
Abstract
Persistent hyperglycemia is causally associated with pancreatic β-cell dysfunction and loss of pancreatic insulin. Glucose normally enhances β-cell excitability through inhibition of KATP channels, opening of voltage-dependent calcium channels, increased [Ca2+]i, which triggers insulin secretion. Glucose-dependent excitability is lost in islets from KATP-knockout (KATP-KO) mice, in which β-cells are permanently hyperexcited, [Ca2+]i, is chronically elevated and insulin is constantly secreted. Mouse models of human neonatal diabetes in which KATP gain-of-function mutations are expressed in β-cells (KATP-GOF) also lose the link between glucose metabolism and excitation-induced insulin secretion, but in this case KATP-GOF β-cells are chronically underexcited, with permanently low [Ca2+]i and lack of glucose-dependent insulin secretion. We used KATP-GOF and KATP-KO islets to examine the role of altered-excitability in glucotoxicity. Wild-type islets showed rapid loss of insulin content when chronically incubated in high-glucose, an effect that was reversed by subsequently switching to low glucose media. In contrast, hyperexcitable KATP-KO islets lost insulin content in both low- and high-glucose, while underexcitable KATP-GOF islets maintained insulin content in both conditions. Loss of insulin content in chronic excitability was replicated by pharmacological inhibition of KATP by glibenclamide, The effects of hyperexcitable and underexcitable islets on glucotoxicity observed in in vivo animal models are directly opposite to the effects observed in vitro: we clearly demonstrate here that in vitro, hyperexcitability is detrimental to islets whereas underexcitability is protective.
Similar content being viewed by others

Introduction
In the pancreatic β-cell, ATP-sensitive K+ (KATP) channels play a critical role in coupling glucose metabolism to insulin secretion via control of membrane excitability, thereby maintaining blood glucose within a narrow physiologic range1. Increase in glucose metabolism leads to elevated intracellular ATP/ADP ratio and closure of KATP channels, with consequent membrane depolarization, leading to opening of voltage-dependent calcium channels and increased [Ca2+]i, which in turn triggers insulin secretion. Conversely, decrease in the metabolic signal opens KATP channels and suppresses the electrical trigger for insulin secretion1. Normally, the β-cell responds to chronic high glucose with a compensatory increase in β-cell mass to match the insulin secretory requirement. However, in genetically or otherwise predisposed individuals, β-cells are unable to sustain appropriate insulin secretory response and diabetes develops2. Persistent hyperglycemia may also lead to β-cell dysfunction and loss of insulin content3. Underlying mechanisms of this so-called glucotoxicity remain unclear, but the unique capacity of β-cells to increase oxidative phosphorylation in response to glucose availability makes them prone to increased reactive oxygen species production, which may underlie increased levels of oxidative stress markers and augmented apoptotic cell-death found in islets from type-2 diabetic organ donors4,5,6,7.
The importance of electrical activity in β-cell function is highlighted by the fact that KATP gain-of-function (GOF) mutations cause human Neonatal Diabetes Mellitus (NDM)8, KATP GOF polymorphisms are highly associated with type-2 diabetes9, and KATP loss-of-function (LOF) mutations underlie congenital hyperinsulinism (CH) (reviewed in1,10,11). Mouse models of NDM and CH due to genetically altered β-cell membrane excitability have been generated12,13,14,15,16,17,18,19. As expected, KATP-GOF mice, with underexcitable β-cells and permanently low [Ca2+]i, show low circulating insulin levels and develop diabetes12,20. Importantly, as diabetes progresses, KATP-GOF mice demonstrate a marked loss of insulin content, a typical secondary consequence of glucotoxicity12. However, the paradigmatic understanding that this glucotoxic loss of insulin content is the result of β-cell death is challenged by the demonstration of β-cell dedifferentiation, with no significant increase in apoptosis in pancreases from either diabetic mice or human type-2 diabetic organ donors21,22,23. Critically, we have demonstrated that this loss of β-cell mass in KATP-GOF mice is caused by loss of mature β-cell identity and dedifferentiation to neurogenin3+/insulin negative cells, rather than apoptotic cell death22.
Chronic hyperglycemia will lead to hyperstimulated metabolism, which will be constitutively coupled to hyperexcitability and chronically elevated [Ca2+]i, factors that have all been suggested to play a role in diabetic loss of β-cell function (Fig. 1). Moreover, decreased insulin secretion and mRNA and increased β-cell death have been demonstrated in multiple in vitro studies2,3,24,25. However, KATP-LOF and KATP-knockout (KO) mice, with chronically hyperexcitable β-cells and persistently elevated [Ca2+]i, do not show any obvious changes in insulin content or β-cell mass15,16,18,26,27, and KATP-KO islets have been reported to be less susceptible to the toxic effects of high glucose, oxidative stress and death28. Conversely, as discussed, there is dramatic secondary loss of insulin content in KATP-GOF mice that is not predicted as a direct consequence of their permanent underexcitability and low [Ca2+]i.

Schematic representation of the proposed key factors involved in pancreatic β-cell damage. In type-2 diabetes, hyperglycemia-induced hypermetabolism is constitutively coupled to membrane hyperexcitability, high [Ca2+]i and insulin hypersecretion but in KATP-LOF (congenital Hyperinsulinism) or KATP-KO mice hypermetabolism is uncoupled from hyerexcitability and high [Ca2+]i, which are persistently high in these mice. Conversely, in KATP-GOF induced neonatal diabetes, hyperglycemia-induced hypermetabolism is uncoupled from hyerexcitability and high [Ca2+]i since they are constitutively low in these mice. Grey boxes represent the initial changes, and some of them have been attributed to cause beta-cell damage, as pointed by red arrows.
In this study, we sought to determine the role of excitability in glucotoxic β-cell failure, and to ask whether this glucotoxicity is induced by hypermetabolism per se, or by the normally obligatorily coupled hyperexcitability and high [Ca2+]i. To do this, we have examined insulin content and secretion under glucotoxic conditions in islets isolated from mice in which the link between β-cell metabolism and excitability is lost. We have achieved this both genetically, using (i) islets from KATP-GOF mice with underexcitable β-cells and permanently low [Ca2+]i, and (ii) from KATP-KO mice with hyperexcitable β-cells with chronically elevated [Ca2+]i (Fig. 1), and pharmacologically by treatment of wild type islets with chronic KATP channel activator (diazoxide) or inhibitor (glibenclamide). We demonstrate that underexcitability is protective against glucotoxic conditions in vitro, while hyperexcitability induces loss of insulin content even under low-glucose conditions. These results do not correlate with those obtained in vivo, where animal models of altered β-cell excitability show the opposite effects, highlighting the importance of whole body physiology.
