
Abstract
The question of whether the iconic avialan Archaeopteryx was capable of active flapping flight or only passive gliding is still unresolved. This study contributes to this debate by reporting on two key aspects of this fossil that are visible under ultraviolet (UV) light. In contrast to previous studies, we show that most of the vertebral column of the Berlin Archaeopteryx possesses intraosseous pneumaticity, and that pneumatic structures also extend beyond the anterior thoracic vertebrae in other specimens of Archaeopteryx. With a minimum Pneumaticity Index (PI) of 0.39, Archaeopteryx had a much more lightweight skeleton than has been previously reported, comprising an air sac-driven respiratory system with the potential for a bird-like, high-performance metabolism. The neural spines of the 16th to 22nd presacral vertebrae in the Berlin Archaeopteryx are bridged by interspinal ossifications, and form a rigid notarium-like structure similar to the condition seen in modern birds. This reinforced vertebral column, combined with the extensive development of air sacs, suggests that Archaeopteryx was capable of flapping its wings for cursorial and/or aerial locomotion.
Similar content being viewed by others

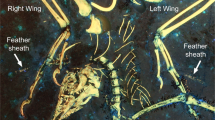

Introduction
Living birds have extensively pneumatized postcrania, due to their unique and extremely efficient respiratory system1,2. This adaptation includes a series of pneumatic diverticula that derive from large air sacs, encompass the lungs, and invade different parts of the skeleton3, leaving clear marks (pneumatic foramina) found in both avian and non-avian dinosaurs4,5,6. Reconstructions of pneumatic structures in fossil archosaurs have been previously based on comparisons with extant birds1,2,5,6,7,8. Evidence used to infer intraosseous pneumaticity for dinosaurs, in particular isolated foramina in compact bone or “blind” fossae, remains ambiguous however, as these cavities can house other structures such as muscles, fat, or neurovascular tissue1. The only unambiguous, reliable indicators of intraosseous pneumaticity are cortical foramina and communicating fossae connected with larger cavities within the bone1. Furthermore, the locations of foramina in the vertebrae or limb bones of dinosaurs that are comparable to those in extant birds support the interpretation of such structures as pneumatic1,2,7. The large cavities within pneumatic vertebrae of dinosaurs have been descriptively separated according to their architecture into larger and rounded camerae, and smaller and more angular-walled camellae8,9,10. Both types of internal pneumatic structures can occur in the same skeletal element, and multiple camellae can create a specific honeycomb-like pattern as in extant birds, described as somphospondylous10. The anteroposterior extension and spatial distribution of intraosseous pneumaticity provides critical anatomical evidence enabling reconstructions of the respiratory apparatus. The extent of pneumaticity also enables assessment of body weight, locomotor preferences, and metabolic activity, and therefore provides insights on the paleobiology and behavior of extinct animals5,6,7.
In Archaeopteryx, both the presence and extent of intraosseous pneumaticity has been controversially discussed ever since the 19th century11. A recent microtomographic study on the Daiting specimen (Archaeopteryx albersdoerferi) showed that all cranial bones, shoulder girdles, and wing bones contain internal pneumatic cavities12. Similar observations have been based on the Berlin, London, and Eichstätt specimens, which preserve foramina on the surfaces of their presacral vertebrae and pubes1,7,13,14,15. Pneumatic foramina have been described in the 2nd to 5th cervical vertebrae1,7,13,15 and posterior presacral vertebrae of the Berlin Archaeopteryx16,17; this specimen is also known to have hollow thoracic ribs13. The most unequivocal pneumatic structure seen in the postcranial skeleton of the Berlin specimen is the pneumatic foramen in the body of the 5th cervical vertebra1, exemplifying the ‘common pattern’ seen in earlier-diverging theropod dinosaurs7.
Here, we describe intact and incomplete postcranial bone surfaces in the Berlin Archaeopteryx (MB.Av.101; MB = Museum Berlin, Av = Collection of fossil birds), utilizing long-wave UV light15,18 to reveal pneumatic structures that are hidden, or difficult to discern, under visible light conditions (Fig. 1). Previous work has shown that UV light is far more sensitive than visible light to the increased contrast between fossilized bone, rocky infill, and surrounding matrix15,18,19,20,21. A previous description based on UV observations of the Berlin Archaeopteryx (MB.Av.101)15 has corroborated other reports1,13 regarding the presence of pneumatic foramina in the cervical vertebral column and hollow vertebral bodies for the 7th to 10th presacral vertebrae. The new UV findings we present here extend these results, enable a detailed account of all unambiguous pneumatic structures in the postcranial skeleton of MB.Av.101, and confirm that numerous postcranial bones of Archaeopteryx were reduced in mass via hollow interiors (Table S1).

Photographs and interpretative drawing of cervical vertebrae of the Berlin specimen of Archaeopteryx, MB.Av.101. (a) UV photograph of the neck and skull, (b) photograph of the neck under visual light, structures previously described and figured12 are marked, (c) magnified frame of (a) showing details of pneumatic structures in the neck, (d) interpretative drawing of (c) with recognized pneumatic structures. Anatomical abbreviations used: abs, area of abraded bone surface with regular spongy internal structure; amf, area with unidentified pores or putative camellae; cam, pneumatic camellae; CR, cervical rib; for, foramen; fos, fossa; nc, neural canal; pf, pneumatic foramen. Pneumatic foramina and camellae are unambiguous pneumatic structures and labeled with bold face. Foramina and pneumatic foramina are filled with black; fossae are filled with grey but without margin. Scale bars are 10 mm.
