
Abstract
Fertilization and early embryo development are regulated by a unique maternal-gamete/embryo cross-talk within the oviduct. Recent studies have shown that extracellular vesicles (EVs) within the oviduct play important roles in mediating this developmental process. Here, we examined the influence of oviductal EVs on sperm function in the domestic cat. We demonstrated that (1) EVs are enriched in proteins related to energy metabolism, membrane modification, and reproductive function; (2) EVs bound and fused with the membranes of the acrosome and mid piece; and (3) incubating sperm with EVs improved motility, fertilizing capacity of cat spermatozoa and prevented acrosomal exocytosis in vitro. These findings indicated that oviductal EVs mediate sperm function and fertilization in the cat and provides new insights to improve sperm cryopreservation and in vitro fertilization in the domestic and wild felids and human.
Similar content being viewed by others

Introduction
Due to the loss of natural habitat1, nearly half of wild felid species are listed as threatened by the International Union for Conservation of Nature (IUCN, 2018)2. Many of these species are kept in zoos and breeding centers for public awareness, research purposes and/or as security populations3, but successful management of small populations is threatened by losses in genetic diversity due to poor reproduction and/or health. Assisted reproductive technologies (ARTs), including artificial insemination and in vitro fertilization (IVF) have been applied to retain gene diversity and avoid inbreeding depression4. Although live offspring have been produced by IVF in a number of wild felids, including the Indian desert cat, African wildcat, serval, caracal, fishing cat, black-footed cat, tiger and the sand cat5, this technology has not been widely used in breeding programs, partly due to limited information on species-specific reproductive endocrinology, gamete biology and embryogenesis6. Furthermore, live birth rates from in vitro derived embryos is low in these species, varying from 0 to 40%7. Therefore, there is a critical need to establish improved in vitro conditions supportive of normal fertilization and embryonic development.
The oviduct (or fallopian tube in the human) provides the physiological environment that is essential for sperm storage and capacitation8,9,10,11,12,13,14,15, fertilization11,16,17,18 and embryo development19,20,21,22. After mating, sperm travel into the uterus, enter the oviduct and attach to the epithelium of the isthmus, a process which is necessary for extending sperm viability within the female reproductive tract23. Within the isthmus, sperm acquire hyperactivated motility and the ability to fertilize an oocyte (a process known as ‘capacitation’)12. It has been demonstrated that the oviduct facilitates gamete function and fertilization by the secretion of extracellular vesicles (EVs)21,24,25. EVs are membrane encapsulated particles containing regulatory molecules that contribute to cell-cell communication by carrying proteins, peptides, RNA species (microRNAs, mRNAs, and long non-coding RNAs), lipids, and DNA fragments26,27,28. EVs have been isolated from prostate, epididymal, uterine, follicular and oviductal fluids21,24,29,30,31,32,33,34,35,36. Yet, only a hand full of studies have investigated the role of oviductal EVs on gamete maturation and embryo production21,24,32,37,38,39. Previous studies have shown that the presence of oviductal EVs during embryo culture increases blastocyst rate and improves embryo quality during in vitro incubation in cattle21. Incubation of mouse sperm with oviductal fluid or exosomes increases a Ca2+ efflux pump protein, PMCA4, which is important for the acquisition of hyperactivated motility and fertilizing ability24. Recently, it also has been shown that oviductal EVs supplemented in embryo transfer medium increased birth rates after transferring in vitro derived embryos in the mouse38.
To date, there are no studies about the effects of oviductal EVs on sperm function and its fertilizing ability in vitro. Because of its importance as a biomedical model and for the applicability of these data to wild felids4,40,41,42 and humans40, we isolated cat oviductal EVs and investigated their protein content, interaction with sperm and influence on sperm function.
Results and Discussion
Exosomes are the most abundant EVs in the cat oviduct
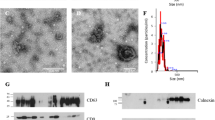
Previous studies have developed methods that allow consistent yield of high quality and quantity EVs26,43,44. In this study, we used Total Exosome Isolation Kit (Invitrogen, USA) to recover EVs from cat oviductal fluid. This method was shown to be superior to commonly used ultracentrifugation protocols in terms of EVs recovery yield, without changes in EVs quality44. Transmission electron microscopy (TEM) analysis showed the presence of EVs with sizes ranging from 40–150 ηm (Fig. 1a), and ZetaView nanoparticle tracking analysis (NTA) confirmed an average EV size of 129.7 ± 89.4 ηm. Overall, EV concentrations were similar among cats, except for one donor (Ovi EVs 3), from which higher EV levels were recovered (Table 1). The concentration of EVs ranged from 1.4 × 109 to 8.6 × 109 particles mL−1 (Fig. 1b and Table 1). Statistical analysis (ANOVA, F(4) = 33.474, p = 0.01, n = 5) revealed that there were significant differences in the concentration of EVs collected among donor cats and post hoc evaluations showed that the level observed in cat 3 was significantly different from cats 2, 4 and 5 (Tukey test p = 0.022, 0.03 and 0.048, respectively). Although there are large variations in EV size, (15–895 ηm), the majority (74%) of EVs were 40–150 ηm in diameter which is typical for exosomes44 (Fig. 1b and Table 1).

Transmission electron microscopy (TEM) and size distribution of cat oviductal EVS. (a) TEM image of oviductal EVs (stars); scale bar depicts 200 nm. (b) Size distribution of oviductal EVs measured by nanoparticle tracking analysis using the ZetaView. EVs were collected from five cats (Ovi1-5) the quadratic interpolation of the mean number of particles isolated from the oviducts of each individual donor cat is represented in the graph.
It’s known that steroids hormones influence the oviductal environment, promoting changes on its transcriptome and proteome, both in vivo and in vitro45,46,47. EVs were previously identified in the bovine oviductal fluid at different stages of the estrous cycle, and demonstrated that their composition is also under hormonal regulation25. For bovine, RNA-sequencing identified 903 differentially expressed transcripts in EVs across the estrous cycle25. Moreover, from the 336 clusters of EVs proteins, 170 were differentially abundant among stages of the estrous cycle25. In the present study, we collected oviductal EVs from cats during the follicular phase of reproductive cycle (neither corpora hemorrhagica nor corpora lutea was present). Therefore, it is unlikely that the high EV particles observed in cat 3 was due to a variation in reproductive stage. Trauma or inflammation can increase the number of EVs secreted in the plasma48, likewise, the increased EV number in cat 3 might have been due to an injury or pathology that was not visibly detected during oviduct flushing.
