
Abstract
The large conductance, calcium- and voltage-activated potassium channel, known as the BK channel, is one of the central proteins that mediate alcohol intoxication and tolerance across species. Although ethanol targets BK channels through direct interaction, how ethanol-mediated BK channel activation causes behavioral intoxication is poorly understood. In. C. elegans, loss of function in SLO-1, the BK channel ortholog, confers profound ethanol resistance in movement and egg-laying behaviors. Here, we show that depletion of SLO-1 channels clustered at the active zones with no change in the overall channel expression level results in locomotory resistance to the intoxicating effect of ethanol, equivalent to that of slo-1 loss-of-function mutants. Likewise, depletion of clustered SLO-1 channels in the sarcolemma and neurons leads to ethanol-resistant egg-laying behavior. By contrast, reduction in the overall SLO-1 channel level by over 70% causes only moderate ethanol resistance in movement, and minimal, if any, resistance in egg laying. Our findings strongly suggest that behavioral ethanol sensitivity is conferred by local, but not global, depression of excitability via clustered BK channels. Given that clustered BK channels are functionally coupled to, and localize near, calcium channels, ethanol may mediate its behavioral effects by targeting BK channels and their coupled calcium channels.
Similar content being viewed by others

Introduction
BK channels have emerged as a major mediator of alcohol response across species from C. elegans to humans1,2,3. In C. elegans genetic screens designed to identify genes that mediate the intoxicating effects of ethanol, slo-1 mutants were most frequently identified and showed profound resistance4. Moreover, ethanol-mediated BK channel activation is commonly observed in mice, rats, and humans, indicating that the effect of ethanol on BK channels is an evolutionarily conserved phenomenon5. Recent studies of C. elegans, mouse, and human BK channels have demonstrated that single amino acid mutations in the cytoplasmic C-terminal domain abolish ethanol-mediated BK channel activation without altering overall channel properties6,7, suggesting that ethanol activates BK channels through direct interaction with these amino acids. Importantly, the fact that these potential ethanol-binding sites are positioned away from known calcium binding sites suggests that ethanol may act on BK channels independently of calcium binding, although it is difficult to completely exclude the possibility that they may affect calcium binding indirectly through an allosteric gating mechanism. However, it is worthwhile to note that the consequence of the interaction between BK channels and ethanol is not straightforward in mammals2,3. While ethanol activates BK channels at calcium ion concentration below 20 μM, ethanol inhibits BK channels as calcium ion concentrations further increase8. Furthermore, β subunits of mammalian BK channels influence the calcium sensitivity of the channels and thus shift the range of calcium concentrations at which ethanol results in potentiation9. Given that the range of intracellular calcium concentrations is highly variable among different subcellular compartments, it is an open question whether ethanol uniformly targets all of the BK channels present in the plasma membrane or prefers certain types of BK channels that mediate the behavioral effects of ethanol.
Studies on SLO-1 in C. elegans provided insights into its localization and function in muscles and neurons10,11. CTN-1, an α-catulin ortholog that shares homology with α-catenin and vinculin12, clusters SLO-1 channels at either muscle excitation sites or presynaptic terminals by direct physical interaction13,14,15,16, and ERG-28 (a homolog of ergosterol biosynthetic protein 28), an endoplasmic reticulum (ER) membrane protein, promotes the trafficking of SLO-1 channels from the ER17. Consistent with these cell biological data, ctn-1 and erg-28 mutants were initially isolated as suppressors of slo-1 (gf, gain-of-function) phenotypes and were found to have deficits in evoked synaptic responses in electrophysiological recordings at the neuromuscular junction13,15. Despite their common involvement in the SLO-1-mediated synaptic response, ERG-28, but not CTN-1, is required for the inhibitory function of SLO-1 channels in calcium signaling during the establishment of the left-right asymmetry of AWC (Amphid Wing C cell) olfactory neurons13,17,18. This raised the question of how these two different genes distinctively contribute to SLO-1 function.
In this study, we used C. elegans to investigate how alcohol intoxication at the behavioral level is mediated via SLO-1 channels. Using different mutants that affect either SLO-1 channel clustering or density, we discovered that clustered, but not diffuse, SLO-1 channels at presynaptic terminals and presumed muscle excitation sites mediate the intoxicating effect of ethanol.
Results
ctn-1 mutants exhibit more prominent ethanol-resistant locomotory behavior than erg-28 mutants
The ctn-1 and erg-28 genes were previously identified as regulators of SLO-1 in a genetic screen to identify suppressors of sluggish movement of the slo-1(ky399) gain-of-function mutant13,17. While mutations in either gene suppress the sluggish movement of the slo-1(ky399gf) mutant, only an erg-28, but not a ctn-1, mutation suppresses the defective asymmetric differentiation of the AWC neuron pair in a slo-1(ky399gf) mutant13,17,18. This prompted us to determine whether mutations in these two genes have differential effects on ethanol intoxication, another well-known slo-1 function. Although we have recently reported that mutations in erg-28 confer resistance to the intoxicating effect of ethanol17, the effect of ctn-1 mutation on ethanol-mediated behavior has not been investigated. As previously reported, at an exogenous concentration that causes complete intoxication of the wild-type animals, slo-1(lf, loss-of-function) mutants exhibited a strongly ethanol-resistant locomotory phenotype4, and erg-28(gk697770) mutants exhibited a moderately ethanol-resistant locomotory phenotype (Fig. 1, Supplementary Video 1). We found that ctn-1(eg1167) mutants showed as much resistance as slo-1(lf) mutants (Fig. 1, Supplementary Video 1). The average speeds of these tested animals in the absence of ethanol did not differ by genotype (wild-type: 191 ± 12.3, erg-28: 197 ± 11.8, ctn-1: 187 ± 8.8, slo-1: 197 ± 7.6 μm sec−1, mean ± sem). Thus, whereas erg-28 mutation, but not ctn-1 mutation, suppresses defective asymmetric differentiation of the AWC neuron pair in the slo-1(ky399gf) mutant17, ctn-1 mutation is more effective than erg-28 mutation in abolishing the function of SLO-1 in ethanol intoxication.

