
Abstract
Since the dawn of agriculture, crop yield has always been impaired through abiotic stresses. In a field trial across five locations worldwide, we tested three abiotic stresses, nitrogen deficiency, drought and salinity, using HEB-YIELD, a selected subset of the wild barley nested association mapping population HEB-25. We show that barley flowering time genes Ppd-H1, Sdw1, Vrn-H1 and Vrn-H3 exert pleiotropic effects on plant development and grain yield. Under field conditions, these effects are strongly influenced by environmental cues like day length and temperature. For example, in Al-Karak, Jordan, the day length-sensitive wild barley allele of Ppd-H1 was associated with an increase of grain yield by up to 30% compared to the insensitive elite barley allele. The observed yield increase is accompanied by pleiotropic effects of Ppd-H1 resulting in shorter life cycle, extended grain filling period and increased grain size. Our study indicates that the adequate timing of plant development is crucial to maximize yield formation under harsh environmental conditions. We provide evidence that wild barley alleles, introgressed into elite barley cultivars, can be utilized to support grain yield formation. The presented knowledge may be transferred to related crop species like wheat and rice securing the rising global food demand for cereals.
Similar content being viewed by others

Introduction
One of the major challenges that mankind faces is the ability to feed the ever-growing population, especially in the face of increased stresses due to climate change and reduced availability of arable land1,2. Different climate prediction models indicate severe effects for large parts of Africa, the Arabian Peninsula and Central South America3,4, where barley (Hordeum vulgare ssp. vulgare) still has an crucial role as human food5. Barley is mainly used for animal feed and for malt production in large parts of the world. It represents the fourth most important cereal crop on a global scale5,6.
Barley inherently exhibits a higher level of abiotic stress tolerance than other crops7,8,9, which offers the possibility to extend its future production to areas suffering from climate change. Furthermore, the relatively simple diploid genetics of barley and the tight relationship between the members of the Triticeae tribe facilitate the transfer of knowledge gained from barley research to other major cereals, for instance, bread wheat, durum wheat and rye10. Wild barley (Hordeum vulgare ssp. spontaneum), originating from the Fertile Crescent and from a second area some 1,500–3,000 km farther east, was used to domesticate modern elite barley (Hordeum vulgare ssp. vulgare) more than 10,000 years ago11,12,13. The usefulness of wild germplasm for future breeding has often been emphasized14,15,16, mostly as a source to improve biotic resistance and abiotic stress tolerance rather than to directly increase grain yield17. Recent studies in wild barley indicate the existence of vast phenological variation for important agronomic traits18,19,20,21,22,23,24,25. Wild barely may thus be an appropriate source to replenish the barley gene pool with novel genetic variation. This variation may be valuable to cope with the challenges arising from climate change26.
Grain yield depends on developmental phases of a plant’s life cycle27. In this regard, flowering time is a key event as plants shift from vegetative to reproductive growth, moving towards providing the harvestable yield28,29,30. The optimal timing of this event is crucial as it should occur in the absence of adverse effects like abiotic stresses31 but also ensuring completion of yield accumulation without encountering further adverse effects in most growing seasons. Therefore, the targeted timing of this phase provides one approach to improve stress tolerance, through stress avoidance, and thus to increase grain yield32. Flowering time is mainly controlled by environmental cues like day length (photoperiod) and temperature (especially the exposure to cold temperatures, also termed vernalization)33,34,35. Flowering time is highly heritable and, so far, several major genes controlling flowering time have been discovered in model species and in crop plants36. Generally, flowering time genes are classified into at least three families: [I] photoperiod genes (e.g. Ppd-H1)37, [II] vernalization genes (e.g. Vrn-H1, Vrn-H2 and Vrn-H3)33,38 and [III] earliness per se (eps) genes, the last controlling flowering independently from photoperiod and temperature (e.g. Sdw1)39,40.
Here, we present data of a large field study with the HEB-YIELD population, a selected subset of the wild barley nested association mapping (NAM) population HEB-2518. The aim of the study was to examine the interplay between flowering time, stress tolerance and yield. For this purpose, HEB-YIELD was studied at five locations worldwide and during two years under locally relevant abiotic stress conditions. We investigated the role of known flowering time genes on developmental and yield-related traits, as well as how they account for yield and stress tolerance.
Results and Discussion
HEB-YIELD exhibits strong phenotypic variation as well as environmental and treatment variation
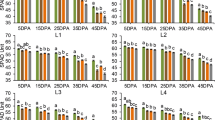
The wild barley introgression population HEB-YIELD comprises a diverse subset of lines selected from the NAM population HEB-2518 (Supplementary Table S1a). We studied eleven agronomically traits in a HEB-YIELD trial conducted in Dundee, Halle, Al-Karak, Dubai and Adelaide (Fig. 1; Supplementary Table S2a), where climate data for day length, temperature and precipitation varied considerably between locations Supplementary Figs S1 and S2). The parameters studied included developmental and yield-related traits, used to capture growth variation among HEB-YIELD lines (Supplementary Table S3). At each location, the traits were measured under site-specific abiotic stress conditions, i.e. nitrogen deficiency in Dundee and Halle, drought stress in Al-Karak and Adelaide and salt stress in Dubai. In total, 3,207 field plots were evaluated over all sites, seasons, treatments, and replicates (Supplementary Tables S2c and S4a). Considerable phenotypic variation within locations and treatments was observed for all investigated traits (Fig. 2; Supplementary Table S5).

Global macroclimate map with information on the five experimental locations. The position of the five (1–5) test locations are indicated on a simplified map of the Köppen-Geiger climate classification system provided by LordToran “Clickable world map with climate classification”, https://en.wikipedia.org/wiki/World_map#/media/File:K%C3%B6ppen-vereinfacht.svg, copyright: CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0/). General information about the test locations are given in the table on the lower left-hand side including the nearest town, country, stress treatment and the years of field trials. Insets next to map positions depicts long-term climate information for each test location. The average monthly precipitation in millimeters (blue bars), the average monthly temperature in degrees Celsius (red line) and the course of the day length during the year in hours (yellow line) are displayed. In addition, the sowing and harvesting dates are indicated with empty and filled circles, respectively. The Adelaide inset on the right-hand side serves as a legend for the insets.

Box-Whisker plots illustrating HEB-YIELD trait variation per location and treatment. Trait names and trait units are indicated in the grey rectangle above each subplot. Trait abbreviations are listed in Supplementary Table S3. The locations Dundee (DUN), Halle (HAL), Al-Karak (ALK), Dubai (DUB) and Adelaide (ADE) are indicated with blue, grey, green, red and yellow box-whiskers, respectively, and, in addition, at the bottom of the plot. Empty and filled boxes refer to control and stress treatments, respectively. Significant differences between treatments are indicated with red asterisks above boxes with *P < 0.05, **P < 0.01 and ***P < 0.001. The relative increase/decrease (in %) of the stress treatment compared to the control treatment is given below the asterisks.
The ANOVA revealed that all investigated factors (genotype, year and location) were significant for all traits except plant height (HEI) where the year effect was not significant (Supplementary Table S6a). Interestingly, only the traits shoot elongation phase (SEL) and HEI showed comparable values across locations, whereas for the majority of traits pronounced location effects were observed (Supplementary Table S6c). For instance, flowering time varied from 57 to 144 days and grain yield from 0.14 dt/ha to 74 dt/ha (Supplementary Table S5), reflecting a strong diversity in yield potential among the trial sites (Fig. 2).
Irrespective of the diverging agricultural practices at the trial sites, developmental trait heritabilities were high with an average of 0.87, ranging from 0.10 (ripening phase (RIP) under control treatment in Al-Karak) to 0.99 (shooting (SHO) under control treatment in Adelaide as well as flowering (HEA) and SEL under both treatments in Dubai, Supplementary Table S5). In general, yield-related traits revealed lower heritabilities with an average of 0.65. The most complex trait, grain yield (YLD), revealed average heritabilities of 0.73, ranging from 0.05 (YLD under stress treatment in Dubai) to 0.93 (YLD under control treatment in Dundee).
Trait performance in HEB-YIELD is usually a linear transformation from control to stress treatments indicating low genotype by treatment interaction
To gain insights into how abiotic stresses may affect plant development and grain yield, we cultivated HEB-YIELD under contrasting stress conditions, which are relevant for the respective test locations (Supplementary Table S2c). The applied stresses exhibited only minor effects on plant development traits except for HEI. In contrast, strong effects on all measured yield-related traits were observed at all test locations, for instance, reducing yield under stress between 16% in Halle and 65% in Adelaide (Fig. 2; Supplementary Table S5). We observed a weak trend, that HEB-YIELD lines under drought and salt stress exhibited an accelerated plant development, presumably to escape the stress condition, which is in agreement with other studies in cereals32,41. Based on our findings we suggest that plant development in the wild barley population HEB-YIELD is mainly determined by genetic factors and to a lesser extend modified by abiotic stresses. This is further supported by the observation that plant developmental traits showed a nearly linear shift between control and stress conditions, as indicated by high correlation coefficients (0.99 > r > 0.59) between stress and control treatments of developmental traits, except for SEL in Adelaide (r = 0.12; Table 1; Supplementary Table S7a).