Oviductal EVs protein content
A total of 1,511 protein groups were identified, out of 4,879 protein entries (Supplementary Data 1). The numbers of proteins identified in cat EVs were more than three-times higher than the amount previously observed in bovine oviductal exosomes and fluids (315 and 482 total proteins, respectively)21,46, but less than the level reported in the human (5,177 total proteins)49. The difference in the amount of proteins between bovine and cat oviductal EVs may be due to the mass spectrometry method used in the two studies. In the present study, oviductal EVs were analyzed using ultraperformance liquid chromatography and tandem mass spectrometry (UPLC-MS/MS). In the bovine study, the samples were analyzed by SDS-PAGE combined with nanoLC–MS/MS. It has been shown that improved sensitivity and efficiency of UPLC-MS/MS increases the ability to detect different proteins isoforms, with higher quantification limits50. Moreover, a single-pot, solid-phase-enhanced sample preparation (SP3) for protein isolation was used, which has been shown to have a high recovery of protein input51.
In the cat oviductal EVs, specific EVs markers, such as transmembrane- or lipid-bound extracellular proteins (CD9, CD63, CD109, CADM1, ITGA3, ITGA6, ITGB1, ITGB3, ITGB4, ESP8, and MFGE8), and cytosolic proteins (TSG101, ANXA1-7, ANXA11, RAB1A, RAB1B, RAB2A, RAB5B, RAB5C, RAB6A, RAB6B, RAB10, RAB11A, RAB11B, EEA1, and SDCBP) were detected, indicating the EV origin43 of the samples analyzed. Furthermore, two functionally grouped gene ontology (GO) cellular component pathways related to EVs were identified: (1) extracellular exosome (GO:0070062) and (2) extracellular vesicle (GO:1903561); these pathways had 709 and 713 total proteins respectively, with a fold enrichment of 2.64 each.
Functional analysis using PANTHER database revealed that the identified proteins were part of different GO biological processes, including metabolism (30.2%), cellular component organization (18.5%), biological regulation (17.1%), localization (15.1%), cellular function (10.3%), response to a stimulus (4.7%), multicellular organism process (3.9%), development (3.6%), biological adhesion (2.5%), signaling (1.3%), immune system (1.1%), reproduction (0.7%), locomotion (0.4%), multi organism process (0.2%), and growth (0.1%). These pathways were similar to the ones identified in bovine oviductal EVs proteins21. Related to the potential role of oviductal EVs on sperm characteristics and fertilizing ability, three main clusters of GO molecular function, cellular components and biological process pathways were recognized by DAVID database: These include pathways related to (1) energy metabolism (Fig. 2a); (2) membrane modification (Fig. 2b); and (3) reproductive function (Fig. 2c). Supplementary Data 2 shows all GO molecular functions, cellular components and biological process pathways identified.

GO pathways related to the potential role of oviductal EVs on sperm characteristics and fertilizing ability. GO molecular function (green), cellular components (yellow) and biological process (blue) pathways related to (a) membrane modification, (b) energy metabolism, and (c) reproductive function.
Proteins related to sperm motility (ATP1A4, ATP2B4, DNAJA1, APOB, CCDC39, CCDC40, DPCD, DNAH1, DNAH5, NPHP4, PGK2, PLTP, TEKT2, and TEKT3), sperm-egg recognition (NME5, NME7, NME9, NME1, CAD, MTOR, and UPRT), and sperm binding to zona pellucida (CCT2, CCT3, CCT4, CCT5, CCT7, CCT8, OVGP1, and TCP1) were included in the functionally grouped GO pathways determined by DAVID. A diagram of proteins and their network pathways can be seen in Supplementary Fig. 1.
Oviductal EVs bind to sperm acrosome and mid-piece
Small EVs (40–150 ηm; exosomes) facilitate the transportation of proteins, microRNAs and other factors from luminal fluids to the sperm surface24,34,35,36,52. We used fluorescence microscopy and TEM to investigate the interaction between oviductal EVs and cat spermatozoa. Incubating cat epididymal sperm with BODIPY® TR Ceramide labeled EVs (1:2 v/v sperm to EVs, n = 3) for 1 hour resulted in a higher percentage of EVs bound spermatozoa (>96%) than samples incubated for 15 (64%) and 30 (84%) minutes (paired sample t-test, p = 0.02 and 0.04, respectively). Cat oviductal EVs bound to the acrosomal region of the sperm head and mid-piece (Fig. 3a). TEM confirmed the fusion of 60 to 180 ηm EVs to the outer acrosomal membrane and mid piece (n = 2, Fig. 3b–e). Bound vesicles were not detected in samples analyzed in the absence of oviductal EVs. This binding pattern is similar to that described previously for mouse sperm incubated with oviductal, epididymal and uterine EVs32,53.

Uptake of oviductal EVs by spermatozoa. (a) Sperm incubated with BODIPY® TR Ceramide labeled oviductal EVs, note that EVs bind to sperm acrosome (arrow) and mid-piece (dashed box). (b–e) TEM images of sperm incubated with EVs (black arrows), in (b,c) EVs bound to sperm head, showing nucleus (N) and acrosome (A); and on (d,e) EVs bound to sperm mid-piece, showing axoneme (Ax) and mitochondria (M). Scale bar: yellow = 15 μm, black = 200 ηm.
Incubation with oviductal EVs increased sperm motility and sustained acrosomal integrity
The functionality of a spermatozoon generally depends on its (1) progressive motility54, (2) acrosomal integrity55, and (3) the ability to undergo capacitation and acrosomal exocytosis56. It is known that the oviductal epithelium influences sperm function as well as supports gamete interaction and early embryo development9,12,18,57,58,59,60. A previous study has shown that human sperm sustain motility longer when they were incubated with homologous oviductal epithelium than those without co-incubation61. Here, we investigated whether oviductal EVs could sustain cat sperm motility in vitro. Sperm were incubated for 1 h with EVs collected from cat oviducts (n = 5 cats). Incubation of sperm with EVs sustained a greater percentage of motile sperm (paired sample t-test, p = 0.016, 0.000, 0.030, and 0.018, for 1, 2, 18 and 24 h, respectively; n = 5; Fig. 4a) than those without EVs, and this trend was maintained throughout the 24-hour incubation. These results are in agreement with previous studies in which incubating bovine sperm with oviductal fluid improved sperm motility for as long as 6 hours9. Incubation of cat spermatozoa with oviductal EVs did not affect progressive motility (Fig. 4b) and only increased hyperactive motility at 1 h post-incubation (paired sample t-test, p = 0.007; Fig. 4c).

