
Abstract
The global dispersal of forests and soils has been proposed as a cause for the Late Devonian mass extinctions of marine organisms, but detailed spatiotemporal records of forests and soils at that time remain lacking. We present data from microscopic and geochemical analyses of the Upper Devonian Chattanooga Shale (Famennian Stage). Plant residues (microfossils, vitrinite and inertinite) and biomarkers derived from terrestrial plants and wildfire occur throughout the stratigraphic section, suggesting widespread forest in the southern Appalachian Basin, a region with no macro plant fossil record during the Famennian. Inorganic geochemical results, as shown by increasing values of SiO2/Al2O3, Ti/Al, Zr/Al, and the Chemical Index of Alteration (CIA) upon time sequence, suggest enhanced continental weathering that may be attributed to the invasion of barren lands by rooted land plants. Our geochemical data collectively provide the oldest evidence of the influences of land plants from the southernmost Appalachian Basin. Our synthesis of vascular plant fossil record shows a more rapid process of afforestation and pedogenesis across south-central Euramerica during the Frasnian and Famennian than previously documented. Together, these results lead us to propose a new hypothesis that global floral dispersal had progressed southward along the Acadian landmass rapidly during the Late Devonian.
Similar content being viewed by others

Introduction
The Late Devonian is known for the rapid and global radiation of early forests and soils such as spodosols and alfisols1,2,3. The development of land plants and soils is hypothesized to have been either a trigger or consequence of a series of global changes in the lithosphere (e.g., increased weathering and erosion), hydrosphere (e.g., anoxic oceans, global transgression and regression), atmosphere (e.g., global changes in O2 and CO2), and biosphere (i.e. mass extinctions of marine life) during the Middle to Late Devonian4,5,6,7,8,9,10. One compelling hypothesis is that forest radiation mobilized a tremendous amount of soils and associated nutrients (N and P) to coastal oceans for the first time in Earth’s history and led to dysoxic/anoxic oceans globally4,11,12,13. Testing this hypothesis, however, requires data on stratigraphic occurrences of soils and plants in specific paleogeographic areas.
Current knowledge on the paleogeographic distribution of Devonian forests is largely based on macrofossils, such as tree trunks, stems, leaves, and roots, as well as some microfossils such as spores14,15,16,17. The oldest tree stems and stumps in the Euramerica are reported from the uppermost Givetian (Middle Devonian) strata of New York (i.e., Gilboa Park and Cairo) representing the central Euramerican landmass18 (Fig. 1). To date, the record of Devonian trees and shrubs assigned to pteridophytes, which were likely the primary component for the first forest, is very limited from the southern Appalachian Basin along the southern Acadian landmass. Only a few uncertain remains have been reported as small wood fragments of possible Callixylon19 and Foerstia20 from Tennessee. In contrast, some tree or shrub fossils are known from the northern Appalachian Basin (e.g., New York, Pennsylvanian, West Virginia) and the Baltica and Avalonia landmasses (e.g., United Kingdom, Belgium)21,22,23,24,25,26,27. This gap in the paleogeographic occurrence between the northern and southern parts of the Appalachian Basin implies that forests originated from the central Euramerica in the late Middle Devonian and dispersed southward during the Late Devonian. Better understanding of the spatiotemporal occurrence of land plants in the southern Appalachian Basin will provide better understanding of the dispersal pattern of early land forest.

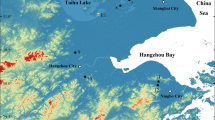
Late Devonian paleogeography. Left: the global scale; right: south-central Euramerica. The yellow star indicates the study site (the present location of Alabama), which represents the southern part of the Chattanooga Sea (dashed elliptical area). Maps are adapted from original map (360 Ma) from Global Paleogeography and Tectonics in Deep Time Series by Ron Blakey [© 2016 Colorado Plateau Geosystems Inc.].
To date, fluvial sedimentary sequences are the most frequently reported units containing tree and shrub fossils (lycopsids, cladoxylopsids, progymnosperms, and possibly stem spermatophytes) and soils in the Appalachian Basin1,18,28,29. Those sandstone, siltstone, and limestone deposits are geographically distributed along the eastern side of the basin (close to the modern-day Appalachian Mountains)30. By comparison, very few tree fossils have been reported from the extensive Upper Devonian black shale deposits further offshore from the western margin of the Appalachian Basin, including the Ohio Shale, New Albany Shale, Cleveland Shale, Huron-Dunkirk shales, Millsboro Shale, and Chattanooga Shale. Typically, the preservation of land plant fossils in offshore marine deposits is not as common as that in nearshore environments (e.g., fluvial, deltaic), because plant remains can be easily broken into pieces and decomposed through taphonomic process. However, if information on the distribution of early forests could be obtained from the abundant and regionally extensive Upper Devonian marine black shales, it would significantly increase the amount of data on the occurrence of land plants available to geologists from the Paleozoic rock record. Such efforts would allow for the reconstruction of a far more spatially and stratigraphically detailed record of afforestation than is possible using the rarely preserved fluvial deposits alone.
In the present study, we present identifiable signatures of forests and soils preserved in unfossiliferous black shales in the southernmost Appalachian Basin. We further demonstrate the potential of using these signatures to generate new understanding of the dispersal patterns of Famennian forest (land plant) and pedogenesis along the southern Acadian Orogen. We analyzed a complete section of the Chattanooga Shale in northeastern Alabama (Fig. 1). The Chattanooga Shale and other Upper Devonian black shale units in the Appalachian Basin are interpreted to have accumulated in a basin-like depositional environment further offshore than equivalent sandstone or siltstone dominated formations (i.e., alluvial plain or basin margin-like environment)30,31. Plant macrofossils such as stems, leaves, stumps, and roots are nearly absent in the Chattanooga Shale, apart from a few brief notes from central Tennessee19. Using microscopic investigation and a comprehensive set of geochemical analyses (e.g., inertinite and vitrinite, mineral composition, trace metals, stable carbon isotope ratio, and biomarker assemblages), we investigate whether early forests had left detectable signals in the Chattanooga Shale of Alabama. The multiple geochemical-tracer approach we use here can overcome the preservation limit of macrofossils, and thus extend our knowledge of the spatiotemporal pattern of the early forest radiation in the southernmost Euramerican continent. Our data can set the foundation for new hypotheses regarding how afforestation progressed during the Late Devonian.
Results
Geological background
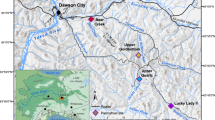
The outcrop of the Chattanooga Shale for this study is located in DeKalb County, northeastern Alabama (Fig. 1) (Material and Methods). The Chattanooga Shale is exposed with disconformable boundaries above the Upper Silurian Red Mountain Sandstone and below the Lower Mississippian Maury Shale. The Chattanooga Shale in this outcrop is 11.3 m thick and is subdivided here into a lower and upper unit based on lithological characteristics (Fig. 2). The lower unit is 4.3 m thick and characterized by thinly laminated, pyritic, fissile rocks composed of layers of interbedded gray and black shale. The upper unit is 7.0 m thick and is composed of dark gray to black, silty, blocky shales. Similar stratigraphic features have been reported from other Chattanooga Shale sections, entirely or in part, in northeastern Alabama32,33.

No macrofossils (visible to the naked eye) have been reported from the Chattanooga Shale in Alabama, aside from a single bivalve shell from a different nearby section (personal observation in 2015). Detailed field work indicates that the studied outcrop is largely unfossiliferous (i.e., no macrofossils were observed with the naked eyes), confirming previous studies. In contrast, macrofossils of marine invertebrates (e.g., brachiopods, crinoids, corals) are relatively common in the Chattanooga Shale in Tennessee20,31. A few possible plant remains have also been reported from Tennessee, but, to date, nothing that has been identified to a specific taxon has been documented from Alabama or Tennessee19.
Biostratigraphy
The phosphatic tooth-like remains of conodonts can be found in almost all marine deposits from the Lower Paleozoic to the Triassic34. For this reason, conodonts are the primary biostratigraphic fossils in most Paleozoic stages, including the Upper Devonian35,36,37,38,39. Conodonts were identified on bedding plane breaks throughout the studied outcrop to provide a biostratigraphic framework to constrain the timing of geochemical variation. The zonation names used in this section are from the updated Famennian zonation of Spalletta et al. (ref.39). For clarity and for easier comparison to previous studies (e.g., ref.35), the older zonation names are also included in parenthesis.