Results
Morphological changes induced by high glucose in islets with normal or altered membrane excitability
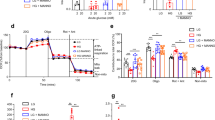
To directly test the influence of membrane excitability on glucotoxicity in β-cells, islets with normal (wild-type, KATP-WT), or with genetically increased- (Kir6.2 knockout: KATP-KO) or decreased-membrane excitability (Pdx-Cre/Kir6.2[K185Q, ΔN30] gain-of-function: KATP-GOF) were isolated from adult mice and chronically incubated in low (3 mM) or high (30 mM, mimicking the glucose concentration that islets are exposed in KATP-GOF mice) glucose for up to two weeks. Independent of the glucose concentration, or the genetically altered membrane excitability, the number of intact islets remaining in the dish did not change significantly over the period of time tested (at time 0: 30 ± 0 islets/dish on each genotype, after 10 day incubation: KATP-WT: 27.4 ± 0.91 islets/dish, KATP-GOF: 25.8 ± 1.2 islets/dish and KATP-KO: 24.6 ± 1.6 islets/dish). However, KATP-WT islets exposed to chronic high glucose became noticeably more translucent than islets incubated in low glucose (Fig. 2a,b) and were larger in diameter (Fig. 2c). KATP-KO islets (with increased membrane excitability) incubated in low glucose were more translucent at low or high glucose compared to control and KATP-GOF islets under the same conditions (Fig. 2a,b). Conversely, islets from KATP-GOF mice (with decreased excitability) maintained their morphology and diameter (Fig. 2) suggesting that membrane underexcitability is protective.

Islet Morphology. (a) Representative picture of pancreatic islets from KATP-WT or genetically altered mice 9 days after incubation in low or high glucose. (b) Quantification of mean gray values (A.U) for islet translucency on KATP-WT (black), KATP-KO (red) or KATP-GOF (green) islets chronically incubated in low- or high-glucose. n = 3 experiments from 9 independent mice, experiments were done in triplicates. Significant differences *p < 0.05, **p < 0.01, ***p < 0.001 and ****p < 0.0001 under the same condition; non-significant differences are not indicated in the figure. (c) Quantification of islet area (mm2) on KATP-WT (black), KATP-KO (red) or KATP-GOF (green) islets chronically incubated in low- or high-glucose. n = 3 experiments from 9 independent mice, experiments were done in triplicates.
Electrophysiology of islets chronically incubated in low and high glucose
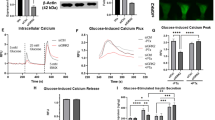
We performed current clamp recordings of membrane potential and voltage clamp recording of membrane conductance and capacitance using β-cells isolated from KATP-WT islets chronically incubated in low and high glucose. Figure 3 shows representative traces (a) and averages (b) of the membrane potential of β-cells (i) chronically incubated in low (3 mM) glucose following break-in in 1 mM glucose (blue), or (ii) chronically incubated in high (30 mM) glucose following break-in in 10 mM glucose (orange). As predicted for normal KATP behavior, the first are hyperpolarized (mean Vm = −64 ± 2 mV) due to active KATP channels (blue), whereas the second are depolarized (mean Vm = −49 ± 4 mV), but then hyperpolarize as KATP spontaneously activates following break-in (orange). Figure 3c shows voltage-clamp recording of membrane currents in response to voltage ramps (from −120 to −40 mV) from a cell chronically incubated in low (3 mM) glucose after break-in (in 1 mM glucose, blue), then after with spontaneous activation to maximum KATP (orange), then following subsequent KATP rundown (grey). The average maximum KATP current density in cells chronically incubated in 3 or 30 mM glucose is shown in Fig. 3d. KATP currents are present in both after the 10 day incubation period, although smaller in cells chronically incubated in high glucose. Figure 3e shows that cell capacitance, directly related to cell surface area, is approximately doubled in cells from islets chronically incubated in high glucose, correlating with the observed increased in islet area (Fig. 2c).

Beta-cell electrophysiology. (a) Representative traces of membrane potential during current clamp recordings from β-cells of KATP-WT islets chronically incubated in 3 mM glucose (blue) or 30 mM glucose (orange) in bath solution containing 1 mM or 10 mM glucose, respectively. (b) Average membrane potential upon break into β-cells from islets chronically incubated in 3 mM or 30 mM glucose. (c) Representative traces of current density in β-cell from islets chronically incubated in 3 mM glucose showing break-in current (blue), current at maximal KATP activation (orange) and current after KATP rundown (grey). (d) Average current density at −120 and −40 mV during maximal KATP activation is shown from islets chronically incubated at 3 mM (n = 13, blue) and 30 mM (n = 23, orange) glucose. (e) Capacitance of β-cells from islets chronically incubated in 3 mM (blue) and 30 mM (orange) glucose.
Insulin content in islets from genetically altered mice chronically exposed to high glucose
KATP-WT islets incubated in high glucose demonstrated a marked and rapid reduction of initial insulin content (to ~40%), over the first 3 days, which was subsequently maintained with no further decline over longer incubation times (Fig. 4a, black circles and squares, solid lines). This early reduction in insulin content was dramatically reversed in islets that were switched back to low glucose after 3 days incubation in high glucose, indicating a reversible effect with no permanent damage (Fig. 4a, black dashed line). Conversely, KATP-WT islets incubated in low glucose showed only a minimal reduction of insulin content over a period of 10 days. Strikingly, islets from KATP-KO mice (with increased membrane excitability and chronically elevated [Ca2+]i16) also demonstrated a rapid (3 days) reduction of insulin content to a similar plateau as KATP-WT islets, independent of whether incubated in low or high glucose (Fig. 4a, red circles and squares, solid lines). The reduction in insulin content observed in KATP-KO islets at low glucose was slightly greater when incubated in high glucose, and again this was reversed to the level maintained in low glucose when the media was switched from high to low glucose (Fig. 4a, red dashed line).