Total (a), progressive (b) and hyperactive (c) motility of sperm incubated with or without oviductal extracellular vesicles (EVs) at 0, 1, 2, 18 and 24 hours. Symbol indicates statistically significant difference (p < 0.05, paired samples t-test) for sperm + EVs vs Sperm for each time point (n = 5).
Sperm acquires energy from molecules present in the seminal plasma and in the female reproductive tract62. As in other mammalian cells, most of this energy is transformed into adenosine tri-phosphate (ATP), which is critical for flagella movement, membrane fusion events during the acrosome reaction, and transportation of ions and other molecules through the sperm membranes62. We demonstrated that, similar to bovine21, cat oviductal exosomes carry proteins related to energy metabolism, such as ATP synthase, V-type proton ATPase subunit E, ATP binding, plasma membrane calcium-transporting ATPase 2, ADP/ATP translocase 1, mitochondrial ATP synthase, cytochrome c oxidase, acyl-CoA dehydrogenase, succinate-CoA ligase [ADP/GDP-forming] subunit alpha, NADPH, ATPase Na+/K+ transporting, ATPase plasma membrane Ca2+ transporting, among others. Because energy production is essential for sperm flagella movement, we speculated that these proteins from cat oviductal EVs can be delivered to the sperm mid-piece and could explain the higher sperm motility observed. Moreover, HSP70, OVGP1 and different apolipoproteins (APOA1, APOB, APOE) were also present in the EVs, which were previously reported to be positively correlated with sperm motility in bulls and human63,64,65. Cumulative findings from our and previous studies suggests that HSP70, OVGP1 and apolipoproteins may also contribute to the higher motility observed in the cat sperm incubated with oviductal EVs.
Another requisite event for fertilization in mammals is the ability of spermatozoa to undergo acrosomal exocytosis (AE)66. AE starts during the sperm transit through the oviduct, and is modulated by the oviductal and follicular fluids, cumulus oophorous cells and the zona pellucida9,55,66. In contrast to the previous reports in cows and human, in which oviductal fluid induced AE9,66, incubating cat sperm with oviductal EVs prevented in vitro acrosome exocytosis of spermatozoa incubated under a non-capacitation condition. The percentages of cat sperm with intact acrosome after 30 min incubation in PBS were 34 ± 3.1 and 23 ± 4.7% for samples incubated with and without EVs, respectively, both of which were less than 49 ± 3.7% observed at 0 hour incubation (p = 0.0041 and 0.0003, respectively; n = 5, paired samples t-test) (Fig. 5). Extending the incubation period to 1 hour significantly decreased the percentage of sperm with intact acrosome in the absence of EVs (P = 0.03), whereas acrosome integrity of EVs exposure samples remained constant throughout the culture period. We have shown in the earlier experiment that the ability of EVs to bind to sperm plasma membrane is time dependent with maximum binding (96%) observed at 1 hour incubation. Therefore, the reduction in acrosome integrity in the presence of EVs during the first 30 min is likely due to insufficient time for EVs to bind to the sperm plasma membrane to fully exert their protective effect. The domestic cat is an induced ovulator, with ovulation occurring within 25–50 h after mating67. As early as 30 min after mating, spermatozoa can be observed in the oviduct of the cat68. Cat sperm must maintain acrosomal integrity for 25–50 h within the female reproductive tract for them to successfully fertilize an oocyte. Therefore, we hypothesize that oviductal EVs, especially during the preovulatory period, play roles in preventing sperm from undergoing acrosomal exocytosis prematurely.

(a) Percentages of sperm with intact acrosome in samples incubated with or without oviductal EVs for 0, 0.5,1, 2 and 3 hours. There were statistically significant differences (paired samples t-test) between the two treatments at 1, 2 and 3 h (p = 0.0001, 0.0001 and 0.0188, respectively; n = 5). In (b), sperm with an intact (arrow) or non-intact (arrow heads) acrosome stained with FITC-PNA. Scale bar = 20 μm.
Steps for capacitation, including acquisition of hyperactive motility and acrosome reaction, are also dependent on the lipid composition of the sperm membrane and its interaction with membrane proteins69. Oviductal EVs were rich in proteins that are part of important membrane modification processes, including, but not limited to, regulation of protein localization to plasma membrane (GO:1903076), membrane raft organization (GO:0031579), and regulation of cholesterol metabolic process (GO:0090181), which could also influence the differences observed in sperm function. Furthermore, sperm membrane lipid composition is also correlated with survival success after cryopreservation70. Significant sperm membrane lipid changes are associated with freezing/thawing procedures71. Incubating sperm with oviductal EVs, or specific EVs factors, prior to cryopreservation or during thawing procedure could influence the sperm membrane lipids composition and improve its functionality post-thawing.
Oviductal EVs improve sperm’s fertilizing ability
Here we examined the effect of cat oviductal EVs on IVF and embryo development. Cumulus-oocyte complexes (COCs) were inseminated with fresh spermatozoa that was previously incubated for 1 h with or without oviductal EVs. Our data revealed that incubating cat sperm with EVs increased cleavage rate by 23% compared to the non-EV samples (General linear model - GLM, p = 0.013; Fig. 6). As a consequence of a higher cleavage rate, a 12% increase in the number of >8 cell-embryos at Day 4 and an 8% increase in blastocyst rates were observed (GLM, p = 0.027 and p = 0.014, respectively). Cat oviductal exosomes contain proteins known to be important for fertilization, such as OVGP1 (improves sperm viability and motility in vitro)65, CD9, TCP1, CCT3, CCT4, CCT7, CCT8, (important for sperm-oocyte binding)72,73, HSP70 (improves sperm survival in vitro)74 and HSP90 (mediates sperm hyperactivation and acrosome exocytosis)21,75. The transfer of these proteins from the oviductal EVs to the spermatozoa could explain the increased fertilization rates observed.

Effect of sperm incubation with (Sperm + EVs) or without (Sperm) oviductal extracellular vesicles (EVs) on fertilization rate (cleavage), percentage of embryos of 8 or more cells (>8-cells) and blastocyst rate (blastocyst). Symbol indicates statistically significant difference (General Linear Model, after correction for male effect) for sperm + EVs vs Sperm (p = 0.0013, 0.027 and 0.014 for cleavage, >8 cells and blastocyst respectively).