Most conodonts observed in the study outcrop of the Chattanooga Shale are external molds (Supplementary Fig. S6). Some phosphatic remains of conodont elements are present, but are often weathered, resulting in a white, partially dissolved element. Conodonts are most abundant in the lower unit, within the lowermost 1–2 m interval (Fig. 2). The absence of Palmateolepis glabra unca and P. lobicornis combined with the presence of P. superlobata in the 0–0.75 m interval indicates Palmateolepis termini Zone (Middle crepida Zone) in age. P1 elements of P. lobicornis, P. minuta, P. superlobata, and P. glabra unca were identified from the 1.25 m and 1.75 m levels, suggesting that this interval is Palmatolepis glabra prima Zone (Upper crepida Zone) in age35,39.
Conodonts were found through the study section. Although those remains tend to be scattered or poorly preserved in the upper unit (except well-preserved Palmateolepis perlobata at the 7.5 m-level), relatively abundant materials were recovered from the lower unit (esp., in the 0 m to 5 m interval from the base) (Fig. 2). Tasmanitid algal cysts are abundant from 5.5 to 7.5 m, and much more silty bedding planes were observed from 7.5 m to the top. Some skeletal fragments were observed under a Scanning Electron Microscope (SEM) that may be derived from brachiopods, bivalves, gastropods, and/or probable spicules of sponges. The abundant Famennian conodonts observed within the basal 2 m of the outcrop show that the studied section comprises only the Famennian Gassaway Member, and that the older Frasnian age Dowelltown Member is missing. Furthermore, the two lithological units identified in the study section resemble the lower and middle units of the Gassaway Member of the Chattanooga Shale in central Tennessee described previously19,40.
Plant residue
Plant residue were identified under microscopy. Those include fragments of leaves, branches, roots, and spores. Tubular or irregularly shaped, carbon-rich fragments are more common in the lower unit (Figs 3, Supplementary Fig. S7). Those tubular fragments display an overall curved shape with a smooth cortex. Spore-like particles and woody fragments were only observed in the upper unit. One well-preserved piece is elongated with a dimension of 30 μm in length and 10 μm in width and likely represents the remains of a spore. Spore-like particles have a rounded or elliptical shape with a bulged surface and are likely derived from trilete spore. Woody fragments are typically stick-like in shape, have a smooth surface, and have dimensions of 20–70 μm in length and about 10 μm in width. Some fragments with xylem- or phloem-like structures indicate functionally conducting wood tissues.

Scanning electron microscope (SEM) images of land plant remains from the Upper Devonian Chattanooga Shale in northeastern Alabama. (a) A tubular particle from the lower unit; (b) a deformed trilete spore from the upper unit; (c) a piece of wood fragment; and (d) a wood fragment with conducting tissues inside. The associated energy-dispersive X-ray spectroscopy (EDS) spectra are presented in Supplemental Material S7.
Organic petrographic analyses show that more than 50% of macerals are amorphous organic matter (Supplementary Fig. S8) that is generally considered to be a degradation product of organic materials of mainly marine origin41,42. Figured components are mainly vitrinite and inertinite, which are commonly used as an indicator of plant residue in both fluvial and marine strata (e.g., refs43,44,45,46,47,48). In our samples, the majority of inertinite fragments are of high reflectance and sharply angular in shape (Fig. 4) that are suggested as typical features resulting from plant combustion41,49. The relative abundance of vitrinite + inertinite is hence used to represent terrestrial inputs. Our data show a significantly increasing trend from the lower unit (mean ± standard deviation = 15.6 ± 5.4%) to the upper unit (24.5 ± 6.4%) (Mann-Whitney U test: P = 0.026).

Variations in inorganic and palynological proxies across the Upper Devonian Chattanooga Shale in northeastern Alabama. Light grey color marks the upper unit of the section. Box plots show comparison of upper vs. lower unit, and asterisks indicate that significance is detected between the lower and upper units (α = 0.05). The higher values of SiO2/Al2O3 and vitrinite + inertinite in the upper unit reflect enhanced terrestrial plant inputs accompanied by high siliciclastic input. Correspondingly, the weathering indices (CIA and CPA) values are higher in the upper unit, reflecting enhanced weathering of land materials. The higher Ti/Aland Zr/Al values in the upper unit suggest increased contributions of heavy detrital sediment inputs, and the higher TP contents in the upper unit likely indicate a higher marine primary productivity.
Bulk and molecular characteristics of organic matter
Total organic carbon (TOC) content of the rocks in the section range from 2.8% to 13.7% (Fig. 5). The TOC of black shales averages 9.6 ± 3.3% in the lower unit, which is significantly higher than that in the upper unit (6.6 ± 1.3%) (Mann-Whitney U test: P = 0.017). The δ13C values of TOC fluctuate between −29.9 and −27.9‰ with a significantly high value in the upper unit (−28.7 ± 0.4‰) than in the lower unit (−29.5 ± 0.5‰) (Mann-Whitney U test: P = 0.01).

Variations in bulk and molecular organic proxies across the Upper Devonian Chattanooga Shale in northeastern Alabama. Light grey color marks the upper unit. Box plots show comparison of upper vs. lower unit, and asterisks indicate that significance is detected between the lower and upper units (α = 0.05). TOC contents are lower in the upper unit, reflecting the dilution by detrital, inorganic materials. Higher δ13C values in the upper unit may reflect increased phytoplankton growth. Normal alkane parameters all show a significantly higher contribution of higher land plant-derived organic matter in the upper unit, and PAHs from higher plants or plant combustion are present in all samples, indicating the contributions of organic matter from higher plants throughout the Famennian.
The distribution of normal alkanes shows a carbon range from n-C13 to n-C32 with an enrichment in low molecular weight n-alkanes (n-C15 − n-C19: 45 ± 9.4% of total n-alkanes), which is typical for marine black shales50,51,52. The δ13C values of n-alkanes of selected samples demonstrate that short chain n-alkanes are on the average of 0.71‰ more enriched in 13C than long chain n-alkanes (Fig. 6). This suggests different biological origins of the short and long chain n-alkanes. The terrigenous-to-aquatic ratio (TAR), defined as (n-C27 + n-C29 + n-C31)/(n-C15 + n-C17 + n-C19), has been widely used to quantify terrestrial versus aquatic source contributions in sedimentary organic matter (e.g., refs53,54,55,56). The TAR values range between 0.07 and 0.75 and average 0.27 ± 0.20 (Fig. 5a). The upper unit has TAR values (0.43 ± 0.23) that are significantly higher than the lower unit (0.16 ± 0.07) (Mann-Whitney U test: P = 0.005).

δ13C values of individual normal alkanes of two representative samples from the Upper Devonian Chattanooga Shale in northeastern Alabama. The left and right panels show samples from the upper and lower units, respectively. The δ13C values of short chain alkanes are more positive than those of long chain alkanes, supporting our interpretation that the two homologues represent different source organisms. Error bars represent standard deviation of replicate measurements.
A range of polycyclic aromatic hydrocarbons (PAHs) were identified, and the terrestrial plant-derived compounds are presented in Figs 5b, Supplementary Fig. S2. Retene, perylene, 1,7-dimethylphenanthrene, 1,2,5,6-tetramethylnaphthalene, and 1-methylphenanthren are compounds that potentially indicate the contributions of organic matter from land plants to aquatic sediments13,57,58,59,60. Although these compounds were detected in all samples, their concentrations do not show a statistically significant difference between the lower and upper unit. The relative concentrations of perylene and the three alkyl PAHs are overall higher in the upper unit, which are 1.1 ± 0.3% and 4.6 ± 0.7% relative to total PAHs, respectively, than those in the lower unit, which are 0.3 ± 0.4 and 4.5 ± 0.4%, respectively (Fig. 5b). The relative concentrations of retene are overall stable in both the upper (0.9 ± 0.3%) and lower (0.9 ± 0.2%) units. PAHs that have been previously used to reconstruct land plant burning and forest fires in geological history were detected in all samples. These include pyrene, benzo(a)pyrene, benzo(e)pyrene and coronene (Supplementary Fig. S2)13,61. The relative concentrations of these combustion-derived PAHs show no significant change from the lower to the upper unit (Mann-Whitney U test: P = 1.000).
