluated using permutational MANOVA via the Adonis test (permutation = 999). (b) Divergence (the spread within each group) is significantly higher in BF infants during the exclusive feeding period. The measurement of divergence was calculated as 1- the average spearman correlation between samples and the overall group-wise average. The group difference was evaluated using Kruskal-Wallis H test follow by post-hoc Dunn test, p < 0.05.
The major difference between the two FF groups was revealed during the exclusive feeding period, with lower levels of amino acids (ornithine, isoleucine, glutamate, phenylalanine, tyrosine, valine and glycine), and 2-hydroxyisovalerate (branched-chain amino acid degradation product), cadaverine (lysine breakdown product), and 4-hydroxyphenyllactate (tyrosine breakdown product) in the EF group (Fig. 5, Cliff’s Delta, Kruskal Wallis H test). Lactate and succinate, the two fecal metabolite markers that are positively correlated with amount of water in stool (Fig. 3b,c), were also lower in EF group (Fig. 5), and corresponded to the lower fecal water % (Fig. 3a). Intermediate metabolites from purine degradation metabolism, including hypoxanthine, were significantly lower, and inosine tended to be lower in the EF group. Although glycosylation of MFGM protein has been reported36, supplementing a bovine MFGM concentrate to the EF did not drive the fecal metabolome to the extent expected for human oligosaccharides (HMOs), thus the differences observed were likely derived from other active ingredients.

Fecal metabolites that are significantly different between formula-fed infants who consumed standard formula (SF, red) and experimental formula (EF, orange).
In general, acetate was the most dominant metabolite in the stool (SI Fig. 4). The appearance of larger and more diverse fecal sugar monomers are unique markers of breastfeeding, and include N-acetylglucosamine, fucose, galactose and glucose. In comparison, the major sugar monomers in the stool of FF infants were mostly galactose and glucose, sugar monomers from lactose. By 12 months of age, glucose was the major sugar monomer in infant stool. Only a few BF infants at 2 months of age had high levels of detectable HMOs in the stool, and the majority of these infants had lower concentrations of by-products from complex carbohydrate degradation (SI Fig. 4).
As breastfeeding continued, although more complementary food was introduced, the signature of breastfeeding persisted in the stool. BF infants exhibited higher levels of 4-hydroxyphenyllactate, fucose, lactate, 1,2-propanediol, pyruvate, and myo-inositol than the FF infants (Fig. 6a). In contrast, the FF-specific metabolic signature, included higher levels of butyrate, propionate, isobutyrate, isovalerate, phenylacetate and 4-hydroxyphenylacetate, which were no longer significant when more “carbohydrate-rich” complementary foods were introduced (Fig. 6b, Kruskal-Wallis H test). The complete list of differential metabolites between BF and FF stool is present in SI Fig. 5.

Significantly differentiating fecal metabolites that are (a) higher and (b) lower in breast-fed (BF, green) than formula-fed (FF, red) infants. The group differences were evaluated using Kruskal-Wallis H test by FDR correction at p < 0.05. The complete list of differentiating metabolites is in SI Fig. 5. Abbreviation: 4-HPLA: 4-hydroxyphenyllactate, 4-HPA: 4-hydroxyphenylacetate.
Discussion
Intestinal microbiota adapts and responds to the availability and distribution of fermentable substrates delivered from food. After escape from digestion, lactose and complex carbohydrates (including free HMOs, protein-derived and host-derived glycans) become substrates for gut microbial fermentation. Introduction of complementary food to either BF or FF infants disrupts the BF- or FF- specific fecal profile (Figs 2a, 4a). As weaning begins, the infant gut microbiota is exposed for the first time to a new range of nutrients, and as a result, inter-individual variation becomes less pronounced and the composition starts to resemble that of an adult-like microbiota. The abundance of Bifidobacterium, the predominant genus of the gut microbiota of BF infants, was no longer significantly higher than in FF infants following introduction of complementary food (Fig. 2c). However, before 6 months of age, if breastfeeding continued, complementary food introduction had a negligible effect on the key BF-specific fecal metabolite markers (Fig. 6a).
In this study, a bovine MFGM concentrate was supplemented to infant formula as an attempt to narrow the metabolic and microbial gap between BF and FF infants. Although some of the MFGM proteins are glycosylated36, which could provide a substrate for microbial fermentation, the amount of bovine MFGM supplemented in the current study is lower (approximately 0.48 g/L) compared to a recent study on rat pups (1.2 g/L) that showed a significant change in colon microbial composition and diversity with MFGM supplementation37. In comparison to the difference in the serum metabolome and other biochemical, clinical and cognitive measurements, the influence of MFGM supplementation on the overall oral microbiota38 and fecal microbiota (Figs 2a, 4a) was moderate and did not override the effect of formula, as infants who consumed MFGM-supplemented formula were more similar to those who consumed SF than those who were breast-fed. However, during the exclusive feeding period, lower concentrations of a few metabolites were observed in the EF in compared to the SF group (Fig. 5), providing a complementary view toward a potential functional change in the gut microbial community. Fecal hypoxanthine was lower in the EF-fed infants (Fig. 5), which is in line with the presence of hypoxanthine dehydrogenase/oxidase in bovine MFGM39. The reduced level of fecal hypoxanthine, together with the high abundance of Akkemansia in a few of the EF-fed infants (SI Fig. 3a), should be further confirmed in a larger clinical study. At 12 months of age, the incidence of having a high prevalence of fecal Haemophilus, a genus consisting of several pathogens, was lower in EF-fed infants (SI Fig. 3b). This observation suggests a potential long term impact of MFGM supplementation on the fecal microbiota that has not been reported elsewhere, and builds on previous studies in this cohort showing a lower incidence of acute otitis media in EF-fed infants at 6 months of age25. Further studies with larger sample sizes and targeted pathogen detection are needed to confirm these observations.