Another known benefit of the oviductal environment is the reduction of polyspermic fertilization18,76,77. In this study, no difference in polyspermy was detected between samples incubated with oviductal EVs and no-EVs control (17 ± 9 vs 11 ± 5%, sperm and sperm + EVs respectively; GLM, p = 0.097). The incidence of polyspermy in cats is similar to that seen in the human (ranging from 3 to 30%) but it is lower than the rate observed in the pig (ranging from 14 to 93%)76. Our results suggest that the prevention of polyspermy in the cat likely depends on the effects of the oviductal environment on the oocyte. Indeed, incubating oocytes with oviductal fluid or its specific protein, the oviductal glycoprotein 1 (OVGP1) reduced polyspermy rates in the pig77,78. Recently, it was also shown that the use of a 3D culture system that mimics the oviductal microenvironment during IVF, significantly reduces polyspermy rates in the cow18. Future studies should examine the effect of incubating oocytes with oviductal EVs on the incidence of polyspermic fertilization in the domestic cat.
Conclusions
This study showed that exosomes were the most abundant oviductal EVs in cats, and that they bind to the acrosomal region of the sperm head as well as the mid-piece. We also demonstrated that incubating sperm with EVs enhanced and sustained motility and acrosome integrity as well as improved fertilizing ability of cat epididymal sperm. The increased motility could be due to the transfer of important proteins from EVs to sperm, such as HSP70, OVGP1, apolipoproteins and proteins related to energy metabolism. Furthermore, increased fertilization was likely due to proteins related to sperm-oocyte recognition and binding, such as CD9, TCP1, and CCTs. The use of oviductal EVs in vitro has the potential to improve ART results for domestic and wild felids especially in individuals/species exhibiting poor seminal characteristics. Moreover, the findings serve as the foundation for future studies aimed at identifying oviductal factors that enhance sperm survival and fertilizing ability. Such studies offer promising avenues for the development of strategies for long-term sperm preservation and for designing improved in vitro culture conditions for IVF and embryo development.
Methods
All male and female reproductive tracts were opportunistically collected from local veterinary clinics after routine neutering and spaying procedures of household and stray cats. No additional permissions were required since these biological materials were designated for disposal via incineration.
Oviductal extracellular vesicles (EVs) isolation
Oviducts of domestic cats (7 months to 4 years old, n = 16) in follicular phase (ovaries without the presence of corpora hemorragica and/or lutea;) were recovered after routine ovariohysterectomy at local veterinary clinics and transported (at 4 °C) to the laboratory within 6 h of excision. After being washed three times in Phosphate Buffer Saline solution (PBS, GIBCO, USA), a 23G needle was inserted through the fimbria opening and the entire oviduct was flushed with 1 ml of PBS. The flush was centrifuged at 2,000 × g (room temperature) for 30 minutes to remove cells and debris, then the supernatant was mixed with 500 μL of the Total Exosome Isolation Reagent (Invitrogen, USA) and incubated overnight at 4 °C. The samples were centrifuged at 10,000 × g for 1 hour, and the pellet resuspended in 50 μL of PBS. EVs were then purified by Exosome Spin Columns (Invitrogen, USA), following manufacturer’s instructions and kept at −20 °C until use.
Oviductal EVs quantification
Nanoparticle tracking analysis was done using the ZetaView S/N 17–332 (Particle Metrix, Meerbusch, Germany) and data analyzed using its software (ZetaView 8.04.02) by Alpha Nano Tech (Research Triangle Drive, NC, USA) as previously described44. For each donor cat (n = 5), oviductal EVs sample (1 ml) was diluted 100X in PBS, loaded into the cell, and the instrument measured each sample at 11 different positions throughout the cell, with three cycles of readings at each position. The pre-acquisition parameters were: sensitivity of 85, frame rate of 30 frames per second (fps), shutter speed and laser pulse duration of 100, temperature of 19.81 °C, and pH of 7.0. Post-acquisition parameters were set to a minimum brightness of 22, a maximum area of 1000 pixels, and a minimum area of 10 pixels44. All parameters (temperature, conductivity, electrical field, and drift measurements) were documented for quality control. After software analysis, the mean, median, and mode (indicated as diameter) sizes, as well as the concentration of the sample, were calculated, excluding outliers44. The number of particles per particle size curves was created using quadratic interpolation44.
Oviductal EVs proteomics
Oviductal EVs (n = 3 cats, a total of 6 oviducts, in follicular phase) were pooled, frozen at −20 °C and sent to Bioproximity LLC (Chantilly, VA) on dry ice for protein extraction and proteomic analysis, according to the company’s protocols. Protein samples were prepared using a single-pot, solid phase-enhanced sample-preparation (SP3) technology51.
Analysis was performed using ultraperformance liquid chromatography and tandem mass spectrometry (UPLC - Thermo Easy-nLC 1200 fitted with a heated, 25 cm Easy-Spray column – MS/MS - Thermo Q-Exactive HF-X quadrupole-Orbitrap mass spectrometer). The peptide dataset (mzML format) were exported to Mascot generic format (.mgf) and searched using X!!Tandem79 using both the native and k-score scoring algorithms80, and by OMSSA81. RAW data files were compared with the protein sequence libraries available for the domestic cat (Felis catus, taxa 9685). Label free quantification (MS1-based) was used and peptide peak areas were calculated using OpenMS82. Proteins were required to have one or more unique peptides across the analyzed samples with E-value scores of 0.01 or less.
Functional GO clustering
Data Entrez Gene IDs were mapped for all identified proteins using the R package rentrez (ver 1.2.1)83. The background dataset for all analyses was the cat (Felis catus) genome. Gene ontology (GO) analyses were performed using DAVID (https://david.ncifcrf.gov)84 and PANTHER (http://pantherdb.org)85 web-based software. For enrichment analysis, the cut off was set to p < 0.05. The Cytoscape 3.5.1 plugin ClueGO86 was used to visualize interactions of EVs proteins and networks integration, by GO terms “biological processes” and “cellular components” using the Felis catus genome. The evidence was set to “Inferred by Curator (IC)”, and the statistical test was set to a right-sided hypergeometrical test with a κ score of 0.7–0.9 using Bonferroni (step down). The function “GO Term fusion” was selected, the GO term restriction levels were set to 3, and a minimum of three genes or 5% genes in each GO term was used.
Oviductal EVs labeling
Oviductal EVs (n = 3 cat oviducts) were stained by phospholipid bilayer BODIPY® TR Ceramide (Invitrogen, USA). In brief, TR Ceramide stock solution (dissolved in DMSO) was added to 50 μl of total EVs (final concentration of 5 µM) and incubated for 30 minutes at room temperature. Labeled EVs were then purified to remove free dye by Exosome Spin Columns (Invitrogen, USA), following manufacturer’s instructions. Labeled EVs were immediately incubated with sperm as described below.