Grain yield correlations indicate that yield formation depends on a location-specific interplay between developmental traits and yield components
We observed Pearson correlations coefficients between plant developmental stages shooting, flowering and maturity ranging from r = 0.67 to r = 0.96 (apart from shooting correlations in Dubai Supplementary Tables S6b and S7b), indicating a nearly colinear regulation of plant developmental phases. Thus, HEB-YIELD lines early or late in shooting have the tendency to stay early or late respectively until maturity. This observation is in agreement with previous findings in the wild barley NAM population HEB-25, studied in Halle20 and Dundee25. Consequently, early developmental stages may be used as an indirect criterion to select HEB-YIELD lines for early or late maturity.
Following these findings, we explored the relationship between plant development and yield formation in HEB-YIELD (Table 2). We observed a trend that late plant development is beneficial for increased grain yield under Dundee, Halle and Adelaide growth conditions, indicated by positive correlation coefficients of r(HEAxYLD) = 0.59/0.66, 0.32/0.20 and 0.57/0.51, respectively, under control/stress treatments. This trend fits the general observation that late lines have the potential to exploit a prolonged growing season if the environmental conditions including temperature and precipitation are beneficial29,42,43. In contrast, under the harsh environmental conditions at Al-Karak and Dubai, HEB-YIELD lines with accelerated plant development were favored. Consequently, we observed negative correlations between flowering and grain yield at Al-Karak and Dubai with r(HEAxYLD) = −0.30/−0.72 and −0.51/−0.44, respectively, under control/stress treatments (Table 2). Here, elevated temperatures and low rainfall restricted plant growth to a few months and thus earliness is a major breeding goal44,45. In future, this situation may intensify, since climate change is expected to further shorten the growing period in drought and heat prone locations like in Jordan26,32,46.
We also observed strong location-specific correlations between flowering time and yield components. For example, in Halle and Dundee, flowering time was positively correlated with grain number per ear (GNE), with r(HEAxGNE) = 0.67/0.67 and 0.71/0.64, respectively (Table 2). Here the extended vegetative growth phase allowed more spikelet primordia to be maintained. In contrast, in Al-Karak flowering time negatively affected thousand grain weight (TGW) with r(HEAxTGW) = −0.56/−0.68, reflecting a grain filling penalty for later flowering genotypes. These findings suggest that flowering time controls final grain yield to a certain degree. By comparing correlations between grain yield and yield components, we observed that apparently GNE is the key determinant of grain yield in HEB-YIELD – irrespective of location and treatment. This result is in agreement with earlier studies47,48, indicating that any increase in number of grains may also improve grain yield49,50. We thus reason that improving GNE may offer the best route to increase grain yield in HEB-YIELD independent of the environmental conditions.
The highest positive correlations of yield were found with harvest index (HI; scored only in Dubai and Adelaide with r(YLDxHI) = 0.87 and 0.83, respectively, Table 2). A previous study noted the importance of increasing harvest index to improve yield during the past century51. However, a further improvement of grain yield through raising harvest index may be a dead end, since barley is supposed to have reached an optimum with a harvest index of approximately 0.6251,52. Therefore, future grain yield improvements may be achieved through increasing plant biomass51,53. This suggestion is in accordance with our finding that grain yield exhibited a slightly positive correlation with shoot elongation phase in those environments where lateness was beneficial to increase yield (Table 2). During shoot elongation, which captures the growth period between establishing awn primordia and ear emergence, the leaf growth rate and the potential grain number per area are defined47,49,54. An extended shoot elongation phase may thus improve grain yield by increasing leaf size, i.e. biomass, and grain number per area. On the other hand, ripening phase under drought stress exhibited positive and negative correlations with grain yield in Al-Karak and in Adelaide, respectively. Whereas the Adelaide finding fits the assumption that early maturity and thus a short ripening phase may improve grain yield under terminal drought, the Al-Karak finding is unexpected. Under drought stress conditions in Al-Karak, an extended ripening phase was associated with an increase in grain weight, ultimately resulting in elevated grain yields. We conclude that fine-tuning of plant development, especially their sub-phases, may contribute to a better adaptation of improved varieties to their target environment. The latter notion is supported by the finding that in the first instance climate change is expected to impair flowering time32, which is crucial for plant adaptation and yield formation49,55. In addition, our stress treatments confirmed the known association between inflorescence development and stress tolerance/avoidance32,56. This offers the possibility to use the genetically relatively well-understood trait flowering time as a proxy to select for improved grain yield under abiotic stresses57.
Flowering time genes exhibit pleiotropic effects on yield formation in HEB-YIELD
In order to explore the interplay between flowering time regulation and yield formation, we investigated the effects of four major flowering time genes, Ppd-H1, Sdw1, Vrn-H1 and Vrn-H3, in HEB-YIELD (Supplementary Table S8a). The relevance of these candidate genes has been reported in various studies34,36, including the wild barley NAM population HEB-2518,20,22,25. The wild barley lines of HEB-YIELD were selected to compare the effects of wild and cultivated alleles at these four flowering time loci (Supplementary Table S1a). In the following, we report on the pleiotropic effects associated with the four flowering genes studied.
Ppd-H1
Flowering under long days is promoted by the photoperiod responsive Ppd-H1 (PHOTOPERIOD-H1) allele, an orthologue of the Arabidopsis pseudoresponse regulator gene PRR7, which is present in wild barley and winter barley cultivars37. In contrast, spring barley cultivars like Barke possess the recessive non-responsive ppd-H1 allele, resulting in late flowering. Allelic variation in Ppd-H1 follows a latitudinal cline, where a large proportion of the dominant, long day responsive allele is present in cultivars from Southern Europe and the recessive, non-responsive allele, predominates in Northern Europe58. Selection of the non-responsive alleles found in elite spring barley allowed barley cultivation in northern latitudes, under more humid and colder conditions59. During early plant development, the photoperiod signal is transmitted from the circadian clock oscillator Ppd-H1 through mediation of the CONSTANS (CO) protein to the floral inducer Vrn-H3, an orthologue of the Arabidopsis FLOWERING LOCUS T (FT) gene.
In order to locate sequence variants in Ppd-H1 discriminating between wild and cultivated barley in the set of HEB-YIELD parents, we used exome capture sequence data (Supplementary Table S1b). The data set comprised nine spring barley cultivars (including Barke), six winter barley cultivars and 25 wild barley accessions (H. v. ssp. spontaneum and H. v. ssp. agriocrithon) including 19 wild barley donors present in HEB-YIELD. We found 107 sequence variants in Ppd-H1 (Supplementary Table S1b). Fourteen sequence variants in Ppd-H1 are notable. Variant 97 at chr2H_29.127.381 bp discriminated between all tested wild and spring barleys. This variant is identical with SNP 22 cited in Turner et al.37. SNP 22 separates winter and wild barley from spring barley and is located in the CCT domain of Ppd-H1, where the G to T substitution causes a glycine to tryptophan change resulting in a photoperiod non-responsive mutant phenotype37. In addition, 13 Ppd-H1 variants (15, 27, 30, 37, 39, 42, 51, 62, 64, 88, 99, 112, 116 46) discriminated between the tested wild barleys and all tested spring barleys except HEB-25 donor HID138, originating from Iran but not present in HEB-YIELD18. The variants mentioned are located in three introns and five exons and potentially indicate a different functional response of the Ppd-H1 allele compared to the donor alleles present in HEB-YIELD.