Results
In the Berlin Archaeopteryx, the vertebral cortex has been abraded in most presacral vertebrae, possibly due to repeated preparation15,18, exposing the internal bone structure of the vertebrae and ribs. UV light conditions expose pneumatic structures more clearly than under visible light, and reveal a far more extensive distribution of intraosseous pneumaticity than has been described since the 19th century. Sharp-lipped pneumatic foramina, which are distinguished from nutrient foramina by their larger diameter13, are present at the vertebral body and neural arch of the 2nd to 5th cervical vertebrae, as reported before1,13,15. Pneumatic foramina can also be identified on the 6th, 16th, and 22nd presacral vertebrae (Figs 1 and 2, Table S1). In the 19th presacral vertebra, a combination of a fossa with a communicating foramen is visible at the vertebral body (Fig. 2d). In contrast, small foramina visible in most of the vertebrae likely represent nutrient foramina, and are therefore not evidence for intraosseous pneumaticity1,13. In the 8th to 14th presacral vertebrae (Fig. 2), and the 1st to 3rd, 12th, and 14th to 16th caudal vertebrae (Fig. 3), the interiors exhibit large internal pneumatic chambers, or camerae. The 3rd, 8th to 14th, 16th, and 20th presacral vertebrae, and the 2nd, 5th, 6th and 11th caudal vertebrae expose pneumatic camellae. These camellae are small, sometimes rounded but mostly angular voids, separated from each other by thin bone walls (Figs 1 and 2b,c); they form a characteristic honeycomb-like pattern that can also be observed in the pneumatic vertebrae of extant birds1. In most vertebrae, the camellae are combined with unambiguous pneumatic foramina and large internal camerae (Table S1) typical for pneumatized bones, and are more often preserved in the neural arches than in the vertebral bodies.

Photographs and interpretative drawing of presacral (thoracic) vertebrae with pneumatic structures of the Berlin specimen of Archaeopteryx, MB.Av.101. (a) UV photograph, (b) magnification of 16th presacral vertebra under UV light, and (c) the same vertebra under visible light, (d) interpretative drawing of region a. Anatomical abbreviations used: abs, area of abraded bone surface with regular spongy internal structure; amf, area with unidentified pores or putative camellae; cam, pneumatic camellae; for, foramen; fos, fossa; ipc, internal pneumatic camerae; nc, neural canal; pf, pneumatic foramen. Pneumatic foramina, internal camerae and camellae are unambiguous pneumatic structures and labeled with bold face. Foramina and pneumatic foramina are filled with black; fossae are filled with grey but without margin. Scale bars are 10 mm.

UV photographs and interpretative drawings of the pelvis and sacral and caudal vertebrae with pneumatic structures of the Berlin specimen of Archaeopteryx, MB.Av.101. (a) Pelvic region in lateral view, with the last (22nd) presacral vertebra, (b) 1st to 5th caudal vertebra, (c) 5th to 9th caudal vertebrae, (d) 10th to 15th caudal vertebrae, (e) 14th to 21st (last) caudal vertebrae. Anatomical abbreviations and filling of structures as in Figs 1 and 2. Scale bars are 10 mm.
At the vertebral bodies, the abraded cortical bone exposes an internal structure that consists of regular, small, and mostly oval pores that are only half as large as the camellae (abs). Rounded and irregularly distributed voids the same size as camellae are visible (amf) in other areas, mostly along the neural arches and in particular all presacral vertebrae and the 3rd and 6th to 8th caudal vertebra (Fig. 1–3). These structures cannot be unambiguously identified as pneumatic structures, as they do not correspond to a typical camellate pattern as described above, but might instead represent areas of spongy bone. The presence of spongy internal bone structure can be taken as evidence for lightweight vertebrae in MB.Av.101. The thoracic ribs also exhibit a pattern comprising small regular and irregular voids in their heads and along their shafts, while their internal shafts are hollow13 (Fig. 2a,d; Table S1). A pattern of small and irregular pores can be observed in the posterior process and ventral margin of the ilium, and the head of the pubis (Fig. 3). Only small putative nutrient foramina are observed within the humerus, pelvic bones, femur, and tibia (Figs 2a,d and 3a, Table S1), so unambiguous evidence for intraosseous pneumaticity in the appendicular skeleton is still unknown1,8.
The overall skeletal pneumaticity in Archaeopteryx was calculated using the Pneumaticity Index (PI), as it provides a quantitative measure for the standardized comparison of postcranial pneumaticity among multiple avian species, independent of phylogeny, body size, and behavior22. Mapping of intraosseous pneumaticity in the Berlin Archaeopteryx yielded a PI of at minimum 0.39 (7 of 18 anatomical units being pneumatic). This value is comparable to the PI of a few of the extant anseriform birds previously reported, such as Mergus merganser and Anas georgica22 (both 0.41).