ctn-1 and erg-28 mutations confer differential resistance to the intoxicating effect of ethanol. (A) The locomotory speeds of wild-type, slo-1(eg142), erg-28(gk697770), or ctn-1(eg1167) mutant animals were measured in the presence of ethanol and their values were divided by the average speed of untreated animals. Error bars represent S.E.M. One-way ANOVA with Tukey’s post-hoc analysis (wild-type vs. erg-28: p = 0.0473, wild-type vs. ctn-1: p < 0.001, wild-type vs. slo-1: p < 0.001, slo-1 vs. ctn-1: p = 0.8136). Data points represent three independent trials of 10 animals. See Supplementary Video 1. (B) Snapshots of single animals in the presence of an intoxicating dose of ethanol show sinusoidal posture.
CTN-1 is essential for SLO-1 channel clustering, while ERG-28 is necessary to maintain the levels of SLO-1 channels
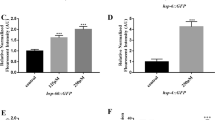
What could be the mechanism for these differential effects of erg-28 and ctn-1 mutation? ERG-28 is required for efficient trafficking of SLO-1 to the plasma membrane and erg-28 mutation leads to a drastic reduction in the number of SLO-1 channels, but SLO-1 puncta are still present at presynaptic terminals. CTN-1 is necessary for SLO-1 to cluster at presynaptic terminals. This led us to hypothesize that the function of SLO-1 in ethanol intoxication is critically dependent on its clustering at presynaptic terminals, while its function in AWC neuron differentiation is not. In a previous study on CTN-115, it was not conclusive whether ctn-1 mutation affect the SLO-1 protein level in addition to clustering because a transgenic line overexpressing SLO-1 in a select group of motor neurons was used. Hence, we used a genome-edited line expressing GFP-tagged SLO-1 to determine SLO-1 expression levels and localization in the ctn-1 mutant and compared them with the erg-28 mutant. This genome-edited line, cim105[slo-1::GFP], is indistinguishable from wild-type animals in locomotive speed, body posture and egg-laying behavior17. ctn-1 mutation disrupts SLO-1 clustering at presynaptic terminals and muscle excitation sites (Figs 2 and 3). In erg-28(gk697770) mutants, the number of clustered SLO-1 channels was reduced to approximately 25% of the quantity in wild type animals. In ctn-1(eg1167) mutants, the number of clustered SLO-1 channels in the dorsal nerve cord was reduced to a level that was not reliably quantifiable (Fig. 2). In body wall muscle cells, most SLO-1 channels aligned as distinct punctal structures in the sarcolemma. In erg-28(gk697770) mutants, the clusters of SLO-1 channels were reduced in number compared to those of wild-type animals. In ctn-1(eg1167) mutants, numerous low, diffused fluorescent signals were present but they did not form high-intensity clusters similar to those in wild-type animals (Figs 2 and 3). These low-intensity fluorescent signals are not readily detectable with a line-scanning quantification method. To further understand the defect of SLO-1 cluster formation in ctn-1 mutants, we tracked low-level fluorescent signals using time-lapse imaging. Unlike those of wild-type animals, the low-level fluorescent signals of mutants were highly mobile and occasionally formed slightly intense puncta, which quickly dissipated (Supplementary Video 2, Supplementary Fig. 2). Likewise, highly mobile low-level fluorescent signals were observed in the dorsal cord of ctn-1 mutant animals (Supplementary Video 2). Weak, mobile SLO-1 signals in ctn-1 mutants could be the result of either endosome-mediated SLO-1 degradation or diffusion of SLO-1 channels in the plasma membrane due to the lack of anchoring. To distinguish these two possibilities, we measured the amount of SLO-1::GFP using Western blot analysis. The total amount of SLO-1::GFP was reduced in erg-28(gk697770) mutants by over 70% but was not significantly different in ctn-1(eg1167) mutants when compared to wild-type animals (Fig. 3). This indicates that CTN-1 is absolutely required for SLO-1 clustering and that its absence causes SLO-1 channels to adopt diffuse, mobile forms without promoting degradation.

Mutation of the ctn-1 gene depletes clustered SLO-1 channels in neurons, whereas mutation of the erg-28 gene reduces the number of clustered SLO-1 channels. (A) SLO-1 localization pattern in the dorsal cord of wild-type, erg-28(gk697770), and ctn-1(eg1167) mutant animals. The slo-1(cim105[slo-1::GFP]) strain, which was generated using genome-editing17, was used in the analysis. Scale bar, 5 μm. (B) The number of SLO-1 puncta representing clustered SLO-1 channels in the dorsal cord. n = 10, Sample numbers represent independent biological replicates.; F(9, 9) = 4.173, p = 0.0447, ANOVA, t(10.23) = 18, p < 0.0001, unpaired two-tailed t-test. ctn-1 mutants were excluded from the test, since we could not detect stable puncta.