Inorganic geochemical characteristics of the chattanooga shale
Based on X-ray fluorescence (XRF) analyses, the samples contain SiO2 (64.8 ± 12.8 wt.%), Al2O3 (11.3 ± 1.2 wt.%), K2O (2.9 ± 0.9 wt.%), Na2O (0.6 ± 0.2 wt.%) and CaO (0.5 ± 0.5 wt.%). The Chemical Index of Alteration (CIA = [Al2O3/(Al2O3 + Na2O + CaO + K2O)] × 100) and the Chemical Proxy of Alteration (CPA = [Al2O3/(Al2O3 + Na2O)] × 100) were calculated as previously described62,63. These two proxies provide a quantitative measure for silicate rock and soil weathering, since alumina becomes increasingly enriched throughout the weathering process and sodium, calcium and potassium are more preferentially removed62,63. CIA values differ significantly between the two stratigraphic units (Mann-Whitney U test: P = 0.008), showing a lower mean value (67.5 ± 5.0) in the lower unit than the upper unit (73.1 ± 4.3) (Fig. 4). Similarly, CPA values show a significantly lower value (91.6 ± 1.1) in the lower unit than in the upper unit (93.3 ± 1.7, Mann-Whitney U test P = 0.006) (Fig. 4).
X-ray diffraction (XRD) analyses show that quartz and clay minerals are the main components in the Chattanooga Shale section. Quartz is the most abundant mineral; it varies in a large range, however, from 6.6% to 90.5% (57.9 ± 20.2%). Clay minerals are the next abundant, ranging from 12.0% to 92.2% (36.3 ± 21.6%), and chlorite and illite are the two most abundant minerals. The quartz-to-clay ratio increases significantly from 1.7 ± 1.0 in the lower unit to 3.6 ± 2.4 in the upper unit (Mann-Whitney U test: P = 0.189). This pattern is in agreement with the XRF data that show an increasing trend in the ratios of SiO2 to Al2O3, from 5.1 ± 1.1 in the lower unit to 7.2 ± 1.3 in the upper unit (Mann-Whitney U test: P = 0.001) (Fig. 4). Crystal forms examined under SEM show that the quartz component is primarily made up of detrital grains (Supplementary Fig. S9), instead of originating from silica-filled cysts of green algae Tasmanites, which instead appear as flattened organic spheres lacking early quartz cement. Therefore, the variation in SiO2 primarily reflects changes in the relative amount of terrigenous materials. On the other hand, Al-normalized concentrations of Ti and Zr, Ti/Al and Zr/Al ratios in the lower unit are (0.07 ± 0.01 and (2.19 ± 0.27) × 10−3 respectively) significantly lower relative to the values for the upper unit (0.09 ± 0.01 and (3.28 ± 0.11) × 10−3, respectively; Mann-Whitney U test: P ≤ 0.003) (Fig. 4). Ti and Zr are thought to be contributed by high-density minerals such as zircon, rutile, sphene, and ilmenite64. This increasing trend in the studied section suggests a change in mineral assemblages during the deposition of the Chattanooga Shale.
Discussion
Although black shales in eastern North America may not preserve abundant macrofossils of Devonian trees and shrubs, our geochemical and microscopic data show that the signatures of afforestation can be identified in offshore environments. Here, we present three main lines of geochemical evidence that demonstrate land plants contributed organic and inorganic terrestrial material to offshore environments observed throughout the deposition of the Chattanooga Shale. First, plant body parts and combustion residue (e.g., wood pieces, spores, vitrinite, inertinite) throughout the entire Alabama section, provide the most direct, visible evidence of the land plant contribution to the Chattanooga Shale (Figs 3, Supplementary Figs S7, S8). Vitrinite is thought to be derived from wood tissues41,65,66, and inertinite represents highly oxidized materials generated from slow oxidation of organic matter or rapid oxidation during wildfires49,67,68,69. In the studied section, vitrinite and inertinite show an overall increasing trend from the lower to upper unit (Fig. 4). Similarly, Rimmer et al. (ref.48) also reported this pattern of increases in inertinite from the uppermost Devonian terrestrial and marine rocks (including black shales) in the northern Appalachian Basin, and it was interpreted to be a result of an increasing occurrence of wildfires.
The second line of geochemical evidence is based on biomarkers including PAHs and normal alkanes. PAH compounds including pyrene, benzo(a)pyrene, benzo(e)pyrene and coronene (Fig. 5b) possibly indicate land plant burning. These PAH compounds have been used to indicate wildfire events throughout the Phanerozoic from a diverse type of rocks and sediments, including Devonian marine sedimentary rocks13,46,52,70. For example, benzo(a)pyrene, benzo(e)pyrene, pyrene and coronene co-occurring with inertinite were reported from Upper Devonian marine rocks in Poland as the evidence of paleo-wildfires in the eastern Avalonia46. In addition to combustion-related PAHs, a range of compounds that may indicate the occurrence of terrestrial plant material are present in our samples, including retene, perylene, long chain n-alkanes, 1,7-dimethylphenanthrene, 1,2,5,6-tertramethylnaphathalene, and 1-methylphenanthrene (Fig. 5). Retene is structurally similar to abietane that is derived from the conifer biomarker abietic acid. Although the oldest macrofossil record of conifers was reported from the Late Carboniferous71, the earliest tracheophytes may also produce the conifer biomarkers60. The occurrence of retene in ancient rocks has been considered to be strong evidence for the contributions of early terrestrial higher plant58,72,73,74. Perylene is believed to originate from the activity of wood-degrading fungi59,75. It is frequently found in sediments and crude oils dating back to the Mesozoic but appears to be largely absent in marine sediments lacking terrestrial input and samples deposited before the rise of vascular plants59. Previous studies have used perylene in Devonian marine formations to reflect organic matter contributions from terrestrial higher plants (e.g., refs13,60,76). In our samples, retene and perylene were detected throughout the studied section, supporting the presence of organic matter from terrestrial higher plants to the Chattanooga Shale of Alabama (Fig. 5b).
Other biomarkers that are less source-specific but may also indicate terrestrial plants, were also evaluated. Traditionally, short chain (C15–C19) n-alkanes in aquatic sediments have been used to represent contributions from algae and microorganisms77,78,79, whereas long chain n-alkanes (≥n-C27) are thought to originate primarily from terrestrial vascular plants80,81. Compound-specific stable carbon isotopes of n-alkanes can further differentiate the biological sources of short versus long chain n-alkanes50,82,83,84. Based on this assumption, TAR was applied to represent organic matter contributions of land plants relative to marine microorganisms, and it shows an increasing trend from the lower to the upper unit in the Chattanooga Shale (Fig. 5a). The assumption that long chain and short chain n-alkanes originate from different biological origins is supported by the observation that the δ13C values of long chain n-alkanes are more depleted than the short chain counterparts (Fig. 6). It also needs to be noted that although long chain n-alkanes are among the most widely utilized biomarkers for terrestrial higher land plants80,85,86, mosses and the non-marine microalgae, Botryococcus braunii, also produce these compounds87,88. Despite this source ambiguity, the TAR ratios in our samples show a strong covariation with the abundance of inertinite and vitrinite (Pearson’s r = 0.702, P = 0.004), supporting the idea that TAR can be used as a proxy of variation in land plant input to the Chattanooga Shale. Additionally, several PAH compounds, 1,7-dimethylphenanthrene, 1,2,5,6-tertramethylnaphathalene, and 1-methylphenanthrene, are present in our samples (Figs 5, Supplementary Fig. S2). These compounds are generally not considered as source-specific terrestrial plant biomarkers as they may be derived from aromatization of organic matter of various biological origins. Terpenoids structures that are prevalent among land plants is one likely source89, and these compounds have been previously used to indicate land plant input into marine sediments (e.g., refs58,60,90).
The third line of evidence lies in inorganic geochemical proxies, which show that continental weathering became more intense during the deposition of the Chattanooga Shale. SiO2/Al2O3, CIA, and CPA all show an increasing trend from the lower to upper unit (Fig. 4). SiO2/Al2O3 is a useful indicator for changes of detrital input into marine environments91,92. CIA and CPA calculated from marine sediments have been widely used to evaluate the chemical weathering intensity of source areas and rocks (e.g., refs93,94,95,96). Both CIA and CPA are positively correlated with SiO2/Al2O3 (CIA vs. SiO2/Al2O3: Pearson’s r = 0.662, P = 0.001; CPA vs. SiO2/Al2O3: Pearson’s r = 0.621, P = 004), and their increasing trends indicate an increase in terrigenous quartz input accompanied by intensification of weathering on land during the deposition of the Chattanooga Shale. Correspondingly, TOC concentrations in the upper unit are lower, probably reflecting an increasing dilution of in situ produced marine organic material caused by increasing amount of continental clastic material. It needs to be acknowledged that marine productivity declines can also lead to the TOC concentration reduction, but this interpretation does not agree with the higher δ13C values and total phosphorous in the upper unit (Figs 4 and 5). Additionally, the upward increases in Ti/Al and Zr/Al suggest that heavier, coarse minerals were deposited over time. This reflects a stronger force of mobilizing allocthonous minerals97,98 and further confirms the increased contribution of terrigenous sediments. The Ti/Al and Zr/Al ratios have also been used in other Upper Devonian marine sedimentary sequences to indicate the relative contribution of heavy minerals and the strength of material transportation from land to sea52,92,99,100,101.
