
Abstract
The Prince Creek Formation of Alaska, a rock unit that represents lower coastal plain and delta deposits, is one of the most important formations in the world for understanding vertebrate ecology in the Arctic during the Cretaceous. Here we report on an isolated cranial material, supraoccipital, of a lambeosaurine hadrosaurid from the Liscomb Bonebed of the Prince Creek Formation. The lambeosaurine supraoccipital has well-developed squamosal bosses and a short sutural surface with the exoccipital-opisthotic complex, and is similar to lambeosaurine supraoccipitals from the Dinosaur Park Formation in having anteriorly positioned squamosal bosses. Affinities with Canadian lambeosaurines elucidate more extensive faunal exchange between the Arctic and lower paleolatitudes which was previously suggested by the presence of Edmontosaurus, Pachyrhinosaurus, tyrannosaurids, and troodontids in both regions. The presence of one lambeosaurine and nine hadrosaurine supraoccipitals in the Liscomb Bonebed suggests hadrosaurine dominated faunal structure as in the Careless Creek Quarry of the USA that was also deposited under a near-shore environment. It differs from the lambeosaurine dominant structures of localities in Russia and China interpreted as inland environments. This may suggest that lambeosaurines had less preference for near-shore environments than hadrosaurines in both Arctic and lower paleolatitudes.
Similar content being viewed by others

Introduction
Vertebrate animals in the Arctic have experienced physiological, behavioral, and morphological adaptations to survive in an extreme environment1,2,3. Rocks from the North Slope of Alaska are important for the understanding of the ecology of fossil vertebrates in the Arctic during the Cretaceous Period4,5. There are abundant fossiliferous exposures of the lower part of the Prince Creek Formation on the North Slope, which range from Campanian to early Maastrichtian in age6. The Prince Creek Formation is a non-marine succession deposited on a high-latitude, low-gradient alluvial/coastal plain. An integrated reconstruction of pedogenic processes and biota7 suggests that this ancient Arctic coastal plain was influenced by seasonally fluctuating water table levels and floods, and in distal areas, marine waters. The formation has yielded a diverse dinosaur assemblage that includes ceratopsids, dromaeosaurids, hadrosaurids, basal ornithopods, pachycephalosaurids, troodintids, and tyrannosaurids4,5,8,9,10,11,12,13,14,15,16.
The Liscomb Bonebed is one of the most prolific dinosaur bearing localities within this rock unit. Radiometric dating based on tephra near the bonebed indicates that the Liscomb Bonebed is early Maastrichtian with estimated ages of 71–68 Ma17, 69.1 ± 0.3 Ma18, and younger than 69.2 ± 0.5 Ma19. These dates are concordant with palynomorph analyses which suggest early Maastrichtian age20,21. The rocks were deposited at an estimated paleolatitude of 74.5° ± 7.5°22. The Liscomb Bonebed is from the distal area of coastal plain and is represented by lower delta plain facies7. Further, the stratigraphic interval containing the Liscomb Bonebed represents a series of episodic floods19,23 and specifically these episodic flood events created deposition by fine-grained viscous hyperconcentrated flows that transported the remains of scores of juvenile dinosaurs onto floodplains adjacent to distributary channels. The Liscomb Bonebed is characterized by high specimen density (up to 160–220 elements/m2) and has yielded over six thousand bones15,24. It is a monodominant multitaxic bonebed consisting of three theropod taxa (dromaeosaurid, troodontid, and tyrannosaurid)7,10,25 and hadrosaurid skeletal elements which comprises 98.5% of dinosaur skeletal elements23.
The Hadrosauridae is a derived clade of Hadrosauroidea and comprise two stem-sister lineages, Lambeosaurinae and Hadrosaurinae26. The Liscomb hadrosaurid materials were initially identified as Lambeosaurinae20, although osteological features to support the identification were not provided. Comparisons based on isolated cranial elements later demonstrated that the Liscomb hadrosaurs showed close similarity to the hadrosaurine Edmontosaurus saskatchewanensis27, which is now considered a junior synonym of Edmontosaurus annectens28. Since then, a general consensus formed that the Liscomb hadrosaur bones represent specimens of Edmontosaurus5,23,29,30,31,32,33,34. Recently, it was proposed that the Liscomb hadrosaur bones represent a new distinct hadrosaurine taxon, Ugrunaaluk kuukpikensis15. However, subsequent workers argued that the proposed new taxon was invalid in part because it was diagnosed on immature growth stage features preserved in the known specimens35. Despite of the taxonomic controversy, these studies have agreed upon the presence of a hadrosaurine hadrosaurid in the Liscomb Bonebed.
Here we report the first definitive lambeosaurine hadrosaurid fossil from the Liscomb Bonebed (DMNH 2014–12–266), represented by an isolated cranial material, a supraoccipital. The supraoccipital demonstrates that the Liscomb Bonebed contains both lambeosaurine and hadrosaurine materials. While co-occurrences of hadrosaurine and lambeosaurine are widely known in the northern hemisphere (e.g., Careless Creek Quarry36,37,38 and Jack’s Birthday Site39 of Montana, United States; Blagoveschensk locality40,41 and Kundur42,43 localities of southern Amur region, Russia; and Wulaga locality44 of northern Heilongjiang Province, China), the Liscomb Bonebed is the first to demonstrate the co-occurrence in the Arctic. Therefore, the new discovery offers an important opportunity to infer possible determinant factors of hadrosaurid taxonomic structure in the Arctic, in comparison with lower latitude regions.