In comparison to formula-feeding, human milk consumption yielded higher levels of lactate, pyruvate, 1,2-propanediol, myo-inositol, fucose and 4-hydroxyphenyllactate in the stool (Fig. 6a). The large amounts of lactate in BF infant stool reported here and elsewhere40, are expected to be produced by Bifidobacterium, Bacteroides, Enterococcus and Streptococcus41,42 that are also primary colonizers of the infant gut (Fig. 1). However, in comparison to other bacterial genera, the metabolic capability of the species belonging to the Bifidobacteria genus is highly conserved with enriched saccharolytic modules in their genomes that are specialized for efficient HMO utilization43. Several Bifidobacteria subspecies (including B. longum subsp. infantis44, B. longum subsp. suis BSM11-545, B. kashiwanohense45, and B. bifidium46) harbor α-fucosidases and demonstrate the ability to produce 1,2-propanediol from fucose and fucosyllactose45. Bifidobacterium has been shown to produce a considerable amount of 4-hydroxyphenyllactate in vitro47, providing an explanation for the high 4-hydroxyphenyllactate found in the stool of BF infants.
While breastfeeding-derived fecal metabolic markers are associated with the utilization of carbohydrates, FF induces higher levels of branched chain fatty acids (BCFAs, isobutyrate and isovalerate) and phenylacetate (Fig. 6b), which are microbial end-products of peptide and amino acid fermentation. These have been shown to be elevated in in vitro incubation of intestinal content48,49, colon content from rodents, and in stool from individuals consuming high protein diets50,51,52. The higher fecal levels of propionate and butyrate in FF infants (Fig. 6b and elsewhere53,54) may be due to degradation of certain amino acids55 or production by species from the Eubacterium genus (Fig. 2d), which demonstrate an ability to utilize lactate and produce butyrate and propionate56,57.
In accordance with the higher protein intake provided by formula, stool ammonia has been reported to be significantly higher in exclusively FF infants compared to their BF counterparts58 which can be explained by excess dietary protein that escapes digestion59. This further implies that the intestinal microbiota in FF infants is likely to participate in significant catabolism of amino acids in the intestinal lumen. To an extent, the presence of ammonia reduces the uptake and utilization of short chain fatty acids (SCFAs) by colonocytes, and subsequently decreases the production of ketone bodies and CO2 50,60,61,62; therefore, the elevated butyrate level in the stool of FF infants (Fig. 6b) could also, in part, be due to reduced uptake by the colonocytes. Furthermore, ammonia can be easily absorbed by colonocytes, enter the circulation, and be detoxified to urea in the liver. This may contribute to the high urea concentration observed in the serum metabolome of FF infants13.
Infant formula contains considerably more protein than human milk63. A high protein content in the diet not only provides substrates for amino acid degradation, but may also induce protease activity in the colon50. In addition, proteases are more active in the neutral to alkaline pH range64. From the proximal to the distal colon, intestinal luminal pH65 and protease activity66,67 progressively increases as the amount of SCFA declines65,68. Gut microbial carbohydrate fermentation is also higher in the proximal colon69, whereas protein fermentation mostly takes place in the distal colon67. We speculate that carbohydrate fermentation in the proximal colon influences protein fermentation in the distal colon. When a substantial amount of HMOs reach the colon, rapid microbial fermentation is induced. As high total acids are produced from complex carbohydrate fermentation, intestinal bicarbonate buffering capacity is insufficient, and thus induces significant acidification of the lumen70,71. It has been shown that BF infants with greater utilization of HMOs exhibit a lower stool pH72, and rats consuming inulin alone or with Bifidobacterium longum have significantly reduced caecal pH and ammonia concentration73. This further supports the observation that acidic pH can decrease microbial uptake of most amino acids, reduce the rate of amino acid fermentation and subsequently decrease the net production of ammonia and BCFA48.
Since the amount of oligosaccharides is low in infant formula, the lack of complex carbohydrate substrates from the diet results in a higher stool pH in FF infants compared to BF infants40,54. In contrast, incorporating galacto-oligosaccharides and fructo-oligosaccharides in infant formula reduces the stool pH to a level more similar to BF infants53,54,74,75,76. Interestingly, none of the key metabolites that are different between BF and FF infants (Fig. 6) were observed in our previous work when comparing BF and FF rhesus monkeys77. Rhesus monkey milk contains lower oligosaccharide concentrations than human milk78,79. Therefore, the difference in fecal metabolome of BF and FF human infants observed here could be largely explained by the availability of complex carbohydrates in the diet for microbial utilization.
During the exclusive feeding period, FF infants had less water content in the stool (Fig. 3a). In addition, a harder stool and a longer intestinal transit time have been associated with carbohydrate deprivation and a more extensive proteinaceous substrate breakdown80,81. Dietary inclusion of fermentable complex carbohydrates has been shown to be a promising approach to reduce the rate of microbial protein fermentation82,83,84. Indeed, infants who consumed formula containing non-digestible oligosaccharides (fructo-oligosaccharides, galacto-oligosaccharides) had faster transit times and softer and more frequent stools22,74,75,76,85,86,87. Here, we observed that a watery stool is associated with an excess amount of sugar monomers and by-products from oligosaccharide fermentation (Fig. 3b–f). These microbial-derived products, if not utilized by the host or other microbes, may also act as organic osmolytes, drawing water from the epithelium into the lumen, leading to an even softer stool.