Glucotoxic changes in islet insulin content in vitro depend on membrane excitability. (a) Insulin content over time in islets from KATP-WT (black), KATP-KO (red) or KATP-GOF (green) mice chronically incubated in 3 mM (circles, solid line) or 30 mM (squares, solid line) glucose, or when media has been switch from high to low glucose (dashed lines). n = 3 experiments from 6 independent mice, experiments were done in triplicates. Significant differences *p < 0.05 respect to control under the same condition; non-significant differences are not indicated in the figure. (b) Total insulin content per islet at day 0 of incubation from KATP-WT (black), KATP-KO (red) or KATP-GOF (green) mice. (c) Insulin secreted into the media from KATP-WT (black), KATP-KO (red) or KATP-GOF (green) islets chronically incubated in 3 mM or 30 mM glucose at day 3.
As we previously described, KATP-GOF mice (with reduced β-cell excitability12 and chronically low [Ca2+]i20), demonstrate severe diabetes within 2 weeks of tamoxifen induction of transgene expression. In vivo, this is followed by reduction in β-cell mass and insulin content due to dedifferentiation from mature β-cells to progenitor-like cells, secondary consequences of chronic systemic diabetes12,22, which include correlate loss of insulin and elevation of glucose. To specifically isolate the influence of electrical activity in glucotoxic conditions, islets from KATP-GOF mice were harvested 10 days after tamoxifen-induction (i.e. prior to development of secondary consequences)12 and then chronically incubated in low or high glucose in vitro. In contrast to WT and KATP-KO islets, insulin content of KATP-GOF islets was completely preserved at low glucose, and only slightly reduced at high glucose (Fig. 4a, green circles and squares, solid lines), and again was completely reversed by switching the media from high to low glucose (Fig. 4a, green dashed line). These results indicate that membrane underexcitability protects from loss of insulin content in these glucotoxic conditions. KATP-WT, KATP-GOF and KATP-KO islets all had similar total islet insulin content on day 0 (Fig. 4b). As predicted, islets from KATP-KO mice demonstrate insulin hypersecretion when chronically incubated in low (3 mM) glucose, while KATP-WT and KATP-GOF do not secrete insulin in this condition (Fig. 4c, left). Moreover, while islets from KATP-WT mice secrete insulin when incubated in 30 mM glucose media, KATP-GOF show blunted secretion due to the mutation as expected (Fig. 4c, right).
High glucose effect on islets with pharmacologically altered membrane excitability
An alternative approach to manipulate membrane excitability is pharmacologically. WT islets were chronically incubated with the sulfonylurea glibenclamide (KATP channel inhibitor) to increase membrane excitability, or with diazoxide (KATP channel activator) to decrease membrane excitability, in parallel to the genetic approach above. As above, WT islets chronically incubated in high glucose demonstrated a marked decrease in insulin content compared to islets incubated in low glucose (Fig. 5a). Glibenclamide treatment caused a significant loss of insulin content in WT islets incubated in either low or high glucose (Fig. 5a). Conversely, diazoxide-treatment of WT islets resulted in preservation of insulin content in both low and high glucose conditions (Fig. 5a). These results mirror those obtained from KATP-KO and KATP-GOF islets, respectively (Fig. 4). To examine the consequences of insulin depletion per se in these experiments, exogenous insulin was added to WT islets incubated in low and high glucose. We demonstrate here that insulin prevented the high glucose-induced loss of insulin content (Fig. 5a).

Chronic pharmacologic manipulation of membrane excitability alters insulin content and secretion. (a) Insulin content in WT islets incubated for 10 days in 3 mM and 30 mM glucose, or plus the addition of the KATP channel inhibitor glibanclamide (1 µM) or the activator diazoxide (250 mM), or insulin (20 nM). Significant differences *p < 0.05 with respect to control under the same condition, non-significant are not indicated in the figure. Insulin secretion response to acute low (light grey bars) or high (dark grey bars). Glucose stimulated insulin secretion on WT islets chronically exposed to low glucose (b) or high glucose (c) plus glibenclamide or diazoxide. Significant differences *p < 0.05 with respect to chronic glucose alone under the same stimulatory condition, non-significant differences are not indicated in the figures. Inserts represent insulin secretion as a fraction of content.
Effects of chronic pharmacologically increased or decreased excitability on glucose-dependent insulin secretion
We examined the insulin secretory response to glucose challenge in WT islets incubated for 10 days in low or high glucose, in the absence or presence of KATP channel inhibitors or activators. WT islets chronically incubated in low glucose secreted insulin normally in response to acute high glucose stimulation (Fig. 5b). However, WT islets that had been chronically incubated in high glucose showed an unexpectedly high basal insulin secretion in response to acute low glucose, but blunted response to acute high glucose (Fig. 5c). Importantly, WT islets chronically incubated in low or high glucose in the presence of glibenclamide also showed increased insulin secretion when acutely exposed to low glucose (Fig. 5b), and a marked decrease in insulin secretion when exposed to high glucose for one hour (Fig. 5c). Conversely, islets chronically incubated with diazoxide (KATP activator, which results in electrical ‘rest’) demonstrated both increased basal and glucose-stimulated insulin secretion, compared to islets exposed to glucose alone (Fig. 5b,c). When insulin secretion was calculated as a fraction of insulin content, it is clear that chronic glibenclamide acutely stimulates increased basal secretion, whereas diazoxide inhibits glucose-dependent secretion, in both cases (Fig. 5b,c, inserts).
Proinsulin is increased in islets exposed to chronic high glucose
Because of the dramatic decrease in insulin content, we tested whether proinsulin biosynthesis was altered in genetically altered or pharmacologically treated islets. All islets exposed to chronic high glucose demonstrated a significant increase in proinsulin content, independent of the genotype (Fig. 6a) or pharmacologic treatment (Fig. 6b). At time 0, KATP-KO islets showed lower proinsulin content than WT (Fig. 6a, red circles and squares), whereas KATP-GOF islets demonstrated a markedly higher proinsulin level (Fig. 6a, green circles and squares). Conversely, all islets exposed to chronic low glucose demonstrated a significant decrease in proinsulin content over time, independent of the genotype (Fig. 6a) or pharmacologic treatment (Fig. 6b). These results demonstrate quite clearly that there is a positive effect of high glucose on proinsulin content, irrespective of membrane excitability.