Our UV light observations also corroborate the presence of a notarium-like structure of fused presacral vertebrae in the Berlin Archaeopteryx. Fused anterior thoracic vertebrae have been mentioned briefly before23, but never described in detail. Our images reveal the presence of a massive, rod-shaped ossification above the apex of the 16th presacral neural spine and connecting to the anterior spine of the 17th (Fig. 4a,b). The caudodorsal corner of the 17th neural spine and the craniodorsal corner of its 18th counterpart comprise a long, thin, rod-like structure that bridges and tightly binds the neural spines together (Fig. 4a,b). The neural spines of the 19th to 20th presacral vertebrae are equally drawn out in their anterodorsal and posterodorsal corners, and fused dorsally with their adjacent neural spines. However, the connection between these elements is partly broken (Fig. 4a,b), while the neural spine of the 21st presacral vertebra has extended anterodorsal and posterodorsal corners.

UV photographs of the Berlin specimen of Archaeopteryx, MB.Av.101, with magnified region of the fused presacral neural spines forming a notarium-like structure. (a) Axial skeleton of Archaeopteryx, white frame demarks area of fused neural spines; scale bar is 20 mm. (b) Magnification of the 16th to 22nd presacral vertebrae, white arrows mark ossifications between the neural spines and remnants of broken spinal processes; scale bar is 10 mm.
Discussion
It is generally accepted that the clearest marker of skeletal pneumaticity is the presence of intraosseous pneumatic structures correlated with foramina1,7,24. Hitherto, the presacral distribution of pneumatic structures in Archaeopteryx has been discussed solely on the basis of described pneumatic foramina, which is considered controversial1,7. Previous workers have noted the existence of such pneumatic foramina on the surfaces of the cervical and thoracic vertebrae, humerus, and pubis of the Berlin, London, Eichstätt, and Thermopolis specimens of Archaeopteryx1,7,8,12,13,14,15,16,17,25 (Table S2). However, the most recent integrative study concluded that just the cervical and anteriormost thoracic vertebrae were pneumatized in Archaeopteryx, analogous with the plesiomorphic ‘common pattern’ seen in basal theropod dinosaurs7. In this study, we expand our knowledge of the Berlin Archaeopteryx with the first evidence of unambiguous intraosseous pneumatic structures (i.e., internal cavities such as camerae and camellae). The UV findings now enable convincing identification of internally pneumatic presacral vertebrae (2nd to 14th, 16th, 20th, 22nd) and caudal vertebrae (1st to 3rd, 5th, 6th, 11th, 12th, 14th to 16th), demonstrating the extension of pneumatic foramina and intraosseous pneumatic structures to most the vertebral column. Unambiguous traces of vertebral pneumaticity are absent in the remaining thoracic vertebrae, which is consistent with other specimens of Archaeopteryx1,7,13,14,26. The thoracic ribs are hollow, which has also been reported for the12th specimen of Archaeopteryx26. The presence of intraosseous pneumaticity in the remaining caudal vertebrae remains unclear, due to the lack of a combination of unambiguous pneumatic structures. We found evidence for at least a spongy and therefore light bone architecture in the 1st and 2nd sacral vertebra, the ilium and the pubis, as well as in some other caudal vertebrae (Fig. 3).
Our survey revealed furthermore that the abraded and broken lumbar vertebra of the Maxberg specimen27 also includes a pneumatized internal vertebral body including camellae (Table S2). Additionally, X-ray images of the thoracic vertebrae of the Maxberg Archaeopteryx27 and the cervicals of the Thermopolis specimen25 corroborate the presence of camellae within these vertebrae (Table S2). Additionally, camellate intraosseous pneumatic structures in the last two cervical vertebrae, as well as a pneumatic foramen in the 1st thoracic vertebra (Rauhut et al. 2018, p.30; Fig. 17), have been described for the most recently reported 12th specimen of Archaeopteryx26. Therefore, the presence of intraosseous postcranial pneumaticity in at least four specimens of Archaeopteryx including the Berlin, Maxberg, Thermopolis, and the 12th specimen, is documented (Table S2). At least in two specimens, the Maxberg and the Berlin Archaeopteryx, pneumatic structures extend outside the region of the anterior thoracic vertebrae. A hollow caudal vertebra is described for the 12th Archaeopteryx26, although it is unclear whether this represents intraosseous pneumaticity. Clearly seen in the Berlin Archaeopteryx, the pattern of intraosseous pneumaticity comprises a combination of closely-spaced honeycomb-shaped camellae bounded by thin bone walls (Fig. 2b), essentially similar to that of extant birds1,6,8,10. Camellate pneumatic bone is also found in a range of non-avialan dinosaurs (e.g. Aerosteon28, Tyrannosaurus29, and other neotheropods8) and in the avialan Rahonavis30, whereas large internal pneumatic camerae have been less frequently reported (e.g., in megalosauroids31, dromaeosaurids8 and ornithomimosaurs32).