Mutation of the ctn-1 gene depletes clustered SLO-1 channels in muscle without affecting the overall SLO-1 levels. (A,B) Representative SLO-1 localization patterns in the sarcolemma of wild-type, erg-28(gk697770), and ctn-1(eg1167) mutant animals. The right panels are zoomed-in images of the boxed areas of the left panel images. Note that ctn-1 mutants exhibit low-level fluorescent SLO-1 signals that fail to form large high-intensity clusters. Scale bar: 5 μm. Comparison of head muscle SLO-1 channel clusters between wild-type and erg-28(gk697770) animals: F(17, 23) = 1.683, p = 0.2429, ANOVA, t(9.140) = 40, p < 0.0001, unpaired two-tailed t-test. (C) Western blot analysis shows that the total SLO-1::GFP ratio is not reduced in ctn-1 mutants. SLO-1::GFP intensities in wild-type, erg-28(gk697770), and ctn-1(eg1167) were normalized to α-tubulin, and their intensities relative to wild-type are indicated. N2 represents a wild type animal that does not express GFP and serves as a negative control for Western blot with an anti-GFP antibody. The blot was probed first with anti-GFP antibody (anti-rabbit monoclonal) and then with anti-α-tubulin antibody (anti-mouse monoclonal) (Supplementary Fig. 3).
The inability of ctn-1 mutation to suppress the defective AWC neuron differentiation of slo-1(gf) mutants indicates that SLO-1 function is normal in AWC neurons of ctn-1 mutant animals. Thus, we examined SLO-1 localization in AWC neurons using transgenic animals expressing SLO-1::GFP under the odr-1 promoter, which is active in AWC and AWB (amphid wing B cell) neurons. Consistent with the results from the nerve cords, SLO-1::GFP is not clustered but diffused in the axons of the ctn-1 mutant (Supplementary Fig. 1). This suggests that the overall level of SLO-1, rather than clustering, is critical for its function in controlling asymmetric differentiation of AWC neurons. These results further support the idea that SLO-1 channels in ctn-1 mutant animals remain in the plasma membrane.
SLO-1 channels clustered at the presynaptic terminals confer ethanol sensitivity in locomotion
A previous study showed that neuronal SLO-1 function is critical for the intoxicating effect of ethanol on locomotory behavior4. Expression of the slo-1 gene in neurons converts the ethanol-resistant locomotory phenotype of slo-1(lf) mutants to a wild-type-like sensitive phenotype. Therefore, we asked whether the clustering of SLO-1 channels at presynaptic terminals of neurons is essential for mediating alcohol-induced sedation. To answer this question, we first expressed a wild-type copy of ctn-1 in either muscles or neurons of ctn-1(eg1167) mutant animals and determined whether SLO-1 channel clusters were restored. Muscular and neuronal expression restored SLO-1 channel clusters in each transgenic line (Fig. 4A–D). Next, we determined whether we could reverse ethanol-resistant locomotory behavior of ctn-1 mutants in neuronal and muscular transgenic lines (Fig. 4E). We found that neuronal expression, but not muscular expression, of the ctn-1 gene reversed the ethanol-resistant locomotory behavior of the ctn-1 mutants (Fig. 4E). Taken together, these results indicate that clustered SLO-1 channels in neurons are responsible for mediating the intoxicating effects of ethanol.

CTN-1-mediated SLO-1 clustering in neurons is critical for alcohol intoxication. (A,B) Muscle expression of a wild-type ctn-1 in ctn-1(eg1167) mutant animals restores SLO-1 clustering in the sarcolemma, but not the neurons in the dorsal cord. The white arrows indicate the dorsal cord. +muscle: myo-3 promoter-driven expression of ctn-1. Scale bar, 10 μm. Comparison of muscle SLO-1 channel clusters between wild-type and muscle-rescued ctn-1 mutant animals: F(29, 29) = 1.393, p = 0.3770, ANOVA, t(1.296) = 58, p = 0.2002, unpaired two-tailed t-test. (C,D) Neuronal expression of a wild-type ctn-1 in ctn-1(eg1167) mutant animals restores SLO-1 clustering at presynaptic terminals, but not sarcolemma. +neuron: rgef-1 promoter-driven expression of ctn-1. Scale bar,10 μm. Comparison of neuronal SLO-1 channel clusters between wild-type and neuron-rescued ctn-1 mutant animals: F(9, 9) = 1.406, p = 0.6197, ANOVA, t(5.405) = 18, p < 0.0001, unpaired two-tailed t-test. (E) Neuronal, but not muscular, ctn-1 expression restores normal ethanol-sensitive locomotory behavior. Locomotory speed was measured as in Fig. 1. +muscle: myo-3 promoter-driven expression of ctn-1. +neuron: H20 pan-neuronal promoter-driven expression of ctn-1. Error bars represent S.E.M. One-way ANOVA with Tukey’s post-hoc analysis (ctn-1 ethanol treated vs. ctn-1(+neuron) ethanol treated: p = 0.0145, ctn-1 ethanol treated vs. ctn-1(+muscle) ethanol treated: p = 0.8477). Data points represent three independent trials of 10 animals.