Changes in proinsulin content in vitro in islsts genetically altered or pharmacologically treated with KATP channel inhibitors and activators. (a) Proinsulin content at 0, 3 and 9 days on KATP-WT (black), KATP-KO (red) or KATP-GOF (green) islets chronically incubated in low or high glucose. (b) Proinsulin content at 0, 3 and 9 days on WT islets chronically incubated in low or high glucose with or without the addition of glibencalmide (pink) or diazoxide (blue).
Discussion
Chronic effects of hyperglycemia on islet function
Progressive deterioration in β-cell function, increased levels of oxidative stress markers, loss of insulin content and decreased β-cell mass in vivo have long been recognized in the pathogenesis of type-2 diabetes4,5,6,24, regardless of therapy7,29. Chronic hyperglycemia and elevated free fatty-acids negatively impact β-cell function and insulin secretion, but mechanisms underlying secondary loss of insulin remain unclear. In otherwise normal islets, chronic hyperglycemia will cause a persistent increase in membrane excitability and permanently elevated [Ca2+]i, potential key factors in deterioration of β-cell function ultimately leading to β-cell death30,31. However, this paradigm of β-cell death in diabetes has now been challenged by the demonstration of loss of pancreatic β-cell identity and cell dedifferentiation in both mouse and human diabetes21,22,23,32. The largely unknown and potentially detrimental effects of abnormally high or low [Ca2+]i prompts us to ask the question: What role does membrane hyper- or hypo-excitability per se, and the consequently high or low [Ca2+]i, play in the deleterious effects of high glucose on islet function. By incubating islets from mice with genetically enhanced or reduced membrane excitability (i.e. KATP-KO and KATP-GOF mice respectively) in chronic high glucose in vitro, we can separate the role of excitability from otherwise linked in vivo factors in any glucotoxic response.
The role of membrane excitability in glucotoxicity
The unique capacity of β-cells to increase oxidative phosphorylation in response to glucose availability makes them prone to increased reactive oxygen species production, which may lead to mitochondrial dysfunction and ultimately cell death. In WT islets, the marked reduction of insulin content and glucose-stimulated insulin secretion (GSIS) when chronically incubated in high glucose is consistent with earlier studies demonstrating that rodent β-cells adapt to chronic high glucose by reducing maximal GSIS, but without alterations in pancreatic islet mass33. The maintenance of insulin content in KATP-GOF islets exposed to high glucose indicates that reduced membrane excitability protects them against such a loss of insulin content in vitro. In addition, this finding argues against any primary role of underexcitability and low [Ca2+]i20 in the loss of β-cell mass and especially insulin content that are observed in KATP-GOF islets in vivo12,22,34.
Conversely, reduced insulin content in KATP-KO mouse islets after chronic incubation in either low or high glucose suggests that chronic membrane hyperexcitability is a major determinant of insulin loss in vitro. Although there is no significant reduction in β-cell mass in normoglycemic KATP-KO mice15,16,35, these results suggest that elevated [Ca2+]i will be detrimental in the setting of hyperglycemia, consistent with studies indicating that high [Ca2+]i is a key factor in reduction of β-cell functionality in diabetes15,16,30,31,35.
Is increased membrane excitability, independent of altered metabolism, responsible for reduced insulin content?
Consistent with the idea that chronic β-cell hyperexcitability leads to a persistently elevated [Ca2+]i, and insulin hypersecretion with depletion of insulin stores24,36, our in vitro data show that increasing membrane excitability, either genetically (KATP-KO islets) or pharmacologically (glibenclamide-treated islets), does indeed lead to marked reduction in insulin content over ~3 days. Conversely, islets with genetically (KATP-GOF islets) or pharmacologically (diazoxide-treated) reduced membrane excitability maintain insulin content over the full 10 days, even when they are chronically incubated in high glucose. Persistent inhibition of WT KATP channels with glibenclamide induces similar reduction of insulin content to that observed in KATP-KO islets. These results correlate with studies demonstrating reduction of GSIS in rodent and human islets chronically incubated with sulfonylurea drugs, although these effects have only been observed in vitro, and not with consistency36,37,38. In addition, these results correlate with our previous observations that, in islets chronically exposed to glibenclamide, there is a mild increase in DNA synthesis; an effect that was completely abrogated by the presence of nifedipine (a calcium channel inhibitor), arguing that it is indeed driven by elevated [Ca2+]i,39. Moreover, our results are also consistent with those showing that islets chronically incubated in, or re-exposed to, sulfonylureas exhibited insulin secretion that occurs at lower [glucose] and accompanied by reduction in the maximal insulin response40,41.
Preserved insulin content and maintained GSIS (Fig. 4) in WT islets exposed to high glucose in the presence of diazoxide mimicked the effect observed in underexcited KATP-GOF islets, supporting the hypothesis that ‘rest’ from hyperexcitability and therefore from chronically elevated [Ca2+]i is protective against depletion of insulin content, and potentially against Ca2+-dependent apoptosis24,36. WT islets incubated in the presence of high glucose plus diazoxide have been reported to be more glucose sensitive, and to show a left-shift in glucose-stimulated insulin secretion, as well as a marked increase in DNA synthesis without changes in plasma KATP channels39,42,43. However, changes in β-cell oscillatory [Ca2+]i and KATP conductance and a shift in glucose sensitivity have also been reported in islets incubated overnight with high glucose plus diazoxide44 which is consistent with altered KATP trafficking to the plasma membrane. Our results also correlate with decreased insulin content and secretion in human islets incubated in high glucose for 4 days, effects that were partially reversed by addition of diazoxide in the incubation media45.