Our new data demonstrate that the pneumaticity status of Archaeopteryx does not, in fact, conform to the plesiomorphic “common pattern” of non-avialan theropod dinosaurs7. Instead, Archaeopteryx possesses what can be termed “extended pattern of pneumaticity” (EPP)7, which includes the posterior thoracic vertebrae, and at least some anterior caudal vertebrae. The EPP is also seen in basal Neotheropoda6,7, as well as in most neotheropod clades, including Tyrannosauroidea28, Oviraptorosauria33,34,35, Dromaeosauridae36, and Troodontidae37,38, where it is thought to have evolved independently7. Among Maniraptora, only Oviraptorosauria possesses a stronger pneumatized vertebral column, including the complete tail39. Apart from Archaeopteryx, evidence of postcranial pneumaticity within Avialae is patchy, possibly related to the small body size and fragmentary preservation of most specimens. Pneumatic foramina are documented for the cervical vertebrae of confuciusornithid birds40, and for the cervical and anterior thoracic vertebrae of Rahonavis and some enantiornithid birds30, which would conform to a “common pattern” of intraosseous pneumaticity. One report of a pneumatic foramen in the humerus of an enantiornithid bird41 might indicate an EPP, but the evolutionary development of this pattern within Avialae still remains unclear. In any case, the level of pneumaticity we found in Archaeopteryx already corresponds with the EPP seen in some extant neornithine birds, including non-diving anseriforms22.
The new evidence of intraosseous pneumatization in Archaeopteryx presented here indicates that the respiratory system of this avialan included an air sac distribution similar to that of living birds1,3,6,22 (Table S1). The extensive intraosseous postcranial pneumatic structures provide unambiguous evidence for the presence of cervical air sacs that pneumatize cervical vertebrae and ribs, as well as anterior to mid-thoracic vertebrae and thoracic ribs. Intraosseous pneumatization of the mid- to posterior thoracic and caudal vertebrae provide unambiguous evidence for the existence of abdominal air sacs1,3,22 (Fig. 5). There is no osteological evidence for the presence of clavicular and anterior thoracic air sacs surrounding the lungs, which would pneumatize the humerus22 (Fig. 2, Table S1). The exact position of the lungs also cannot be reconstructed, because this organ is not associated with osteological traces; the lack of pulmonary foramina in thoracic vertebrae of Archaeopteryx makes it unlikely that lung tissue pneumatized these vertebrae on a large scale1. Similarly, the presence of posterior thoracic air sacs, present in extant birds, cannot be reconstructed for Archaeopteryx because these air sacs do not pneumatize the skeleton1,3,22. The reconstruction of air sacs in Archaeopteryx (Fig. 5) confirms the already assumed1,7 presence of a bird-like, high-compliance lung-air sac respiration system, incorporating lungs fixed at the rigid thoracic vertebral column and ventilated by air sacs positioned anteriorly and posteriorly to it.

Skeleton of Archaeopteryx (Berlin specimen, MB.Av.101) with schematic representation of air sacs and pneumatic postcranial elements as deduced from this study. Skeleton is shown from left lateral side and paired elements are represented only from the left body side. Black arrows indicate the path of pneumatization within air sacs. Light blue = cervical air sac (a), red = lung (b), dark blue = abdominal air sac (c). Grey = clavicular (d), anterior thoracic (e), and posterior thoracic (f) air sacs; these have no osteological evidence in Archaeopteryx, making their presence uncertain. Skeleton redrawn from Wellnhofer26.
The rostrad extension of skeletal pneumatization in theropod dinosaurs is a “centrum-first” pattern7, which means that first the vertebral bodies are pneumatized via pneumatic foramina (“intramural pneumatization”)42, followed by the neural arches. This is consistent with the occurrence of pneumatic foramina throughout the cervical vertebral column of the Berlin specimen (Fig. 1). In the posterior direction, pneumatization proceeds in a “neural arch-first” pattern, with the neural arches being pneumatized first, followed by pneumatization of the vertebral bodies1,7. Pneumatic diverticula can proceed through the medullary region first, and enter the vertebral body internally, as in extant birds1,10,43. This pattern likely explains the lack of pneumatic foramina observed in the thoracic vertebral bodies in Archaeopteryx (Fig. 2), which contrasts its rich intraosseous pneumatic structures. Such an absence of pneumatic foramina in the mid- thoracic region is consistent with many extant birds, in which pneumatic foramina are reduced in the thoracic vertebral series towards the sacrum1.
Pneumatization of the postcranial skeleton has a direct weight-reducing effect, by replacing bone and marrow with air-filled cavities42,44,45. A correlation between the degree of vertebral pneumaticity and body size was found in large-bodied, non-avian theropod dinosaurs7. In extant birds, the degree of postcranial pneumaticity has traditionally been linked to body mass and diving behavior, with the assumption that larger-bodied birds are more pneumatic than smaller-bodied birds, and the known absence of intraosseous pneumaticity in diving birds such as penguins22. At least within anseriform birds, no statistically significant relationships between body size and Pneumaticity Index has been found22, so that reduced or absent intraosseous pneumaticity (such as in diving taxa) seems not to be generally correlated with the absence of active flight. In smaller extant birds, the threshold of a critical body mass related to the energetic requirements of flight is not reached by a reduction or increase of intraosseous pneumaticity, meaning that the amount, presence, or absence of postcranial intraosseous pneumaticity gives no information on the flight capability of an extant bird22. However, if the metabolic activity of a fossil avialan such as Archaeopteryx was different than that of extant birds, then the increase of intraosseous pneumaticity might very well have had an effect on the critical body mass, and thereby also the flight capability in Archaeopteryx. Nevertheless, given the remaining uncertainty in the reconstruction of both the total extension of postcranial intraosseous pneumaticity and the presence of all pulmonary air-sacs, the precise estimation of body mass in Archaeopteryx remains speculative. A hypothetical adult (somatically mature) Archaeopteryx has been reconstructed to exceed 500 mm in body length and have a body mass between 0.8 and 1 kg, comparable to a raven (Corvus corax)46, values which we assume to be within the possible ranges for this taxon and not contradicted by the new data on intraosseous pneumaticity in this study.