Results
Systematic paleontology
Dinosauria Owen, 184245
Ornithischia Seeley, 188746
Cerapoda Sereno, 198647
Ornithopoda Marsh, 188148
Iguanodontia Dollo, 188849
Hadrosauridae Cope, 187050
Lambeosaurinae Parks, 192351
Description
The new supraoccipital (DMNH 2014-12-266; Fig. 1) is nearly complete but missing both anterior processes and anterodorsal end of the ascending process. Its maximum width along the posteroventral margin is 44.8 mm, which is slightly larger than those of Edmontosaurus sp. specimens from the Liscomb Bonebed, nearly equivalent with that of the indeterminate lambeosaurine CMN 017052,53, and smaller than that of Prosaurolophus maximus MOR 447-8-8-7-1452 (Table 1). The width is also much less than the posteriorly exposed supraoccipital surfaces of adult articulated skulls of Edmontosaurus regalis (100.3 mm, CMN 2278), Gryposaurus notabilis (102.4 mm, CMN 2288), Hypacrosaurus stebingeri (87.1 mm, MOR 553 s; 98.0 mm, MOR 455), and Lambeosaurus lambei (79.5 mm, ROM 1218). Its small size may indicates that the supraoccipital (DMNH 2014-12-266) belonged to an immature individual.

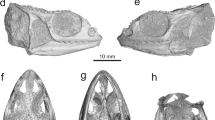
Lambeosaurine supraoccipital (DMNH 2014-12-266) from the Liscomb Bonebed. (a) Dorsal view. (b) Ventral view. (c) Left lateral view. (d) Posterior view. (e) Anterior view. (f) Right lateral view. Abbreviations: asp, ascending process; eo, articulation surface for the exoccipital-opisthotic complex; eog, exoccipital groove; ptg, post-temporal groove; sqb, squamosal boss. Scale = 2 cm.
The ascending process is well-developed, taller and wider anteriorly than posteriorly, and divides the bone along the midline (Fig. 1a,d). It extends posterior to the posterior margin of the articulation surface with the exoccipital-opisthotic complex (Fig. 1a,b), unlike the anteriorly positioned ascending process of Edmontosaurus sp. (DMNH 22807, UAMES 4291, UAMES 12727, UAMES 21544; Fig. 2). The ascending process is convergent posteroventrally (Fig. 1a,d) as in Hypacrosaurus stebingeri (USNM 1189354,55), while those of Edmontosaurus sp. (DMNH 22807, UAMES 21544, UAMES 4291, UAMES 12727; Fig. 2b–e) and Prosaurolophus maximus52 are nearly parallel or divergent, and those of non-hadrosaurid hadrosauroids (Bactrosaurus johnsoni56, Batyrosaurus rozhdestvenskyi57, Eolambia caroljonesa58, Eotrachodon orientalis59) are strongly divergent posteroventrally. The dorsal surface of the ascending process is rounded (Fig. 1d) unlike the bi-lobed ascending process of Hypacrosaurus stebingeri (USNM 1189354,55). The dorsal surface is rugose and lacks the nuchal crest. On either side of the ascending process, a deep post-temporal groove53 runs anteroposteriorly (Fig. 1a,d) unlike in supraoccipitals of non-hadrosaurid hadrosauroids which have no distinct post-temporal groove56,57,58,59. The grooves are strongly divergent anteriorly as in the indeterminate lambeosaurine (CMN 017052,53; Fig. 2n), differing from those of Prosaurolophus maximus (MOR 447-8-8-7-1452) and Edmontosaurus sp. (DMNH 22807, UAMES 4291, UAMES 12727, UAMES 21544; Fig. 2b–e) which run nearly parallel to or only slightly divergent from each other. Lateral to the groove, an anterolaterally oriented squamosal boss is present (Fig. 1a,d). The squamosal bosses are well-developed unlike in Prosaurolophus maximus (MOR 447-8-8-7-1452) and Edmontosaurus sp. (DMNH 22807, UAMES 4291, UAMES 12727, UAMES 21544; Fig. 2b–e). The squamosal bosses of DMNH 2014-12-266 are formed solely by the supraoccipital without participation of the exoccipital-opisthotic complex. This morphology of the squamosal boss differs from those of Hypacrosaurus altispinus (AMNH FARB 524860), Hypacrosaurus stebingeri (USNM 1189354,55), the indeterminate lambeosaurine (CMN 017052,53; Fig. 2n), and non-hadrosaurid hadrosauroids (Bactrosaurus johnsoni56, Batyrosaurus rozhdestvenskyi57, Eolambia caroljonesa58), in which the boss is also formed in part of the exoccipital-opisthotic complex.

Hadrosaurid supraoccipitals. DMNH 2014-12-266 (a,f). Edmontosaurus sp: cast of UAMES 21544 (b,g); cast of UAMES 4291 (c,h); DMNH 22807 (d,i); cast of UAMES 12727 (e,j). Indeterminate lambeosaurines: UALVP 48 (k,q); UALVP 53092 (l,r); UALVP 53106 (m,s); CMN 0170 (n,t); UALVP 55300 (o,u); UALVP 54569 (p,v). Dorsal (a–e,k–p) and ventral (f–j,q–v) views. Abbreviations: asp, ascending process; ap, anterior process; eo, articulation surface for the exoccipital-opisthotic complex; eog, exoccipital groove; nc, nuchal crestp, articulation surface for the parietal, ptg, post-temporal groove; sqb, squamosal boss. Scale = 2 cm.