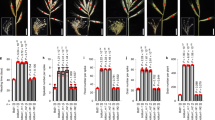
Among the candidate genes, Ppd-H1 revealed the most pronounced effects on plant development in Dundee, Halle and Al-Karak (Fig. 3; Supplementary Table S8a). This finding is in accordance with several other studies conducted in barley20,23,25,60,61. At these locations, the wild allele of Ppd-H1 accelerated plant development in HEB-YIELD (SHO, HEA and maturity (MAT)) with a maximum effect of −9.0 days in Halle. In contrast, no significant effect of the Ppd-H1 wild allele was observed in Dubai and Adelaide. Most wild barley accessions carry the dominant allele, which is responsive to a long day photoperiod, accelerating plant development through upregulating of Vrn-H3/HvFT162,63. One possible explanation for contrasting effects between locations is the different day lengths at these sites. Dundee and Halle are more than 5,700 km distant from the equator and are clearly exposed to long day conditions indicated by average day lengths of more than 15 hours during shooting phase, which is necessary to trigger the effect of Ppd-H137. In Al-Karak, with a day length of appr. 12 hours during shooting phase, we still observed strong Ppd-H1 effects, although this location is more than 2,000 km closer to the equator than Halle. In Dubai, where day length is shorter with less than 11 hours during shooting phase, only one minor Ppd-H1 effect on plant development and yield formation was observed (Supplementary Table S8b). Apparently, this is because Ppd-H1 is only active under long-day condition37. Presumably, the short-day signal to initiate flower development in Dubai is transmitted through Ppd-H2, the FLOWERING LOCUS T3 (FT3) gene64. The dominant functional allele of Ppd-H2 promotes spikelet formation under short-day in spring barley and winter barley originating from Southern Europe whereas the recessive non-functional allele carries a large deletion in the transcribed coding region, which is typically present in Northern European winter barley cultivars65,66,67. Based on exome capture sequence data, we found nine sequence variants in Ppd-H2 (Supplementary Table S1b). No sequence variant could be identified, which perfectly discriminated between cultivated and wild barley. However, Ppd-H2 variant 3 at chr1H_514.098.364 bp and variant 7 at chr1H_514.098.702 bp discriminated between the tested winter barleys and the tested spring barleys except Morex. The wild barley HEB-YIELD donors showed SNPs in common but not restricted to spring or winter alleles.

Estimates of Ppd-H1 wild allele effects on plant developmental and yield-related traits. The trait names are given in the grey rectangles above each subplot and at the bottom where, in addition, the units of the traits are indicated. Trait abbreviations are listed in Supplementary Table S3. The color of the bars represents the location, blue for Dundee, grey for Halle, green for Al-Karak, red for Dubai and yellow for Adelaide. Ppd-H1 wild allele effects under control and stress treatments are depicted with a bright blue (top) and a bright red background (bottom), respectively. Statistically significant wild allele effects are indicated by red asterisks above or below the bars with *P < 0.05, **P < 0.01, ***P < 0.001. The height of the bars indicates the size of the Ppd-H1 wild allele effect, obtained by calculating the difference between the mean performances of HEB-YIELD lines carrying two wild alleles versus two elite alleles.
In future, follow-up field studies with double HIFs (heterogeneous inbred families) may assist to further characterize the interplay between the Ppd-H1 and Ppd-H2 photoperiod receptors under short-day and long-day conditions68. For this, four HEB-25 lines, HEB_05_044, HEB_08_149, HEB_16_063 or HEB_22_039, which were simultaneously heterozygous at both loci in generation BC1S3, can be chosen based on Maurer et al.18. Subsequently, four homozygous allele combinations at the two loci can be selected in the available HEB-25 selfing generation BC1S3:11. After seed multiplication, epistatic effects on flowering time and yield formation can be tested in replicated field trials using the resulting four nearly isogenic double HIFs.
In HEB-YIELD, Ppd-H1 acted in a location-specific manner on yield-related traits. The most pronounced Ppd-H1 effect was present in Al-Karak where the day length-sensitive wild barley allele was associated with an increase of grain yield by 4.1 dt/ha (+15%) and 5.3 dt/ha (+30%) under control and drought stress conditions, respectively (Fig. 3; Supplementary Table S8a). The yield effect may be explained through pleiotropic effects of the wild barley Ppd-H1 allele, which shortened the overall growing season, increased the period of grain filling (RIP) and increased grain size (TGW). A tendency of the Ppd-H1 wild barley allele towards enhanced grain yields was also observed in Dubai and Adelaide, however, only significant in Adelaide under drought stress (+4.8 dt/ha = +29%). Usually, the location-specific effects of Ppd-H1 on yield-related traits are in agreement with the preferred length of the growing period. At those locations where earliness is beneficial, the responsive wild allele of Ppd-H1 exerted increasing effects on yield-related traits, for example in Al-Karak, where early plants escaped higher temperatures and terminal drought at the end of the growing season. On the other hand, where lateness is preferable to achieve higher yields, the elite barley ppd-H1 allele increased yield-related traits, for example, in Dundee and Halle. At those locations late HEB-YIELD lines benefited from the extended growing period since the environmental conditions supported plant growth under suitable conditions.
Sdw1
Sdw1 belongs to the group of so-called semi-dwarfing genes40, which are responsible for yield elevations during the ‘Green Revolution’69. Wild barley accessions possess the functional and dominant Sdw1 allele, a gibberellic acid 20 oxidase (GA20ox) gene, which promotes plant growth. In contrast, the recessive, GA-deficient sdw1 allele70,71 is present in barley cultivars like Barke, causing a semi-dwarf phenotype. Several studies have shown that semi-dwarfs exhibit reduced plant height, late maturity, increased tiller numbers and an improved harvest index, ultimately resulting in elevated grain yields40,72,73. Based on exome capture sequence data, we found 46 sequence variants in Sdw1 (Supplementary Table S1b). No Sdw1 sequence variant could be identified to completely discriminate cultivated and wild barley. However, variant 130 at chr3H_634.078.282 bp discriminated all tested cultivated spring and winter barleys from the tested wild barleys except of the three HEB-25 donors HID003, HID099 and HID114, originating from Iraq, Syria and Lebanon, respectively18. It is, thus, possible that new GA20ox mutations are present in wild barley.
The reported pleiotropic effects of Sdw1 are also supported by HEB-YIELD field data (Fig. 4; Supplementary Table S8a). Throughout plant development, we detected an accelerating effect of the wild barley Sdw1 allele in HEB-YIELD, accelerating grain maturity by 4.0 to 8.9 days in Dundee, Al-Karak, Dubai and Adelaide, compared to the semi-dwarfing allele of Barke. Most striking was the pronounced delay of development in Adelaide under the control condition (precipitation = 484 mm), with up to 13 days for SHO. Whereas under stress (precipitation = 159 mm) the effects were on a similar level as in the other locations. The Adelaide effect might be explained by different environmental cues between the two years, resulting from the earlier sowing date and the prolonged growing period of 50 days in 2016. So far, there is no evidence that day length or precipitation affects the function of Sdw140,71. However, a wheat survey under controlled conditions already reported that temperature can modify GA dependent responses, where elevated temperatures increase the abundance of GA74.

Estimates of Sdw1 wild allele effects on plant developmental and yield-related traits. The trait names are given in the grey rectangles above each subplot and at the bottom where, in addition, the units of the traits are indicated. Trait abbreviations are listed in Supplementary Table S3. The color of the bars represents the location, blue for Dundee, grey for Halle, green for Al-Karak, red for Dubai and yellow for Adelaide. Sdw1 wild allele effects under control and stress treatments are depicted with a bright blue (top) and a bright red background (bottom), respectively. Statistically significant wild allele effects are indicated by red asterisks above or below the bars with *P < 0.05, **P < 0.01 or ***P < 0.001. The height of the bars indicates the size of the Sdw1 wild allele effect, obtained by calculating the difference between the mean performances of HEB-YIELD lines carrying two wild alleles versus two elite alleles.
The most prominent effect of semi-dwarfing genes is their control of plant architecture, in particular, plant height40,75. We confirmed this effect in HEB-YIELD since the wild barley allele increased plant height at all locations and under both treatments with a maximum increase of 33.2 cm in Dundee under control condition. The dominance of semi-dwarf genes in modern crop cultivars indicates their global importance for agriculture69,76.
This notion is also confirmed in HEB-YIELD where the Barke semi-dwarf allele was associated with an increase in grain yield (Fig. 4; Supplementary Table S8a). In turn, the wild barley allele significantly reduced grain yield, for instance under control conditions in Dundee, Halle and Adelaide by up to 15.8 dt/ha. Under drought stress conditions in Adelaide, the Barke semi-dwarf allele revealed the strongest impact, accounting for 60% of the final yield level. The observed yield increase may be attributed to an accumulation of several positive effects, including an extended growing period, more tillers, a higher harvest index and less lodging and head loss (Fig. 4; Supplementary Table S8a).
Vrn-H1
In addition to the aforementioned photoperiod and GA dependent pathways, flowering time is also regulated through the vernalization pathway, where exposure to cold temperatures accelerates flowering33,77,78. In barley, the response to cold temperatures is mainly controlled by interaction of the two vernalization genes, Vrn-H179 and Vrn-H280. Vrn-H2 acts as a strong repressor of flowering under long day conditions, preventing winter barley cultivars and wild barley accessions to flower during winter80. The expression of the APETALA1 MADS-box gene Vrn-H1 is only induced after extended periods of cold exposure81, resulting in down-regulation of Vrn-H2 and induction of flower initiation through direct binding of the Vrn-H1 protein to the promoters of Vrn-H2 (repression) and Vrn-H3 (activation)82. In spring barley cultivars like Barke, the dominant Vrn-H1 allele promotes flowering whereas the recessive winter barley and wild barley alleles delay flowering if cold exposure is imperfect.