The documentation of stiffening structures in neural spines of the thoracic vertebral column of Archaeopteryx is rather novel, and we argue that these ossifications represent the earliest known occurrence of a notarium-like structure on the line to extant birds, pushing back its origin to Paraves. Previously, spinal processes in the thoracic vertebrae had only been described in late Cretaceous enantiornithine birds41. It is also interesting to note the absence of stabilization structures in basal birds more derived than Archaeopteryx, such as the Confuciusornithidae, as well as the heterogeneous distribution of notaria in extant birds47,48,49,50,51. The apparent absence of these structures in other specimens of Archaeopteryx must remain speculative, and might be due to differing degrees of tendon ossification and/or preservation in these specimens. In several members of extant avian groups such as Tinamiformes, Podicipediformes, Phoenicopteriformes, Galliformes, Columbiformes, Pelecaniformes, Falconiformes, Gruiformes, and Caprimulgiformes, the vertebrae of the thorax form a notarium; this is a rigid structure created by 2–5 ankylosed vertebrae, including ossified tendons and ligaments, and separated in most cases from the synsacrum by at least one “free” or unankylosed vertebra47,48,49,50,51. Importantly, the fusion of neural spines in Archaeopteryx lies in the thoracic vertebral column between the 16th and the 22nd presacral vertebrae (Fig. 4a,b), corresponding to the notarium region in extant avians51, and there is also a vertebral gap between the fused and sacral regions. The narrow, ‘rod-like structures along the neural spines are interpreted to represent ossified ligaments52 or tendons of the epaxial trunk musculature. These would have acted as bony splints52, and likely contributed to an incipient ankylosis between the vertebrae (although the vertebral bodies themselves are not fused with each other).
A relatively rigid thoracic vertebral column is required for a bird-like respiration mechanism3, and is therefore an important structure to be documented in Archaeopteryx. In extant birds, the biological importance of the notarium is explained mostly as a rigid stabilization structure, counteracting mechanical stress occurring during wing-driven flapping flight and acting as a shock absorber during landing50. A notarium has also been described for some pterosaurs as an adaptation to active flight53,54. In any case, fusion of the neural spines creates a more stable (but less flexible) region in the vertebral column, which acts as a unit and can be more effectively stabilized by the available muscles and ligaments against the mechanical forces during locomotion.
Ultimately, reinforcement structures in the thoracic vertebral column can be interpreted to facilitate novel locomotor modes, in particular the development of active flapping flight or bipedal running supported by flapping wings, that increases stresses acting on the vertebral column. There has been controversial anatomical evidence on the ability of Archaeopteryx for gliding or flapping flight27,55,56,57,58,59. Nevertheless, the reinforcement structures observed in the trunk is indirect evidence for the increased use of forelimbs in Archaeopteryx, and depicts a stabilization trend that continues on the line to extant birds with the reduction in the number and increasing intervertebral fusion of thoracic vertebrae into a regional notarium47,48,52.
Conclusions
We demonstrate that Archaeopteryx possessed a derived, bird-like postcranial pneumatization pattern that comprises intraosseous pneumatic camellae and camerae within the presacral vertebral column and caudal vertebrae. The new observations furthermore confirm that expanded postcranial pneumatic structures and a bird-like respiratory system, both important physiological adaptations for powered flight, were present in Archaeopteryx. Whereas the number of pneumatic bones has a direct effect on weight reduction in large-bodied dinosaurs5,7, the camerate and camellate architecture of pneumatized bone is more likely determined by mechanical factors in the vertebral column, in concert with the evolutionary development and phylogenetic integration of the taxa. In Archaeopteryx, the direct effect on weight-reduction of the vertebral column by intraosseous pneumatic structures was moderate. As an indirect effect, the lightened pneumatic vertebral column would have needed less muscle forces to be stabilized against the mechanical loads that occur during all locomotor modes. Intraosseous pneumatization in Archaeopteryx was important for the metabolism of the animal, because the pneumatic epithelium replaces metabolically costly and massive bone, which reduces metabolic energy consumption and locomotion costs, and helped to increase the metabolic performance of the animal7,45,59,60. The putative high energetic advantage of a pneumatic postcranium7,24 demonstrates the particular importance for a correct establishment of the pneumaticity status in Archaeopteryx. The presence of an expanded pneumatization pattern in Archaeopteryx indicates facilitation of an active lifestyle7,12 and allows characterizing Archaeopteryx as a taxon that already had the metabolic prerequisites for this highly demanding active lifestyle and an avian-like, high-performance endothermy. The presence of a notarium-like stabilizing structure in the vertebral column in Archaeopteryx is fully in line with the new evidence on its expanded pneumaticity pattern, and adds another piece of evidence for the potential of this taxon for active, wing-driven flapping flight.