The anteroposterior length of the ventral sutural surface is short (Fig. 1b), being 26.8% of the mediolateral width along the posteroventral margin. The ratio is much smaller than those of the Liscomb Edmontosaurus sp. (DMNH 22807, UAMES 4292, UAMES 21544, UAMES 12727), Edmontosaurus annectens (DMNH EPV 127701), and Prosaurolophus maximus (MOR 447-8-8-7-1452), but resembles lambeosaurines (Table 1). The sutural surface with the exoccipital-opisthotic complex is bowed ventrally toward the midline (Fig. 1d) as in the largest supraoccipital of Edmontosaurus sp. from the Liscomb Bonebed (UAMES 12727), but unlike in the smaller three. The exoccipital groove, located laterodorsal to the ventral sutural surface with the exoccipital-opisthotic complex, faces lateroventrally (Fig. 1b,c,f). The exoccipital groove is mediolaterally narrower than those of Prosaurolophus maximus (MOR 447-8-8-7-1452) and Edmontosaurus sp. (DMNH 22807, UAMES 4291, UAMES 12727, UAMES 21544; Fig. 2g–j), but resembles the indeterminate lambeosaurine (CMN 017052,53; Fig. 2t). The anterior surface of the supraoccipital is smooth and slightly concave to form a part of the endocranial wall (Fig. 1e). However, detailed morphology of the endocranial wall is uncertain because of the missing anterior processes. The height of the supraoccipital is 55.6% of its posteroventral width (Fig. 1d,e; Table 1).
Discussion
The new hadrosaurid supraoccipital DMNH 2014-12-266 largely differs from those of the Liscomb Edmontosaurus sp. in the presence of the well-developed squamosal bosses (Fig. 2a–e) and the short exoccipital articulation surface (Fig. 2f–j; Table 2). The length of the exoccipital articulation surface is equivalent with a phylogenetic character that differentiates hadrosaurines from lambeosaurines and non-hadrosaurid hadrosauroids (degree of the caudal extension of the supraoccipital-exoccipital shelf35,61,62). The well-developed squamosal bosses are widely seen in lambeosaurines as well as in a few non-hadrosaurid hadrosauroids, but has never been reported in hadrosaurines (Fig. 3; Table 2). The appearance of squamosal bosses is an ontogenetic change in the non-hadrosaurid hadrosauroid Bactrosaurus johnsoni56; however, the presence of well-developed squamosal bosses in both juvenile (AMNH FARB 5461, skull length approximately 30% of the holotype MOR 549; Fig. 3g) and adult (MOR 455) individuals of Hypacrosaurus stebingeri suggests that the well-developed squamosal boss of DMNH 2014-12-266 is unlikely to be a result of ontogenetic variation but more likely is a taxonomic difference.

Supraoccipitals of Liscomb hadrosaurid DMNH 2014-12-266 (a,d), an indeterminate lambeosaurine CMN 0170 (b,e), and Edmontosaurus annectens DMNH EPV 127701 (c,f) in dorsal (a–c) and posterior (e–f) views. Posterior views of articulated skulls of Hypacrosaurus stebingeri (g) and Edmontosaurus annectens (h). Abbreviations: asp, ascending process; eo, articulation surface for the exoccipital-opisthotic complex; ptg, post-temporal groove; sqb, squamosal boss; Ex, Exoccipital-opisthotic complex, Sq, Squamosal. Scale = 2 cm (a–f), 5 cm (g,h). Dashed line represents the boundary of supraoccipital in (g,h).
DMNH 2014-12-266 shows a posteroventrally convergent ascending process (Fig. 2a), which is seen only in lambeosaurines (Hypacrosaurus stebingeri USNMH 11893; indeterminate lambeosaurines UALVP 48, UALVP 55300, UALVP 54569; Fig. 2k,o,p), but different from a posteroventrally divergent ascending process in non-hadrosaurid hadrosauroids57,58,59,63 and a parallel or posteroventrally divergent ascending process in hadrosaurines (e.g., Edmontosaurus annectens, DMNH EPV 127701; Prosaurolophus maximus52) (Table 2). Additionally, the gently curved posterodorsal border of the ascending process (Fig. 1c,f) suggests anterior inclination of the posterior surface of the supraoccipital in articulation, which is a synapomorphic character of hadrosaurids26,35. Therefore, the combination of the four characters mentioned above (the short exoccipital articular surface, well-developed squamosal bosses, posteroventrally convergent ascending process, and anteriorly inclined posterodorsal surface of the ascending process) is unique to Lambeosaurinae (Table 2), suggesting DMNH 2014-12-266 is a supraoccipital of a lambeosaurine hadrosaur.