One strength of the present study is the double-blind randomized design among FF infants. Comparisons between FF and BF infants in the present study should be made with caution since these two groups were not randomized and potential confounding factors involving feeding choice cannot be ruled out. Another limitation of this study is the lack of information on the volume of human milk consumed and mother’s milk composition. It is well established that the composition of HMOs is driven by maternal secretor status and Lewis Blood group status88,89. Furthermore, by 12 months of age, only 6 infants in the BF group had a record of breast milk consumption. Although Bäckhed et al. have shown that the influence of breastfeeding on the gut microbiome remains at 12 months if breastfeeding persists1, we were not able to confirm this in the present study. Additionally, a large number of infants in all dietary groups consumed probiotics (containing Lactobacillus) during the study intervention period. Although the relative abundance of Lactobacillus in stool was low (<3%), we cannot rule out its potential contribution to the infant fecal metabolome90.
Conclusions
The results obtained in the present study clearly show the importance of early life nutrition on the community profile and functional characterization of intestinal microbiota. Breastmilk or formula-feeding as well as complementary food influence gut microbiota and the luminal environment. Building on previous work13,25,28,29,38,91 on this cohort, this study provides information on the effect of feeding on gut microbiota in the first year of life. The different fecal microbiomes are reflected by differences in fecal water % and concentration of microbial by-products in the stool. With limited oligosaccharides present and high protein levels in infant formula, we speculate that the difference observed between the BF and FF infant fecal microbiome and metabolome can be explained by an alteration in the availability and microbial utilization of carbohydrates and proteins as energy substrates. The difference between the fecal microbial taxonomic profiles of MFGM supplemented EF and SF was moderate in comparison to the distinct difference between BF and FF infants. However, an influence of MFGM on the fecal metabolome was observed, and may be associated with a change in microbial activity and function. Further studies are needed to explore the linkage between the potential antimicrobial property of MFGM proteins and the specific alteration we observed in the fecal metabolome.
Methods
Study population
This double-blinded, parallel randomized controlled trial took place at Umeå University Hospital, Umeå, Sweden after approval by the Regional Ethical Review Board in Umeå (Dnr 07-083 M) for all experimental protocols, and obtaining both oral and written informed consent from the parents/caregivers before inclusion. The study was registered at clinicaltrials.gov (NCT00624689, February 27, 2008), and was conducted in accordance with the Declaration of Helsinki and Good Clinical Practice guidelines.
Study formula
SF infants consumed BabySemp1 (Semper AB, Sundbyberg, Sweden) and EF infants consumed a formula modified from BabySemp1 with the addition of bovine MFGM-enriched whey protein concentrate (Lacprodan® MFGM-10; Arla Foods Ingredients, Viby J, Denmark) to account for 4% of the total protein in the EF. The macronutrient composition of the EF and SF has been described previously and also available in SI Table 3.
Estimation of nutrient intake
3-day food diary was recorded by parents every month over 3 consecutive days at 2, 3, 4, 5, 6 and 12 months of age. All food and drink that a child consumed were recorded either by weight (grams) or volume (milliliters, deciliters, tablespoons or teaspoons) by the parents using household measures and kitchen utensils. Taste portions were noted as if the intake volume was <15 mL. Brand name and manufacturer name were recorded if the food/drink consumed was a commercial product. The nutrient intake of each complementary food was calculated according to the Swedish National Food Agency Database and food labels. Use of medications, vitamins or dietary supplementations was documented. Persistence of breastfeeding was reported but the volume was not.
Stool collection procedure
Before or at the time of each visit, stool samples were collected by parents into a plastic container. The samples were immediately frozen at −80 °C or, if the sample was collected at home, frozen at −20 °C in a home freezer before transportation to the clinic and storage at −80 °C. The maximum storage time at −20 °C was 3 days.
Fecal metabolite extraction
After manual homogenization with a sterile microspatula, avoiding undigested food, approximately 250 mg of fecal material were weighted and then extracted by vigorously vortexing in 1.5 ml ice-cold Dulbecco’s phosphate buffered saline (DPBS, 1X, pH 7.4) followed by centrifugation. The pellet was collected, frozen at −80 °C, and subsequently lyophilized to determine fecal dry weight (Labconco FreeZone 4.5 L Freeze Dry System, Labconco, Kansas city, MO). The supernatants were carefully collected and sequentially filtered through a syringe filter with 0.22 µm pore size (Millex-GP syringe filter, Millipore, Billerica, MA) and an ultracentrifugal filter with a 3k Da molecular weight cut-off (Amicon ultra centrifugal filter, Millipore, Billerica, MA) to remove microbes and excess proteins. 23 µl of internal standard (DSS-d6 in 5 mM, with 0.2% sodium azide in 99.8% D2O) was added to 207 µL filtrate. The pH of each sample was adjusted to 6.8 ± 0.1 by adding small amounts of NaOH and HCl to minimize pH-based peak movement. 180 µL aliquots were subsequently transferred to 3 mm Bruker NMR tubes (Bruker, Brillerica, MA) and stored at 4 °C until spectral acquisition.
NMR acquisition, data processing and quantification
1H NMR spectra were acquired at 298K using the NOESY 1H presaturation experiment (‘noesypr1d’) on Bruker Avance 600 MHz NMR spectrometer (Bruker BioSpin, Germany). Spectral acquisition and processing parameters are identical to our previous work from this cohort13. Spectra were manually processed and profiled using Chenomx NMR Suite v8.3 (Chenomx Inc, Edmonton, Alberta, Canada). The concentration of fecal metabolites were estimated as:
where fecal water estimate is wet weight (mg) − dry weight (mg).
Stool water % and bristol score
Stool water % was calculated as (wet − dry)/wet stool weight * 100. Bristol stool score92, a scale from category 1 to 7 that indicates from hard to watery stool, was documented by one laboratory technician to ensure consistency.