The enhancement in continental weathering during the deposition of the Chattanooga Shale may be caused by a climatic shift to warmer and wetter conditions, yet this explanation contradict previous data suggesting that the early and middle Famennian climate become cooler and drier globally and near the study area25,102. More likely, the intensified continental weathering is due to land plant invasion onto unvegetated, barren lands. The early development and invasion of rooted land plants to barren lands have been suggested to accelerate physical and chemical weathering of bedrocks through the Devonian4,103. The roots of land plants evolved from being small (1–3 mm in diameter and up to 30 cm long), having limited geochemical effects on soils during the Early Devonian104,105,106,107, to being large (>2.5 cm in diameter), deep (reaching >1 m in depth), and effective in breaking down rocks during the Late Devonian4,103,104,108. Our data show significant positive correlations between the proxies of terrestrial plant abundance (TAR, vitrinite and inertinite) and the proxies of continental input and weathering (SiO2/Al2O3, CIA and CPA) (Fig. 7), providing further evidence supporting the interrelated connections among land plants, continental weathering and soil development. The combustion and land plant related PAH compounds show a more scattered pattern (Fig. 5) and do not correlate significantly with the weathering proxies, but their low concentrations make reliable quantifications difficult. Nevertheless, their occurrences throughout the studied section strongly support that land plants were widespread in the southern Acadian land during the Late Devonian.

Cross plots of terrestrial plant proxies versus continental weathering proxies from the Upper Devonian Chattanooga Shale in northeastern Alabama. Pearson’s P and r values are presented. Red lines denote linear regression lines. The positive significant correlations between the indicators for plant inputs and weathering intensity suggest that early land plants likely intensified continental weathering.
Combining geochemical analyses, including multiple organic and inorganic geochemical proxies, our data provide the first evidence of afforestation on the southern Acadian land and the associated changes in land-ocean biogeochemical linkages during the Famennian. The occurrences of microfossils (wood fragments and spores) and biomarkers indicate that forests were present during the Famennian time on the southern Acadian land – a paleogeographic region and time that is largely absent of plant records based on conventional investigations of fossils. Several proxies (vitrinite and inertinite, TAR, retene, perylene, chemical weathering indices) further demonstrate that terrestrial plants became an increasing source of organic matter that likely intensified continental weathering and better mobilized clastic materials during the deposition of the Gassaway Member of the Chattanooga Shale.
Current knowledge of the paleogeographic distribution of Devonian forests and associated soils is primarily based on the fossil record paleobiogeographic occurences of vascualr siltstone successions (e.g., refs15,16,28,109,110,111) (Supplementary Table S1). Our data from black shales, therefore, make an important addition to the scarce records of paleogeographic occurrences of the early forest and soil formations during the Late Devonian by presenting clear evidence of afforestation and the associated input to marine sediments in a paleogeographic area with no previously known records (i.e., the southern Acadian Orogen). Because upper Devonian black shale units are geographically distributed in a large area from the northernmost to southernmost margins of the Appalachian Basin30, they overcome the limitation due to the poor preservation of terrestrial deposits and can place the record of afforestation within a detailed biostratigraphic framework. Although the geochemical and microscopic data do not provide diagnostic characteristics to identify specific plant taxa, accumulated information on biostratigraphic and paleobiogeographic occurences of vascular plant fossils (e.g., refs28,109,112) can offer a reasonable clue. In southcentral Euramerica along the Acadian landmass, plants with wood tissues, assigned to first shrubs or trees, appeared and soon diversified during the latest Middle Devonian to the end of the Famennian (Fig. 8 and Supplementary Table S1). Because woody tissue (taller and robust stem, megallophyles, and deeper roots) is thought to be advantageous for adapting to or invading drier and more inland environments, those species are thought to be a major contributor of the earliest forests in this paleogeographic region29,109.

Spatiotemporal dispersal pattern of land forests in southcentral Euramerican landmass during the Devonian. Vascular plant occurrence is summarized by taxa (specific-level), time (Emsian to Famennian ages), and space (state, province, and/or country). Circle size indicates stratigraphic distribution: the older record with a larger symbol (see the legend on left bottom). Three gradients in states/region/country (dark grey, intermediate, and light grey) represent dispersal time-sequence: the older with the dark to the younger with the light color), showing a southern dispersal pattern (the ‘south path’ arrow). A simplified cladogram of higher-level euphyllophytes (selected tree and shrub taxa) show a diversity pattern through the time (i.e., Famennian expansion). Raw data are available in Supplementary Table S2.
To place our geochemical data in the context of the spatiotemporal evolution of Devonian forest, we synthesize the fossil record of vascular plants (Eutracheophytes) based on the published data, museum specimens, and the Paleobiology Database (https://paleobiodb.org/). Figure 8 shows a summary of the generic- and species-level occurrence across time (three Age- or Stage-based intervals) and space (state-province-country), which characterizes the Devonian afforestation and pedogenesis (raw data of the figure available in Supplementary Table S1). Stratigraphically, the oldest trees and shrubs (within euphyllophytes) appeared in the Givetian (later Middle Devonian) strata; only a few basal eutracheophtyes, such as the trimerophytes, are known from Emisian-Eifelian strata of the northern Acadian-southern Caledonian orogens (indicated by states/region/country in dark grey on the map of Fig. 8). Although cladoxylopsids were the most dominant group in the Middle Devonian, aneurophytales eventually took over the niches through the Givetian–Frasnian transition. Paleogeographically, the two main tree-shrub bearing groups (taxa assigned to Cladoxylopsida and the progymnosperm Aneurophytopsida) were restricted in the northern Acadian region by the end of the Frasnian (see the stratigraphic occurrence with a cladogram in Fig. 8 and Supplementary Table S2). However, through the Famennian Age, the single taxon of Archaeopteridopsida, Archaeopteris–Callixylon, had spread from 2 to 12 states/provinces. This pattern is unlikely to result from differential preservation in space and time because of two reasons. First, plant fossils have been observed in formations composed of various rock types (e.g., sandstone, siltstone, shale, see Supplementary Table S1), suggesting that vascual plant fossil preservation in the Appalachiain Basin is not selective for rock types. Second, we observe temporal changes in plant fossils for a given formation with little change in lithology. For example, Frasnian shales (e.g., those lower members of the Chattanooga Shale, the Fynn Creek Formation) in the Southern Appalachian have no known wood fossils, but Famennian shales of the Chattanooga Shale contain some well-preserved large tree fossils in central Tennessee (i.e., in the Gassaway Member) (Supplementary Fig. S10). Rather than reflecting preservation differences, these stratigraphic and paleogeographic data reveal two important trends of floral turnover that occurred on a large scale through the Frasnian–Famennian transition near the southcentral Euramerican landmass. First, the global floral turnover had progressed southward (see the red arrow of ‘south path’ on Fig. 8). Second, this global southward dispersal had progressed in a relatively short time during the Frasnian–Famennian transition. Our geochemical data provide the first strong evidence for the southern end of this southward path. We further hypothesize that this transition could have extended further into the southern American landmass of Gondwana as part of global-scale afforestation, progressing from north to south during the Famennian (e.g., refs18,113) especially if the physical landbridge between Euramerica and Gondwana existed by the end of the Devonian114. The widespread, yet under-utilized, unfossiliferous Devonian black shales may hold the key to test this hypothesis by filling temporal and spatial gaps in the global path of afforestation and pedogenesis.
Material and Methods
Samples
Rock samples were collected from 45 layers at a 25-cm interval, which covered all identified or visible lithological changes in the Chattanooga Shale section (Fig. 2). Weathered rocks (i.e., generally turning to light gray) were avoided for sampling, but freshly exposed rocks (i.e., darker color) were chosen. Prior to geochemical analyses, samples were washed in sonicating baths sequentially using ultra-pure carbon-free water, dichloromethane, and hexane.