Isolated lambeosaurine supraoccipitals from the Oldman and Dinosaur Park formations can be divided into two morphotypes by the position of the squamosal bosses. While the squamosal bosses of the first morphotype (UALVP 48, UALVP 53092, and UALVP 53106 from the Oldman Formation and CMN 170 from the Dinosaur Park Formation; Fig. 2k,l,m,n) are posteriorly positioned, those of the other morphotype (UALVP 55300 and UALVP 54569 from the Dinosaur Park Formation; Fig. 2o,p) are anteriorly positioned, which are also seen in the Liscomb lambeosaurine (Fig. 2a). Although the Liscomb lambeosaurine shares this character with UALVP 55300 and UALVP 54569, it differs from UALVP 55300 in having posteriorly extended ascending process (Fig. 2a,f,o,u,p,v). Additionally, the Liscomb lambeosaurine differs from all other lambeosaurine supraoccipitals from the Oldman and the Dinosaur Park formations in having a rugose surface of the ascending process, the laterally completed squamosal bosses, and the ventrally bowed posteroventral margin (Figs 1 and 2). Furthermore, the Liscomb lambeosaurine also differs from penecontemporaneous lambeosaurine Hypacrosaurus altispinus (AMNH FARB 5248) from the Horseshoe Canyon Formation, which has weakly developed ascending process and squamosal bosses that are partly formed by the exoccipital-opisthotic complex60. Comparisons of supraoccipital characters with the Canadian specimens indicate that the Liscomb lambeosaurine is distinct from the Canadian specimens but shows affinities with the supraoccipitals from the Dinosaur Park Formation.
Previous studies suggested presence of lambeosaurine in the Arctic29,64,65 with no definitive descriptions of fossil materials. Russell65, cited by Rich and others66, noted occurrence of lambeosaurine from the Bylot Island of Canada, but details of the record are unknown. Russell64 and Gangloff29 mentioned possible lambeosaurine records from the North Slope of the Alaska, but the identification in the former was based on a personal communication (by John R. Horner) and the latter did not provide a specimen number or the basis for the identification. The Liscomb lambeosaurine is the first definitive occurrence of this group from the Arctic and confirms that lambeosaurines inhabited the ancient Arctic terrestrial environment. This greatly expands the paleogeographic distribution of lambeosaurines much further north than previously known from taxa such as Hypacrosaurus altispinus from southern Alberta, Canada (Fig. 4). At the same time, the morphological affinities with the Canadian lambeosaurines elucidate more extensive faunal exchange between the Arctic and lower paleolatitudes within North America than previously suggested, which is also supported by the presence of Edmontosaurus, Pachyrhinosaurus, tyrannosaurids, and troodontids in both regions4,12,13,15,25,35.

Paleogeographical records records of lambeosaurines during the Late Cretaceous. The red star represents the Liscomb lambeosaurine reported herein. Map is redrawn from Deep Time Maps77. The paleocoordinates are obtained from the Paleobiology Database (www.paleobiodb.org).
The co-occurrence of hadrosaurine and lambeosaurine supraoccipitals from the Liscomb Bonebed suggests that the validity of Ugrunaaluk kuukpikensis should be treated with caution because hadrosaur bones from the bonebed may consist of these hadrosaurid sub-families as well as different ontogenetic stages35 and, more importantly, indicates that hadrosaurine and lambeosaurine dinosaurs co-existed in the Cretaceous Arctic region. The presence of one lambeosaurine supraoccipital and eight previously reported hadrosaurine supraoccipitals15, as well as additional unpublished hadrosaurine specimens in the Perot Museum of Nature and Science collections, suggests numerical dominance of hadrosaurines over lambeosaurines in the ancient Liscomb region. While the hadrosaurine dominance may indicate their better adaptation to Arctic environment than lambeosaurines, hadrosaurine dominance is known from lower latitudes marine deposits67 and regions closer to paleoshorelines of North America68 and eastern Asia34,40,41,42,43,44,69, indicative of near-shore environment preferences by hadrosaurines. Consequently, the hadrosaurine dominant faunal structure of the Liscomb Bonebed, deposited in lower coastal environment, may indicate that Arctic hadrosaurids performed environment preferences similar to those in the lower latitudes (Figs 5 and 6).

Schematic drawing of differential habitat preference between hadrosaurines and lambeosaurines.

Life reconstruction of lambeosaurine-hadrosaurine co-occurrence based on the Liscomb Bonebed hadrosaurids. Artwork by Masato Hattori.
Material and Method
DMNH 2014-12-266, collected from the Liscomb Bonebed and stored in the collection of the Perot Museum of Nature and Science, Dallas, USA, was examined and described herein. Its symmetrical shape and the endocranial wall suggest that the bone is a sagittal endocranial element such as basioccipital, basisphenoid, and supraoccipital. Absences of structures present in basioccipital and basisphenoid (occipital condyle, sphenoccipital tubera, foramina for cranial nerves, basipterygoid process) leaves supraoccipital the only possible candidate. Although multiple large tetrapods are known from the Prince Creek Formation, complete exclusion of supraoccipital from the foramen magnum, suggested by the rugose sutural surface for the exoccipital-opisthotic complex, indicate that the supraoccipital does not belong to basal ornithopod70, dromaeosaurids71, pachycephalosaurines72, troodintids73, or tyranosaurids74. Additionally, the absence of the rostrodorsal process suggest that it does not belong to ceratopsids75. On the other hand, DMNH 2014-12-266 resembles the supraoccipitals of hadrosaurids in complete exclusion from foramen magnum26 and lambeosaurines and non-hadrosaurid hadrosauroids in presence of well-developed squamosal bosses52,53,54,56,57,59. Therefore, DMNH 2014-12-266 is identified as a supraoccipital of hadrosauroid.