Epididymal sperm isolation and incubation with oviductal EVs
Testes from domestic cats (8 months to 4 years old, n = 15) were recovered after routine neutering at local veterinary clinics and transported (at 4 °C) to the laboratory within 6 h of excision. Corpus and caudal epididymides as well as vas deferens were isolated and cleaned off blood vessels. Transversal dissections of the epididymis were performed and the epididymides were immersed in PBS after which the vas deferens were squeezed and sperm were allowed to swim out for 5 minutes at 38 °C. PBS containing spermatozoa was collected, centrifuged at 700 × g for 5 minutes, and supernatant was discarded. The sperm pellet was then suspended in PBS to achieve the concentration of 1 × 106 sperm/mL and incubated with oviductal EVs or PBS (1 volume of sperm:2 volumes of EVs/PBS, 2.26 ± 1.03 × 106 total particles) for 1 hour at 38 °C and 5% CO2 in air. After incubation, sperm suspension was pelleted by centrifuging at 700 x g for 5 minutes and: (1) fixed in 2% glutaraldehyde +4% paraformaldehyde in PBS and processed for transmission electron microscopy (TEM); (2) diluted to 5 × 106 sperm/mL in synthetic oviductal fluid (SOF) medium and used for sperm motility analyses; (3) kept in PBS and collected at 0, 0.5, 1, 2 and 3 h for acrosome integrity analysis; or (4) processed for in vitro fertilization. Sperm cells were also incubated with BODIPY labeled EVs (as described above), fixed in 4% paraformaldehyde for 15 minutes and visualized by a fluorescence microscope (EVOS FL auto 2, Invitrogen, USA) with a 1,000x Plan-Apochromat oil immersion objective to evaluate EVs bound to sperm cell.
Oviductal EVs and spermatozoa transmission electron microscopy
For transmission electron microscopy (TEM), the EVs suspensions (n = 2 cats) were fixed for 1 hour in 4% paraformaldehyde. EVs (5 μl) of each donor were dropped onto copper mesh Formvar coated carbon stabilized grids and allowed to be adsorbed to the grid for 4–5 min after which extra liquid was wicked off with filter paper44. A 2% Aqueous Uranyl Acetate (5 μl) solution was applied to the grid for 30 seconds (negative staining of exosomes), then whisked off with filter paper, and grids were air dried before imaging44. Epididymal sperm incubated with or without EVs (n = 4 cats) were fixed as described above and processed for TEM. The sperm-EVs samples preparation and imaging were performed at the Advanced Microscopy Laboratory at the University of Virginia (AMLUVA) and imaged on a Jeol JEM1230 transmission electron microscope (TEM) (40 kV) at AMLUVA.
Sperm motility and acrosomal analyses
Sperm samples from five cats (three replicates) were incubated at 38.5 °C in SOF medium +1000 IU of Penicillin, 10 μg/ml streptomycin, 10 μg/ml of heparin, 20 µM penicillamine, 10 µM hypotaurine, 1 µM epinephrine for up to 24 h and samples collected at 0, 1, 2, 18, and 24 h for motility analysis; or in PBS with or without oviductal EVs and samples collected at 0, 0.5, 1, 2 and 3 h for acrosome integrity. For motility analysis, 10 μl of sperm samples were dropped onto a glass slide, covered with a coverslip and observed at a magnification of 200x on a positive phase-contrast microscope (Leika Leitz DM1L; Wetzlar, Germany) with a warmed (38 °C) stage. Ten fields per sample were recorded using a Nikon DS-F camera (Japan), and a minimum of 100 spermatozoa were analyzed in each sperm sample for sperm total motility (percentage of moving spermatozoa), hyperactive (star spin moving spermatozoa) and progressive (forward moving spermatozoa) motility analyses. For acrosome integrity assessment, 10 μl of sperm were fixed in 10 μl of 4% paraformaldehyde for 15 min and labeled with HOECHST33342 (for DNA) and the acrosome-specific fluorochrome fluorescein isothiocyanate-labeled peanut (Arachis hypogaea) agglutinin (FITC-PNA). At least one hundred spermatozoa were counted at 1,000x magnification (EVOS FL auto 2, Invitrogen, USA),
In vitro maturation, In vitro fertilization and embryo culture
Ovaries from adult domestic cats were recovered after routine ovariohysterectomy at local veterinary clinics and transported (at 4 °C) to the laboratory within 6 h of excision. Cumulus cell–oocyte complexes (COCs) were mechanically isolated into HEPES (Sigma-Aldrich, USA)-buffered minimum essential medium (MEM; Life Technologies, USA) supplemented with 2 mM L-glutamine, 1 mM pyruvate (Sigma-Aldrich, USA), 100 IU/ml penicillin (Sigma-Aldrich, USA), 100 μg/ml streptomycin (Sigma-Aldrich, USA) and 4 mg/ml bovine serum albumin (BSA; Sigma-Aldrich, USA). COCs (n = 429) were incubated in 50 μl microdrops (15–25 COCs per drop) of IVM medium consisting of 1 μg/ml FSH (Vetoquinol, USA) and 1 μg/ml porcine LH (National Hormone and Pituitary Program, USA) in SAGE blastocyst medium87. After a 24 h in vitro maturation (38 °C in 5% CO2), COCs were inseminated with 1 × 106 sperm/ml of fresh domestic cat epididymal spermatozoa incubated with (n = 231 oocytes) or without (n = 198 oocytes; Control) oviductal EVs. At 24 h post-insemination, oocytes were denuded and cleaned by gentle pipetting. Presumptive zygotes were cultured (38 °C in 5% CO2) in 50 μl droplets of SAGE blastocyst medium (15–25 embryos per drop) for up to 8 days. The numbers of embryos developing to 2–4 cell, (cleavage), >8 cell, and blastocyst stage were recorded on Day 2, 4 and 7 post-IVF, respectively using an inverted light microscope at a 200x magnification (Leika Leitz DM1L; Wetzlar, Germany).
In a separate study, IVF was performed as described and presumptive zygotes were collected 24 hours post fertilization and fixed in 4% paraformaldehyde. The fixed zygotes were stained with HOECHST33342 (5 μg/mL) for 30 min and visualized under a fluorescence microscope (EVOS FL auto 2, Invitrogen, USA) at a 400× magnification. The zygotes were classified as one of the two following categories: normal fertilization (presence of two pronuclei or one maternal pronuclei and one sperm head, inside the oolemma, plus the detection of two polar bodies in the perivitelline space) or polyspermic zygotes (presence of three or more pronuclei or one maternal pronucleus and multiple sperm heads, inside the oolemma plus the detection of two polar bodies in the perivitelline space). Three replicates were performed and the total number of COCs used were 68 for the control and 87 for sperm incubated with exosomes.