Fecal microbial DNA extraction and library preparation
To minimize the freeze-thaw cycle, the fecal metabolite and microbial DNA were extracted on the same day for each sample. Approximately 250 mg of fecal material was extracted according to the HMP protocol using MoBio PowerLyzer PowerSoil DNA isolation kit (MoBio, Carlsbad, CA)93. The HMP protocol was modified from the manufacturer’s instruction as follows: (1) samples were heated at 65 °C for 10 min and then 95 °C for 10 min after adding C1 solution; (2) bead-beating of the samples was performed at 6.5 m/s for 1 min followed by 5 min rest, and bead-beating again for 1 min (FastPrep-24 bead beater, MP Bio, Solon OH); and (3) after bead beating, samples were centrifuged for 3 min to ensure pellets fully formed. The DNA purity was determined spectrophotometrically (NanoDrop 2000C Spectrophotometer, Thermo Fisher Scientific, Waltham, MA, USA).
The V4 region of the 16S rRNA gene was targeted using F515/R806 primers94 modified according to Bokulich et al.95 to contain an illumina adapter sequence on the forward primer. An 8 bp Hamming error-correcting barcode was attached to the 3′ end of the forward primer to enable sample multiplexing. PCR reactions were performed in 20 µL reaction volumes following the protocol developed by Gohl et al.96. PCR reactions contained DNA template, DMSO (Fisher Scientific, Waltham, MA), 5X KAPA HiFi Buffer (KAPA Biosystems, Woburn, MA), dNTP Mix (10 mM), KAPA HiFi HotStart Polymerase (KAPA Biosystems, Woburn, MA), PCR-grade water (MoBio, Carlsbad, CA), DNA template and primers. The ideal volume of DNA template in the PCR reaction was estimated using a dilution series experiment followed by a quality check via gel electrophoresis of the amplicon product. The PCR reaction consisted of an initial 95 °C for 5 min followed by a 25-cycle program of 20 s at 98 °C for denaturation, 15 s at 55 °C for annealing, 60 s at 72 °C for primer extension and a final extension of 72 °C for 10 min. All reactions were amplified in duplicate and combined prior to purification.
All amplified PCR products were quality checked by gel electrophoresis. The band intensity (around 380 bp) was visualized using SYBR safe DNA stain (Invitrogen) and its quantity (in ng/µL) was estimated using a molecular ladder with known concentration (BioRad EZ ladder 1 kb) through ImageLab software (v5.2.1, BioRad, Hercules, CA). Amplicons were pooled at equimolar ratio and purified using the QIAquick PCR Purification Kit (QIAGEN, Hilden, Germany) using a modified protocol from the manufacturer’s instruction. To maximize DNA yield and purity in the final library, amplicons were first combined into 12 separate libraries (not exceeding the column DNA capacity) and each were cleaned according to the manufacturer’s instruction. The final elusion was made by eluting the first column with 60 µL of elusion buffer and all subsequent column were eluted from the filtrate from the previous column. A Purified amplicon library was quality checked by Bioanalyzer and submitted to the UC Davis Genome Center DNA Technologies Core for 250 bp paired-end sequencing on the Illumina MiSeq platform.
To avoid airborne contamination during extraction as described in Salter et al.97, the DNA extraction was performed using PCR clean and sterile filter pipette tips and in a cleaned biosafety cabinet sanitized by UV for at least 30 min prior to the experiment followed by surface decontamination by DNAaway (Thermo Scientific). One negative control “blank sample” from extraction of PCR-grade water (MoBio, Carlsbad, CA) was included for each batch. This batch sample was subsequently amplified via PCR, visually inspected using gel electrophoresis and included in the final DNA library.
Analysis of 16 s amplicon sequence
A sliding window trimming of 4 bases wide on the low quality bases (quality score <20) was performed using Trimmomatic98 (version 0.33) at the end of both forward and reverse sequences. About 0.5% poor quality sequences were excluded. The subsequent sequence data was analyzed using the Quantitative Insights Into Microbial Ecology (QIIME) platform99 (version 1.9.0). Forward and reverse reads were joined using Fastq-join100 via join_paired_end.py where at least 50 bp of overlapping was required and the maximum difference was below 5%. About 74.16% of the sequences remained after joining paired end reads. The subsequent joined sequences were preprocessed and demultiplexed according to the following criteria: (1) removal of primer sequences; (2) minimum acceptable Phred quality score Q > 20: and (3) no barcode mismatch allowed. Closed OTU picking at 97% identity was done using sortmerna101 against the greengenes database (greengenes 13_8)102. After removing singletons (0.011% of total sequences), the OTU table was normalized by the copy number using normalize_by_copy_number.py function developed in PICRUSt (version 1.1.3)103.
Statistical analysis
Statistical computing and graphical generation were performed using the R programing environment. OTU table handling was achieved using phyloseq and the microbiome package. α-diversity and β-diversity distance matrices were computed using DiversitySeq and the vegan package, respectively. Generalized log transformation (defined as log(y + sqrt(y2 + lambda))) was applied to all metabolomics data where lambda is 1. Principal coordinate analysis was computed using the pcoa function from the ape package. Adonis, a nonparametric implementation of a permutational analysis of variance, was computed using the adonis function from vegan with permutation set at 999. The spread (homogeneity) within a group was determined as 1 - cor where cor is the average spearman correlation between samples and the overall group-wise average. This feature is implemented in the divergence function in the microbiome package.
The differential analysis for the microbiome was computed at the genus level using ANCOM104 and controls for False Discovery Rate (FDR). A pseudo count value of 0.001 was added for the relative abundance data. For other data, significance between groups was evaluated using the Kruskal-Wallis test (kruskal.test function) and p-values from the Kruskall-Wallis test were further adjusted by FDR (p.adjust(, method = ‘fdr’)). The overall level of significance was set at p < 0.05.