Comparisons with isolated supraoccipitals of hadrosaurines from the Liscomb Bonebed (DMNH 22807 and casts of UAMES 4291, UAMES 12727, UAMES 21544, housed at the Canadian Museum of Nature), Prosaurolophus maximus MOR 447-8-8-7-1452, Hypacrosaurus stebingeri USNM 1189354,55, an indeterminate lambeosaurine CMN 017052,53, and non-hadrosaurid hadrosauroids (Bactrosaurus johnsoni56, Batyrosaurus rozhdestvenskyi57, Eolambia caroljonesa58, Eotrachodon orientalis59) were made for taxonomic identification. To further investigate similarities and differences with the late Cretaceous lambeosaurines in Canada, DMNH 2014-12-266 is compared with isolated lambeosaurine supraoccipitals from the Campanian Oldman Formation (UALVP 48, UALVP 53092, UALVP 53106) and the Campanian Dinosaur Park Formation (CMN 0170, UALVP 55300, UALVP 54569). Because Xing and others35 argued that Ugrunaaluk kuukpikensis is a nomen dubium, we conservatively regard the hadrosaurine specimens from the Liscomb Bonebed as Edmontosaurus sp. as they were in prior works5,7,23,29,30,31,32,33,76.
References
Prestrud, P. Adaptation by the Arctic Fox (Alopex lagopus) to the Polar Winter. Arctic 44, 132–138, https://doi.org/10.14430/arctic1529 (1991).
Scholander, P. F., Hock, R., Walters, V. & Irving, L. Adaptation to cold in arctic and tropical mammals and birds in relation to body temperature, insulation, and basal metabolic rate. Biol Bull 99, 259–271, https://doi.org/10.2307/1538742 (1950).
Blix, A. S. Arctic animals and their adaptations to life on the edge. (Tapir Academic Press, 2005).
Fiorillo, A. R. Alaska Dinosaurs: An ancient arctic world. (CRC Press, 2018).
Fiorillo, A. R. Dinosaurs of Alaska: Implications for the Cretaceous origin of Beringia. Geological Society of America Special Papers 442, 313–326, https://doi.org/10.1130/2008.442(15) (2008).
Mull, C. G., Houseknecht, D. W. & Bird, K. J. Revised Cretaceous and Tertiary stratigraphic nomenclature in the Colville Basin, Northern Alaska. Report No. 1673 (2003).
Fiorillo, A. R., McCarthy, P. J. & Flaig, P. P. A multi-disciplinary perspective on habitat preferences among dinosaurs in a Cretaceous Arctic greenhouse world, North Slope, Alaska (Prince Creek Formation: lower Maastrichtian). Palaeogeography, Palaeoclimatology, Palaeoecology 441, 377–389, https://doi.org/10.1016/j.palaeo.2015.07.024 (2016).
Davies, K. L. Duck-bill dinosaurs (Hadrosauridae, Ornithischia) from the North Slope of Alaska. Journal of Paleontology 61, 198–200, https://doi.org/10.1017/s0022336000028341 (1987).
Clemens, W. A. & Nelms, L. G. Paleoecological implications of Alaskan terrestrial vertebrate fauna in latest Cretaceous time at high paleolatitudes. Geology 21, 503, https://doi.org/10.1130/0091-7613 (1993)021<0503:pioatv>2.3.co;2 (1993).
Fiorillo, A. R. On the occurrence of exceptionally large teeth of Troodon (Dinosauria: Saurischia) from the late Cretaceous of Northern Alaska. Palaios 23, 322–328, https://doi.org/10.2110/palo.2007.p07-036r (2008).
Fiorillo, A. R., Tykoski, R. S., Currie, P. J., McCarthy, P. J. & Flaig, P. Description of two partial Troodon braincases from the Prince Creek Formation (Upper Cretaceous), North Slope Alaska. Journal of Vertebrate Paleontology 29, 178–187, https://doi.org/10.1080/02724634.2009.10010370 (2009).
Fiorillo, A. R. & Tykoski, R. S. A new Maastrichtian species of the centrosaurine ceratopsid Pachyrhinosaurus from the North Slope of Alaska. Acta Palaeontologica Polonica 57, 561–573, https://doi.org/10.4202/app.2011.0033 (2012).
Fiorillo, A. R. & Tykoski, R. S. An immature Pachyrhinosaurus perotorum (Dinosauria: Ceratopsidae) nasal reveals unexpected complexity of craniofacial ontogeny and integument in Pachyrhinosaurus. PLoS One 8, e65802, https://doi.org/10.1371/journal.pone.0065802 (2013).
Fiorillo, A. R. & Tykoski, R. S. A diminutive new tyrannosaur from the top of the world. PLoS One 9, e91287, https://doi.org/10.1371/journal.pone.0091287 (2014).
Mori, H., Druckenmiller, P. & Erickson, G. A new Arctic hadrosaurid (Dinosauria: Hadrosauridae) from the Prince Creek Formation (lower Maastrichtian) of northern Alaska. Acta Palaeontologica Polonica 61, 15–32, https://doi.org/10.4202/app.00152.2015 (2016).
Brown, C. M., Druckenmiller, P. & Sues, H.-D. Basal ornithopod (Dinosauria: Ornithischia) teeth from the Prince Creek Formation (early Maastrichtian) of Alaska. Canadian Journal of Earth Sciences 48, 1342–1354, https://doi.org/10.1139/e11-017 (2011).
McKee, E. H., Conrad, J. E. & Turin, B. D. Better dates for Arctic dinosaurs. Eos, Transactions American Geophysical Union 70, 74, https://doi.org/10.1029/89eo00042 (1989).
Conrad, J. E., McKee, E. H. & Turrin, B. D. Age of tephra beds at the Ocean Point dinosaur locality, North Slope, Alaska, based on K-Ar and 40Ar/39Ar analyses. Report No. 1990C (1992).