X-ray diffraction (XRD), X-ray fluorescence (XRF) and scanning electron microscope (SEM)
Fifteen samples collected at intervals of 25–50 cm were selected for X-ray diffraction analysis on a Bruker D8 Advance XRD. Mineral identification was based on diffraction patterns using the DIFFRACPLUSEVA 4.0 library (Bruker AXS), and the abundances of different mineral components were determined using the Rietveld Method115.
Total element contents were determined by X-ray fluorescence spectrometry (Philips PANalytical PW2424, Netherland) at the ALS Chemex Lab, Ltd (Guangzhou, China). Before the analysis, the powdered samples were dissolved using lithium metaborate mixed with lithium nitrate and heated at 1050 °C for an hour. The mixtures were then transferred into a platinum mould and analyzed by XRF spectrometry.
Selected samples were fixed on stubs and coated with carbon for further examination of minerals and microfossils using a JOEL SEM (JSM-6010PLUS/LA). The SEM magnification was set to 500X to 3500X, depending on the size of the particles. The elemental composition of the samples was analyzed on a JEOL 7000 FE SEM equipped with EDX at the Central Analytical Facility, The University of Alabama.
Trace element
Measurements of trace elements were made using a PerkinElmer Elan9000 element inductively coupled plasma mass spectrometry (ICP-MS) at the ALS Chemex Lab, Ltd (Guangzhou, China). Ground samples were prepared using a four-acid (HF, HNO3, HClO4, HCl) digestion method116. Analytical precision for all elements is better than 7%, and accuracy was evaluated relative to international reference materials, including GBM398-4c, GBM908-10, MRGeo08 and OGGeo08.
Organic petrography
Samples at an interval of 50 cm were selected for organic petrography analysis. 10 g of each sample was demineralized using cold 10% HCl for 24 h and then cold 48% HF for 48 h. The samples were then treated with hot Schultz’s solution and sodium hydroxide, followed by a water rinse until a neutral pH was achieved. The residues of the samples were embedded in epoxy resin, polished, and observed using reflectance microscopy under a Nikon Microphot microscope. The samples were examined under immersion oil using a ×40 objective lens, and the abundances of vitrinite and inertinite particles were point-counted (500 points). All samples were analyzed in duplicate and mean values of the results are reported in this study.
TOC and stable carbon isotope of TOC
Samples at intervals of 25–100 cm were analyzed for total organic carbon (TOC) and stable isotopes of total organic carbon (δ13C). Samples were ground into 100–200 mesh powder, and approximately 10 mg of powdered samples were placed into tin capsules. Samples were treated with 5% sulfurous acid to remove carbonate and then dried overnight in an oven at 50 °C. The samples were analyzed on a Micro Cube elemental analyzer (Elementar Analysensysteme GmbH, Hanau, Germany) interfaced to a PDZ Europa 20–20 isotope ratio mass spectrometer (The Sercon Ltd., Cheshire, UK) at The University of California Davis Stable Isotope Facility (California, USA). The analytical precision was <0.2‰ based on internal standards (including nylon, bovine liver, peach leaves, and glutamic acid) calibrated against NIST Standard Reference Materials (USGS-40, USGS-41). Our data were reported as δ13C values (‰) relative to V-PDB.
Biomarker quantification and compound-specific stable carbon isotope
Samples at an interval of 25–100 cm were selected for biomarker analysis at the Organic Geochemistry Laboratory, Department of Geological Sciences, University of Alabama. Duplicate samples were analyzed in every five samples, and solvent blanks were taken through the whole procedure for each run. Approximately 5 g powdered samples were ultrasonically extracted (20 minutes) three times with a mixture of 18 mL dichloromethane (DCM) and 2 mL methanol. Blanks (i.e., only the solvent mixture) were analyzed every 5 samples. In order to remove sulfur, short copper turnings were added to the extracts during the extraction process (20 °C) and overnight storage (−20 °C). The extracts were then concentrated to a volume of ca. 1 mL with a gentle ultrahigh purity (UHP) nitrogen stream using a Zymark Turbo Vap LV Evaporator, and the concentrates were then transferred into GC vials. The extracts were further blown dry gently, diluted with 300–350 μL of hexane, and run on an Agilent 7890B gas chromatograph interfaced with an Agilent 5977 A mass selective detector (MSD). The MSD was operated at a full scanning mode in the mass range of m/z 50–700 at 2.3 scans per second at ionizing electron energy of 70 eV. A fused silica capillary column (Agilent Technologies: 30 m × 0.32 mm, DB-5, 0.25 μm) was used with helium as the carrier gas at a rate of 0.9 mL/min. Sample injection was operated in a pulsed splitless mode at 320 °C. The oven temperature was set at 60 °C, held for 1 minute, and increased at a rate of 6 °C/min to 325 °C, held for 20 minutes. External standards are a mixture of C7–C40 saturated n-alkanes (Sigma Aldrich 49453-U, St. Louis, Missouri) and a PM-610 PAH (Ultra Scientific, North Kingstown, Rhode Island) for quantifying aliphatic and aromatic compounds, respectively. The concentration was calculated using a five-point, peak area vs. concentration calibration curve constructed from standard mixtures with known concentrations (concentration from 0.1 to 20 ng/μL). Compound concentrations were reported in values normalized to TOC contents (μg/g TOC) or the relative percentages. Selected samples were separated into aliphatic and aromatic fractions after the precipitation of asphaltenes. The de-asphalted extracts were then separated into saturate, aromatic, and polar fractions using hexane, benzene, and methanol respectively.
For compound-specific carbon isotopes of normal alkanes, saturated hydrocarbon fractions were further separated into n-alkanes and branched/cyclic alkanes by urea adduction117. The δ13C values of n-alkanes were measured in duplicate on a Thermo Fisher Trace GC Ultra coupled with a Thermo Fisher MAT-253 mass spectrometer. The GC was fitted with a 60 m × 0.32 mm i.d. A DB-1MS fused silica capillary column with a film thickness of 0.25 μm leading directly into the combustion furnace was used. The GC oven temperature was programmed from 50 °C (1 min) at 1.5 °C/min to 125 °C, then increased to 300 °C at 5 °C/min, and finally held at 300 °C for 30 min. Helium was used as the carrier gas. The isotopic values were calibrated against the reference gas and were reported in the usual “del” notation relative to VPDB. The precision of the measurements was typically <0.5‰. The accuracy of the instrument was evaluated two to three times daily via analyzing a mixture of n-alkanes with known δ13C values acquired from Indiana University, USA.
References
Retallack, G. J. Early forest soils and their role in Devonian global change. Science 276, 583–585 (1997).
Algeo, T. J., Berner, R. A., Maynard, J. B. & Scheckler, S. E. Late Devonian oceanic anoxic events and biotic crises: “rooted” in the evolution of vascular land plants. GSA Today 5, 45–66 (1995).
Le Hir, G. et al. The climate change caused by the land plant invasion in the Devonian. Earth Planet. Sci. Lett. 310, 203–212 (2011).
Algeo, T. J. & Scheckler, S. E. Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events. Philos. Trans. R Soc. Lond B Biol. Sci. 353, 113–130 (1998).
Berner, R. A. The rise of plants and their effect on weathering and atmospheric CO2. Science 276, 544–546 (1997).
Turgeon, S. C., Creaser, R. A. & Algeo, T. J. Re-Os depositional ages and seawater Os estimates for the Frasnian–Famennian boundary: implications for weathering rates, land plant evolution, and extinction mechanisms. Earth Planet. Sci. Lett. 261, 649–661 (2007).
Błażejowski, B., Gieszcz, P., Brett, C. E. & Binkowski, M. A moment from before 365 Ma frozen in time and space. Sci. Rep. 5, 14191, https://doi.org/10.1038/srep14191 (2015).
Brom, K. R., Salamon, M. A. & Gorzelak, P. Body-size increase in crinoids following the end-Devonian mass extinction. Sci. Rep. 8, 9606, https://doi.org/10.1038/s41598-018-27986-x (2018).
Haworth, M. et al. Impaired photosynthesis and increased leaf construction costs may induce floral stress during episodes of global warming over macroevolutionary timescales. Sci. Rep. 8, 6206, https://doi.org/10.1038/s41598-018-24459-z (2018).