The concept of deficient insulin stores as a contributing factor to β-cell dysfunction in type-2 diabetes arose many years ago, based on the observation that type-2 diabetic patients show restoration of insulin response to oral sulfonylurea stimulation after a period of treatment with diazoxide or insulin, in order to induce β-cell ‘rest’46,47,48,49. Diazoxide treatment has also been shown to improve subsequent β-cell secretory function in 90% pancreatectomized diabetic rats and in streptozotocin-diabetic rats50, and to reverse the marked loss of glucose-induced rise in [Ca2+]i in human islets exposed to chronic high glucose51. Taken together, these in vitro and in vivo studies implicate electrical excitability-dependent loss of β-cell insulin content in high glucose conditions, which can be restored after exposure to low glucose and consequent suppression of excitation. Normally, islets do not accumulate large amounts of proinsulin since it is processed into mature insulin, which is then secreted. We began culturing islets from KATP-GOF mice at day 10 post tamoxifen at which point these mice are already hyperglycemic (>30 mM blood glucose levels) due to lack of insulin secretion. This will drive enhanced glucose metabolism, and since proinsulin biosynthesis is induced in response to glucose metabolism, elevated proinsulin in these islets may be expected52. In the KATP-KO model, since insulin is persistently secreted (reflected in accumulation of insulin in the media in low glucose condition), proinsulin accumulation is minimal. As expected, when any islets (controls, KATP-GOF or KATP-KO) are exposed to chronically high glucose, they showed increased proinsulin synthesis in response to elevated glucose in the media.
While precise mechanisms underlying the decrease in insulin content are not clear, our finding of increased proinsulin in all islets incubated in high glucose is consistent with inefficient conversion of proinsulin to insulin, potentially due to chronic glucose-induced oxidative and endoplasmic reticulum stress. It has been shown that, while glibenclamide increases and diazoxide decreases insulin secretion, there is no effect of these drugs on proinsulin biosynthesis (reviewed in53). These results are consistent with our observations that islets chronically incubated in high glucose, whether in the presence of either glibenclamide or diazoxide (which induce opposite effects on excitability and [Ca]i), leads to increased proinsulin, thus highlighting the effect of high glucose itself on proinsulin accumulation, rather than membrane excitability per-se. Our results also correlate with those demonstrating decreased proinsulin/insulin ratio in human islets treated with diazoxide at 11 mM glucose, which, by reducing excitability, preserves insulin content45. In obese mouse models of type-2 diabetes, islet proinsulin synthesis is typically increased and is accompanied by decreased mature insulin granules, due to dysfunctional insulin processing, effects that are reversed following incubation in low glucose media54. Consistent with the findings in rodent islets, human islets exposed to high glucose show accumulation and preferential secretion of proinsulin, secondary to depletion of mature insulin granules55.
Exogenous insulin prevention of high glucose-induced loss of insulin content
We have previously demonstrated that inclusion of insulin in the incubation media can protect islets from loss of endogenous insulin content39. Preservation of insulin content in WT islets that were co-incubated with high glucose and insulin is consistent with studies that demonstrate better preservation of β-cell function in newly diagnosed type-2 diabetic patients subjected to intensive insulin therapy than in those treated with sulfonylureas7,46. It has also previously been shown that insulin can acutely induce membrane hyperpolarization by activation of KATP channels56, although such a mechanism seems unlikely to contribute to the chronic preservation of insulin in the current experiments. At higher concentrations, DNA synthesis and islet proliferation can be induced by exogenous insulin39, but this is not likely at the concentration used in the present study.
In vitro versus in vivo models of glucotoxicity
We demonstrate here that chronic membrane hyperexcitability induced by high glucose conditions underlies loss of insulin content, and that membrane underexcitability protects islets from high glucose-induced loss of insulin content in vitro. It is important to note that pancreatic β-cells from mice with altered membrane excitability show the opposite long-term outcomes in vivo. We and others have previously demonstrated a marked reduction of insulin content, β-cell mass and insulin mRNA in islets from KATP-GOF diabetic mice with reduced membrane excitability12,13,14,22,34. This is the result of chronic systemic diabetes and correlates with findings demonstrated in other forms of diabetes21,33,57. The primary mechanism for loss of insulin was shown to be pancreatic β-cell dedifferentiation to progenitor-like cells in the severely diabetic state, with re-differentiation to mature β-cells following lowering of blood glucose by intensive insulin therapy22,33. Conversely, islets with chronically increased membrane excitability and increased [Ca2+]i (from KATP-KO15,16,18 and KATP-LOF mice20,26) show preserved insulin content in vivo. Moreover, chronic pharmacologic treatment with glibenclamide leading to increase in membrane excitability did not demonstrate any significant reduction in pancreatic islet insulin content in vivo35.
These differences observed in vivo versus in vitro may arise from the shorter duration of exposure to high glucose typically examined in vitro, including in this study, which might not be sufficient for the long-term consequences detected in vivo to occur. In addition, the in vitro absence of other nutrients such as amino acids and incretin hormones, which are known modulators of β-cell sensitivity to glucose, may contribute to in vivo consequences. We previously demonstrated that the in vivo environment is important for the islet response in KATP-GOF mice. Severely diabetic KATP-GOF mice show increased glucose metabolism at non-stimulatory glucose concentrations20, whereas KATP-GOF mice treated acutely with glibenclamide at disease onset can enter a sustained remission and maintain near normoglycemia34, effects that are characteristic of other animal models of β-cell overstimulation and diabetes34,35,58. Finally, it is also important to note that the types of stresses that β-cells are exposed to in vitro (e.g. islet isolation from vascularization, loss of endothelial cells, and changes in intracellular matrix) are absent in vivo and could influence the response to high glucose exposure. Thus, other in vivo factors could also play a role in the marked loss of drug responsiveness, β-cell function deterioration, reduction of insulin content and loss of β-cell mass in diabetes11.
β-cell rest from hypermetabolism, hyperexcitability or hypersecretion? What does ‘rest’ mean in potential recovery from β-cell dysfunction?