Effect size between BF and FF and between SF and EF was evaluated using Cliff’s delta (δ) statistics using the cliff.delta function from effsize package. The 95% confidence interval of each computed Cliff’s delta was estimated assuming a normal distribution (use.normal = TRUE). The magnitude was assessed using the thresholds suggested by Romano et al.105, where |δ| < 0.147 corresponds to negligible, |δ| < 0.33 corresponds to small, |δ| < 0.474 corresponds to medium, otherwise large.
The square of Pearson’s correction coefficient (r2) was computed using cor, (method = “pearson”) to evaluate the strength of the correlation. All plots were generated using ggplot2.
Data Availability
16S sequencing data from this study was deposited in the European Nucleotide Archive (accession code ERP112481) and Qiita106 (study ID: 12021). NMR spectra and relevant metadata are available from the corresponding author on request.
References
Bäckhed, F. et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host & Microbe 17, 690–703 (2015).
Forbes, J. D. et al. Association of exposure to formula in the hospital and subsequent infant feeding practices with gut microbiota and risk of overweight in the first year of life. JAMA Pediatr. 172, e181161 (2018).
Bode, L. Human milk oligosaccharides: prebiotics and beyond. Nutr. Rev. 67(Suppl 2), S183–191 (2009).
Smilowitz, J. T., Lebrilla, C. B., Mills, D. A., German, J. B. & Freeman, S. L. Breast milk oligosaccharides: structure-function relationships in the neonate. Annu. Rev. Nutr. 34, 143–169 (2014).
Demmelmair, H., Prell, C., Timby, N. & Lönnerdal, B. Benefits of lactoferrin, osteopontin and milk fat globule membranes for infants. Nutrients 9, 817 (2017).
Rivière, A., Selak, M., Lantin, D., Leroy, F. & De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: importance and strategies for their stimulation in the human gut. Front. Microbiol. 7, 979 (2016).
Gibson, G. R. & Wang, X. Regulatory effects of bifidobacteria on the growth of other colonic bacteria. J. Appl. Bacteriol. 77, 412–420 (1994).
Mohan, R. et al. Effects of Bifidobacterium lactis Bb12 supplementation on body weight, fecal pH, acetate, lactate, calprotectin, and IgA in preterm infants. Pediatr. Res. 64, 418–422 (2008).
Sugahara, H. et al. Probiotic Bifidobacterium longum alters gut luminal metabolism through modification of the gut microbial community. Sci. Rep. 5, 13548 (2015).
Slupsky, C. M. et al. Postprandial metabolic response of breast-fed infants and infants fed lactose-free vs regular infant formula: A randomized controlled trial. Sci. Rep. 7, 3640 (2017).
Kirchberg, F. F. et al. Dietary protein intake affects amino acid and acylcarnitine metabolism in infants aged 6 months. J. Clin. Endocrinol. Metab. 100, 149–158 (2015).
Socha, P. et al. Milk protein intake, the metabolic-endocrine response, and growth in infancy: data from a randomized clinical trial. Am. J. Clin. Nutr. 94, 1776S–1784S (2011).
He, X. et al. Metabolic phenotype of breast-fed infants, and infants fed standard formula or bovine MFGM supplemented formula: a randomized controlled trial Sci. Rep. 9, 339 (2019).
Weber, M. et al. Lower protein content in infant formula reduces BMI and obesity risk at school age: follow-up of a randomized trial. Am. J. Clin. Nutr. 99, 1041–1051 (2014).
Koletzko, B. et al. Can infant feeding choices modulate later obesity risk? Am. J. Clin. Nutr. 89, 1502S–1508S (2009).
Gaudichon, C. et al. Ileal losses of nitrogen and amino acids in humans and their importance to the assessment of amino acid requirements. Gastroenterology 123, 50–59 (2002).
Davila, A.-M. et al. Intestinal luminal nitrogen metabolism: role of the gut microbiota and consequences for the host. Pharmacol. Res. 68, 95–107 (2013).
Metges, C. C. et al. Availability of intestinal microbial lysine for whole body lysine homeostasis in human subjects. Am. J. Physiol. 277, E597–607 (1999).
Metges, C. C. et al. Incorporation of urea and ammonia nitrogen into ileal and fecal microbial proteins and plasma free amino acids in normal men and ileostomates. Am. J. Clin. Nutr. 70, 1046–1058 (1999).
Torrallardona, D., Harris, C. I., Coates, M. E. & Fuller, M. F. Microbial amino acid synthesis and utilization in rats: incorporation of 15N from 15NH4Cl into lysine in the tissues of germ-free and conventional rats. Br. J. Nutr. 76, 689–700 (1996).
Barouei, J. et al. Microbiota, metabolome, and immune alterations in obese mice fed a high-fat diet containing type 2 resistant starch. Mol. Nutr. Food Res. 61 (2017).
Closa-Monasterolo, R. et al. Safety and efficacy of inulin and oligofructose supplementation in infant formula: Results from a randomized clinical trial. Clin. Nutr. 32, 918–927 (2013).
Fuller, K. L., Kuhlenschmidt, T. B., Kuhlenschmidt, M. S., Jiménez-Flores, R. & Donovan, S. M. Milk fat globule membrane isolated from buttermilk or whey cream and their lipid components inhibit infectivity of rotavirus in vitro. J. Dairy Sci. 96, 3488–3497 (2013).
Sprong, R. C., Hulstein, M. F. E. & van der Meer, R. Bovine milk fat components inhibit food-borne pathogens. Int. Dairy. J. 12, 209–215 (2002).
Timby, N. et al. Infections in infants fed formula supplemented with bovine milk fat globule membranes. J. Pediatr. Gastroenterol. Nutr. 60, 384–389 (2015).