Statistical analysis
The data were analyzed using IBM SPSS Statistics (version 24). A Shapiro-Wilk test was performed, and all data proved to be normally distributed. Data are presented as mean ± standard deviations. ANOVA followed by a Tukey test was used to compare EVs concentration among cats. A paired samples t-test with 95% confidence interval was used to compare motility and acrosomal integrity between samples incubated with and without exosomes. Effects of sperm-EVs incubation on polyspermy, cleavage, percentage of >8-cells and blastocyst rates (dependent variables) were analyzed by a General Linear Model (GLM) where incubation of sperm with/without oviductal EVs was the fixed factor, and the male used for fertilization was the co-variate factor.
Data Availability
The authors declare that all data supporting the findings of this study are available within the article, Supplementary Files, or from the corresponding author upon reasonable request. UPLC-MS/MS (mzML) file has been deposited in FIGSHARE database under https://doi.org/10.6084/m9.figshare.7837331 (https://figshare.com/s/85c874c0d1076a1d5894).
References
Loveridge, A. J., Wang, S. W., Frank, L. G. & Seidensticker, J. People and wild felids: conservation of cats and management of conflicts. In Biology and conservation of wild felids (Oxford University Press).
IUCN 2018. The IUCN Red List of Threatened Species. Version 2018–2, http://www.iucnredlist.org Downloaded on 14 November 2018.
Tribe, A. & Booth, R. Assessing the Role of Zoos in Wildlife Conservation. Hum. Dimens. Wildl. 8, 65–74 (2003).
Comizzoli, P. et al. Advances in reproductive science for wild carnivore conservation. Reprod. Domest. Anim. 44, 47–52 (2009).
Herrick, J. R. et al. In Vitro Fertilization and Sperm Cryopreservation in the Black-Footed Cat (Felis nigripes) and Sand Cat (Felis margarita)1. Biol. Reprod., https://doi.org/10.1095/biolreprod.109.081034 (2010).
Jewgenow, K. & Songsasen, N. Reproduction and Advances in Reproductive Studies in Carnivores. Advances in experimental medicine and biology, https://doi.org/10.1007/978-1-4939-0820-2_10 (Springer US, 2014).
Veraguas, D., Echeverry, D., Castro, F. O. & Rodriguez-Alvarez, L. Applied Biotechnologies in the Conservation of Wild Felids: In Vitro Embryo Production and Cellular Regenerative Therapies. In Big Cats (eds Shrivastav, A. B. & Singh, K. P.), https://doi.org/10.5772/intechopen.71311 (Intech Open, 2017).
Talevi, R. & Gualtieri, R. Molecules involved in sperm-oviduct adhesion and release. Theriogenology 73, 796–801 (2010).
Mcnutt, T. & Killian, G. Influence of Bovine Follicular and Oviduct Fluids on sperm Capacitation In Vitro. J. Androl. 12, 244–252 (1991).
Ellington, J. E. et al. Coculture of human sperm with bovine oviduct epithelial cells decreases sperm chromatin structural changes seen during culture in media alone. Fertil. Steril. 69, 643–9 (1998).
Pollard, J. W. et al. Fertilizing capacity of bovine sperm may be maintained by binding of oviductal epithelial cells. Biol. Reprod. 44, 102–107 (1991).
Rodriguez-Martinez, H. Role of the oviduct in sperm capacitation. Theriogenology 68, 138–146 (2007).
Parrish, J. J., Susko-Parrish, J. L., Handrow, R. R., Sims, M. M. & First, N. L. Capacitation of bovine spermatozoa by oviduct fluid. Biol. Reprod. 40, 1020–1025 (1989).
Kawakami, E., Kashiwagi, C., Hori, T. & Tsutsui, T. Effects of canine oviduct epithelial cells on movement and capacitation of homologous spermatozoa in vitro. Anim. Reprod. Sci., https://doi.org/10.1016/S0378-4320(01)00135-X (2001).
Zhang, B., Yin, T. L. & Yang, J. A novel microfluidic device for selecting human sperm to increase the proportion of morphologically normal, motile sperm with uncompromised DNA integrity. Anal. Methods 7, 5981–5988 (2015).
Hunter, R. H. Oviduct function in pigs, with particular reference to the pathological condition of polyspermy. Mol. Reprod. Dev. 29, 385–91 (1991).
Gabler, C. et al. Exploring cumulus-oocyte-complex-oviductal cell interactions: Gene profiling in the bovine oviduct. J. Physiol. Pharmacol. 59, 29–42 (2008).
Ferraz, M. A. M. M. et al. Improved bovine embryo production in an oviduct-on-a-chip system: Prevention of poly-spermic fertilization and parthenogenic activation. Lab Chip, https://doi.org/10.1039/C6LC01566B (2017).
Besenfelder, U., Havlicek, V. & Brem, G. Role of the oviduct in early embryo development. Reprod. Domest. Anim. 47, 156–163 (2012).
Enright, B. P. et al. Culture of in vitro produced bovine zygotes in vitro vs in vivo: implications for early embryo development and quality. Theriogenology 54, 659–73 (2000).
Almiñana, C. et al. Oviduct extracellular vesicles protein content and their role during oviduct–embryo cross-talk. Reproduction 154, 253–268 (2017).
Cebrian-Serrano, A. et al. Effect of the bovine oviductal fluid on in vitro fertilization, development and gene expression of in vitro-produced bovine blastocysts. Reprod. Domest. Anim. 48, 331–338 (2013).
Suarez, S. S. Mammalian sperm interactions with the female reproductive tract. Cell Tissue Res. 363, 185–194 (2016).
Al-Dossary, A. A., Strehler, E. E. & Martin-DeLeon, P. A. Expression and secretion of plasma membrane Ca2+ -ATPase 4a (PMCA4a) during murine estrus: Association with oviductal exosomes and uptake in sperm. PLoS One 8, 1–14 (2013).
Almiñana, C. et al. Deciphering the oviductal extracellular vesicles content across the estrous cycle: Implications for the gametes-oviduct interactions and the environment of the potential embryo. BMC Genomics 19, 1–27 (2018).
Raposo, G. & Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. Journal of Cell Biology, https://doi.org/10.1083/jcb.201211138 (2013).
Kalra, H., Drummen, G. P. C. & Mathivanan, S. Focus on extracellular vesicles: Introducing the next small big thing. International Journal of Molecular Sciences, https://doi.org/10.3390/ijms17020170 (2016).
Xu, R. et al. Extracellular vesicles in cancer - implications for future improvements in cancer care. Nat. Rev. Clin. Oncol. 15, 617–638 (2018).