Finally, there has been much interest in the idea that exogenous insulin (and insulin plus diazoxide) treatment leads to β-cell ‘rest’, and that this permits restoration of β-cell function. It is unclear exactly what ‘rest’ means in this context: Does it mean rest from hyperstimulated metabolism, hyperexcitation, elevated [Ca2+]i, or from insulin secretion? Some studies clearly lean towards rest meaning rest from secretion. The present study provides one clear answer: our results show that rest from hyperexcitability (and its downstream consequences) is the key determinant of maintenance of insulin content in vitro. KATP-GOF islets (or WT islets treated with diazoxide), which are permanently in the ‘resting’ state in terms of excitability, [Ca2+]i, and insulin secretion maintain insulin content even in the chronic metabolically stimulated state of hyperglycemia. Conversely, KATP-KO islets (or WT islets treated with glibenclamide), which are chronically excited, lose insulin content rapidly in both high and low glucose conditions.
Methods
Mouse models of altered membrane electrical activity
All experiments were performed in compliance with the institutional guidelines of, and approved by, the Washington University Animal Studies Committee. KATP knockout16 and tamoxifen-inducible Pdx1PBCreERTM β-cell specific KATP-GOF (Kir6.2 [K185Q, ΔN30])12 mutant mice were previously generated. Control littermates were used in all experiments.
Pancreatic islet isolation
Mice were anesthetized with Isofluorane (0.2 ml) and killed by cervical dislocation, and the bile duct was cannulated and perfused with Hank’s solution (Sigma) containing collagenase (Collagenase Type XI, Sigma). The pancreas was removed and digested at 37 °C, hand shaken and washed in cold Hank’s solution. Islets were isolated by hand under a dissecting microscope and maintained overnight in CMRL-1066 (5.6 mM glucose) culture medium (GIBCO) supplemented with fetal calf serum (10%), penicillin (100 U/ml), and streptomycin (100μg/ml)12.
Islet morphology measurements
Following overnight incubation in CMRL media containing 5.6 mM glucose, 30 islets per group were chronically incubated for 10 days in CMRL-1066 containing low (3 mM) or high (30 mM) glucose (experiment done in triplicates). On day 9, islets images were obtained using the Leica DMI 4000B inverted microscope (Lecia microsystems, IL) under bright field. Islet mean grey value and islet area were analyzed by Fiji (ImageJ).
Electrophysiological experiments
Whole cell recordings were made using an Axopatch 200B amplifier and Digidata 1200 (Molecular Devices), as reported previously59. Voltage clamp recordings were performed using a ramp protocol from −120 mV to +40 mV with cells held at −70 mV during the inter-pulse interval. The bath solution was Tyrode’s solution contained 137 mM NaCl, 5.4 mM KCl, 1 mM MgCl2, 2 mM CaCl2, 0.33 mM NaH2PO4, 5 mM HEPES and 1 mM Glucose with pH adjusted to 7.4 mM with NaOH. The pipette solution contained 120 mM KCl, 10 mM HEPES and 1 mM K-EGTA, with pH adjusted to 7.4. For current clamp recordings current was held at 0pA. The bath solution used for current clamp recordings contained 10 mM glucose for cells from islets chronically incubated in 30 mM glucose and was unaltered for cells from islets chronically incubated at 3 mM glucose. Glass electrodes were pulled from Kimble-Chase 2502 plain capillary tubes using a P-87 puller (Sutter instruments). For voltage clamp recordings electrodes with 1–3 MΩ tips were used and for current clamp recordings 5–7 MΩ tips were used.
Islet manipulation and measurement of insulin secretion and content
30 islets per group were incubated at low (3 mM) or high (30 mM) glucose and were collected at day 3, 6 and 9 for measurement of insulin content. Insulin secreted in the media under 3 and 30 mM glucose conditions was measured on day 3. For reversibility experiments, isolated islets were incubated with high glucose for several days and then the media was changed to low glucose for the time indicated in the figures. Additionally, wild-type islets were chronically incubated for 10 days in low or high glucose in the presence of 1 μM glibenclamide or 250 µM diazoxide (to block or activate KATP, and thereby pharmacologically increase or decrease membrane excitability, respectively), or in the presence of 20 nM insulin. After the chronic treatment period, the islets were counted, and then pre-incubated in groups of 10 per well in 12 well plates in glucose-free CMRL-1066 plus 3 mM glucose, then incubated for 1 hour at 37 °C in CMRL-1066 plus low (3 mM) or high (30 mM) glucose for insulin secretion experiments. After the incubation period, the medium was removed and assayed for released insulin. Experiments were repeated in triplicate. For islet insulin content, groups of 10 islets were disrupted using ethanol-HCl extraction and sonicated on ice for estimation of insulin content. Insulin secretion and content were measured using either Rat Insulin radioimmunoassay (RIA, Millipore, St. Charles, MO)22 or Ultrasensitive Mouse Insulin Elisa kit (Crystal Chem, Elk Grove Village, IL)60 according to manufacturer’s procedure22.
Measurement of proinsulin content in islets
Islets were incubated overnight in CMRL media containing 5.6 mM glucose prior to all experiments. On days 0, 3, and 9, replicates of 10 islets per genotype was collected in microcentrifuge tubes, washed with PBS and re-suspended in acid-ethanol extraction buffer. Proinsulin content was calculated by utilizing the Mouse Proinsulin Elisa (Mercodia, Salem NC).
Statistics
Data are presented as mean ± SEM. Differences among groups were tested using analysis of variance (ANOVA) and post-hoc Duncan’s test. When only two groups were compared, unpaired t-tests were used to assess significance. *Indicates significant differences, with p < 0.05 respect to control condition. **Indicates significant differences p < 0.01, ***p < 0.001 and ****p < 0.0001.
References
Remedi, M. S. & Koster, J. C. KATP channelopathies in the pancreas. Pflugers Arch 460, 307–320 (2010).
Prentki, M. & Nolan, C. J. Islet beta cell failure in type 2 diabetes. J Clin Invest 116, 1802–1812 (2006).
Poitout, V. & Robertson, R. P. Glucolipotoxicity: fuel excess and beta-cell dysfunction. Endocr Rev 29, 351–366 (2008).
Cnop, M. et al. Mechanisms of pancreatic beta-cell death in type 1 and type 2 diabetes: many differences, few similarities. Diabetes 54(Suppl 2), S97–107 (2005).
Del Guerra, S. et al. Functional and Molecular Defects of Pancreatic Islets in Human Type 2 Diabetes. Diabetes 54, 727–735 (2005).
Butler, A. E. et al. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes 52, 102–110 (2003).