Zavaleta, N. et al. Efficacy of an MFGM-enriched complementary food in diarrhea, anemia, and micronutrient status in infants. J. Pediatr. Gastroenterol. Nutr. 53, 561–568 (2011).
Lee, H., Zavaleta, N., Chen, S.-Y., Lönnerdal, B. & Slupsky, C. Effect of bovine milk fat globule membranes as a complementary food on the serum metabolome and immune markers of 6-11-month-old Peruvian infants. npj Science of Food 2 (2018).
Timby, N., Domellöf, E., Hernell, O., Lönnerdal, B. & Domellöf, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: a randomized controlled trial. Am. J. Clin. Nutr. 99, 860–868 (2014).
Timby, N., Lönnerdal, B., Hernell, O. & Domellöf, M. Cardiovascular risk markers until 12 mo of age in infants fed a formula supplemented with bovine milk fat globule membranes. Pediatr. Res. 76, 394–400 (2014).
Good food for infants under one year. National Food Agency, Sweden Available at, https://www.livsmedelsverket.se/globalassets/publikationsdatabas/andra-sprak/good-food-for-infants-under-one-year-livsmedelsverket2.pdf.
Ferretti, P. et al. Mother-to-infant microbial transmission from different body sites shapes the developing infant gut microbiome. Cell Host Microbe 24, 133–145.e5 (2018).
Hunt, K. M. et al. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS One 6, e21313 (2011).
Madan, J. C. et al. Association of cesarean delivery and formula supplementation with the intestinal microbiome of 6-week-old infants. JAMA Pediatr. 170, 212 (2016).
Li, K., Bihan, M., Yooseph, S. & Methé, B. A. Analyses of the microbial diversity across the human microbiome. PLoS One 7, e32118 (2012).
Singhal, A. et al. Dietary nucleotides and fecal microbiota in formula-fed infants: a randomized controlled trial. Am. J. Clin. Nutr. 87, 1785–1792 (2008).
O’Riordan, N., Kane, M., Joshi, L. & Hickey, R. M. Structural and functional characteristics of bovine milk protein glycosylation. Glycobiology 24, 220–236 (2014).
Bhinder, G. et al. Milk Fat Globule membrane supplementation in formula modulates the neonatal gut microbiome and normalizes intestinal development. Sci. Rep. 7, 45274 (2017).
Timby, N. et al. Oral microbiota in infants fed a formula supplemented with bovine milk fat globule membranes - a randomized controlled trial. Plos One 12, e0169831 (2017).
Le, T. T. et al. Distribution and isolation of milk fat globule membrane proteins during dairy processing as revealed by proteomic analysis. Int. Dairy. J. 32, 110–120 (2013).
Ogawa, K., Ben, R. A., Pons, S., de Paolo, M. I. & Bustos Fernández, L. Volatile fatty acids, lactic acid, and pH in the stools of breast-fed and bottle-fed infants. J. Pediatr. Gastroenterol. Nutr. 15, 248–252 (1992).
Heinken, A., Sahoo, S., Fleming, R. M. T. & Thiele, I. Systems-level characterization of a host-microbe metabolic symbiosis in the mammalian gut. Gut Microbes 4, 28–40 (2013).
Vaughan, E. E., Heilig, H. G. H. J., Ben-Amor, K. & de Vos, W. M. Diversity, vitality and activities of intestinal lactic acid bacteria and bifidobacteria assessed by molecular approaches. FEMS Microbiology Reviews 29, 477–490 (2005).
Vieira-Silva, S. et al. Species-function relationships shape ecological properties of the human gut microbiome. Nat. Microbiol. 1, 16088 (2016).
Sela, D. A. et al. The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc. Natl. Acad. Sci. USA 105, 18964–18969 (2008).
Bunesova, V., Lacroix, C. & Schwab, C. Fucosyllactose and L-fucose utilization of infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol. 16, 248 (2016).
Ashida, H. et al. Two distinct alpha-L-fucosidases from Bifidobacterium bifidum are essential for the utilization of fucosylated milk oligosaccharides and glycoconjugates. Glycobiology 19, 1010–1017 (2009).
Beloborodova, N. et al. Effect of phenolic acids of microbial origin on production of reactive oxygen species in mitochondria and neutrophils. J. Biomed. Sci. 19, 89 (2012).
Smith, E. A. & Macfarlane, G. T. Enumeration of amino acid fermenting bacteria in the human large intestine: effects of pH and starch on peptide metabolism and dissimilation of amino acids. FEMS Microbiol. Ecol. 25, 355–368 (1998).
Smith, E. A. & Macfarlane, G. T. Enumeration of human colonic bacteria producing phenolic and indolic compounds: effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Bacteriol. 81, 288–302 (1996).
Andriamihaja, M. et al. Colon luminal content and epithelial cell morphology are markedly modified in rats fed with a high-protein diet. Am. J. Physiol. Gastrointest. Liver Physiol. 299, G1030–1037 (2010).
David, L. A. et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 505, 559–563 (2014).
Russell, W. R. et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am. J. Clin. Nutr. 93, 1062–1072 (2011).
Holscher, H. D. et al. Effects of prebiotic-containing infant formula on gastrointestinal tolerance and fecal microbiota in a randomized controlled trial. JPEN J Parenter. Enteral. Nutr. 36, 95S–105S (2012).
Knol, J. et al. Colon microflora in infants fed formula with galacto- and fructo-oligosaccharides: more like breast-fed infants. J. Pediatr. Gastroenterol. Nutr. 40, 36–42 (2005).
Smith, E. A. & Macfarlane, G. T. Dissimilatory amino acid metabolism in human colonic bacteria. Anaerobe 3, 327–337 (1997).
Duncan, S. H., Louis, P. & Flint, H. J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 70, 5810–5817 (2004).