Methods
The main slab of the Berlin specimen of Archaeopteryx, MB.Av.101, was observed directly (by the naked eye) and with microscopy. The images taken by Helmut Tischlinger with the help of longwave UV light (365–366 nm) in combination with selective filter techniques15,21 were examined carefully and compared to the original specimen. The UV photographs are available at the MfN. Additionally, new photographs under longwave UV radiation were taken by M.K. to supplement the older images and compare both of them. Comparative photographs under visible light were provided by R.C.
A survey of other described specimens of Archaeopteryx has been done mainly by literature to get an idea of potentially preserved pneumatic structures. The Maxberg specimen of Archaeopteryx is lost and therefore could only be examined by photographs from Wellnhofer27. Data on the Thermopolis specimen of Archaeopteryx were taken from Mayr et al.25.
We assume that the first preserved vertebra visible in the head region represents the 2nd cervical because of its integration with the occiput; thus, the first vertebra with a thoracic rib is the 11th presacral vertebra (which has its rib slightly caudoventrally emplaced beneath the scapula), and so the first sacral vertebra (the rostralmost vertebra without a thoracic rib) is medial to the anterior iliac blade. We count therefore 22 presacral vertebrae in this specimen (i.e., 10 cervicals, and 12 rib-bearing thoracic vertebrae), one less than in the other known specimens of Archaeopteryx14,27.
We calculated a Pneumaticity Index22 (PI) as a quantitative measure to compare the amount of pneumatic elements throughout the postcranial skeleton with some extant birds. The PI is defined as the ratio between the number of pneumatic postcranial elements and the total number of postcranial elements22. Postcranial elements are subdivided into anatomical units (AU), yielding the equation of PI = #AU pneumatic/#AU total. The following AUs (see also Table S3) were defined for Archaeopteryx: Composite units = ANC, Anterior Cervical Vertebrae (1st to 3th cervical); MC, Middle Cervical Vertebrae (4th to 6th cervical); POC, Posterior Cervical Vertebrae (7th to 10th cervical); ANT, Anterior Thoracic Vertebra (1st to 6th thoracic); POT, Posterior Thoracic Vertebrae (7th to 12th thoracic); SS, Synsacral Vertebrae; CAA, Anterior Caudal Vertebrae; CAP, Posterior Caudal Vertebrae; TR, Thoracic Ribs; CAR, Caudal Ribs and Chevrons; PE, Pelvis (Ilium, Ischium, Pubis); DFL, distal forelimb elements (i.e., bones distal to elbow joints); DHL, distal hind limb elements (i.e., bones distal to knee joints); Individually scored units = CC, coracoids; FU, furculae; FM, femora; HU, humeri; SC, scapulae. In the Berlin specimen of Archaeopteryx, 7 (ANC, MC, POC, ANT, POT, CAA, TR) of the listed 18 AUs are unambiguously pneumatic, yielding a minimum PI of 7:18 = 0.39.
Data Availability
The original Berlin specimen of Archaeopteryx, MB.Av.101, is housed in the collection of fossil vertebrates at the Museum für Naturkunde in Berlin (MfN) and can be studied upon request at the institution. The authors declare that the data supporting the findings of this study are available within the paper and its supplementary information files.
References
O’Connor, M. P. Postcranial pneumaticity: An evaluation of soft-tissue influences on the postcranial skeleton and the reconstruction of pulmonary anatomy in archosaurs. J. Morphol. 267, 1199 (2006).
O’Connor, M. P. The postcranial axial skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Soc. Vertebr. Paleontol. Mem. 8(27), 127 (2007).
Duncker, H.-R. The lung air sac system of birds. Adv. Anat. Embryol. Cel. 45, 1 (1971).
Perry, S. F. Method for reconstructing the respiratory system of extinct animals. Sauropod dinosaurs as a case in point. Comp. Biochem. Physiol. A 143, 61 (2006).
Wedel, M. J. Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs. Paleobiology 29, 243 (2003).
O’Connor, M. P. & Claessens, L. P. A. M. Basic avian pulmonary design and flow-through ventilation in nonavian theropod dinosaurs. Nature 436, 253 (2005).
Benson, R. B. J., Butler, R. J., Carrano, M. T. & O’Connor, M. P. Air-filled postcranial bones in theropod dinosaurs: physiological implications and the ‘reptile’–bird transition. Biol. Rev. 87, 168 (2012).
B. B. Britt Thesis, University of Calgary (1993).
Wedel, M. J., Cifelli, R. I. & Sanders, R. K. Osteology, paleobiology, and relationships of the sauropod dinosaur. Sauroposeidon. Acta Palaeontol. Pol. 45, 343 (2000).
Wedel, M. J. The evolution of vertebral pneumaticity in sauropod dinosaurs. J. Vertebr. Paleontol. 23, 344 (2003).
Owen, R. On the Archaeopteryx of von Meyer, with a description of a new long tailed species from the lithographic stone of Solenhofen. Philosophical Transactions of the Royal Society of London 153, 33 (1864).
Kundrát, M., Nudds, M. J., Kear, B., Lü, J. & Ahlberg., P. The first specimen of Archaeopteryx from the Upper Jurassic Mörnsheim Formation of Germany. Historical. Biology 1, 3 (2018).