Wajchenberg, B. L. beta-cell failure in diabetes and preservation by clinical treatment. Endocr Rev 28, 187–218 (2007).
Gloyn, A. L. et al. Activating Mutations in the Gene Encoding the ATP-Sensitive Potassium-Channel Subunit Kir6.2 and Permanent Neonatal Diabetes. N Engl J Med 350, 1838–1849 (2004).
Villareal, D. T. et al. Kir6.2 variant E23K increases ATP-sensitive K+ channel activity and is associated with impaired insulin release and enhanced insulin sensitivity in adults with normal glucose tolerance. Diabetes 58, 1869–1878 (2009).
Stanley, C. A. Perspective on the Genetics and Diagnosis of Congenital Hyperinsulinism Disorders. The Journal of clinical endocrinology and metabolism 101, 815–826 (2016).
Remedi, M. S. & Emfinger, C. Pancreatic beta-cell identity in diabetes. Diabetes Obes Metab 18(Suppl 1), 110–116 (2016).
Remedi, M. S. et al. Secondary consequences of beta cell inexcitability: identification and prevention in a murine model of K(ATP)-induced neonatal diabetes mellitus. Cell Metab 9, 140–151 (2009).
Koster, J. C., Marshall, B. A., Ensor, N., Corbett, J. A. & Nichols, C. G. Targeted overactivity of beta cell K(ATP) channels induces profound neonatal diabetes. Cell 100, 645–654 (2000).
Girard, C. A. et al. Expression of an activating mutation in the gene encoding the KATP channel subunit Kir6.2 in mouse pancreatic beta cells recapitulates neonatal diabetes. J Clin Invest 119, 80–90 (2009).
Seghers, V., Nakazaki, M., DeMayo, F., Aguilar-Bryan, L. & Bryan, J. Sur1 knockout mice. A model for K(ATP) channel-independent regulation of insulin secretion. The Journal of biological chemistry 275, 9270–9277 (2000).
Miki, T. et al. Defective insulin secretion and enhanced insulin action in KATP channel- deficient mice. Proc Natl Acad Sci USA 95, 10402–10406 (1998).
Miki, T. et al. Abnormalities of pancreatic islets by targeted expression of a dominant-negative KATP channel. Proc Natl Acad Sci USA 94, 11969–11973 (1997).
Shiota, C. et al. Sulfonylurea receptor type 1 knock-out mice have intact feeding-stimulated insulin secretion despite marked impairment in their response to glucose. Journal of Biological Chemistry 277, 37176–37183 (2002).
Remedi, M. S. et al. Hyperinsulinism in mice with heterozygous loss of K(ATP) channels. Diabetologia 49, 2368–2378 (2006).
Benninger, R. K. et al. Defects in beta cell Ca(2)+ signalling, glucose metabolism and insulin secretion in a murine model of K(ATP) channel-induced neonatal diabetes mellitus. Diabetologia 54, 1087–1097 (2011).
Talchai, C., Xuan, S., Lin, H. V., Sussel, L. & Accili, D. Pancreatic beta Cell Dedifferentiation as a Mechanism of Diabetic beta Cell Failure. Cell 150, 1223–1234 (2012).
Wang, Z., York, N. W., Nichols, C. G. & Remedi, M. S. Pancreatic beta cell dedifferentiation in diabetes and redifferentiation following insulin therapy. Cell Metab 19, 872–882 (2014).
Cinti, F. et al. Evidence of beta-cell Dedifferentiation in Human Type 2 Diabetes. The Journal of clinical endocrinology and metabolism 101, 1044–1054 (2015).
Robertson, R. P., Harmon, J., Tran, P. O., Tanaka, Y. & Takahashi, H. Glucose toxicity in beta-cells: type 2 diabetes, good radicals gone bad, and the glutathione connection. Diabetes 52, 581–587 (2003).
Nichols, C. G. & Remedi, M. S. The diabetic β-cell: hyperstimulated vs. hyperexcited. Diabetes, Obesity and Metabolism 14 (2012).
Koster, J. C. et al. Hyperinsulinism induced by targeted suppression of beta cell KATP channels. Proc Natl Acad Sci USA 99, 16992–16997 (2002).
Rocheleau, J. V. et al. Critical Role of Gap Junction Coupled KATP Channel Activity for Regulated Insulin Secretion. PLoS Biology 4 (2006).
Gier, B. et al. Suppression of KATP channel activity protects murine pancreatic beta cells against oxidative stress. J Clin Invest 119, 3246–3256 (2009).
UKPDS-group. Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33).UK Prospective Diabetes Study Group. Lancet (London, England) 352, 837–853 (1998).
Grill, V. & Bjorklund, A. Overstimulation and beta-cell function. Diabetes 50(Suppl 1), S122–124 (2001).
Poitout, V. Glucolipotoxicity of the pancreatic beta-cell: myth or reality? Biochemical Society transactions 36, 901–904 (2008).
Brereton, M. F. et al. Reversible changes in pancreatic islet structure and function produced by elevated blood glucose. Nat Commun 5, 4639 (2014).
Jonas, J. C. et al. Glucose regulation of islet stress responses and beta-cell failure in type 2 diabetes. Diabetes Obes Metab 11(Suppl 4), 65–81 (2009).
Remedi, M. S., Agapova, S. E., Vyas, A. K., Hruz, P. W. & Nichols, C. G. Acute Sulfonylurea Therapy at Disease Onset Can Cause Permanent Remission of KATP-Induced Diabetes. Diabetes (2011).
Remedi, M. S. & Nichols, C. G. Chronic antidiabetic sulfonylureas in vivo: reversible effects on mouse pancreatic beta-cells. PLoS Med 5, e206 (2008).
Efanova, I. B. et al. Glucose and tolbutamide induce apoptosis in pancreatic beta-cells. A process dependent on intracellular Ca2+ concentration. The Journal of biological chemistry 273, 33501–33507 (1998).
Maedler, K. et al. Sulfonylurea induced beta-cell apoptosis in cultured human islets. The Journal of clinical endocrinology and metabolism 90, 501–506 (2005).
Del Guerra, S. et al. Effects of prolonged in vitro exposure to sulphonylureas on the function and survival of human islets. Journal of Diabetes & its Complications 19, 60–64 (2005).