Ethanol-mediated egg-laying suppression is primarily mediated by clustered SLO-1 channels
In addition to locomotory behavior, slo-1 mutants exhibit robust ethanol-resistant egg-laying behavior. The neural circuits for egg-laying behavior is different from the circuits for locomotory behavior. While locomotory behavior is mediated by striated body wall muscle that receives cholinergic and GABAergic inputs from motor neurons, egg laying is mediated by smooth muscle cells that receive serotonergic and cholinergic inputs from the HSN (Hermaphrodite Specific Neuron) and VC (Ventral Cord) neurons19. We examined whether erg-28(gk697770) and ctn-1(eg1167) mutants also exhibit ethanol-resistant egg-laying behavior (Fig. 5). In the presence of an ethanol dose that suppresses the egg laying of wild-type animals almost completely, slo-1 and ctn-1 mutants laid significantly more eggs over 60 min than wild-type animals (Fig. 5). However, erg-28 mutants show minimal, if any, egg laying under the same conditions. These results indicate that clustered SLO-1 channels are responsible for ethanol-mediated suppression of egg laying. Next, we asked in which tissue the clustering of SLO-1 channels is responsible for ethanol-mediated suppression of egg laying. Specifically, we investigated whether the ethanol-mediated change in egg laying behavior is mediated through egg-laying muscles or upstream neurons (Fig. 6). Muscle-specific expression of the ctn-1 gene in ctn-1(eg1167) mutants restored the SLO-1 clusters in egg laying muscles (Fig. 6A). This restored SLO-1 channel clustering reduced egg laying in the presence of ethanol within a 60-min time period. These results indicate that SLO-1 channel clusters in egg-laying muscles significantly contribute to ethanol sensitivity (Fig. 6B). Likewise, animals expressing a wild-type copy of the ctn-1 gene under the control of a pan-neuronal promoter in ctn-1 mutants, which restores SLO-1 clustering in the neuron (Fig. 2), laid moderately fewer eggs than ctn-1(eg1167) mutants within a 60-min time period in response to the same ethanol dose. Taken together, these results indicate that, unlike locomotory behavior, ethanol-mediated suppression of egg laying is mediated by clustered SLO-1 channels present in both egg-laying muscles and neurons.

Mutation of the ctn-1 gene, but not the erg-28 gene, causes strong ethanol-resistant egg-laying behavior. Egg-laying behavior of wild-type, slo-1(eg142), erg-28(gk697770), or ctn-1(eg1167) mutant animals was quantified by measuring the number of eggs laid over a 60 min period in the presence or absence of ethanol. Egg laying was measured in 10 age-matched animals (30 h post L4) and repeated three times (biological replicates). The data (mean + SEM) are presented as the number of eggs laid by individual animals. One-way ANOVA with Tukey’s post-hoc analysis (wild-type N2 vs. wild-type N2 ethanol treated: ****p < 0.0001, slo-1 vs. slo-1 ethanol treated: p = 0.1174, erg-28 vs. erg-28 ethanol treated: ****p < 0.0001, ctn-1 vs. ctn-1 ethanol treated: p < 0.1517).

CTN-1-mediated SLO-1 clustering in neurons and egg-laying muscles contributes to ethanol-mediated suppression of egg laying behavior. (A) Muscle expression of a wild-type copy of ctn-1 in ctn-1(eg1167) mutant animals restores SLO-1 clustering at egg-laying muscles. The images represent a maximum projection of Z-stack image sections. The asterisks, arrows, and arrowheads denote the ventral nerve cord, the vulval slit, and SLO-1 clusters present in egg-laying muscle cells, respectively. The number of SLO-1 channel clusters were quantified from a region of a single muscle cell, which was not overlapped with body wall muscle or the ventral cord. Comparison of vulval muscle SLO-1 channel clusters between wild-type and muscle-rescued ctn-1 mutant animals: F(6, 6) = 1.039, p = 0.96, ANOVA, t(1.482) = 12, p < 0.1642, unpaired two-tailed t-test. WT: wild-type, +muscle: myo-3 promoter-driven expression of ctn-1. (B) Neuronal and muscular SLO-1 channel clustering contributes to ethanol-mediated suppression of egg laying. Egg laying was measured as in Fig. 5. Egg laying was measured in 10 age-matched animals (30 h post L4) and repeated three times (biological replicates). +muscle: myo-3 promoter-driven expression of ctn-1. +neuron: H20 pan-neuronal promoter-driven expression of ctn-1. Error bars represent S.E.M. One-way ANOVA with Tukey’s post-hoc analysis (ctn-1 ethanol treated vs. ctn-1(+neuron) ethanol treated: p = 0.0086, ctn-1 ethanol treated vs. ctn-1(+muscle) ethanol treated: p < 0.0001).
Discussions
BK channels are known to mediate ethanol sensitivity across a wide variety of species, including C. elegans. Here, we demonstrate in a C. elegans genetic study that the behavioral effects of ethanol are mediated by clustered, but not dispersed, SLO-1 channels. We show that ctn-1 mutation causes a drastic elimination of clustered SLO-1 channels and, as a consequence, an increase in diffuse SLO-1 channels without affecting their overall level. We found that this ctn-1 mutation confers ethanol-resistant locomotory and egg-laying behaviors equivalent to slo-1 null mutations, indicating that the behavioral effect of ethanol is mediated by clustered SLO-1 channels.