Britt, B. B., Makovicky, P. J., Gauthier, J. & Bonde, N. Postcranial pneumatization in Archaeopteryx. Nature 395, 374 (1998).
Christiansen, P. & Bonde, N. Axial and appendicular pneumaticity in Archaeopteryx. Proc. Roy. Soc. Lond., Series B 267, 2501 (2000).
Tischlinger, H. & Unwin, D. M. UV-Untersuchungen des Berliner Exemplars von Archaeopteryx lithographica H. v. MEYER 1961 und der isolierten Archaeopteryx-Feder. Archaeopteryx 22, 17 (2004).
Ostrom, J. H. Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society 8, 91 (1976).
Elzanowski, A. In Mesozoic Birds: Above the Heads of Dinosaurs, Chiappe, L. & Witmer, L. M., Eds (University of California Press), pp. 129–159 (2002).
Tischlinger, H. Neue Informationen zum Berliner Exemplar von Archaeopteryx lithographica H.v.Meyer 1861. New information regarding the Berlin example of Archaeopteryx lithographica H.v.Meyer 1861. Archaeopteryx 23, 33–50 (2005).
Hone, D. W. E., Tischlinger, H., Xu, X. & Zhang, F. The extent of the preserved feathers on the four-winged dinosaur Microraptor gui under ultraviolet light. PLoS One 5, 1 (2010).
Rauhut, O. W. M., Foth, C., Tischlinger, H. & Norell, M. A. Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany. PNAS 109, 11746–11751 (2012).
Tischlinger, H. & Arratia, G. Ultraviolet light as a tool for investigating Mesozoic fishes, with a focus on the ichthyofauna of the Solnhofen archipelago. in Mesozoic Fishes 5 – Global Diversity and Evolution, Arratia, G., Schultze, H.-P. & Wilson, H., Eds (Verlag Dr. Friedrich Pfeil, München), pp. 549–560 (2013).
O’Connor, M. P. Pulmonary pneumaticity in the postcranial skeleton of extant Aves: a case study examining Anseriformes. J. Morphol. 261, 141 (2004).
Martin, L. D. The beginning of the modern avian radiation. Documents des Laboratoires de. Geologie de la Faculte des Sciences de Lyon 99, 9–19 (1987).
O’Connor, M. P. Evolution of archosaurian body plans: Skeletal adaptations of an air-sac-based breathing apparatus in birds and other archosaurs. J. Exp. Zool. 311A, 629 (2009).
Mayr, G., Pohl, B., Hartman, S. & Peters, D. S. The tenth skeletal specimen of Archaeopteryx. Zool. J. Linn. Soc. 149, 97 (2007).
Rauhut, O. W. M., Foth, C. & Tischlinger, H. The oldest Archaeopteryx (Theropoda: Avialae): a new specimen from the Kimmeridgian/Tithonian boundary of Schamhaupten, Bavaria. PeerJ 6, e4191 (2018).
Wellnhofer, P. Archaeopteryx. Der Urvogel von Solnhofen. (Verlag Dr. Friedrich Pfeil, München), pp. 256 (2008).
Sereno, P. C. et al. Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina. PLoS One 3, e3303 (2008).
Brochu, C. A. Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and highresolution computed tomographic analysis of the skull. Soc. Vertebr. Paleontol. Mem. 7(22), 1 (2002).
Forster, C. A., Sampson, S. D., Chiappe, L. M. & Krause, D. W. The theropod ancestry of birds: New Evidence from the Late Cretaceous of Madagascar. Science 279, 1915 (1998).
Ewers, S. W., Rauhut, O. W. M., Milner, A. C., McFeeters, B. & Allain, R. A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco. PeerJ 3, e1323 (2015).
Watanabe, A. et al. Vertebral pneumaticity in the ornithomimosaur Archaeornithomimus (Dinosauria: Theropoda) revealed by computed tomography imaging and reappraisal of axial pneumaticity in Ornithomimosauria. PLoS One 10, e0145168 (2015).
Lü, J. et al. A new oviraptorid dinosaur (Dinosauria: Oviraptorosauria) from the Late Cretaceous of southern China and its paleobiological implications. Sci. Rep. 5, 11490 (2015).
Lü, J. et al. High diversity of the Ganzhou oviraptorid fauna increased by a new “cassowary-like” crested species. Sci. Rep. 7, 6393 (2017).
Lü, J. C. A new oviraptorosaurid (Theropoda: Oviraptorosauria) from the Late Cretaceous of southern China. J. Vertebr. Paleontol. 22, 871 (2002).
Norell, M. A. & Makovicky, P. J. Important features of the dromaeosaur skeleton. II. Information from newly collected specimens of Velociraptor. Am. Mus. Novit. 3282, 1 (1999).
Shen, C., Zhao, B., Gao, C.-L., Lü, J. & Kundrát, M. A new troodontid dinosaur, Liaoningvenator curriei gen. et sp. nov., from the Early Cretaceous Yixian Formation of western Liaoning Province, China. Acta Geosci. Sin. 38, 1–13 (2017).
Shen, C. et al. A new troodontid dinosaur from the Lower Cretaceous Yixian Formation of Liaoning Province. China. Acta Geol. Sin. 91, 763–780 (2017).