Kwon, G. et al. Glucose-stimulated DNA synthesis through mammalian target of rapamycin (mTOR) is regulated by KATP channels: effects on cell cycle progression in rodent islets. The Journal of biological chemistry 281, 3261–3267 (2006).
Anello, M., Gilon, P. & Henquin, J. C. Alterations of insulin secretion from mouse islets treated with sulphonylureas: perturbations of Ca2+ regulation prevail over changes in insulin content. British journal of pharmacology 127, 1883–1891 (1999).
Rustenbeck, I., Dickel, C. & Grimmsmanns, T. Desensitization of insulin secretory response to imidazolines, tolbutamide, and quinine. II. Electrophysiological and fluorimetric studies. Biochem Pharmacol 62, 1695–1703 (2001).
Ma, Z., Portwood, N., Brodin, D., Grill, V. & Bjorklund, A. Effects of diazoxide on gene expression in rat pancreatic islets are largely linked to elevated glucose and potentially serve to enhance beta-cell sensitivity. Diabetes 56, 1095–1106 (2007).
Grill, V., Radtke, M., Qvigstad, E., Kollind, M. & Bjorklund, A. Beneficial effects of K-ATP channel openers in diabetes: an update on mechanisms and clinical experiences. Diabetes Obes Metab 11(Suppl 4), 143–148 (2009).
Glynn, E. et al. Chronic Glucose Exposure Systematically Shifts the Oscillatory Threshold of Mouse Islets: Experimental Evidence for an Early Intrinsic Mechanism of Compensation for Hyperglycemia. Endocrinology 157, 611–623 (2016).
Song, S. H., Rhodes, C. J., Veldhuis, J. D. & Butler, P. C. Diazoxide attenuates glucose-induced defects in first-phase insulin release and pulsatile insulin secretion in human islets. Endocrinology 144, 3399–3405 (2003).
Alvarsson, M. et al. Effects of insulin vs. glibenclamide in recently diagnosed patients with type 2 diabetes: a 4-year follow-up. Diabetes Obes Metab 10, 421–429 (2008).
Torella, R. et al. Restoration of sensitivity to sulfonylurea after strict glycaemic control with insulin in non-obese type 2 diabetic subjects. Diabete et Metabolisme 17, 443–447 (1991).
Weng, J. et al. Effect of intensive insulin therapy on beta-cell function and glycaemic control in patients with newly diagnosed type 2 diabetes: a multicentre randomised parallel-group trial. Lancet (London, England) 371, 1753–1760 (2008).
Greenwood, R. H., Mahler, R. F. & Hales, C. N. Improvement in insulin secretion in diabetes after diazoxide. Lancet (London, England) 1, 444–447 (1976).
Leahy, J. L., Bumbalo, L. M. & Chen, C. Diazoxide causes recovery of beta-cell glucose responsiveness in 90% pancreatectomized diabetic rats. Diabetes 43, 173–179 (1994).
Bjorklund, A., Lansner, A. & Grill, V. E. Glucose-induced [Ca2+]i abnormalities in human pancreatic islets: important role of overstimulation. Diabetes 49, 1840–1848 (2000).
Boland, B. B., Rhodes, C. J. & Grimsby, J. S. The dynamic plasticity of insulin production in beta-cells. Molecular metabolism 6, 958–973 (2017).
Uchizono, Y., Alarcon, C., Wicksteed, B. L., Marsh, B. J. & Rhodes, C. J. The balance between proinsulin biosynthesis and insulin secretion: where can imbalance lead? Diabetes Obes Metab 9(Suppl 2), 56–66 (2007).
Alarcon, C. et al. Pancreatic beta-Cell Adaptive Plasticity in Obesity Increases Insulin Production but Adversely Affects Secretory Function. Diabetes 65, 438–450 (2016).
Bjorklund, A. & Grill, V. Enhancing effects of long-term elevated glucose and palmitate on stored and secreted proinsulin-to-insulin ratios in human pancreatic islets. Diabetes 48, 1409–1414 (1999).
Khan, F. A., Goforth, P. B., Zhang, M. & Satin, L. S. Insulin activates ATP-sensitive K(+) channels in pancreatic beta-cells through a phosphatidylinositol 3-kinase-dependent pathway. Diabetes 50, 2192–2198 (2001).
Laybutt, D. R. et al. Critical reduction in beta-cell mass results in two distinct outcomes over time. Adaptation with impaired glucose tolerance or decompensated diabetes. The Journal of biological chemistry 278, 2997–3005 (2003).
Leahy, J. L., Bumbalo, L. M. & Chen, C. Beta-cell hypersensitivity for glucose precedes loss of glucose-induced insulin secretion in 90% pancreatectomized rats. Diabetologia 36, 1238–1244 (1993).
Emfinger, C. H. et al. Expression and function of ATP-dependent potassium channels in zebrafish islet beta-cells. R Soc Open Sci 4, 160808 (2017).
Yan, Z. et al. High-fat-diet-induced remission of diabetes in a subset of KATP -GOF insulin-secretory-deficient mice. Diabetes Obes Metab (2018).
Acknowledgements
This work was supported by NIH R01 DK098584 to M.S.R., NIH R01 DK109407 to C.G.N. Fellowship support was provided by NIH T32 DK108742 to Z.A.S. and NIH T32 HL125241 to N.W.Y. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. We thank Theresa M. Harter and Zihan Yan (Department of Cell Biology and Physiology and Medicine, Washington University School of Medicine, Saint Louis, MO) for assistance with mouse breeding, maintenance and genotyping.
Author information
Authors and Affiliations
Contributions
M.S.R., Z.W., Z.A.S. designed the study and C.G.N. designed the study. M.S.R., Z.W., N.W.Y., C.G.N. and Z.A.S. carried out the experiments and analyzed the data. M.S.R. wrote the paper, Z.A.S. provided comments, and C.G.N. and M.S.R. edited it. M.S.R. is a guarantor of this manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shyr, Z.A., Wang, Z., York, N.W. et al. The role of membrane excitability in pancreatic β-cell glucotoxicity. Sci Rep 9, 6952 (2019). https://doi.org/10.1038/s41598-019-43452-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-43452-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