Our study provides evidence that some of the functions of SLO-1, such as alcohol intoxication, are mediated uniquely by clustered SLO-1 channels, while other functions, such as asymmetry of olfactory neurons, are mediated by diffuse, scattered SLO-1 channels. Our previous study showed that ctn-1 mutation abolishes SLO-1 puncta in neurons and muscles, although we were not able to definitively conclude whether ctn-1 mutation causes any change in diffuse, scattered SLO-1 channels13,15. Using a SLO-1::GFP genome-edited strain we now show that ctn-1 mutation disperses SLO-1 channels without affecting their overall abundance. This steady level of SLO-1 channels in the ctn-1 mutant is in contrast with the dramatically reduced SLO-1 channel level observed in the erg-28 mutant. Mutation in the erg-28 gene causes ineffective exit of SLO-1 channels from the ER, thus resulting in the degradation of a majority of SLO-1 channels and a consequent reduction in SLO-1 channel density17. Mutation in the ctn-1 gene does not suppress the defect in the left-right asymmetry of AWC olfactory neurons observed in slo-1 gain-of-function mutants, while erg-28 mutation does17. These results indicate that the left-right asymmetry of AWC olfactory neurons is associated with the total number of SLO-1 channels, but not the clustering of SLO-1 channels. On the other hand, ctn-1 mutants exhibit prominent ethanol-resistant locomotory and egg-laying behaviors, whereas erg-28 mutants show moderate ethanol-resistant locomotory and weak, if any, egg-laying behaviors.
Another interesting result obtained in our study is how the density of BK channels is regulated. The density of ion channels is often regulated by the retrieval from the plasma membrane and subsequent degradation by proteasomes or endocytosis/lysosomes20,21,22. It is conceivable that SLO-1 channels similarly undergo degradation via proteasome- or endocytosis/lysosome-mediated degradation when they cannot form clusters at presynaptic terminals and presumed muscle excitation sites. Contrary to this belief, our data indicate that the density of SLO-1 channels is not regulated by the retrieval from the plasma membrane. ctn-1 mutation abolishes SLO-1 channel clustering and causes SLO-1 channels to disperse through neurons and muscles but does not influence the overall levels of SLO-1 channels. Furthermore, because ctn-1 mutation does not suppress the defect in the left-right asymmetry of slo-1(gf) AWC neurons, disperse SLO-1 channels are functional and are likely present at the plasma membrane. By contrast, erg-28 mutation, which disrupts normal SLO-1 channel trafficking in the ER, reduces the abundance of SLO-1 channels by over 70%. Therefore, the major regulatory point of the overall SLO-1 channel level is channel trafficking in the ER.
Why does SLO-1 channel-mediated alcohol intoxication require clustering at presynaptic terminals and the sarcolemma? One possibility is that ethanol can activate both clustered and diffuse SLO-1 channels but only clustered SLO-1 channels are relevant to ethanol sensitivity due to their coupling with calcium channels that mediate behavioral sensitivity to ethanol. It has been reported that BK channels are functionally coupled with a wide variety of calcium channels23,24,25,26,27, which localize to distinct subcellular compartments, including pre- and post-synaptic sites, the ER, and muscle excitation sites28,29. Although clustered and diffuse SLO-1 channels both increase potassium efflux in response to ethanol, only clustered SLO-1 channels lead to an inhibition of behaviorally-relevant calcium channels, thereby causing sedation at the behavioral level. Alternatively, it is possible that, due to their unique locations, SLO-1 channels localized in the clusters bind to ethanol more readily than diffuse channels do. These clustered SLO-1 channels are found at the major entry sites for calcium ions, such as presynaptic terminals and muscle excitation sites, and thus could be exposed to a specific level of calcium ions that makes channel conformation most favorable for ethanol binding. BK channels have indeed been described to adopt multiple complex conformations depending on calcium and voltage8,30,31,32.
Our study demonstrates that the clustering of SLO-1 at presynaptic terminals or presumed muscle excitation sites plays a critical role in mediating ethanol sensitivity. It is generally thought that ethanol-mediated SLO-1 activation reduces overall neural excitability by hyperpolarizing the entire neuron. However, our study indicates that the depressive behavioral effects of ethanol on both locomotory and egg-laying behavior are mediated by clustered SLO-1 channels, suggesting that ethanol inhibits specific molecular steps through a local hyperpolarization mediated by SLO-1 channels. Given that BK channels are functionally coupled with calcium channels in muscles and neurons, these molecular steps likely involve the functions of calcium channels. While our recent study showed that neuron-specific P/Q-type voltage-gated UNC-2 calcium channels, which localize to presynaptic terminals33, indirectly contribute to SLO-1 localization in select cholinergic neurons15, whether alcohol-mediated SLO-1 activation suppresses UNC-2 function is not clear. Furthermore, it is not clear which calcium channel is the target of alcohol-mediated SLO-1 activation in egg-laying muscles. In the future, it will be necessary to identify calcium channels that couple with clustered SLO-1 channels and mediate the behavioral effects of ethanol.
Drug targeting of BK channels has been suggested as a tool to control alcohol dependence or alcohol withdrawal34,35,36,37. Given that both diffuse and clustered BK channels serve overlapping and distinct functions in C. elegans and mammals38,39, targeting clustered BK channels may offer a better therapeutic strategy than indiscriminate drug targeting of BK channels. Blocking BK channel clustering will be an effective way of de-coupling BK channels from calcium channels that mediate behavioral alcohol response while sparing the functions dependent on diffuse, disperse BK channels.
Methods
Worm strains and maintenance
All C. elegans strains were cultured at 20 °C on NGM (nematode growth medium) plates seeded with E. coli OP50. Table 1 lists all strains along with their genotypes and origins. Transgenic strains were constructed using standard DNA microinjection methods with either the coelomocyte unc-122p::GFP(or mCherry) or AIY (Amphid Interneuron Y) neuron ttx-3p::RFP co-injection marker40.