Based on exome capture sequence data, we found a huge number of 377 sequence variants in Vrn-H1 (Supplementary Table S1b). A number of Vrn-H1 sequence variants discriminated between cultivated and wild barley. For example, variants 475 and 499 at chr5H_599.131.041 bp and chr5H_599.131.479 bp, respectively, discriminated the tested spring barleys from the tested wild and winter barleys. Interestingly, we found one variant, 443 at chr5H_599.130.360 bp, which discriminated the tested spring and winter barleys from the tested wild barleys. In addition, ten Vrn-H1 variants (187, 194, 289, 296, 335, 340, 388, 452, 457, 498), discriminated the tested winter barleys from the tested spring and wild barleys. The named variants are exclusively located in introns between exon 1 and exon 4. It was already known that deletions in the first intron of Vrn-H1 result in spring type cultivars, lacking the vernalization need to initiate flowering82. Our findings indicate that additional intron regulatory elements may be present in Vrn-H1 to differentiate winter barley, spring barley and wild barley.
In future, follow-up field studies using nearly isogenic HIFs may assist to characterize developmental and yield formation effects of individual Vrn-H1 variants, which are present in HEB-YIELD lines68. Likewise, the epistatic interaction between selected alleles of Vrn-H1 and Vrn-H2 may be characterized in double HIFs, which can be developed from any of five double heterozygous HEB-25 lines, HEB_09_101, HEB_16_095, HEB_16_099, HEB_23_061 or HEB_24_066, as mentioned before.
In HEB-YIELD, Vrn-H1 exhibited considerable effects on nearly every trait in Dubai and, to a lesser extent, in Adelaide (Supplementary Fig. S3; Supplementary Table S8a). HEB-YIELD lines carrying the wild barley allele at this locus delayed flowering time and maturity by more than 10 days in Dubai. In Adelaide, pronounced effects on plant development were restricted to the control condition (i.e. the Adelaide growing period 2016). Most likely, this effect is caused by warmer temperatures and therefore less vernalization stimuli at the beginning of the growing season (Supplementary Figs S1 and S2; Supplementary Table S9). However, the late development effect of the wild barley Vrn-H1 allele in Adelaide diminished during cultivation from +12 days at shooting, +6 days at flowering to, finally, +4 days at maturity. This tendency was also present in Halle and Al-Karak, although on a much lower level. In contrast, the late development effect of the wild barley Vrn-H1 allele remained stable throughout plant cultivation in Dubai. This may be because the temperature in Dubai never reached a vernalization-triggering level. In Dubai, the HEB-YIELD lines possessing a wild barley winter allele at Vrn-H1 thus responded to the lack of vernalization with a late plant development.
In addition to its developmental effects the wild barley allele of Vrn-H1 exerted significant reducing effects on all yield components of around 25% in Dubai. Consequently, the final grain yield in Dubai was reduced by 3.2 dt/ha under control conditions, which corresponds to 37% of the total yield. At locations where earliness is the preferred breeding goal and vernalizing conditions are rare, the use of the dominant elite barley allele of Vrn-H1 is highly recommended.
Vrn-H3
As mentioned before, the expression level of Vrn-H1 increases with exposure to cold temperatures, resulting in flower induction through repression of Vrn-H2 and activation of Vrn-H329,82. Vrn-H3 corresponds to the HvFT1 gene, which is an orthologue of the Arabidopsis FT gene, the so called ‘florigen’33,83,84,85 Vrn-H3 plays a central role in flower induction integrating photoperiod and vernalization signals84. Barley alleles of Vrn-H3 vary regarding the first intron sequence, promoter sequence and copy number. They are widely distributed over winter and spring growth habits84,85,86. Unfortunately, we could not identify Vrn-H3 variants in our exome capture sequence data. Presumably, Vrn-H3 produced no variants since all reads were identified as multi-mappers, located at two or more genomic regions simultaneously, and, hence, were ignored by the variant caller. (M. Bayer, personal comm.).
HEB-YIELD field data validated the role of Vrn-H3 on plant development throughout the growing period in all locations except from Dubai (Supplementary Fig. S4; Supplementary Table S8a). The wild barley allele of Vrn-H3 slowed down plant development between 2.2 and 6.6 days. Generally, winter genotypes are characterized by carrying a recessive Vrn-H3 allele, which displays a reduced expression85. Most wild barleys possess a winter type62 and probably harbor a recessive vrn-H3 allele, which explains the decelerating developmental effects.
Although Vrn-H3 plays an important role for plant development, we identified only weak, mostly non-significant, impacts on yield-related traits. Only in Al-Karak, the wild allele showed significant reducing effects on grain number per ears (under both treatments) and on grain yield under drought stress (−3.3 dt/ha).
The best wild barley HEB-YIELD lines match the yield performance of high-yielding local check cultivars
The usefulness of wild accessions, related to crop species has been proposed and demonstrated frequently14,15,87. Wild barley accessions, in particular H. v. ssp. spontaneum, the progenitor of cultivated barley have been used to improve disease resistance24,88 and abiotic stress tolerance22,73,89,90, as well as plant developmental traits18,20,25 and quality traits88,91,92. The successful use of wild relatives to increase grain yield of barley has not been reported frequently, some exceptions are available73,93,94. This may be because of the negative impacts of linked deleterious wild alleles, a phenomenon generally referred to as ‘linkage drag’95. The HEB-YIELD lines offer the possibility to estimate potentially positive wild allele effects in an adapted genetic background, since they are embedded through backcrossing into the modern elite barley cultivar Barke. In addition, the elite genetic background enables the direct use of HEB-YIELD lines in barley breeding programs.
Based on our two-year field trials, we identified five high yielding HEB-YIELD lines, which showed acceptable grain yield performance, comparable to the recurrent elite parent Barke, across the tested locations. These HEB-YIELD lines are 01_132, 01_104, 10_184, 10_173 and 05_043 (Supplementary Fig. S5; Supplementary Tables S10a–c). They possessed higher grain yields than Barke in Al-Karak (except HEB_10_184). In addition, HEB-YIELD lines 01_132 and 10_184 surpassed the Barke grain yield in Dundee under both stress and control treatments. Furthermore, we identified HEB-YIELD lines, which reached or surpassed the yield level of locally adapted check cultivars (Supplementary Fig. S5; Supplementary Tables S10a–c). These HEB lines are 10_184 and 01_132 in Dundee (Supplementary Fig. S6), 01_132 and 01_104 in Halle (Supplementary Fig. S7), 05_043 and 10_173 in Al-Karak (Fig. 5), 15_082 and 06_116 in Dubai (Supplementary Fig. S8) and 10_184 & 01_132 in Adelaide (Supplementary Fig. S9). For instance, HEB_01_132 surpassed the grain yield of the established local check cultivar ‘Navigator’ under stress treatment in Adelaide. In addition, under both treatments it was comparable to ‘Compass’ and ‘La Trobe’, which have become the dominant commercial cultivars in South Australia. HEB_01_132 also surpassed the grain yield of the local check ‘58/1 A’ under control treatment in Dubai, indicating that this line may be directly suited for cultivation in the respective environments. Likewise, HEB_05_043 and HEB_10_173 outperformed the check cultivar ‘Rum’ in Al-Karak under drought stress.

Regression of grain yield on flowering in Al-Karak. The yield levels of the 48 HEB-YIELD lines plus checks are depicted as a function of flowering time, separately for control (blue labels) and stress (red labels) treatments. The yield level of the local check cultivar ‘Rum’ is indicated by a dashed red line. On top of each subplot the linear regression equation, the Pearson’s correlation coefficient (r) and the coefficient of determination (r2) are indicated.
Interestingly, HEB-YIELD lines adapted different yield formation strategies at each test location. Compared to the local check cultivars, HEB-YIELD lines had increased numbers of ears (EAR) at Al-Karak, increased thousand grain weights at Dubai in almost all cases, and increased grain numbers per ear under stress at Dundee and Adelaide in many cases (Supplementary Figs S10–S12). This offers the possibility of achieving future yield improvements following a location-specific adaptation route.
The challenges of climate change demand that cultivars need to re-adapt to changing environmental conditions, for instance shorter growing seasons, higher average temperatures during cultivation and more frequently occurring drought periods26,46,96. HEB-YIELD lines exhibited a high phenological variation. For instance, flowering time exhibited a range of 49 days in Dubai and 36 days in Adelaide (Supplementary Table S5), which offers the potential to use this variation to adapt new cultivars to changing environmental conditions by backcrossing favorable HEB-YIELD donor lines with locally adapted elite cultivars. In those areas where drought and heat affect plant development and grain maturation, early maturing lines like HEB_05_043, HEB_15_082 or HEB_10_173 may be beneficial because of their fast development (Fig. 5; Supplementary Fig. S9; Supplementary Table S10a). Moreover, the increased tiller capacity of HEB-YIELD lines may be promising to achieve an improved canopy cover, reducing moisture losses26,45 and to increase biomass yield. The latter trait may be high value in the eastern part of the Mediterranean basin where straw and grains of barley are mainly used for animal feeding97,98.