Flaig, P. P., Fiorillo, A. R. & McCarthy, P. J. Dinosaur-Bearing Hyperconcentrated Flows of Cretaceous Arctic Alaska: Recurring Catastrophic Event Beds on a Distal Paleopolar Coastal Plain. Palaios 29, 594–611, https://doi.org/10.2110/palo.2013.133 (2014).
Brouwers, E. M. et al. Dinosaurs on the North Slope, Alaska: high latitude, latest Cretaceous environments. Science 237, 1608–1610, https://doi.org/10.1126/science.237.4822.1608 (1987).
Flores, R. M. et al. Stratigraphy and facies of Cretaceous Schrader Bluff and Prince Creek Formations in Colville River Bluffs, North Slope, Alaska. Report No. 1748 (2007).
Witte, W. K., Stone, D. B. & Mull, C. In Alaskan North Slope Geology (eds Tailleur, I. & Weimer, P.) 571–579 (Society of Economic Paleontologists and Mineralogists, Pacific Section, 1987).
Gangloff, R. A. & Fiorillo, A. R. Taphonomy and paleoecology of a bonebed from the Prince Creek Formation, North Slope, Alaska. Palaios 25, 299–317, https://doi.org/10.2110/palo.2009.p09-103r (2010).
Fiorillo, A. R., McCarthy, P. J. & Flaig, P. P. Taphonomic and sedimentologic interpretations of the dinosaur-bearing Upper Cretaceous Strata of the Prince Creek Formation, Northern Alaska: Insights from an ancient high-latitude terrestrial ecosystem. Palaeogeography, Palaeoclimatology, Palaeoecology 295, 376–388, https://doi.org/10.1016/j.palaeo.2010.02.029 (2010).
Fiorillo, A. R. & Gangloff, R. A. Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on Arctic Dinosaur paleoecology. Journal of Vertebrate Paleontology 20, 675–682, https://doi.org/10.1671/0272-4634(2000)020[0675:ttftpc]2.0.co;2 (2000).
Horner, J. R., Weishampel, D. B. & Forster, C. A. In The Dinosauria: Second Edition (eds Weishampel, D. B., Dodson, P. & Osmólska, H.) 438–463 (PublisherUniversity of California Press, 2004).
Nelms, L. Late Cretaceous dinosaurs from the North Slope of Alaska. Journal of Vertebrate Paleontology 9, 34, https://doi.org/10.2307/3514763 (1989).
Campione, N. E. & Evans, D. C. Cranial growth and variation in Edmontosaurs (Dinosauria: Hadrosauridae): implications for latest Cretaceous megaherbivore diversity in North America. PLoS One 6, e25186, https://doi.org/10.1371/journal.pone.0025186 (2011).
Gangloff, R. A. Arctic dinosaurs with emphasis on the Cretaceous record of Alaska and the Eurasian-North American connection. Lower and Middle Cretaceous Terrestrial Ecosystems: New Mexico Museum of Natural History and Science Bulletin 14, 211–220 (1998).
Fiorillo, A. R. & Gangloff, R. A. The Caribou Migration Model for Arctic Hadrosaurs (Dinosauria: Ornithischia): A Reassessment. Historical. Biology 15, 323–334, https://doi.org/10.1080/0891296021000037327 (2001).
Chinsamy, A., Thomas, D. B., Tumarkin-Deratzian, A. R. & Fiorillo, A. R. Hadrosaurs were perennial polar residents. The Anatomical Record 295, 610–614, https://doi.org/10.1002/ar.22428 (2012).
Watanabe, A., Erickson, G. M. & Druckenmiller, P. S. An ornithomimosaurian from the Upper Cretaceous Prince Creek Formation of Alaska. Journal of Vertebrate Paleontology 33, 1169–1175, https://doi.org/10.1080/02724634.2013.770750 (2013).
Bell, P. R. & Snively, E. Polar dinosaurs on parade: a review of dinosaur migration. Alcheringa: An Australasian. Journal of Palaeontology 32, 271–284, https://doi.org/10.1080/03115510802096101 (2008).
Xing, H. et al. Comparative Osteology and Phylogenetic Relationship of Edmontosaurus and Shantungosaurus (Dinosauria: Hadrosauridae) from the Upper Cretaceous of North America and East Asia. Acta Geol Sin-Engl 88, 1623–1652, https://doi.org/10.1111/1755-6724.12334 (2014).
Xing, H., Mallon, J. C. & Currie, M. L. Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLoS One 12, e0175253, https://doi.org/10.1371/journal.pone.0175253 (2017).
Fiorillo, A. The vertebrate fauna from the Judith River Formation (Late Cretaceous) of Wheatland and Golden Valley Counties. Montana. The Mosasaur 4, 127–142 (1989).
Prieto-Márquez, A. & Gutarra, S. The ‘duck-billed’ dinosaurs of Careless Creek (Upper Cretaceous of Montana, USA), with comments on hadrosaurid ontogeny. Journal of Paleontology 90, 133–146, https://doi.org/10.1017/jpa.2016.42 (2016).
Fiorillo, A. R. Taphonomy and depositional setting of Careless Creek Quarry (Judith River Formation), Wheatland County, Montana, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 81, 281–311 (1991).
Varricchio, D. J. Taphonomy of Jack’s Birthday Site, a diverse dinosaur bonebed from the Upper Cretaceous Two Medicine Formation of Montana. Palaeogeography, Palaeoclimatology, Palaeoecology 114, 297–323, https://doi.org/10.1016/0031-0182(94)00084-l (1995).