Greening, D. W., Nguyen, H. P. T., Elgass, K., Simpson, R. J. & Salamonsen, L. A. Human Endometrial Exosomes Contain Hormone-Specific Cargo Modulating Trophoblast Adhesive Capacity: Insights into Endometrial-Embryo Interactions1. Biol. Reprod., https://doi.org/10.1095/biolreprod.115.134890 (2016).
Bidarimath, M. et al. Extracellular vesicle mediated intercellular communication at the porcine maternal-fetal interface: A new paradigm for conceptus-endometrial cross-talk. Sci. Rep., https://doi.org/10.1038/srep40476 (2017).
Burns, G. et al. Extracellular vesicles in luminal fluid of the ovine uterus. PLoS One, https://doi.org/10.1371/journal.pone.0090913 (2014).
Al-Dossary, A. A., Bathala, P., Caplan, J. L. & Martin-DeLeon, P. A. Oviductosome-sperm membrane interaction in cargo delivery: Detection of fusion and underlying molecular players using three-dimensional super-resolution structured illumination microscopy (SR-SIM). J. Biol. Chem. 290, 17710–17723 (2015).
Burns, G. W., Brooks, K. E. & Spencer, T. E. Extracellular Vesicles Originate from the Conceptus and Uterus During Early Pregnancy in Sheep1. Biol. Reprod. 94, 1–11 (2016).
Saez, F., Frenette, G. & Sullivan, R. Epididymosomes and prostasomes: their roles in posttesticular maturation of the sperm cells. J. Androl. 24, 149–154 (2003).
Rowlison, T., Ottinger, M. A. & Comizzoli, P. Key factors enhancing sperm fertilizing ability are transferred from the epididymis to the spermatozoa via epididymosomes in the domestic cat model. J. Assist. Reprod. Genet. 35, 221–228 (2018).
Frenette, G. & Sullivan, R. Prostasome-like particles are involved in the transfer of P25b from the bovine epididymal fluid to the sperm surface. Mol. Reprod. Dev. 59, 115–121 (2001).
Lopera-Vásquez, R. et al. Extracellular Vesicles from BOEC in In Vitro Embryo Development and Quality. PLoS One 11, e0148083 (2016).
Qu, P. et al. Extracellular vesicles derived from donor oviduct fluid improved birth rates after embryo transfer in mice. Reprod. Fertil. Dev., https://doi.org/10.1071/RD18203 (2018).
Bathala, P. et al. Oviductal extracellular vesicles (oviductosomes, OVS) are conserved in humans: Murine OVS play a pivotal role in sperm capacitation and fertility. Mol. Hum. Reprod., https://doi.org/10.1093/molehr/gay003 (2018).
Songsasen, N., Comizzoli, P., Nagashima, J., Fujihara, M. & Wildt, D. The Domestic Dog and Cat as Models for Understanding the Regulation of Ovarian Follicle Development In Vitro. Reprod. Domest. Anim. 47, 13–18 (2012).
Wildt, D. E., Comizzoli, P., Pukazhenthi, B. & Songsasen, N. Lessons from biodiversity - The value of nontraditional species to advance reproductive science, conservation, and human health. Mol. Reprod. Dev. 77, 397–409 (2010).
Comizzoli, P., Songsasen, N. & Wildt, D. E. Protecting and extending fertility for females of wild and endangered mammals. Cancer Treat. Res. 156, 87–100 (2010).
Lötvall, J. et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 3, 1–6 (2014).
Helwa, I. et al. A comparative study of serum exosome isolation using differential ultracentrifugation and three commercial reagents. PLoS One 12, 1–22 (2017).
Ferraz, M. A. M. M. et al. An oviduct-on-a-chip provides an enhanced in vitro environment for zygote genome reprogramming. Nat. Commun. 9, 4934 (2018).
Lamy, J. et al. Regulation of the bovine oviductal fluid proteome. Reproduction 152, 629–644 (2016).
Cerny, K. L., Garrett, E., Walton, A. J., Anderson, L. H. & Bridges, P. J. A transcriptomal analysis of bovine oviductal epithelial cells collected during the follicular phase versus the luteal phase of the estrous cycle. Reprod. Biol. Endocrinol. 13, 84 (2015).
Kuravi, S. J. et al. Changes in the pattern of plasma extracellular vesicles after severe trauma. PLoS One 12, e0183640 (2017).
Liu, Y. et al. Human fallopian tube proteome shows high coverage of mesenchymal stem cells associated proteins. Biosci. Rep. 36, e00297–e00297 (2016).
Reguant, J. et al. Comparative study of UPLC–MS/MS and HPLC–MS/MS to determine procyanidins and alkaloids in cocoa samples. J. Food Compos. Anal. 23, 298–305 (2010).
Hughes, C. S. et al. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 14, 68–85 (2019).
Machtinger, R., Laurent, L. C. & Baccarelli, A. A. Extracellular vesicles: Roles in gamete maturation, fertilization and embryo implantation. Hum. Reprod. Update 22, 182–193 (2016).
Griffiths, G. S., Galileo, D. S., Reese, K. & Martin-Deleon, P. A. Investigating the role of murine epididymosomes and uterosomes in GPI-linked protein transfer to sperm using SPAM1 as a model. Mol. Reprod. Dev. 75, 1627–36 (2008).
Drobnis, E. Z., Yudin, A. I., Cherr, G. N. & Katz, D. F. Hamster sperm penetration of the zona pellucida: kinematic analysis and mechanical implications. Dev. Biol. 130, 311–23 (1988).
Cummins, J. M. & Yanagimachi, R. Development of ability to penetrate the cumulus oophorus by hamster spermatozoa capacitated in vitro, in relation to the timing of the acrosome reaction. Gamete Res. 15, 187–212 (1986).
Yanagimachi, R. Fertility of mammalian spermatozoa: its development and relativity. Zygote 2, 371–2 (1994).
McCauley, T. C. Oviduct-Specific Glycoprotein Modulates Sperm-Zona Binding and Improves Efficiency of Porcine Fertilization In Vitro. Biol. Reprod. 69, 828–834 (2003).
Ferraz, M. A. M. M., Henning, H. H. W., Stout, T. A. E., Vos, P. L. A. M. & Gadella, B. M. Designing 3-Dimensional In Vitro Oviduct Culture Systems to Study Mammalian Fertilization and Embryo Production. Ann. Biomed. Eng., https://doi.org/10.1007/s10439-016-1760-x (2016).
Erikson, D. W., Way, A. L., Bertolla, R. P., Chapman, D. A. & Killian, G. J. Influence of osteopontin, casein and oviductal fluid on bovine sperm capacitation. Anim. Reprod. 4, 103–112 (2007).
Killian, G. J. Evidence for the role of oviduct secretions in sperm function, fertilization and embryo development. Anim. Reprod. Sci. 82–83, 141–153 (2004).