In the past, Australian varieties followed a strong focus on earliness but changes in agricultural practices have resulted in earlier sowing dates and thus an extended growing season. The earlier sowing allowed later genotypes to benefit from a longer growing period, enabling HEB-YIELD lines HEB_03_085, HEB_10_184, 20_064 and HEB_04_135 to surpass the grain yield of the local check cultivar ‘Navigator’ in Adelaide under control condition (Supplementary Fig. S9; Supplementary Table S10a). We also identified HEB-YIELD lines that performed quite well in the high yielding environments of Dundee and Halle. Here, lines HEB_01_104, HEB_01_132 and HEB_10_184 accomplished reasonable yields. HEB_10_184, for instance, achieved a maximum grain yield of 74.0 dt/ha under control condition in Dundee, which was almost on par with the local check cultivar ‘Odyssey’ (−0.3%) and 5.1% higher than the recipient cultivar Barke. Our findings indicate that wild barley HEB-YIELD lines can be used as pre-breeding material to further improve plant development and yield formation of elite barley. (Supplementary Figs S1 and S2; Supplementary Tables S9a and S10a).
Conclusion
It is expected that the impact of climate change necessitates the adaptation of our established crop cultivation systems to harsher environmental conditions26,96. Stress avoidance is one promising approach to increase stress tolerance. We explored this relationship by studying the wild barley-derived model population HEB-YIELD in a field experiment, ranging from Dundee in Scotland to Adelaide in South Australia, where the effects of nitrogen deficiency, drought and salinity on plant development and yield-related traits were investigated.
Our findings confirm the crucial relationship between flowering time, plant development and grain yield99. The exact timing of the switch from vegetative to reproductive growth under favorable conditions32, the length of the growing period and the duration of the sub-phases of plant development are crucial to secure yield under abiotic stress conditions. We suggest that adjusting plant development may be a promising breeding strategy to cope with abiotic stresses. To optimize breeding programs, it is thus advisable to first predict the environment-dependent impact of flowering time genes on yield formation and then to select locally advantageous alleles for sustainable crop improvement.
Our HEB-YIELD data indicate that wild germplasm may serve as a resource to increase genetic diversity14,20,22 and to enable the above mentioned adaptation to abiotic stresses, through selection of early or late development alleles of known major flowering time genes, e.g. Ppd-H1, Sdw1, Vrn-H1 and Vrn-H3. We showed that allelic variants of these flowering time genes strongly react to environmental cues. This information can be used to design novel breeding strategies such as precise backcrossing of suitable developmental genes into regionally adapted cultivars. Our data also provide evidence that wild barley germplasm may be useful to improve yield in low-yielding environments, for instance, in the Middle East, as well as in high-yielding environments, for instance, in Northern and Central Europe. This knowledge may be transferred to related crop species like wheat and rice to secure the rising global food demand for cereals.
Materials and Methods
Plant material
HEB-YIELD, a subset of the wild barley nested association mapping (NAM) population Halle Exotic Barley-25 (HEB-2518), was used in yield trials. HEB-25 originated from crossing 25 diverse wild barley accessions (Hordeum vulgare ssp. spontaneum and H.v. ssp. agriocrithon) with the German spring barley elite cultivar Barke (Hordeum vulgare ssp. vulgare). HEB-25 comprises 1,420 BC1S3 derived lines (backcrossed with Barke) grouped into 25 families (for more details see Maurer et al.18).
The HEB-YIELD subset consists of 48 HEB-25 lines that were selected from HEB-25 to ensure the absence of brittleness and a good threshability enabling accurate yield estimation in field trials. In addition, the final HEB-YIELD lines were selected to independently segregate at four major flowering time loci, which exhibited major plant developmental effects in HEB-25: Ppd-H1, Sdw1, Vrn-H1 and Vrn-H318,20,25.
Genotypic data
The complete HEB-25 population was genotyped in generation BC1S3 using the barley Infinium iSelect 9k SNP chip (see)18. The diagnostic markers i_BK_16, i_12_30924, i_11_10705 and i_12_10218, co-segregating with the four flowering time genes Ppd-H1, Sdw1, Vrn-H1 and Vrn-H3, respectively, were used for selection of HEB-YIELD lines carrying homozygous elite versus homozygous wild barley alleles. In total, HEB-YIELD includes wild barley chromosomal segments derived from 20 wild barley donors. (Supplementary Table S1a). These donors contributed wild barley alleles at each of the studied four flowering time genes. Supplementary Table S1b provides exome capture-based sequence data of four flowering time genes, Ppd-H1, Ppd-H2, sdw1 and Vrn-H1, collected through the WHEALBI consortium (https://www.whealbi.eu/) and kindly provided by Drs. Micha Bayer and Joanne Russell, The James Hutton Institute, Dundee, UK.
Field trials
The HEB-YIELD population was grown at five locations worldwide during two years (2015 and 2016), resulting in ten environments. The locations are (from north to south): Dundee (United Kingdom; 56°28′53.71″N 3°6′35.17″W), Halle (Germany; 51°29′46.05″N 11°59′29.58″E), Al-Karak (Jordan; 31°16′34.03″N 35°44′24.94″E), Dubai (United Arab Emirates; 25°5′44.40″N 55°23′24.48″E) and Adelaide (Australia; 35°19′18.5″S 138°53′07.5″E). A detailed description for each location is given in Supplementary Table S2a. The full set of 48 HEB-YIELD lines was cultivated at each location except in Adelaide. Due to lack of seeds, in Adelaide only 34 and 47 HEB-YIELD lines were cultivated in 2015 and 2016, respectively (Supplementary Table S2d). At each location, additional local check cultivars were cultivated, for example: ‘Odyssey’ (Limagrain, 2011) in Dundee, ‘Quench’ (Syngenta, 2006) in Halle, ‘Rum’ (CIMMYT, 1986) in Al-Karak, ‘58/1 A’ (ICBA, 2002) in Dubai and ‘Navigator’ (University of Adelaide, 2012) in Adelaide.
At each location, a control treatment and a site-specific stress treatment was applied. Stress treatments were nitrogen deficiency in Dundee and Halle, drought stress in Al-Karak, salt stress in Dubai and drought stress in Adelaide (see Supplementary Table S2c). Therefore, lines of the stress treatment received no nitrogen fertilizer in Dundee and Halle, no drip irrigation in Al-Karak and a saline water drip irrigation in Dubai. In Adelaide, only one treatment was applied per season due to lack of seeds. In this case, the two contrasting seasons represented the treatments where 2015 was regarded as the drought stress treatment with only 159 mm precipitation during the growing period and 2016 as the control treatment with 484 mm precipitation.
On average, each HEB-YIELD line was replicated three to four times per treatment. A randomized complete block design was chosen as test design for the trials, with the exception of Dubai and Adelaide where a completely randomized design within each treatment was applied. The trials were conducted in accordance to local practices regarding tillage, fertilization and pest management. Additional information on plant cultivation is provided in Supplementary Table S2b.
Phenotypic data
Eleven developmental and yield related traits were investigated. A description of where and how each trait was measured is given in Supplementary Table S3.
Statistical analyses
All statistical analyses were carried out with SAS 9.4 (SAS Institute Inc., Cary, NC, USA)100. Variance components (defined as random) were estimated with PROC VARCOMP and broad sense heritabilities (h2) for each trait within locations and treatments were calculated across years following the formula:
where
For traits analyzed in a single year, repeatability (rep) was calculated following the formula:
The analysis of variance (ANOVA) across locations was calculated with PROC MIXED to test for the presence of genotype, location and year effects. For this purpose, the main effects (genotype, location and year), as well as their corresponding interaction effects were treated as fixed effects in the following model:
where
Fixed effects are written in bold
Best linear unbiased estimators (BLUEs) were estimated using the PROC MIXED procedure. The BLUEs for each HEB-YIELD line were computed across years and for each treatment level and location separately. Genotype and treatment were modelled as fixed effects and year as a random effect:
where
Fixed effects are written in bold
For location Adelaide BLUEs were calculated within years and the model was restricted to a fixed effect of genotype.