Godefroit, P., Bolotsky, Y. L. & Van Itterbeeck, J. The lambeosaurine dinosaur Amurosaurus riabinini, from the Maastrichtian of Far Eastern Russia. Acta Palaeontologica Polonica 49 (2004).
Bolotsky, Y. L. & Godefroit, P. A new hadrosaurine dinosaur from the Late Cretaceous of Far Eastern Russia. Journal of Vertebrate Paleontology 24, 351–365, https://doi.org/10.1671/1110 (2004).
Godefroit, P., Bolotsky, Y. L. & Lauters, P. A new saurolophine dinosaur from the latest cretaceous of Far Eastern Russia. PLoS One 7, e36849, https://doi.org/10.1371/journal.pone.0036849 (2012).
Godefroit, P., Bolotsky, Y. L. & Bolotsky, I. Y. Osteology and relationships of Olorotitan arharensis, a hollow-crested hadrosaurid dinosaur from the latest Cretaceous of Far Eastern Russia. Acta Palaeontologica Polonica 57, 527–560, https://doi.org/10.4202/app.2011.0051 (2012).
Godefroit Shulin, H., Tingxiang, Y. & Lauters, P. New Hadrosaurid Dinosaurs from the Uppermost Cretaceous of Northeastern China. Acta Palaeontologica Polonica 53, 47–74, https://doi.org/10.4202/app.2008.0103 (2008).
Owen, R. Report on British fossil reptiles, Part 2. Report of the British Association for the Advancement of Science 11, 60–204 (1842).
Seeley, H. G. On the Classification of the Fossil Animals Commonly Named Dinosauria. Proceedings of the Royal Society of London 43, 165–171, https://doi.org/10.1098/rspl.1887.0117 (1887).
Sereno, P. C. Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). National geographic research 2, 234–256 (1986).
Marsh, O. C. Principal characters of American Jurassic dinosaurs, part IV. American Journal of Science 21, 417–423, https://doi.org/10.2475/ajs.s3-21.122.167 (1881).
Dollo, L. Iguanodontidae et Camptonotidae. Comptes Rendus hebdomadaires de l’Academie des Sciences de Paris 106, 775–777 (1888).
Cope, E. D. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the American Philosophical Society 14, 1–252 (1870).
Parks, W. A. Corythosaurus intermedius, a new species of trachodont dinosaur. University of Toronto Studies, Geological Series 15, 1–57 (1923).
Horner, J. R. Cranial morphology of Prosaurolophus (Ornithischia: Hadrosauridae) with descriptions of two new hadrosaurid species and an evaluation of hadrosaurid phylogenetic relationships. Museum of the Rockies Occasional Paper 2, 1–119 (1992).
Langston, W. The vertebrate fauna of the Selma Formation of Alabama: Part VI. The Dinosaurs. Fieldiana: Geology Memoirs 3, 313–361 (1960).
Gilmore, C. W. On the Detailed Skull Structure of a Crested Hadrosaurian Dinosaur. Proceedings of the United States National Museum 84, 481–491 (1937).
Horner, J. R. & Currie, P. J. In Dinosaur eggs and babies (eds Carpenter, Kenneth, Hirrsch, Karl F. & Horner, John R.) Ch. 21, 312–336 (Cambridge University Press, 1994).
Godefroit, P., Dong, Z., Bultynck, P., Li, H. & Feng, L. New Bactrosaurus (Dinosauria: Hadrosauroidea) material from Iren Dabasu (Inner Mongolia, PR China). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terra 68, 3–70 (1998).
Godefroit, P., Escuillié, F., Bolotsky, Y. L. & Lauters, P. In Bernissart dinosaurs and Early Cretaceous terrestrial ecosystems (ed. Godefroit, P.) 335–358 (Indiana University Press, 2012).
McDonald, A. T., Bird, J., Kirkland, J. I. & Dodson, P. Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah. PLoS One 7, e45712, https://doi.org/10.1371/journal.pone.0045712 (2012).
Prieto-Marquez, A., Erickson, G. M. & Ebersole, J. A. Anatomy and osteohistology of the basal hadrosaurid dinosaur Eotrachodon from the uppermost Santonian (Cretaceous) of southern Appalachia. PeerJ 4, e1872, https://doi.org/10.7717/peerj.1872 (2016).
Evans, D. C. Cranial anatomy and systematics of Hypacrosaurus altispinus, and a comparative analysis of skull growth in lambeosaurine hadrosaurids (Dinosauria: Ornithischia). Zoological Journal of the Linnean Society 159, 398–434, https://doi.org/10.1111/j.1096-3642.2009.00611.x (2010).
Godefroit, P., Alifanov, V. & Bolotsky, Y. A re-appraisal of Aralosaurus tuberiferus (Dinosauria, Hadrosauridae) from the Late Cretaceous of Kazakhstan. Bulletin de L’institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 74, 139–154 (2004).
Prieto-Márquez, A., Fondevilla, V., Sellés, A. G., Wagner, J. R. & Galobart, À. Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European archipelago. Cretaceous Research 96, 19–37, https://doi.org/10.1016/j.cretres.2018.12.002 (2019).
Lee, Y. N. et al. Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus. Nature 515, 257–260, https://doi.org/10.1038/nature13874 (2014).
Russell, D. A. In Canada’s missing dimension: science and history in the Canadian Arctic Island Vol. 1 (ed. Harington, C. R.) 81–90 (Canadian Museum of Nature, 1990).