Percival, L. et al. Precisely dating the Frasnian–Famennian boundary: implications for the cause of the Late Devonian mass extinction. Sci. Rep. 8, 9578, https://doi.org/10.1038/s41598-018-27847-7 (2018).
Algeo, T. J. & Scheckler, S. E. Land plant evolution and weathering rate changes in the Devonian. J. Earth Sci. 21, 75–78 (2010).
Moreno, C., González, F., Sáez, R., Melgarejo, J. C. & Suárez-Ruiz, I. The Upper Devonian Kellwasser event recorded in a regressive sequence from inner shelf to lagoonal pond, Catalan Coastal Ranges, Spain. Sedimentology 65, 2055–2087 (2018).
Tulipani, S. et al. Changes of palaeoenvironmental conditions recorded in Late Devonian reef systems from the Canning Basin, Western Australia: A biomarker and stable isotope approach. Gondwana Res. 28, 1500–1515 (2015).
Richardson, J. B. & McGregor, D. C. Silurian and Devonian spore zones of the Old Red Sandstone continent and adjacent regions. Geol. Surv. Can. No. 364, 1–79 (1986).
Scheckler, S. E. Geology, floristics and paleoecology of Late Devonian coal swamps from Appalachian Laurentia (USA). Annales de la Société géologique de Belgique, 109, 209–222 (1986).
Scheckler, S. E. Floras of the Devonian-Mississippian transition. Notes for a Short Course: Studies in Geology 15, 81–96 (1986).
Wang, D., Liu, L., Guo, Y., Xue, J. & Meng, M. A Late Devonian fertile organ with seed plant affinities from China. Sci. Rep. 5, 10736 (2015).
Stein, W. E., Mannolini, F., Hernick, L. V., Landing, E. & Berry, C. M. Giant cladoxylopsid trees resolve the enigma of the Earth’s earliest forest stumps at Gilboa. Nature 446, 904–908 (2007).
Conant, L. C. & Swanson, V. E. Chattanooga Shale and related rocks of central Tennessee and nearby areas. USGS Prof. Paper 357, 1–91 (1961).
Schopf, J. M. & Schwietering, J. F. Foerstia zone of the Ohio and Chattanooga shales. US Geol. Surv. Bull. 1294-H, 1–15 (1970).
Long, A. G. XIII. — On the Structure of Deltasperma fouldenense gen. et sp. nov., and Camptosperma berniciense gen. et sp. nov., petrified seeds from the Calciferous Sandstone Series of Berwickshire. Earth Env. Sci. Trans. R. Soc. Edinburgh 64, 281–295 (1961).
Galtier, J. & Meyer-Berthaud, B. The early seed plant Tristichia tripos (Unger) comb. nov. from the Lower Carboniferous of Saalfeld, Thuringia. Rev. Palaeobot. Palynol. 93, 299–315 (1996).
Matten, L. C., Lacey, W. S., May, B. I. & Lucas, R. C. A megafossil flora from the uppermost Devonian near Ballyheigue, Co. Kerry, Ireland. Rev. Palaeobot. Palynol. 29, 241–251 (1980).
Berry, C. M. & Fairon-Demaret, M. The middle Devonian flora revisited. In Plants Invade the Land: Evolutionary and Environmental Perspectives (eds Gensel, P. G. & Edwards, D.), 120–139 (Columbia University Press, Cambridge, 2001).
Streel, M., Caputo, M. V., Loboziak, S. & Melo, J. H. G. Late Frasnian–Famennian climates based on palynomorph analyses and the question of the Late Devonian glaciations. Earth Sci. Rev. 52, 121–173 (2000).
Scheckler, S. E., Skog, J. E. & Banks, H. P. Langoxylon asterochlaenoideum Stockmans: anatomy and relationships of a fern-like plant from the Middle Devonian of Belgium. Rev. Palaeobot. Palynol. 142, 193–217 (2006).
Prestianni, C. & Gerrienne, P. Early seed plant radiation: an ecological hypothesis. Geol. Soc. London Spec. Publ. 339, 71–80 (2010).
Cressler, W. L. Plant paleoecology of the Late Devonian Red Hill locality, north-central Pennsylvania, an Archaeopteris-dominated wetland plant community and early tetrapod site. Geol. Soc. Spec. Pap. 399, 79–102 (2006).
Meyer-Berthaud, B., Soria, A. & Decombeix, A.-L. The land plant cover in the Devonian: a reassessment of the evolution of the tree habit. Geol. Soc. London Spec. Publ. 339, 59–70 (2010).
Woodrow, D., Dennison, J., Ettensohn, F., Sevon, W. & Kirchgasser, W. Middle and Upper Devonian stratigraphy and paleogeography of the central and southern Appalachians and eastern Midcontinent, U.S.A. In Devonian of the World (eds. McMillan, N. J. et al.) Can. Assoc. Petr. Geol. Mem. 14, 277–301 (1988).
Schieber, J. Evidence for high-energy events and shallow-water deposition in the Chattanooga Shale, Devonian, central Tennessee, USA. Sediment. Geol. 93, 193–208 (1994).
Hass, W. H. Age and correlation of the Chattanooga Shale and the Maury Formation. USGS Prof. Paper 286, 2330–7102 (1956).
Pashin, J. C., Kopaska-Merkel, D., Arnold, A. & McIntyre, M. Geological foundation for production of natural gas from diverse shale formations. RPSEA, Final Report 7122, 156 (2011).
Sweet, W. A quantitative conodont biostratigraphy for the Lower Triassic. Senckenbergiana lethaea 69, 253–273 (1988).
Over, D. J. Conodont biostratigraphy of the Chattanooga Shale, Middle and Upper Devonian, southern Appalachian Basin, eastern United States. J. Paleontol. 81, 1194–1217 (2007).
Hansma, J. et al. Late Devonian carbonate magnetostratigraphy from the Oscar and Horse Spring Ranges, Lennard Shelf, Canning Basin, Western Australia. Earth Planet. Sci. Lett. 409, 232–242 (2015).
Hillbun, K. et al. Upper Kellwasser carbon isotope excursion pre-dates the F–F boundary in the Upper Devonian Lennard Shelf carbonate system, Canning Basin, Western Australia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 438, 180–190 (2015).
Day, J. & Brian, J. W. Upper Devonian biostratigraphy, event stratigraphy, and Late Frasnian Kellwasser extinction bioevents in the Iowa Basin: western Euramerica. Stratigraphy & Timescales 2, 243–332 (2017).
Spalletta, C., Perri, M. C., Over, D. J. & Corradini, C. Famennian (Upper Devonian) conodont zonation: revised global standard. Bull. Geosci. 92, 31–57 (2017).
Schieber, J. Developing a sequence stratigraphic framework for the Late Devonian Chattanooga Shale of the southeastern USA: relevance for the Bakken Shale. Williston Basin Symposium, Saskatchewan Geol. Soc. Spec. Publ. 13, 58–68 (1998).
Taylor, G. H. et al. Organic Petrology (Gevruder Borntraeger, Berlin, 1998).
Pickel, W. et al. Classification of liptinite–ICCP System 1994. Int. J. Coal Geol. 169, 40–61 (2017).
Nichols, G. & Jones, T. Fusain in Carboniferous shallow marine sediments, Donegal, Ireland: the sedimentological effects of wildfire. Sedimentology 39, 487–502 (1992).
Falcon-Lang, H. J. The impact of wildfire on an Early Carboniferous coastal environment, North Mayo, Ireland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 139, 121–138 (1998).
Jahren, A. H., Arens, N. C., Sarmiento, G., Guerrero, J. & Amundson, R. Terrestrial record of methane hydrate dissociation in the Early Cretaceous. Geology 29, 159–162 (2001).
Marynowski, L. & Filipiak, P. Water column euxinia and wildfire evidence during deposition of the Upper Famennian Hangenberg event horizon from the Holy Cross Mountains (central Poland). Geol. Mag. 144, 569–595 (2007).
Petersen, H. I., Schovsbo, N. H. & Nielsen, A. T. Reflectance measurements of zooclasts and solid bitumen in Lower Paleozoic shales, southern Scandinavia: correlation to vitrinite reflectance. Int. J. Coal Geol. 114, 1–18 (2013).
Rimmer, S. M., Hawkins, S. J., Scott, A. C. & Cressler, W. L. The rise of fire: fossil charcoal in late Devonian marine shales as an indicator of expanding terrestrial ecosystems, fire, and atmospheric change. Am. J. Sci. 315, 713–733 (2015).