Pearson correlation coefficients (r) between trait BLUEs were calculated via PROC CORR. Furthermore, to test for significant treatment effects a simple t-test (PROC TTEST) and an ANOVA within locations were performed (PROC MIXED). The ANOVA model included the main effects (genotype, treatment and year) and their corresponding interaction effects as fixed effects (comparable to model III). In addition, a one-factorial ANOVA was computed to test for significant location effects within treatments where only the main effect (location) was included, followed by a Tukey test (PROC GLM).
Performance of the HEB-YIELD lines was compared to an adapted check cultivar from the corresponding location (see field trials above) by conducting a Dunnett test101 (PROC MIXED). To enable an easier comparison between the lines the relative performance (RP) was calculated as:
To check for significance and estimate effects of the four flowering candidate genes Ppd-H1, Sdw1, Vrn-H1 and Vrn-H3, a simple linear regression model (PROC GLM) was fitted for each candidate gene applying BLUEs across years. Each model included a single locus-specific SNP mentioned above, modeled as a quantitative variable representing the wild allele dosage20.
All figures were created using R (3.4.2)102 with the package ggplot2 (2.2.1)103.
References
Godfray, H. C. J. et al. Food security: the challenge of feeding 9 billion people. Science 327, 812–818, https://doi.org/10.1126/science.1185383 (2010).
Powell, N., Ji, X., Ravash, R., Edlington, J. & Dolferus, R. Yield stability for cereals in a changing climate. Functional Plant Biol. 39, 539, https://doi.org/10.1071/FP12078 (2013).
Samson, J., Berteaux, D., McGill, B. J. & Humphries, M. M. Geographic disparities and moral hazards in the predicted impacts of climate change on human populations. Global Ecology and Biogeography 20, 532–544, https://doi.org/10.1111/j.1466-8238.2010.00632.x (2011).
Field, C. B. et al. Detection and attribution of observed impacts. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, 2014.
Zhou, M. X. In Genetics and Improvement of Barley Malt Quality, edited by G. Zhang & C. Li (Springer-Verlag Berlin Heidelberg, Berlin, Heidelberg), pp. 1–17 (2010).
FAOSTAT. FAOSTAT. Available at, http://www.fao.org/faostat/en/#home (2017).
Munns, R. & Tester, M. Mechanisms of salinity tolerance. Annual Review of Plant Biology 59, 651–681, https://doi.org/10.1146/annurev.arplant.59.032607.092911 (2008).
Baik, B.-K., Newman, C. W. & Newman, R. K. In Barley: Production, Improvement, and Uses, edited by S. E. Ullrich (Wiley-Blackwell, Oxford, UK, 2011).
Nevo, E. et al. Evolution of wild cereals during 28 years of global warming in Israel. Proceedings of the National Academy of Sciences of the United States of America 109, 3412–3415, https://doi.org/10.1073/pnas.1121411109 (2012).
Sreenivasulu, N., Graner, A. & Wobus, U. Barley genomics. An overview. International Journal of Plant Genomics 2008, 486258, https://doi.org/10.1155/2008/486258 (2008).
Badr, A. et al. On the origin and domestication history of Barley (Hordeum vulgare). Mol Biol Evol 17, 499–510 (2000).
Morrell, P. L. & Clegg, M. T. Genetic evidence for a second domestication of barley (Hordeum vulgare) east of the Fertile Crescent. Proceedings of the National Academy of Sciences of the United States of America 104, 3289–3294, https://doi.org/10.1073/pnas.0611377104 (2007).
Dai, F. et al. Tibet is one of the centers of domestication of cultivated barley. Proceedings of the National Academy of Sciences of the United States of America 109, 16969–16973, https://doi.org/10.1073/pnas.1215265109 (2012).
Zamir, D. Improving plant breeding with exotic genetic libraries. Nature Reviews Genetics 2, 983–989, https://doi.org/10.1038/35103589 (2001).
McCouch, S. et al. Agriculture: Feeding the future. Nature 499, 23–24, https://doi.org/10.1038/499023a (2013).
Zhang, H., Mittal, N., Leamy, L. J., Barazani, O. & Song, B.-H. Back into the wild-Apply untapped genetic diversity of wild relatives for crop improvement. Evolutionary Applications 10, 5–24, https://doi.org/10.1111/eva.12434 (2017).
Hajjar, R. & Hodgkin, T. The use of wild relatives in crop improvement. A survey of developments over the last 20 years. Euphytica 156, 1–13, https://doi.org/10.1007/s10681-007-9363-0 (2007).
Maurer, A. et al. Modelling the genetic architecture of flowering time control in barley through nested association mapping. BMC Genomics 16, 290, https://doi.org/10.1186/s12864-015-1459-7 (2015).
Nice, L. M. et al. Development and Genetic Characterization of an Advanced Backcross-Nested Association Mapping (AB-NAM) Population of Wild × Cultivated Barley. Genetics 203, 1453–1467, https://doi.org/10.1534/genetics.116.190736 (2016).
Maurer, A., Draba, V. & Pillen, K. Genomic dissection of plant development and its impact on thousand grain weight in barley through nested association mapping. Journal of Experimental Botany 67, 2507–2518, https://doi.org/10.1093/jxb/erw070 (2016).
Russell, J. R. et al. Exome sequencing of geographically diverse barley landraces and wild relatives gives insights into environmental adaptation. Nature Genetics 48, 1024–1030, https://doi.org/10.1038/ng.3612 (2016).
Saade, S. et al. Yield-related salinity tolerance traits identified in a nested association mapping (NAM) population of wild barley. Scientific Reports 6, 32586, https://doi.org/10.1038/srep32586 (2016).
Nice, L. M. et al. Mapping Agronomic Traits in a Wild Barley Advanced Backcross–Nested Association Mapping Population. Crop Science 57, 1199, https://doi.org/10.2135/cropsci2016.10.0850 (2017).
Vatter, T. et al. A nested association mapping population identifies multiple small effect QTL conferring resistance against net blotch (Pyrenophora teres f. teres) in wild barley. Plos One 12, e0186803, https://doi.org/10.1371/journal.pone.0186803 (2017).
Herzig, P. et al. Contrasting genetic regulation of plant development in two European environments revealed by wild barley nested association mapping. Journal of Experimental Botany, https://doi.org/10.1093/jxb/ery002 (2018).
Ceccarelli, S. et al. Plant breeding and climate changes. J. Agric. Sci. 148, 627–637, https://doi.org/10.1017/S0021859610000651 (2010).
Slafer, G. A. Genetic basis of yield as viewed from a crop physiologist’s perspective. Ann Applied Biology 142, 117–128, https://doi.org/10.1111/j.1744-7348.2003.tb00237.x (2003).
Worland, A. J. The influence of flowering time genes on environmental adaptability in European wheats. Euphytica 89, 49–57, https://doi.org/10.1007/BF00015718 (1996).
Cockram, J. et al. Control of flowering time in temperate cereals: genes, domestication, and sustainable productivity. Journal of Experimental Botany 58, 1231–1244, https://doi.org/10.1093/jxb/erm042 (2007).
Kamran, A., Iqbal, M. & Spaner, D. Flowering time in wheat (Triticum aestivum L.). A key factor for global adaptability. Euphytica 197, 1–26, https://doi.org/10.1007/s10681-014-1075-7 (2014).
Slafer, G. A., Kantolic, A. G., Appendino, M. L., Miralles, D. J. & Savin, R. In Crop physiology, edited by V. O. Sadras & D. Calderini (Academic Press is an imprint of Elsevier, London, UK, Waltham, M. A., USA), pp. 277–308 (2009).
Kazan, K. & Lyons, R. The link between flowering time and stress tolerance. Journal of Experimental Botany 67, 47–60, https://doi.org/10.1093/jxb/erv441 (2016).
Trevaskis, B., Hemming, M. N., Dennis, E. S. & Peacock, W. J. The molecular basis of vernalization-induced flowering in cereals. Trends in Plant Science 12, 352–357, https://doi.org/10.1016/j.tplants.2007.06.010 (2007).
Andrés, F. & Coupland, G. The genetic basis of flowering responses to seasonal cues. Nature Reviews Genetics 13, 627–639, https://doi.org/10.1038/nrg3291 (2012).
Cho, L.-H., Yoon, J. & An, G. The control of flowering time by environmental factors. The Plant Journal 90, 708–719, https://doi.org/10.1111/tpj.13461 (2017).
Blümel, M., Dally, N. & Jung, C. Flowering time regulation in crops-what did we learn from Arabidopsis? Current Opinion in Biotechnology 32, 121–129, https://doi.org/10.1016/j.copbio.2014.11.023 (2015).
Turner, A. S., Beales, J., Faure, S., Dunford, R. P. & Laurie, D. A. The pseudo-response regulator Ppd-H1 provides adaptation to photoperiod in barley. Science 310, 1031–1034, https://doi.org/10.1126/science.1117619 (2005).