Ethanol-resistant locomotory behavior assay
All animals used for locomotory assays were day 1 adults (20–24 hours post L4). The overall method for the assay has been previously described4,41. The NGM agar plates used for all of the experiments were dried without lids for 25 min in a 60 °C incubator. This drying process was necessary to prevent the animals from exhibiting a swimming-like movement on wet plates, and this process usually reduces the agar to 85–90% of its original volume. After the plates were cooled down to room temperature, a desired amount of ethanol (400 mM), which gives internal ethanol concentration of approximately 50 mM42, was applied to the NGM plates. The plates were immediately sealed with parafilm and incubated for an additional 2 hours to equilibrate with the ethanol. Immediately before the assays, three flame-heated copper rings were embedded in the agar surface of the NGM plates. These copper rings allow for the simultaneous comparison of three different genotypes in the same assay plates. We included wild-type and slo-1(lf) mutant (or ctn-1 mutant) animals in the ethanol assays as positive and negative controls. Ten animals with a given genotype were incubated for 10 min in unseeded plates to remove attached bacteria and then transferred inside a copper ring. After 10 min of exposure to 0 mM and 400 mM ethanol, video frames from three different genotypes were simultaneously acquired from a dissecting microscope fitted with GO-3 camera (Qimaging) with 500 ms intervals for 2 min. For each genotype, the experiments on ethanol plates were repeated three times. We calculated the average speed of the tested animals using the Tracking Objects option in Image-Pro Plus 7.0 (Media Cybernetics, RRID:SCR_016879). The software algorithm automatically generates new tracks when the paths of animals overlap due to collision.
Ethanol-resistant egg-laying behavior assay
All animals used for the egg-laying assays were day 1 adults (30 hours post L4). Unlike the locomotory assays, the egg-laying assay requires bacterial food4,7. NGM agar plates were dried without lids for 2 hours in a 37 °C incubator and then seeded in a triradiate shape with E. coli OP50. The next day, three small circular agar plugs (1 cm diameter) were punched out from the outside of the bacterial lawn and ethanol (500 mM) was then added to the holes. The agar plugs were placed back into the holes after the ethanol was halfway absorbed into the agar plate. During this process, we ensured that the ethanol did not come into direct contact with the bacterial food. After being sealed with parafilm, the plates were equilibrated at room temperature for approximately 2 hours. As in the locomotory assay, three copper rings were embedded on the agar surface, but the rings were placed where the bacterial food was spread. These copper rings allow the simultaneous comparison of three different genotypes in the same assay plates. Ten animals were directly transferred to each copper ring, and the number of eggs laid was counted at 60 min. The assay included wild-type and slo-1(lf) mutant animals in the ethanol assays as negative and positive controls, respectively. The experiments were repeated three times for each genotype.
Microscopy and time-lapse imaging
Images were acquired using a 63x/1.4 numerical aperture on a Zeiss microscope (Axio-Observer Z1) equipped with Spectra X light engine (Lumencor). Images were captured with an 82% quantum efficiency Zyla 4.2 PLUS sCMOS camera (Andor) using MetaMorph 7.8 (RRID:SCR_002368) at 200 ms intervals with 350 ms exposure time. To quantify the intensity of SLO-1 channel clusters, a 150-pixel line was drawn on the linearly aligned fluorescence clusters in head and body-wall muscles, and its pixel intensity was quantified using a line-scanning method in MetaMorph software package (RRID:SCR_002368). To accommodate the variability within each muscle, data obtained from three neighboring parallel lines were averaged for a single data point. For the quantification in egg-laying muscle, the number of fluorescence clusters was counted from an area of a single vulval muscle cell, which was not overlapped with body wall muscle cells or the ventral nerve cord.
Western blotting and quantification
Mixed-stage worms were lysed and solubilized in SDS lysis buffer (2% SDS, 100 mM NaCl, 10% glycerol, and 50 mM Tris-HCl, pH 6.8) with sonication. Worm extracts were separated by SDS-PAGE and transferred onto PVDF membranes. The separated proteins were probed with anti-GFP (Cell Signaling Technology, #2956, RRID: AB_1196615) or anti-tubulin (Developmental Studies Hybridoma Bank, AA4.3, RRID:AB_579793) antibodies. The pixel intensities of GFP and tubulin bands were quantified using Adobe Photoshop CC 2015.
Experimental design and statistical analysis
The speed data (Figs 1A and 4E) and egg-laying data (Figs 5 and 6B) were analyzed using one-way ANOVA with Tukey’s post hoc analysis, and the number of fluorescence clusters was analyzed using a two-tailed t-test (GraphPad Prism 7, RRID:SCR_002798). Statistical analyses for each experiment are detailed in the figure legends.
References
Bettinger, J. C. & Davies, A. G. The role of the BK channel in ethanol response behaviors: evidence from model organism and human studies. Frontiers in physiology 5, 346, https://doi.org/10.3389/fphys.2014.00346 (2014).
Dopico, A. M., Bukiya, A. N. & Martin, G. E. Ethanol modulation of mammalian BK channels in excitable tissues: molecular targets and their possible contribution to alcohol-induced altered behavior. Frontiers in physiology 5, 466, https://doi.org/10.3389/fphys.2014.00466 (2014).
Dopico, A. M., Bukiya, A. N., Kuntamallappanavar, G. & Liu, J. Modulation of BK Channels by Ethanol. International review of neurobiology 128, 239–279, https://doi.org/10.1016/bs.irn.2016.03.019 (2016).