Scott, A. C. & Glasspool, I. J. Observations and experiments on the origin and formation of inertinite group macerals. Int. J. Coal Geol. 70, 53–66 (2007).
Sikes, E. L., Uhle, M. E., Nodder, S. D. & Howard, M. E. Sources of organic matter in a coastal marine environment: evidence from n-alkanes and their δ13C distributions in the Hauraki Gulf, New Zealand. Mar. Chem. 113, 149–163 (2009).
Brown, T. C. & Kenig, F. Water column structure during deposition of Middle Devonian–Lower Mississippian black and green/gray shales of the Illinois and Michigan Basins: a biomarker approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 215, 59–85 (2004).
Riboulleau, A. et al. Organic matter deposition in the Ghadames Basin (Libya) during the Late Devonian—A multidisciplinary approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 497, 37–51 (2018).
Bourbonniere, R. A. & Meyers, P. A. Sedimentary geolipid records of historical changes in the watersheds and productivities of Lakes Ontario and Erie. Limnol. Oceanogr. 41, 352–359 (1996).
Fabbri, D., Sangiorgi, F. & Vassura, I. Pyrolysis–GC–MS to trace terrigenous organic matter in marine sediments: a comparison between pyrolytic and lipid markers in the Adriatic Sea. Analytica Chimica Acta 530, 253–261 (2005).
Lu, Y. & Meyers, P. A. Sediment lipid biomarkers as recorders of the contamination and cultural eutrophication of Lake Erie, 1909–2003. Org. Geochem. 40, 912–921 (2009).
Silva, T. R., Lopes, S. R., Spörl, G., Knoppers, B. A. & Azevedo, D. A. Source characterization using molecular distribution and stable carbon isotopic composition of n-alkanes in sediment cores from the tropical Mundaú–Manguaba estuarine–lagoon system, Brazil. Org. Geochem. 53, 25–33 (2012).
Laflamme, R. E. & Hites, R. A. The global distribution of polycyclic aromatic hydrocarbons in recent sediments. Geochim. Cosmochim. Acta 42, 289–303 (1978).
Armstroff, A., Wilkes, H., Schwarzbauer, J., Littke, R. & Horsfield, B. Aromatic hydrocarbon biomarkers in terrestrial organic matter of Devonian to Permian age. Palaeogeogr. Palaeoclimatol. Palaeoecol. 240, 253–274 (2006).
Grice, K. et al. New insights into the origin of perylene in geological samples. Geochim. Cosmochim. Acta 73, 6531–6543 (2009).
Romero-Sarmiento, M.-F., Riboulleau, A., Vecoli, M. & Versteegh, G. J.-M. Aliphatic and aromatic biomarkers from Gondwanan sediments of Late Ordovician to Early Devonian age: An early terrestrialization approach. Org. Geochem. 42, 605–617 (2011).
Cesar, J. & Grice, K. δ13C of polycyclic aromatic hydrocarbons to establish the facies variations in a fluvial deltaic Triassic record (Dampier sub-Basin, Western Australia). Org. Geochem. 107, 59–68 (2017).
Nesbitt, H. W. & Young, G. Early Proterozoic climates and plate motions inferred from major element chemistry of lutites. Nature 299, 715 (1982).
Buggle, B., Glaser, B., Hambach, U., Gerasimenko, N. & Marković, S. An evaluation of geochemical weathering indices in loess–paleosol studies. Quat. Int. 240, 12–21 (2011).
Calvert, S. & Pedersen, T. Elemental proxies for palaeoclimatic and palaeoceanographic variability in marine sediments: Interpretation and application, in Proxies in Late Cenozoic Paleoceanopgraphy (eds Hillaire-Marcel C. & Vernal A. D.) 568–644 (Elsevier, 2007).
Kennedy, K. L. et al. Lower Devonian coaly shales of northern New Brunswick, Canada: plant accumulations in the early stages of Terrestrial colonization. J. Sediment Res. 83, 1202–1215 (2013).
Killops, S. D. & Killops, V. J. Introduction to Organic Geochemistry. (John Wiley & Sons, Oxford, 2013).
Jones, T. P. & Chaloner, W. G. Fossil charcoal, its recognition and palaeoatmospheric significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 97, 39–50 (1991).
Scott, L. Microscopic charcoal in sediments: Quaternary fire history of the grassland and savanna regions in South Africa. J. Quat. Sci 17, 77–86 (2002).
Uhl, D. & Kerp, H. Wildfires in the Late Palaeozoic of Central Europe–The Zechstein (Upper Permian) of NW-Hesse (Germany). Palaeogeogr. Palaeoclimatol. Palaeoecol. 199, 1–15 (2003).
Kaiho, K. et al. A forest fire and soil erosion event during the Late Devonian mass extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 392, 272–280 (2013).
Scott, A. The earliest conifer. Nature 251, 707 (1974).
Menor-Salván, C., Tornos, F., Fernández-Remolar, D. & Amils, R. Association between catastrophic paleovegetation changes during Devonian–Carboniferous boundary and the formation of giant massive sulfide deposits. Earth Planet. Sci. Lett. 299, 398–408 (2010).
Romero-Sarmiento, M.-F., Riboulleau, A., Vecoli, M., Laggoun-Défarge, F. & Versteegh, G. J. M. Aliphatic and aromatic biomarkers from Carboniferous coal deposits at Dunbar (East Lothian, Scotland): Palaeobotanical and palaeoenvironmental significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 309, 309–326 (2010).
Romero-Sarmiento, M.-F., Riboulleau, A., Vecoli, M. & Versteegh, G. J. Occurrence of retene in upper Silurian–lower Devonian sediments from North Africa: Origin and implications. Org. Geochem. 41, 302–306 (2010).
Jiang, C., Alexander, R., Kagi, R. I. & Murray, A. P. Origin of perylene in ancient sediments and its geological significance. Org. Geochem. 31, 1545–1559 (2000).
Song, D., He, D., Qi, X., Wang, S. & Li, D. Occurrence and composition of solid bitumens from the Bulonggoer Devonian paleo-oil reservoir, North Xinjiang, China. Org. Geochem. 83, 1–15 (2015).
Gelpi, E., Schneider, H., Mann, J. & Oro, J. Hydrocarbons of geochemical significance in microscopic algae. Phytochemistry 9, 603–612 (1970).
Meyers, P. A. & Ishiwatari, R. Lacustrine organic geochemistry—an overview of indicators of organic matter sources and diagenesis in lake sediments. Org. Geochem. 20, 867–900 (1993).
Blokker, P. et al. The chemical structure of Gloeocapsomorpha prisca microfossils: implications for their origin. Geochim. Cosmochim. Acta 65, 885–900 (2001).
Eglinton, G. & Hamilton, R. J. Leaf epicuticular waxes. Science 156, 1322–1335 (1967).
Rieley, G. et al. Sources of sedimentary lipids deduced from stable carbon-isotope analyses of individual compounds. Nature 352, 425–427 (1991).
Freeman, K. H., Trendel, J. M. H. & Albrech, P. Evidence from GC-MS carbon-isotopic measurements for multiple origins of sedimentary hydrocarbons. Nature 353, 254–256 (1990).
Collister, J. W., Rieley, G., Stern, B., Eglinton, G. & Fry, B. Compound-specific δ13C analyses of leaf lipids from plants with differing carbon dioxide metabolisms. Org. Geochem. 21, 619–627 (1994).
Hockun, K. et al. Using distributions and stable isotopes of n-alkanes to disentangle organic matter contributions to sediments of Laguna Potrok Aike, Argentina. Org. Geochem. 102, 110–119 (2016).
Chevalier, N. et al. Precise indices based on n-alkane distribution for quantifying sources of sedimentary organic matter in coastal systems. Org. Geochem. 88, 69–77 (2015).
Yandoka, B. M. S., Abdullah, W. H., Abubakar, M., Hakimi, M. H. & Adegoke, A. K. Geochemical characterisation of Early Cretaceous lacustrine sediments of Bima Formation, Yola Sub-basin, Northern Benue Trough, NE Nigeria: Organic matter input, preservation, paleoenvironment and palaeoclimatic conditions. Mar. Pet. Geol. 61, 82–94 (2015).
Lichtfouse, E., Derenne, S., Mariotti, A. & Largeau, C. Possible algal origin of long chain odd n-alkanes in immature sediments as revealed by distributions and carbon isotope ratios. Org. Geochem. 22, 1023–1027 (1994).