Rollins, J. A. et al. Variation at the vernalisation genes Vrn-H1 and Vrn-H2 determines growth and yield stability in barley (Hordeum vulgare) grown under dryland conditions in Syria. Tag 126, 2803–2824, https://doi.org/10.1007/s00122-013-2173-y (2013).
Gawroński, P. & Schnurbusch, T. High-density mapping of the earliness per se-3Am (Eps-3Am) locus in diploid einkorn wheat and its relation to the syntenic regions in rice and Brachypodium distachyon L. Mol Breeding 30, 1097–1108, https://doi.org/10.1007/s11032-011-9697-0 (2012).
Kuczyńska, A. et al. Effects of the semi-dwarfing sdw1/denso gene in barley. Journal of Applied Genetics 54, 381–390, https://doi.org/10.1007/s13353-013-0165-x (2013).
Barnabás, B., Jäger, K. & Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant, Cell & Environment 31, 11–38, https://doi.org/10.1111/j.1365-3040.2007.01727.x (2008).
Sacks, W. J. & Kucharik, C. J. Crop management and phenology trends in the U.S. Corn Belt. Impacts on yields, evapotranspiration and energy balance. Agricultural and Forest Meteorology 151, 882–894, https://doi.org/10.1016/j.agrformet.2011.02.010 (2011).
Alvarez Prado, S., Gallardo, J. M., Kruk, B. C. & Miralles, D. J. Strategies for yield determination of bread wheat and two-row barley growing under different environments. A comparative study. Field Crops Research 203, 94–105, https://doi.org/10.1016/j.fcr.2016.12.013 (2017).
Shakhatreh, Y., Kafawin, O., Ceccarelli, S. & Saoub, H. Selection of Barley Lines for Drought Tolerance in Low-Rainfall Areas. J Agron Crop Sci 186, 119–127, https://doi.org/10.1046/j.1439-037X.2001.00459.x (2001).
Al-Ajlouni, Z. et al. Impact of Pre-Anthesis Water Deficit on Yield and Yield Components in Barley (Hordeum vulgare L.) Plants Grown under Controlled Conditions. Agronomy 6, 33, https://doi.org/10.3390/agronomy6020033 (2016).
Samarah, N. H. Effects of drought stress on growth and yield of barley. Agron. Sustain. Dev. 25, 145–149, https://doi.org/10.1051/agro:2004064 (2005).
Fischer, R. A. Number of kernels in wheat crops and the influence of solar radiation and temperature. J. Agric. Sci. 105, 447, https://doi.org/10.1017/S0021859600056495 (1985).
Cossani, C. M., Slafer, G. A. & Savin, R. Yield and biomass in wheat and barley under a range of conditions in a Mediterranean site. Field Crops Research 112, 205–213, https://doi.org/10.1016/j.fcr.2009.03.003 (2009).
Sreenivasulu, N. & Schnurbusch, T. A genetic playground for enhancing grain number in cereals. Trends in Plant Science 17, 91–101, https://doi.org/10.1016/j.tplants.2011.11.003 (2012).
Slafer, G. A., Savin, R. & Sadras, V. O. Coarse and fine regulation of wheat yield components in response to genotype and environment. Field Crops Research 157, 71–83, https://doi.org/10.1016/j.fcr.2013.12.004 (2014).
Hay, R. K. M. Harvest index: a review of its use in plant breeding and crop physiology. Ann Applied Biology 126, 197–216, https://doi.org/10.1111/j.1744-7348.1995.tb05015.x (1995).
Fischer, R. A. & Edmeades, G. O. Breeding and Cereal Yield Progress. Crop Science 50, S-85–S-98, https://doi.org/10.2135/cropsci2009.10.0564 (2010).
Reynolds, M. et al. Raising yield potential in wheat. Journal of Experimental Botany 60, 1899–1918, https://doi.org/10.1093/jxb/erp016 (2009).
Alqudah, A. M. & Schnurbusch, T. Barley Leaf Area and Leaf Growth Rates Are Maximized during the Pre-Anthesis Phase. Agronomy 5, 107–129, https://doi.org/10.3390/agronomy5020107 (2015).
Gao, H. et al. Days to heading 7, a major quantitative locus determining photoperiod sensitivity and regional adaptation in rice. Proceedings of the National Academy of Sciences of the United States of America 111, 16337–16342, https://doi.org/10.1073/pnas.1418204111 (2014).
Sultan, S. E. Phenotypic plasticity for plant development, function and life history. Trends in Plant Science 5, 537–542, https://doi.org/10.1016/S1360-1385(00)01797-0 (2000).
Roy, S. J., Tucker, E. J. & Tester, M. Genetic analysis of abiotic stress tolerance in crops. Current Opinion in Plant Biology 14, 232–239, https://doi.org/10.1016/j.pbi.2011.03.002 (2011).
Jones, H. et al. Population-based resequencing reveals that the flowering time adaptation of cultivated barley originated east of the Fertile Crescent. Mol Biol Evol 25, 2211–2219, https://doi.org/10.1093/molbev/msn167 (2008).
Lister, D. L. et al. Latitudinal variation in a photoperiod response gene in European barley. Insight into the dynamics of agricultural spread from ‘historic’ specimens. Journal of Archaeological Science 36, 1092–1098, https://doi.org/10.1016/j.jas.2008.12.012 (2009).
Digel, B. et al. Photoperiod-H1 (Ppd-H1) Controls Leaf Size. Plant Physiology 172, 405–415, https://doi.org/10.1104/pp.16.00977 (2016).
Ejaz, M. & Korff, M. V. The Genetic Control of Reproductive Development under High Ambient Temperature. Plant Physiology 173, 294–306, https://doi.org/10.1104/pp.16.01275 (2017).
Cockram, J., Hones, H. & O’Sullivan, D. M. Genetic variation at flowering time loci in wild and cultivated barley. Plant Genet. Resour. 9, 264–267, https://doi.org/10.1017/S1479262111000505 (2011).
Johansson, M. & Staiger, D. Time to flower: interplay between photoperiod and the circadian clock. Journal of Experimental Botany 66, 719–730, https://doi.org/10.1093/jxb/eru441 (2015).
Faure, S., Higgins, J., Turner, A. & Laurie, D. A. The Flowering Locus T-like gene family in barley (Hordeum vulgare). Genetics 176, 599–609, https://doi.org/10.1534/genetics.106.069500 (2007).
Casao, M. C. et al. Expression analysis of vernalization and day-length response genes in barley (Hordeum vulgare L.) indicates that VRNH2 is a repressor of PPDH2 (HvFT3) under long days. Journal of Experimental Botany 62, 1939–1949, https://doi.org/10.1093/jxb/erq382 (2011).
Casao, C. M. et al. Adaptation of barley to mild winters: a role for PPDH2. BMC Plant Biology 11, 164, https://doi.org/10.1186/1471-2229-11-164 (2011).
Mulki, M. A., Bi, X. & Korff, M. V. Flowering Locus T3 Controls Spikelet Initiation But Not Floral Development. Plant Physiology 178, 1170–1186, https://doi.org/10.1104/pp.18.00236 (2018).
Bergelson, J. & Roux, F. Towards identifying genes underlying ecologically relevant traits in Arabidopsis thaliana. Nature reviews. Genetics 11, 867–879, https://doi.org/10.1038/nrg2896 (2010).
Hedden, P. The genes of the Green Revolution. Tig 19, 5–9 (2003).
Jia, Q. et al. Expression level of a gibberellin 20-oxidase gene is associated with multiple agronomic and quality traits in barley. Tag 122, 1451–1460, https://doi.org/10.1007/s00122-011-1544-5 (2011).
Jia, Q. et al. Molecular characterization and functional analysis of barley semi-dwarf mutant Riso no. 9265. BMC Genomics 16, 927, https://doi.org/10.1186/s12864-015-2116-x (2015).
Coventry, S. J., Barr, A. R., Eglinton, J. K. & McDonald, G. K. The determinants and genome locations influencing grain weight and size in barley (Hordeum vulgare L.). Aust. J. Agric. Res. 54, 1103, https://doi.org/10.1071/AR02194 (2003).
Baum, M. et al. QTLs for agronomic traits in the Mediterranean environment identified in recombinant inbred lines of the cross ‘Arta’ x H. spontaneum 41-1. Tag 107, 1215–1225, https://doi.org/10.1007/s00122-003-1357-2 (2003).
Pinthus, M. J., Gale, M. D., Appleford, N. E. J. & Lenton, J. R. Effect of Temperature on Gibberellin (GA) Responsiveness and on Endogenous GA1 Content of Tall and Dwarf Wheat Genotypes. Plant Physiology 90, 854–859, https://doi.org/10.1104/pp.90.3.854 (1989).
Hellewell, K. B., Rasmusson, D. C. & Gallo-Meagher, M. Enhancing Yield of Semidwarf Barley. Crop Science 40, 352, https://doi.org/10.2135/cropsci2000.402352x (2000).