Davies, A. G. et al. A central role of the BK potassium channel in behavioral responses to ethanol in C. elegans. Cell 115, 655–666 (2003).
Mulholland, P. J. et al. Sizing up ethanol-induced plasticity: the role of small and large conductance calcium-activated potassium channels. Alcoholism, clinical and experimental research 33, 1125–1135, https://doi.org/10.1111/j.1530-0277.2009.00936.x (2009).
Bukiya, A. N. et al. An alcohol-sensing site in the calcium- and voltage-gated, large conductance potassium (BK) channel. Proceedings of National Academy of Science USA 111, 9313–9318, https://doi.org/10.1073/pnas.1317363111 (2014).
Davis, S. J., Scott, L. L., Hu, K. & Pierce-Shimomura, J. T. Conserved single residue in the BK potassium channel required for activation by alcohol and intoxication in C. elegans. J Neurosci 34, 9562–9573, https://doi.org/10.1523/JNEUROSCI.0838-14.2014 (2014).
Liu, J., Vaithianathan, T., Manivannan, K., Parrill, A. & Dopico, A. M. Ethanol modulates BKCa channels by acting as an adjuvant of calcium. Molecular Pharmacology 74, 628–640, https://doi.org/10.1124/mol.108.048694 (2008).
Bukiya, A. N., Liu, J. & Dopico, A. M. The BK channel accessory beta1 subunit determines alcohol-induced cerebrovascular constriction. FEBS Lett 583, 2779–2784, https://doi.org/10.1016/j.febslet.2009.07.019 (2009).
Kim, H. et al. The dystrophin complex controls bk channel localization and muscle activity in Caenorhabditis elegans. PLoS Genetics 5, e1000780, https://doi.org/10.1371/journal.pgen.1000780 (2009).
Kim, H. & Oh, K. H. Protein Network Interacting with BK Channels. International review of neurobiology 128, 127–161, https://doi.org/10.1016/bs.irn.2016.03.003 (2016).
Janssens, B., Staes, K. & van Roy, F. Human alpha-catulin, a novel alpha-catenin-like molecule with conserved genomic structure, but deviating alternative splicing. Biochim Biophys Acta 1447, 341–347 (1999).
Abraham, L. S., Oh, H. J., Sancar, F., Richmond, J. E. & Kim, H. An alpha-catulin homologue controls neuromuscular function through localization of the dystrophin complex and BK channels in Caenorhabditis elegans. PLoS Genetics 6, e1001077, https://doi.org/10.1371/journal.pgen.1001077 (2010).
Oh, H. J. et al. Interaction of alpha-catulin with dystrobrevin contributes to integrity of dystrophin complex in muscle. Journal of Biological Chemistry 287, 21717–21728, https://doi.org/10.1074/jbc.M112.369496 (2012).
Oh, K. H. et al. Presynaptic BK channel localization is dependent on the hierarchical organization of alpha-catulin and dystrobrevin and fine-tuned by CaV2 calcium channels. BMC neuroscience 16, 26, https://doi.org/10.1186/s12868-015-0166-2 (2015).
Chen, B. et al. alpha-Catulin CTN-1 is required for BK channel subcellular localization in C. elegans body-wall muscle cells. EMBO J 29, 3184–3195 (2010).
Oh, K. H. et al. ERG-28 controls BK channel trafficking in the ER to regulate synaptic function and alcohol response in C. elegans. eLife 6, e24733 (2017).
Alqadah, A. et al. SLO BK Potassium Channels Couple Gap Junctions to Inhibition of Calcium Signaling in Olfactory Neuron Diversification. PLoS Genetics 12, e1005654, https://doi.org/10.1371/journal.pgen.1005654 (2016).
White, J. G., Southgate, E., Thomson, J. N. & Brenner, S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos Trans R Soc Lond B Biol Sci 314, 1–340 (1986).
Manna, P. T. et al. Constitutive endocytic recycling and protein kinase C-mediated lysosomal degradation control K(ATP) channel surface density. Journal of Biological Chemistry 285, 5963–5973, https://doi.org/10.1074/jbc.M109.066902 (2010).
Felix, R. & Weiss, N. Ubiquitination and proteasome-mediated degradation of voltage-gated Ca2+ channels and potential pathophysiological implications. Gen Physiol Biophys 36, 1–5, https://doi.org/10.4149/gpb_2016037 (2017).
Conrad, R. et al. Rapid Turnover of the Cardiac L-Type CaV1.2 Channel by Endocytic Recycling Regulates Its Cell Surface Availability. iScience 7, 1–15, https://doi.org/10.1016/j.isci.2018.08.012 (2018).
Berkefeld, H. et al. BKCa-Cav channel complexes mediate rapid and localized Ca2+- activated K+ signaling. Science 314, 615–620 (2006).
Isaacson, J. S. & Murphy, G. J. Glutamate-mediated extrasynaptic inhibition: direct coupling of NMDA receptors to Ca(2+)-activated K+ channels. Neuron 31, 1027–1034 (2001).
Wellman, G. C. & Nelson, M. T. Signaling between SR and plasmalemma in smooth muscle: sparks and the activation of Ca2+ -sensitive ion channels. Cell Calcium 34, 211–229 (2003).
Weaver, A. K., Olsen, M. L., McFerrin, M. B. & Sontheimer, H. BK channels are linked to inositol 1,4,5-triphosphate receptors via lipid rafts: a novel mechanism for coupling [Ca(2+)](i) to ion channel activation. Journal of Biological Chemistry 282, 31558–31568, https://doi.org/10.1074/jbc.M702866200 (2007).