Murray, S. C. & Smith, T. T. Sperm interaction with fallopian tube apical membrane enhances sperm motility and delays capacitation. Fertil. Steril. 68, 351–357 (1997).
Visconti, P. E. Sperm Bioenergetics in a Nutshell1. Biol. Reprod. 87, 1–4 (2012).
Yan, G.-J. et al. Association of heat shock protein 70 with motility of frozen-thawed sperm in bulls. Czech J. Anim. Sci., https://doi.org/10.17221/8239-cjas (2015).
Akerlöf, E., Jörnvall, H., Slotte, H. & Pousette, A. Identification of apolipoprotein A1 and immunoglobulin as components of a serum complex that mediates activation of human sperm motility. Biochemistry 30, 8986–90 (1991).
Abe, H., Sendai, Y., Satoh, T. & Hoshi, H. Bovine oviduct‐specific glycoprotein: A potent factor for maintenance of viability and motility of bovine spermatozoa in vitro. Mol. Reprod. Dev. 42, 226–232 (1995).
Dejonge, C. J. et al. The Acrosome Reaction-Inducing Effect of Human Follicular and Oviductal Fluid. J. Androl. 141, 359–365 (1993).
Dawson, A. B. & Friedgood, H. B. The time and sequence of preovulatory changes in the cat ovary after mating or mechanical stimulation of the cervix uteri. Anat. Rec. 76, 411–429 (1940).
Chatdarong, K., Lohachit, C. & Linde-Forsberg, C. Distribution of spermatozoa in the female reproductive tract of the domestic cat in relation to ovulation induced by natural mating. Theriogenology 62, 1027–1041 (2004).
Flesch, F. M. & Gadella, B. M. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim. Biophys. Acta 1469, 197–235 (2000).
Mandal, R., Badyakar, D. & Chakrabarty, J. Role of Membrane Lipid Fatty Acids in Sperm Cryopreservation. Adv. Androl. 2014, 1–9 (2014).
Schiller, J., Arnhold, J., Glander, H. J. & Arnold, K. Lipid analysis of human spermatozoa and seminal plasma by MALDI-TOF mass spectrometry and NMR spectroscopy - effects of freezing and thawing. Chem. Phys. Lipids 106, 145–56 (2000).
Chen, M. S. et al. Role of the integrin-associated protein CD9 in binding between sperm ADAM 2 and the egg integrin α6β1: Implications for murine fertilization, 2–7 (1999).
Baker, M. A. et al. The Chaperonin Containing TCP1 Complex (CCT/TRiC) Is Involved in Mediating Sperm-Oocyte Interaction. J. Biol. Chem. 286, 36875–36887 (2011).
Elliott, R. M. A. et al. Effects of HSPA8, an evolutionarily conserved oviductal protein, on boar and bull spermatozoa, https://doi.org/10.1530/REP-08-0298 (2005).
Sagare-patil, V., Bhilawadikar, R., Galvankar, M. & Zaveri, K. Progesterone requires heat shock protein 90 (HSP90) in human sperm to regulate motility and acrosome reaction. J Assist Reprod Genet 90, 495–503 (2017).
Coy, P. & Avilés, M. What controls polyspermy in mammals, the oviduct or the oocyte? Biol. Rev. 85, 593–605 (2010).
Mondéjar, I., Martínez-Martínez, I., Avilés, M. & Coy, P. Identification of potential oviductal factors responsible for zona pellucida hardening and monospermy during fertilization in mammals. Biol. Reprod. 89, 67 (2013).
Kouba, A. J., Abeydeera, L. R., Alvarez, I. M., Day, B. N. & Buhi, W. C. Effects of the Porcine Oviduct-Specific Glycoprotein on Fertilization, Polyspermy, and Embryonic Development In Vitro 1. Biology OF Reproduction 63 (2000).
Bjornson, R. D. et al. X!! Tandem, an Improved Method for Running X! Tandem in Parallel on Collections of Commodity Computers (2008).
Maclean, B., Eng, J. K., Beavis, R. C. & Mcintosh, M. General framework for developing and evaluating database scoring algorithms using the TANDEM search engine. 22, 2830–2832 (2006).
Geer, L. Y. et al. Open Mass Spectrometry Search Algorithm. 958–964 (2004).
Sturm, M. et al. OpenMS – An open-source software framework for mass spectrometry. 11, 1–11 (2008).
Winter, D. J. Rentrez: An R package for the NCBI eUtils API. R J. 9, 520–526 (2017).
Huang, D. W., Sherman, B. T. & Lempicki, R. A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57 (2009).
Mi, H., Muruganujan, A., Casagrande, J. T. & Thomas, P. D. Large-scale gene function analysis with the PANTHER classification system. 8, 1551–1566 (2013).
Bindea, G. et al. ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 25, 1091–3 (2009).
Lee, P.-C., Wildt, D. E. & Comizzoli, P. Proteomic analysis of germinal vesicles in the domestic cat model reveals candidate nuclear proteins involved in oocyte competence acquisition. MHR Basic Sci. Reprod. Med. 24, 14–26 (2018).
Acknowledgements
The authors would like to acknowledge the Smithsonian Institution for their support in this project. And Stacey Criswell from UVA for the TEM sample preparation and analysis.
Author information
Authors and Affiliations
Contributions
A.C. performed sperm motility and acrosome analysis experiments. R.D. performed IVF experiments. M.A.M.M.F., designed experiments, supervised A.C. and R.D., performed IVF experiments, data analysis and wrote this manuscript. M.J.N., processed and analyzed proteomics data. N.S. wrote manuscript and performed experiments design. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ferraz, M.d.A.M.M., Carothers, A., Dahal, R. et al. Oviductal extracellular vesicles interact with the spermatozoon’s head and mid-piece and improves its motility and fertilizing ability in the domestic cat. Sci Rep 9, 9484 (2019). https://doi.org/10.1038/s41598-019-45857-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-45857-x
This article is cited by
-
Oviductal extracellular vesicles miRNA cargo varies in response to embryos and their quality
BMC Genomics (2024)
-
Contribution of extracellular vesicles for the pathogenesis of retinal diseases: shedding light on blood-retinal barrier dysfunction
Journal of Biomedical Science (2024)
-
Plasma-derived extracellular vesicles improve mice embryo development
Molecular Biology Reports (2024)
-
A review of the regulatory mechanisms of extracellular vesicles-mediated intercellular communication
Cell Communication and Signaling (2023)
-
The sperm-interacting proteome in the bovine isthmus and ampulla during the periovulatory period
Journal of Animal Science and Biotechnology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