Jia, Q. et al. GA-20 oxidase as a candidate for the semidwarf gene sdw1/denso in barley. Functional & Integrative Genomics 9, 255–262, https://doi.org/10.1007/s10142-009-0120-4 (2009).
Mutasa-Göttgens, E. & Hedden, P. Gibberellin as a factor in floral regulatory networks. Journal of Experimental Botany 60, 1979–1989, https://doi.org/10.1093/jxb/erp040 (2009).
Song, Y. H., Ito, S. & Imaizumi, T. Flowering time regulation: photoperiod- and temperature-sensing in leaves. Trends in Plant Science 18, 575–583, https://doi.org/10.1016/j.tplants.2013.05.003 (2013).
Yan, L. et al. Positional cloning of the wheat vernalization gene VRN1. Proceedings of the National Academy of Sciences of the United States of America 100, 6263–6268, https://doi.org/10.1073/pnas.0937399100 (2003).
Yan, L. et al. The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science 303, 1640–1644, https://doi.org/10.1126/science.1094305 (2004).
Oliver, S. N., Deng, W., Casao, C. M. & Trevaskis, B. Low temperatures induce rapid changes in chromatin state and transcript levels of the cereal VERNALIZATION1 gene. Journal of Experimental Botany 64, 2413–2422, https://doi.org/10.1093/jxb/ert095 (2013).
Deng, W. et al. Direct links between the vernalization response and other key traits of cereal crops. Nature Communications 6, 5882, https://doi.org/10.1038/ncomms6882 (2015).
Corbesier, L. et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 316, 1030–1033, https://doi.org/10.1126/science.1141752 (2007).
Loscos, J., Igartua, E., Contreras-Moreira, B., Gracia, M. P. & Casas, A. M. HvFT1 polymorphism and effect-survey of barley germplasm and expression analysis. Front. Plant Sci. 5, 251, https://doi.org/10.3389/fpls.2014.00251 (2014).
Yan, L. et al. The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proceedings of the National Academy of Sciences of the United States of America 103, 19581–19586, https://doi.org/10.1073/pnas.0607142103 (2006).
Nitcher, R., Distelfeld, A., Tan, C., Yan, L. & Dubcovsky, J. Increased copy number at the HvFT1 locus is associated with accelerated flowering time in barley. Molecular genetics and genomics: MGG 288, 261–275, https://doi.org/10.1007/s00438-013-0746-8 (2013).
Tanksley, S. D. & McCouch, S. R. Seed Banks and Molecular Maps. Unlocking Genetic Potential from the Wild. Science 277, 1063–1066, https://doi.org/10.1126/science.277.5329.1063 (1997).
Schmalenbach, I., Körber, N. & Pillen, K. Selecting a set of wild barley introgression lines and verification of QTL effects for resistance to powdery mildew and leaf rust. TAG 117, 1093–1106, https://doi.org/10.1007/s00122-008-0847-7 (2008).
Honsdorf, N., March, T. J., Berger, B., Tester, M. & Pillen, K. High-throughput phenotyping to detect drought tolerance QTL in wild barley introgression lines. Plos One 9, e97047, https://doi.org/10.1371/journal.pone.0097047 (2014).
Reuscher, S., Kolter, A., Hoffmann, A., Pillen, K. & Krämer, U. Quantitative Trait Loci and Inter-Organ Partitioning for Essential Metal and Toxic Analogue Accumulation in Barley. Plos One 11, e0153392, https://doi.org/10.1371/journal.pone.0153392 (2016).
Korff, M., von, Wang, H., Léon, J. & Pillen, K. AB-QTL analysis in spring barley. III. Identification of exotic alleles for the improvement of malting quality in spring barley (H. vulgare ssp. spontaneum). Mol Breeding 21, 81–93, https://doi.org/10.1007/s11032-007-9110-1 (2008).
March, T. J. et al. Identification of proteins associated with malting quality in a subset of wild barley introgression lines. Proteomics 12, 2843–2851, https://doi.org/10.1002/pmic.201200117 (2012).
Talame, V. et al. Identification of Hordeum spontaneum QTL alleles improving field performance of barley grown under rainfed conditions. Ann Applied Biology 144, 309–319, https://doi.org/10.1111/j.1744-7348.2004.tb00346.x (2004).
Nevo, E. & Chen, G. Drought and salt tolerances in wild relatives for wheat and barley improvement. Plant, Cell & Environment 33, 670–685, https://doi.org/10.1111/j.1365-3040.2009.02107.x (2010).
Feuillet, C., Langridge, P. & Waugh, R. Cereal breeding takes a walk on the wild side. Tig 24, 24–32, https://doi.org/10.1016/j.tig.2007.11.001 (2008).
Battisti, D. S. & Naylor, R. L. Historical warnings of future food insecurity with unprecedented seasonal heat. Science 323, 240–244, https://doi.org/10.1126/science.1164363 (2009).
Brush, S. B. (ed.). Genes in the field. On-farm conservation of crop diversity (International Plant Genetic Resources Centre, Rome Ont., 2011).
Al-Abdallat, A. M. et al. Assessment of genetic diversity and yield performance in Jordanian barley (Hordeum vulgare L.) landraces grown under Rainfed conditions. Bmc Plant Biology 17, 191, https://doi.org/10.1186/s12870-017-1140-1 (2017).
Purugganan, M. D. & Fuller, D. Q. The nature of selection during plant domestication. Nature 457, 843–848, https://doi.org/10.1038/nature07895 (2009).
SAS. SAS (SAS Institute inc., Cary, North Carolina, USA, 2013).
Dunnett, C. W. A Multiple Comparison Procedure for Comparing Several Treatments with a Control. Journal of the American Statistical Association, 1096–1121 (1955).
R Development Core Team. R. A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, Austria, 2008).
Wickham, H. ggplot2. Elegant Graphics for Data Analysis (Springer-Verlag, New York, USA, 2009).
Acknowledgements
This work was financially supported by the German Research Foundation (DFG) via the priority program 1530: ‘Flowering time control - from natural variation to crop improvement’ with grants Pi339/7-1 and Pi339/7-2) and via ERA-NET for Coordinating Action in Plant Sciences (ERA-CAPS) grant Pi339/8-1. Funding from King Abdullah University of Science and Technology (KAUST) is also gratefully acknowledged. We are grateful to a multitude of research assistants, located at the James Hutton Institute in Dundee, the Martin Luther University in Halle, the International Center for Agricultural Research in the Dry Areas and NCARE in Al-Karak, the International Center for Biosaline Agriculture in Dubai and at the University of Adelaide for their excellent technical support in conducting the global field trials. We are grateful to Drs. Micha Bayer and Joanne Russell, The James Hutton Institute, Dundee, UK, providing exome capture-based SNP data of Ppd-H1, Ppd-H2, sdw1 and Vrn-H1, collected through the WHEALBI consortium (https://www.whealbi.eu/).
Author information
Authors and Affiliations
Contributions
M.W. conducted the field trials in 2015 and 2016 in Halle, gathered and analyzed the phenotypic data for all test locations, created the figures, and drafted the manuscript. A.M. planned the field trials in 2015 and 2016 in Halle, supported the analysis of the phenotypic data and drafted the manuscript. A.F., H.B. and W.T. planned and conducted the field trials in 2015 and 2016 in Dundee. M.B. and A.A. planned and conducted the field trials in 2014/15 and 2015/16 in Al-Karak. M.T. and M.S. planned and conducted the field trials in 2014/15 and 2015/16 in Dubai. J.E., A.P. and T.M. planned and conducted the field trials in 2015 and 2016 in Adelaide. K.P. acquired the funding, coordinated the collaboration between the project partners, finalized the exome capture data and drafted the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wiegmann, M., Maurer, A., Pham, A. et al. Barley yield formation under abiotic stress depends on the interplay between flowering time genes and environmental cues. Sci Rep 9, 6397 (2019). https://doi.org/10.1038/s41598-019-42673-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-42673-1
This article is cited by
-
Finding stable and closely linked QTLs against spot blotch in different planting dates during the adult stage in barley
Scientific Reports (2024)
-
Hordatines, dimerised hydroxycinnamoylagmatine conjugates of barley (Hordeum vulgare L.): an appraisal of the biosynthesis, chemistry, identification and bioactivities
Phytochemistry Reviews (2024)
-
Morphological, pathogenic and molecular analysis of Bipolaris sorokiniana inciting spot blotch of barley in India
Indian Phytopathology (2024)
-
Investigating the genetic control of plant development in spring barley under speed breeding conditions
Theoretical and Applied Genetics (2024)
-
A wild barley nested association mapping population shows a wide variation for yield-associated traits to be used for breeding in Australian environment
Euphytica (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}