Russell, D. A. A checklist of North American marine Cretaceous vertebrates including fresh water fishes. Occasional Paper of the Tyrrell Museum of Palaeontology 4, 1–58 (1988).
Rich, T. H., Gangloff, R. A. & Hammer, W. R. In Encyclopedia of Dinosaurs (eds Currie, P. J. & Padian, K.) 562–573 (Academic Press, 1997).
Horner, J. R. Upper Cretaceous dinosaurs from the Bearpaw Shale (marine) of south-central Montana with a checklist of Upper Cretaceous dinosaur remains from marine sediments in North America. Journal of Paleontology, 566–577 (1979).
Béland, P. & Russell, D. A. Paleoecology of Dinosaur Provincial Park (Cretaceous), Alberta, interpreted from the distribution of articulated vertebrate remains. Canadian Journal of Earth Sciences 15, 1012–1024, https://doi.org/10.1139/e78-109 (1978).
Godefroit, P., Zan, S. & Liyong, J. The Maastrichtian (Late Cretaceous) lambeosaurine dinosaur Charonosaurus jiayinensis from North-Eastern China. Bulletin de l’Institut Royal des Sciences Naturelles de Belqique, Sciences de la Terre 71, 108–159 (2001).
Norman, D. B., Sues, H. D., Witmer, L. M. & Coria, R. A. In The Dinosauria: Second Edition (eds Weishampel, D. B., Dodson, P. & Osmólska, H.) 393–412 (University of California Press, 2004).
Barsbold, R. & Osmólska, H. The skull of Velociraptor (Theropoda) from the late Cretaceous of Mongolia. Acta Palaeontologica Polonica 44, 189–219 (1999).
Maryańska, T., Chapman, R. E. & Weishampel, D. B. In The Dinosauria: Second Edition (eds Weishampel, D. B., Dodson, P. & Osmólska, H.) 464–477 (University of California Press, 2004).
Currie, P. J. & Zhao, X.-J. A new troodontid (Dinosauria, Theropoda) braincase from the Dinosaur Park Formation (Campanian) of Alberta. Canadian Journal of Earth Sciences 30, 2231–2247, https://doi.org/10.1139/e93-194 (1993).
Currie, P. J. Cranial anatomy of tyrannosaurid dinosaurs from the late Cretaceous of Alberta, Canada. Acta Palaeontologica Polonica 48, 191–226 (2003).
Dodson, P., Forster, C. A. & Sampson, S. D. In The Dinosauria: Second Edition (eds Weishampel, D. B., Dodson, P. & Osmólska, H.) 494–513 (2004).
Flaig, P. P., McCarthy, P. J. & Fiorillo, A. R. In New Frontiers in Paleopedology and Terrestrial Paleoclimatology: Paleosols and Soil Surface Analog Systems (eds Driese, Steven G. & Nordt, Lee C.) (SEPM Society for Sedimentary Geology, 2013).
Blakey, R. Global paleogeography and techtonics in deep time map, http://deeptimemaps.com/ (2018).
Rozhdestvensky, A. Hadrosaurs of Kazakhstan. Upper Paleozoic and Mesozoic Amphibians and Reptiles. Moscow: Akademia Nauk SSSR, 97–141 (1968).
Gates, T. A. et al. Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the late Campanian Cerro del Pueblo Formation, Coahuila, Mexico. Journal of Vertebrate Paleontology 27, 917–930, https://doi.org/10.1671/0272-4634 (2007)27[917:vcanlh]2.0.co;2 (2007).
Gates, T. A., Horner, J. R., Hanna, R. R. & Nelson, C. R. New unadorned hadrosaurine hadrosaurid (Dinosauria, Ornithopoda) from the Campanian of North America. Journal of Vertebrate Paleontology 31, 798–811, https://doi.org/10.1080/02724634.2011.577854 (2011).
Prieto-Márquez, A. The braincase and skull roof of Gryposaurus notabilis (Dinosauria, Hadrosauridae), with a taxonomic revision of the genus. Journal of Vertebrate Paleontology 30, 838–854, https://doi.org/10.1080/02724631003762971 (2010).
Sues, H. D. & Averianov, A. A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs. Proc Biol Sci 276, 2549–2555, https://doi.org/10.1098/rspb.2009.0229 (2009).
Acknowledgements
We thank Kevin Seymour, Margaret Currie, and Clive Coy for their kind help during collection visits for comparisons. We are also grateful to Greg Funston, Kristen MacKenzie, and Jessica Johnson for their kind help on photography. The first author is thankful to Masaya Iijima, Junki Yoshida, and Yoshihiro Tanaka for their comments on earlier versions of the manuscript. The fieldworks were financially supported by the National Office of Polar Programs (OPP 0424594 and OPP 0425636) and National Geographic Society (W221-12). The collection visits are supported by Grant-in-Aid for JSPS Research Fellow Grant Number 17J06410.
Author information
Authors and Affiliations
Contributions
A.R.F. and P.J.M. conducted the field work and collected the material. R.S.T. prepped the specimen. R.T., Y.K. and A.R.F. wrote the main manuscript text. R.T. prepared the figures. Y.K. and A.R.F. supervised the project. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Takasaki, R., Fiorillo, A.R., Kobayashi, Y. et al. The First Definite Lambeosaurine Bone From the Liscomb Bonebed of the Upper Cretaceous Prince Creek Formation, Alaska, United States. Sci Rep 9, 5384 (2019). https://doi.org/10.1038/s41598-019-41325-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-41325-8
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