Bush, R. T. & McInerney, F. A. Leaf wax n-alkane distributions in and across modern plants: implications for paleoecology and chemotaxonomy. Geochim. Cosmochim. Acta 117, 161–179 (2013).
Pichersky, E. & Raguso, R. A. Why do plants produce so many terpenoid compounds? New Phytologist 220, 692–702 (2016).
Chattopadhyay, A. & Dutta, S. Higher plant biomarker signatures of Early Eocene sediments of North Eastern India. Mar. Pet. Geol. 57, 51–67 (2014).
Caplan, M. L. & Bustin, R. M. Factors governing organic matter accumulation and preservation in a marine petroleum source rock from the Upper Devonian to Lower Carboniferous Exshaw Formation, Alberta. Bull. Can. Petrol. Geol. 44, 474–494 (1996).
Rimmer, S. M., Thompson, J. A., Goodnight, S. A. & Robl, T. L. Multiple controls on the preservation of organic matter in Devonian–Mississippian marine black shales: geochemical and petrographic evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 215, 125–154 (2004).
Visser, J. N. & Young, G. M. Major element geochemistry and paleoclimatology of the Permo-Carboniferous glacigene Dwyka Formation and postglacial mudrocks in southern Africa. Palaeogeogr. Palaeoclimatol. Palaeoecol. 81, 49–57 (1990).
Dingle, R. & Lavelle, M. Late Cretaceous–Cenozoic climatic variations of the northern Antarctic Peninsula: new geochemical evidence and review. Palaeogeogr. Palaeoclimatol. Palaeoecol. 141, 215–232 (1998).
Shen, J. et al. Volcanic perturbations of the marine environment in South China preceding the latest Permian mass extinction and their biotic effects. Geobiology 10, 82–103 (2012).
Fathy, D., Wagreich, M., Zaki, R., Mohamed, R. S. & Gier, S. Geochemical fingerprinting of Maastrichtian oil shales from the Central Eastern Desert, Egypt: Implications for provenance, tectonic setting, and source area weathering. Geol. J. 53, 2597–2612 (2018).
Morton, A. C. & Hallsworth, C. R. Processes controlling the composition of heavy mineral assemblages in sandstones. Sediment. Geol. 124, 3–29 (1999).
Hubert, J. F. A zircon-tourmaline-rutile maturity index and the interdependence of the composition of heavy mineral assemblages with the gross composition and texture of sandstones. J. Sediment Res. 32, 440–450 (1962).
Sageman, B. B. et al. A tale of shales: the relative roles of production, decomposition, and dilution in the accumulation of organic-rich strata, Middle–Upper Devonian, Appalachian basin. Chem. Geol. 195, 229–273 (2003).
Pujol, F., Berner, Z. & Stüben, D. Palaeoenvironmental changes at the Frasnian/Famennian boundary in key European sections: chemostratigraphic constraints. Palaeogeogr. Palaeoclimatol. Palaeoecol. 240, 120–145 (2006).
Lash, G. G. A multiproxy analysis of the Frasnian-Famennian transition in western New York State, USA. Palaeogeogr. Palaeoclimatol. Palaeoecol. 473, 108–122 (2017).
Joachimski, M. et al. Devonian climate and reef evolution: insights from oxygen isotopes in apatite. Earth Planet. Sci. Lett. 284, 599–609 (2009).
Algeo, T. J., Scheckler, S. E. & Maynard, J. B. Effects of the Middle to Late Devonian spread of vascular land plants on weathering regimes, marine biotas, and global climate. In Plants Invade the Land: Evolutionary and Environmental Perspectives (eds Gensel, P. G. & Edwards, D.) 213–236 (Columbia Univeristy Press, New York, 2001).
Driese, S. G., Mora, C. I. & Elick, J. M. Morphology and taphonomy of root and stump casts of the earliest trees (Middle to Late Devonian), Pennsylvania and New York, USA. Palaios 12, 524–537 (1997).
Gensel, P. G., Kotyk, M. E. & Basinger, J. F. Morphology of Above-and Below-Ground Structures in Early Devonian (Pragian–Emsian) Plants. In Plants Invade the Land: Evolutionary and Environmental Perspectives (eds Gensel, P. G. & Edwards, D.), 83–102 (2001).
Hao, S., Xue, J., Guo, D. & Wang, D. Earliest rooting system and root: shoot ratio from a new Zosterophyllum plant. New Phytol. 185, 217–225 (2010).
Kenrick, P. & Strullu-Derrien, C. The origin and early evolution of roots. Plant Physiol. 166, 570–580 (2014).
Retallack, G., Catt, J. & Chaloner, W. Fossil soils as grounds for interpreting the advent of large plants and animals on land. Philos. Trans. R. Soc. Lond B. Biol. Sci. 309, 105–142 (1985).
Morris, J. L. et al. Investigating Devonian trees as geo-engineers of past climates: linking palaeosols to palaeobotany and experimental geobiology. Palaeontology 58, 787–801 (2015).
Arnold, C. A. Observations on fossil plants from the Devonian of eastern North America. IV. Plant remains from the Catskill Delta deposits of northern Pennsylvania and southern New York. Contributions from the Museum of Paleontology, University of Michigan 5, 271–314 (1939).
Matten, L. C. The Givetian flora from Cairo, New York: Rhacophyton, Triloboxylon and Cladoxylon. Bot. J. Linnean Soc. 68, 303–318 (1974).
Beck, C. B. Periastron reticulatum Unger and Aerocortex kentuckiensis, n. gen. et sp., from the New Albany Shale of Kentucky. 65 , Am. J. Bot. 221–235 (1978).
Hammond, S. E. & Berry, C. M. A new species of Tetraxylopteris (Aneurophytales) from the Devonian of Venezuela. Bot. J. Linnean Soc. 148, 275–303 (2005).
Golonka, J. & Gawęda, A. Plate tectonic evolution of the southern margin of Laurussia in the Palozoic. Tectonics-Recent Advances 10, 261–282 (2012).
Rietveld, H. A profile refinement method for nuclear and magnetic structures. J. Appl. Crystallogr. 2, 65–71 (1969).
Ouyang, T. et al. Sensitivity of sediment magnetic records to climate change during Holocene for the northern South China Sea. Front Earth Sci. 4, 54 (2016).
Sun, Y., Chen, Z., Xu, S. & Cai, P. Stable carbon and hydrogen isotopic fractionation of individual n-alkanes accompanying biodegradation: evidence from a group of progressively biodegraded oils. Org. Geochem. 36, 225–238 (2005).
Acknowledgements
The study of the Chattanooga Shale from Alabama was funded by a faculty grant from the Gulf Coast Association of Geological Societies (GCAGS) to Y.H.L., a graduate student grant from GCAGS to M.L., and Newton/Winefordner Scholarship from the Geological Survey of Alabama to M.L. The biomarker analyses were performed on a GC-MS purchased through the support from the Instrumentation and Facilities Programs in the Division of Earth Sciences to Y.H.L. (NSF EAR-1255724). Kim Genareau allowed us (M.L.) to access to the SEM Lab at Geological Sciences, The University of Alabama. We thank Dr. Thomas J. Algeo for providing constructive and detailed comments that have contributed significantly to revisions and improvement of the manuscript. Y.H.L. thanks the six-month Sabbatical Fellowship from the South University of Science and Technology of Shen Zhen, China. The authors gratefully acknowledge use of the resources of the Alabama Water Institute at the University of Alabama.
Author information
Authors and Affiliations
Contributions
M.L., Y.H.L. and T.I. designed this project and wrote the manuscript. Field work was done by M.L., Y.H.L., T.I., N.H., Y.S. and I.C. Geochemical analyses were performed in various labs by M.L., Y.H.L. and Q.W. T.I. took care of the fossil diversity analysis. N.H. determined the conodont biostratigraphy. R.C. and M.L. examined plant remains under microscope. I.C. and J.P. contributed to information of geological features near the study site.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
41598_2019_43993_MOESM1_ESM.pdf
Supplementary material for Geochemical Evidence of the First Forestation in the Southernmost Euramerica from Upper Devonian (Famennian) Black Shales
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lu, M., Lu, Y., Ikejiri, T. et al. Geochemical Evidence of First Forestation in the Southernmost Euramerica from Upper Devonian (Famennian) Black Shales. Sci Rep 9, 7581 (2019). https://doi.org/10.1038/s41598-019-43993-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-43993-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