Schwab, C. et al. Trophic interactions of infant Bifidobacteria and Eubacterium hallii during L-fucose and fucosyllactose degradation. Front. Microbiol. 8 (2017).
Heavey, P. M. et al. Protein-degradation products and bacterial enzyme activities in faeces of breast-fed and formula-fed infants. Br. J. Nutr. 89, 509 (2003).
Pieper, R., Boudry, C., Bindelle, J., Vahjen, W. & Zentek, J. Interaction between dietary protein content and the source of carbohydrates along the gastrointestinal tract of weaned piglets. Arch. Anim. Nutr. 68, 263–280 (2014).
Cremin, J. D., Fitch, M. D. & Fleming, S. E. Glucose alleviates ammonia-induced inhibition of short-chain fatty acid metabolism in rat colonic epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 285, G105–G114 (2003).
Darcy-Vrillon, B., Cherbuy, C., Morel, M. T., Durand, M. & Duée, P. H. Short chain fatty acid and glucose metabolism in isolated pig colonocytes: modulation by NH4+. Mol. Cell. Biochem. 156, 145–151 (1996).
Villodre Tudela, C. et al. Down-regulation of monocarboxylate transporter 1 (MCT1) gene expression in the colon of piglets is linked to bacterial protein fermentation and pro-inflammatory cytokine-mediated signalling. Br. J. Nutr. 113, 610–617 (2015).
Martin, C. R., Ling, P.-R. & Blackburn, G. L. Review of infant feeding: key features of breast milk and infant formula. Nutrients 8 (2016).
Macfarlane, G. T., Allison, C. & Gibson, G. R. Effect of pH on protease activities in the large intestine. Lett. Appl. Microbiol. 7, 161–164 (1988).
Cummings, J. H. & Macfarlane, G. T. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 70, 443–459 (1991).
Evans, D. F. et al. Measurement of gastrointestinal pH profiles in normal ambulant human subjects. Gut 29, 1035–1041 (1988).
Macfarlane, G. T., Gibson, G. R. & Cummings, J. H. Comparison of fermentation reactions in different regions of the human colon. J. Appl. Bacteriol. 72, 57–64 (1992).
Wong, J. M. W., de Souza, R., Kendall, C. W. C., Emam, A. & Jenkins, D. J. A. Colonic health: fermentation and short chain fatty acids. J. Clin. Gastroenterol. 40, 235–243 (2006).
van Beers-Schreurs, H. M. et al. Weaning piglets, microbial fermentation, short chain fatty acids and diarrhoea. Vet Q 20(Suppl 3), S64–68 (1998).
Hasselblatt, P., Warth, R., Schulz-Baldes, A., Greger, R. & Bleich, M. pH regulation in isolated in vitro perfused rat colonic crypts. Pflugers Arch. 441, 118–124 (2000).
Ilhan, Z. E., Marcus, A. K., Kang, D.-W., Rittmann, B. E. & Krajmalnik-Brown, R. pH-mediated microbial and metabolic interactions in fecal enrichment cultures. mSphere 2, e00047-17 (2017).
Frese, S. A. et al. Persistence of supplemented Bifidobacterium longum subsp. infantis EVC001 in breastfed infants. mSphere 2, e00501-17 (2017).
Rowland, I. R., Rumney, C. J., Coutts, J. T. & Lievense, L. C. Effect of Bifidobacterium longum and inulin on gut bacterial metabolism and carcinogen-induced aberrant crypt foci in rats. Carcinogenesis 19, 281–285 (1998).
Mihatsch, W. A., Hoegel, J. & Pohlandt, F. Prebiotic oligosaccharides reduce stool viscosity and accelerate gastrointestinal transport in preterm infants. Acta Paediatr. 95, 843–848 (2006).
Moro, G. et al. Dosage-related bifidogenic effects of galacto- and fructooligosaccharides in formula-fed term infants. J. Pediatr. Gastroenterol. Nutr. 34, 291–295 (2002).
Sierra, C. et al. Prebiotic effect during the first year of life in healthy infants fed formula containing GOS as the only prebiotic: a multicentre, randomised, double-blind and placebo-controlled trial. Eur. J. Nutr. 54, 89–99 (2015).
He, X., Slupsky, C. M., Dekker, J. W., Haggarty, N. W. & Lönnerdal, B. Integrated Role of Bifidobacterium animalis subsp. lactis Supplementation in Gut Microbiota, Immunity, and Metabolism of Infant Rhesus Monkeys. mSystems 1, e00128-16 (2016).
Tao, N. et al. Evolutionary glycomics: characterization of milk oligosaccharides in primates. J. Proteome Res. 10, 1548–1557 (2011).
O’Sullivan, A. et al. Metabolomic phenotyping validates the infant rhesus monkey as a model of human infant metabolism. J. Pediatr. Gastroenterol. Nutr. 56, 355–363 (2013).
Roager, H. M. et al. Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut. Nat Microbiol 1, 16093 (2016).
Vandeputte, D. et al. Stool consistency is strongly associated with gut microbiota richness and composition, enterotypes and bacterial growth rates. Gut 65, 57–62 (2016).
Alles, M. S. et al. Bacterial fermentation of fructooligosaccharides and resistant starch in patients with an ileal pouch-anal anastomosis. Am. J. Clin. Nutr. 66, 1286–1292 (1997).
De Preter, V. et al. Effects of Lactobacillus casei Shirota, Bifidobacterium breve, and oligofructose-enriched inulin on colonic nitrogen-protein metabolism in healthy humans. Am. J. Physiol. Gastrointest. Liver Physiol. 292, G358–368 (2007).
Swanson, K. S. et al. Fructooligosaccharides and Lactobacillus acidophilus modify bowel function and protein catabolites excreted by healthy humans. J. Nutr. 132, 3042–3050 (2002).