Zhang, J. et al. Glutamate-activated BK channel complexes formed with NMDA receptors. Proceedings of National Academy of Science USA 115, E9006–E9014, https://doi.org/10.1073/pnas.1802567115 (2018).
Wang, X., Wang, G., Lemos, J. R. & Treistman, S. N. Ethanol directly modulates gating of a dihydropyridine-sensitive Ca2+ channel in neurohypophysial terminals. J Neurosci 14, 5453–5460 (1994).
Balino, P., Ledesma, J. C. & Aragon, C. M. In vivo study of ethanol-activated brain protein kinase A: manipulations of Ca2+ distribution and flux. Alcoholism, clinical and experimental research 38, 629–640, https://doi.org/10.1111/acer.12289 (2014).
Horrigan, F. T. & Aldrich, R. W. Allosteric voltage gating of potassium channels II. Mslo channel gating charge movement in the absence of Ca(2+). Journal of General Physiology 114, 305–336 (1999).
Geng, Y. & Magleby, K. L. Single-channel kinetics of BK (Slo1). channels. Frontiers in physiology 5, 532, https://doi.org/10.3389/fphys.2014.00532 (2014).
Shelley, C., Niu, X., Geng, Y. & Magleby, K. L. Coupling and cooperativity in voltage activation of a limited-state BK channel gating in saturating Ca2+. Journal of General Physiology 135, 461–480, https://doi.org/10.1085/jgp.200910331 (2010).
Saheki, Y. & Bargmann, C. I. Presynaptic CaV2 calcium channel traffic requires CALF-1 and the alpha(2)delta subunit UNC-36. Nature neuroscience 12, 1257–1265, https://doi.org/10.1038/nn.2383 (2009).
Ghezzi, A., Krishnan, H. R. & Atkinson, N. S. Susceptibility to ethanol withdrawal seizures is produced by BK channel gene expression. Addict Biol 19, 332–337, https://doi.org/10.1111/j.1369-1600.2012.00465.x (2014).
N’Gouemo, P. & Morad, M. Alcohol withdrawal is associated with a downregulation of large-conductance Ca(2)(+)-activated K(+) channels in rat inferior colliculus neurons. Psychopharmacology 231, 2009–2018, https://doi.org/10.1007/s00213-013-3346-8 (2014).
Kreifeldt, M., Le, D., Treistman, S. N., Koob, G. F. & Contet, C. BK channel beta1 and beta4 auxiliary subunits exert opposite influences on escalated ethanol drinking in dependent mice. Frontiers in integrative neuroscience 7, 105, https://doi.org/10.3389/fnint.2013.00105 (2013).
Scott, L. L. et al. A Novel Peptide Restricts Ethanol Modulation of the BK Channel In Vitro and In Vivo. The Journal of pharmacology and experimental therapeutics 367, 282–290, https://doi.org/10.1124/jpet.118.251918 (2018).
Kaufmann, W. A. et al. Large-conductance calcium-activated potassium channels in purkinje cell plasma membranes are clustered at sites of hypolemmal microdomains. J Comp Neurol 515, 215–230, https://doi.org/10.1002/cne.22066 (2009).
Kaufmann, W. A., Kasugai, Y., Ferraguti, F. & Storm, J. F. Two distinct pools of large-conductance calcium-activated potassium channels in the somatic plasma membrane of central principal neurons. Neuroscience 169, 974–986, https://doi.org/10.1016/j.neuroscience.2010.05.070 (2010).
Mello, C. & Fire, A. DNA transformation. Methods in cell biology 48, 451–482 (1995).
Davies, A. G., Blackwell, G. G., Raabe, R. C. & Bettinger, J. C. An Assay for Measuring the Effects of Ethanol on the Locomotion Speed of Caenorhabditis elegans. Journal of visualized experiments: JoVE, https://doi.org/10.3791/52681 (2015).
Alaimo, J. T. et al. Ethanol metabolism and osmolarity modify behavioral responses to ethanol in C. elegans. Alcoholism, clinical and experimental research 36, 1840–1850, https://doi.org/10.1111/j.1530-0277.2012.01799.x (2012).
Acknowledgements
This work is supported by NIH Grant R01GM125749 and R21AA023940 (H.K.). Some strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440).
Author information
Authors and Affiliations
Contributions
K.O. and H.K. performed all of the experiments and wrote the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
41598_2019_46615_MOESM1_ESM.mp4
<Emphasis Type="Italic">ctn-1(eg1167)</Emphasis> mutant animals exhibit ethanol-resistant locomotory behavior comparable to that of <Emphasis Type="Italic">slo-1(eg142)</Emphasis> mutant animals.
41598_2019_46615_MOESM2_ESM.mp4
Low-level fluorescent SLO-1 signals in the dorsal cord of <Emphasis Type="Italic">ctn-1(eg1167)</Emphasis> mutant animals are highly mobile, while clustered SLO-1 channels in the dorsal cord of wild-type animals are stable.
41598_2019_46615_MOESM3_ESM.pdf
BK channel clustering is required for normal behavioral alcohol sensitivity in <Emphasis Type="Italic">C. elegans</Emphasis>
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Oh, K.H., Kim, H. BK channel clustering is required for normal behavioral alcohol sensitivity in C. elegans. Sci Rep 9, 10224 (2019). https://doi.org/10.1038/s41598-019-46615-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-46615-9
This article is cited by
-
Transcriptional analysis of the response of C. elegans to ethanol exposure
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