Osmólska, H., Currie, P. J. & Barsbold, R. In The Dinosauria. 2nd ed., Weishampel, D. B., Dodson, P. & Osmólska, H., Eds. (University of California Press, Berkeley), pp. 165–183 (2004).
Chiappe, L. M., Shu’an, J. & Qiang, J. Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of Northeastern China. Bull. Am. Mus. Nat. Hist. 242, 1 (1999).
Chiappe, L. M. & Walker, C. A. In Mesozoic Birds: Above the Heads of Dinosaurs, Chiappe, L. & Witmer, L. M., Eds (University of California Press), pp. 559–588 (2002).
Witmer, L. M. The craniofacial air sac system of mesozoic birds (Aves). Zoological Journal of the Linnean Society 100, 327 (1990).
Müller, B. The air sacs of the pigeon. Smithsonian Miscellaneous. Collections 50, 365 (1908).
Schwarz-Wings, D., Meyer, C. A., Frey, E., Manz-Steiner, H.-R. & Schumacher, R. Mechanics of pneumatic neck vertebrae in sauropod dinosaurs reveals a key role of vertebral pneumaticity for achieving enormous body sizes. Proc. Roy. Soc. Lond. B 277, 11 (2010).
Fajardo, R. J., Hernandez, E. & O’Connor, M. P. Postcranial skeletal pneumaticity: A case study in the use of quantitative microCT to assess vertebral structure in birds. J. Anat. 211, 138 (2007).
Erickson, G. M. et al. Was dinosaurian physiology inherited by birds? Reconciling slow growth in Archaeopteryx. PLoS One 4, e7390 (2009).
Baumel, J. J. & Witmer, L. M. In Handbook of Avian Anatomy: Nomina Anatomica Avium, Baumel, J. J., King, A. S., Breazile, J. E., Evans, H. E. & Vanden Berge, J. C., Eds (Nuttal Ornithological Club, Cambridge, Massachusetts), pp. 45–132 (1993).
Bellairs, A. D. A. & Jenkins, C. A. In Biology and Comparative Physiology of Birds, Marshall, A. J., Ed. (Academic Press, New York), vol. 1, pp. 241–300 (1960).
Hogg, D. A. Fusions occurring in the postcranial skeleton of the domestic fowl. J. Anat. 135, 501 (1982).
Storer, R. W. Fused thoracic vertebrae in birds: Their occurrence and possible significance. J. Yamashina Inst. Ornithol. 14, 86 (1982).
James, H. F. Repeated evolution of fused thoracic vertebrae in songbirds. The Auk 126, 862 (2009).
Kaiser, G. The inner bird: Anatomy and Evolution. (University of British Columbia) pp. 386 (2007).
Unwin, D. M. Pterosaurs from Deep Time. (Pi Press, New York 2006).
Witton, M. P. Pterosaurs: Natural History, Evolution, Anatomy. (Princeton University Press, Princeton, New Jersey), pp. 304 (2013).
Senter, P. Scapular orientation in theropods and basal birds, and the origin of flapping flight. Acta Palaeont. Pol. 51, 305 (2006).
Longrich, N. R., Vinther, J., Meng, Q., Li, Q. & Russell, A. P. Primitive wing feather arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Curr. Biol. 22, 2262 (2012).
Nudds, R. L. & Dyke, G. Narrow primary feather rachises in Confuciusornis and Archaeopteryx suggest poor flight ability. Science 328, 887 (2010).
Feo, T. J., Field, D. J. & Prum, R. O. Barb geometry of asymmetrical feathers reveals a transitional morphology in the evolution of avian flight. Proc. Roy. Soc. Lond. B 282, 20142864 (2015).
Cubo, J. & Casinos, A. Incidence and mechanical significance of pneumatization in the long bones of birds. Zool. J. Linn. Soc. Lond. 130, 499 (2000).
Currey, J. D. & Alexander, R. M. The thickness of the walls of tubular bones. J. Zool. Lond. 206, 453 (1985).
Acknowledgements
We thank Elke Siebert (MfN) for her help with scanning photographs, and the preparators Lutz Berner and Markus Brinkmann for their assistance in handling the Berlin Archaeopteryx. We are grateful to Peter J. Makovicky and two other anonymous referees for their valuable comments and hints on earlier versions of this manuscript. MK was supported by Scientific Grant Agency VEGA of the Ministry of the Education, Science, Research and Sport of the Slovak Republic (1/0853/17). GD acknowledges support from UEFISCDI Romania [grant number PN-III-P4-ID-PCE-2016-0572]. The publication of this article was funded by the Open Access Fund of the Leibniz Association.
Author information
Authors and Affiliations
Contributions
D.S. and M.K. designed and performed the research; D.S., M.K., H.T., G.D. and R.C. contributed analytical tools and comparative data and/or discussed the manuscript during progress; D.S. and M.K. analyzed data; D.S., M.K., G.D. and R.C. wrote the paper.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schwarz, D., Kundrát, M., Tischlinger, H. et al. Ultraviolet light illuminates the avian nature of the Berlin Archaeopteryx skeleton. Sci Rep 9, 6518 (2019). https://doi.org/10.1038/s41598-019-42823-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-42823-5
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