Costalos, C., Kapiki, A., Apostolou, M. & Papathoma, E. The effect of a prebiotic supplemented formula on growth and stool microbiology of term infants. Early Hum. Dev. 84, 45–49 (2008).
Vivatvakin, B., Mahayosnond, A., Theamboonlers, A., Steenhout, P. G. & Conus, N. J. Effect of a whey-predominant starter formula containing LCPUFAs and oligosaccharides (FOS/GOS) on gastrointestinal comfort in infants. Asia Pac. J. Clin. Nutr. 19, 473–480 (2010).
Ziegler, E. et al. Term infants fed formula supplemented with selected blends of prebiotics grow normally and have soft stools similar to those reported for breast-fed infants. J. Pediatr. Gastroenterol. Nutr. 44, 359–364 (2007).
Kunz, C. et al. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 64, 789–798 (2017).
Smilowitz, J. T. et al. The human milk metabolome reveals diverse oligosaccharide profiles. J. Nutr. 143, 1709–1718 (2013).
Martinic, A. et al. Supplementation of Lactobacillus plantarum Improves Markers of Metabolic Dysfunction Induced by a High Fat Diet. J. Proteome Res. 17, 2790–2802 (2018).
Grip, T. et al. Serum, plasma and erythrocyte membrane lipidomes in infants fed formula supplemented with bovine milk fat globule membranes. Pediatr. Res 84, 726–732 (2018).
Lewis, S. J. & Heaton, K. W. Stool form scale as a useful guide to intestinal transit time. Scand. J. Gastroenterol. 32, 920–924 (1997).
Wesolowska-Andersen, A. et al. Choice of bacterial DNA extraction method from fecal material influences community structure as evaluated by metagenomic analysis. Microbiome 2, 19 (2014).
Caporaso, J. G. et al. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 108(Suppl 1), 4516–4522 (2011).
Bokulich, N. A., Bamforth, C. W. & Mills, D. A. Brewhouse-resident microbiota are responsible for multi-stage fermentation of American coolship ale. PLoS One 7, e35507 (2012).
Gohl, D. M. et al. Systematic improvement of amplicon marker gene methods for increased accuracy in microbiome studies. Nat. Biotechnol. 34, 942–949 (2016).
Salter, S. J. et al. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 12, 87 (2014).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336 (2010).
Aronesty, E. ea-utils: Command-line tools for processing biological sequencing data. Durham, NC: Expression Analysis (2011).
Kopylova, E. et al. Open-source sequence clustering methods improve the state of the art. mSystems 1, e00003-15 (2016).
DeSantis, T. Z. et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72, 5069–5072 (2006).
Langille, M. G. I. et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 31, 814–821 (2013).
Mandal, S. et al. Analysis of composition of microbiomes: a novel method for studying microbial composition. Microb. Ecol. Health Dis. 26, 27663 (2015).
Romano, J., Kromrey, J. D., Coraggio, J. & Skowronek, J. Appropriate statistics for ordinal level data: Should we really be using t-test and Cohen’sd for evaluating group differences on the NSSE and other surveys. In 1–33 (2006).
Gonzalez, A. et al. Qiita: rapid, web-enabled microbiome meta-analysis. Nat. Methods 15, 796–798 (2018).
Acknowledgements
This work was funded by Hero. The funders had no influence on the conduct of the study, collection, management, analysis and interpretation of the data or writing of this manuscript. The authors acknowledge support for CS from the United States Department of Agriculture National Institute of Food and Agriculture Hatch Project [1005945], and the Kinsella Endowed Chair in Food, Nutrition, and Health. The 600 MHz NMR is supported through the National Institutes of Health [1S10RR011973-01].
Author information
Authors and Affiliations
Contributions
Clinical study design: Olle Hernell, Bo Lönnerdal, Magnus Domellöf. Recruitment of participants and metadata collection: Niklas Timby & Tove Grip. Experimental design and supervision: Carolyn M. Slupsky. NMR Sample preparation: Mariana Parenti & Xuan He. Fecal DNA extraction and library preparation: Mariana Parenti & Xuan He. NMR and fecal microbiome data generation and analysis: Xuan He. Interpretation: Xuan He & Carolyn M. Slupsky. Writing of the manuscript: Xuan He. Primary responsibility for final content of the manuscript: Xuan He & Carolyn M. Slupsky. Reading, editing and approval of the final manuscript: all authors.
Corresponding author
Ethics declarations
Competing Interests
He and Parenti declare no personal or financial conflict of interest. Grip, Hernell, Lönnerdal, Slupsky, Timby received travel grants from Hero. Hernell and Lönnerdal are members of Semper and Hero Scientific Advisory Boards.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
He, X., Parenti, M., Grip, T. et al. Fecal microbiome and metabolome of infants fed bovine MFGM supplemented formula or standard formula with breast-fed infants as reference: a randomized controlled trial. Sci Rep 9, 11589 (2019). https://doi.org/10.1038/s41598-019-47953-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-47953-4
This article is cited by
-
Milk oligosaccharide-driven persistence of Bifidobacterium pseudocatenulatum modulates local and systemic microbial metabolites upon synbiotic treatment in conventionally colonized mice
Microbiome (2023)
-
Targeting the gut-lung axis by synbiotic feeding to infants in a randomized controlled trial
BMC Biology (2023)
-
Longitudinal profiles of the fecal metabolome during the first 2 years of life
Scientific Reports (2023)
-
Effect of beta-glucan supplementation on cystic fibrosis colonic microbiota: an in vitro study
Pediatric Research (2023)
-
The gut microbiome-metabolome dataset collection: a curated resource for integrative meta-analysis
npj Biofilms and Microbiomes (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
