
Abstract
The rise of animal life is temporally related to the increased availability of oxygen in the hydrosphere and atmosphere during the Neoproterozoic. However, the earliest metazoans probably needed relatively low oxygen concentrations, suggesting additional environmental and/or biochemical developments were involved. Copper was required in the exploitation of oxygen by the evolving animals, through the development of respiratory proteins and the extracellular matrix required for structural support. We synthesize global data demonstrating a marked enrichment of copper in the Earth’s crust that coincided with the biological use of oxygen, and this new biological use of copper. The copper enrichment was likely recycled into the surface environment by weathering of basalt and other magmatic rocks, at copper liberation rates up to 300 times that of typical granitic terrain. The weathering of basalts also triggered the Sturtian glaciation, which accelerated erosion. We postulate that the coincidence of a high availability of copper, along with increased oxygen levels, for the first time during the Neoproterozoic supported the critical advances of respiration and structural support in evolving animals.
Similar content being viewed by others
Introduction
The precise timing of the first metazoans is uncertain, but the evidence focuses on a time at or shortly after the first of the two major Neoproterozoic glacial intervals (‘Sturtian’) as the time at which metazoans started to appear1,2. This was a time of increasing concentrations of oxygen in the atmosphere, inviting inference of a genetic link between the availability of oxygen and animal evolution3,4,5,6. However, recent assessments of the requirements of the first metazoans conclude that oxygen was only needed at low levels that had already been available for some time5,6,7,8. This implies that a Neoproterozoic oxygenation event was not the sole requirement for the rise of animal life. The use of oxygen required availability of copper9,10,11, which we show was likely being cycled at unprecedented levels in surface environments at that time. Thus, the higher oxygen levels of the late Neoproterozoic were exploited by innovative use of the available copper.
Copper is used by all domains of life12,13,14, and it has been inferred that the availability of copper influenced the timing of the evolution of multicellular life15,16. Proteins evolved to utilize copper in new ways following the oxygenation of the Earth13,17,18. The new copper proteins were, accordingly, used by animals and other eukaryotes (Fig. 1). Copper is used by organisms living in oxygen-rich environments, while most anaerobes do not use it13. There was a marked increase in protein fold domains (biologically functional 3-D structures) related to aerobic metabolism at ~700 Ma17, i.e. at about the time of the Sturtian glaciation, and today up to 160 copper proteins are found in eukaryotes12 Initially, copper proteins helped to bind and neutralize oxygen and negate oxygen toxicity9,10, but copper also conferred benefits to enhance the radiation of metazoans. Cupredoxins catalysed the reduction of oxygen to water in the respiratory chain18,19. The copper-bearing cytochrome c oxidase (COX) is the terminal enzyme in the mitochondrial respiratory chain required for the synthesis of ATP for energy in cells20.Two classes of copper protein are of particular importance to animals in allowing the breathing of oxygen, and the development of body architecture. These are the oxygen-carrying hemocyanin respiratory proteins, and lysyl-oxidases (LOX) whose primary role is in the modelling of extracellular matrix, including the synthesis of collagen and elastin11,21 essential to the transition from unicellular to multicellular organisms, including animals. These copper-dependent proteins diversified in the Neoproterozoic, coincident with the flourishing of animals14,22.

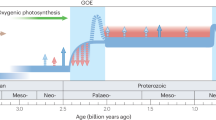
Timeframe for magmatic and sedimentary copper mineralization, enhanced weathering of magmatic rocks, metazoan evolution, and copper protein utilization. Data from refs22,23,29,31,32,40. Oxygenation profile from ref. 4. Copper concentrations also exhibit high levels in Neoproterozoic black shales, shown by Cu/Ti levels47 and copper in pyrite48. First use of copper proteins by metazoans uncertain but at least as old as shown. PAL = Present Atmosphere Level.
Evidence for the antiquity of hemocyanin in molluscs derives from a ~735 Ma molecular clock age23 and an inferred occurrence in the Cambrian Burgess Shale24. This is consistent with a general increase in the biological use of copper at that time15. Sponges are the earliest diverging metazoan group, for which body fossils, molecular clock dates and lipid biomarkers all indicate a record back to the mid-Cryogenian, following the Sturtian glaciation3,25,26. The copper protein hemocyanin was used by the last common metazoan ancestor and accordingly, was used by the earliest sponges27. Sponges were similarly the first organisms in which LOX enzymatic activity may have sculpted extracellular matrix to allow, for example, structural support22. The most abundant protein in animals is collagen, which provides essential mechanical support and was used by the earliest metazoans28,29. A requirement for oxygen in collagen has formerly been viewed as an expression of how animal evolution was dependent upon oxygen30, but recently this constraint has been questioned5. An alternative essential requirement for collagen synthesis is copper11,21, based on the LOX enzymes. Collagen formation may have been a consequence of the detoxification of oxygen31, which copper proteins engendered9,10. The exploitation of these copper-dependent proteins was therefore fundamental to the earliest metazoan physiology.
Whilst copper had always been available, albeit in lower concentrations15, the increased levels of oxygen in combination with anomalous availability of copper allowed a major innovation in how copper was used. There had even been previous episodes of high copper availability32, but before the critical rise in oxygen. This coincidental availability of relatively high levels of both copper and oxygen allowed their integrated use by the new metazoans. Further, the increased oxygen levels also ensured that the copper could be readily recycled at the Earth’s surface. Copper is strongly redox-sensitive, and in oxidizing conditions is mobile33. Thus, the high oxygen facilitated high copper mobility and availability, which in turn could be used by the metazoans to exploit the oxygen. The focus on copper as the element important to the metazoans reflects the critical combination of its abundance in basalts and mobility in oxidizing conditions, which is a combination not shared by other elements.
The Sturtian glaciation at ~700 Ma likely enhanced the availability of copper to a possible role in subsequent evolution. Glacial erosion is especially important as a source of finely ground material with high bioavailability34. The global Sturtian event in particular caused extensive deep erosion of copper-enriched crust, bringing copper to the surface environment just as oxygen levels increased and the first metazoans appeared. The Proterozoic crust had become enriched in copper32 through a succession of Palaeoproterozoic-Mesoproterozoic volcanic massive sulphide (VMS), copper porphyry and other granite-hosted deposits. Then, global Neoproterozoic magmatic activity35 was widely associated with copper mineralization. Volcanic deposits on at least thirteen palaeocontinents host copper mineralization in the 900 to 700 Ma interval, prior to the Sturtian glaciation (Table 1).
Weathering of the Neoproterozoic basalts has been implicated in carbon dioxide drawdown and cooling, which triggered the Sturtian glaciation6,36,37,38. The basalts of this magmatic episode would have been highly weatherable6,36, and the weathering of an atypically large volume of basalt at that time would have liberated anomalous amounts of copper to surface systems. The average basalt has a copper content about seven times that of the average granite (mean values 90 ppm basalt, 13 ppm granite39), which would normally have dominated the detritus from eroding continents. Considering also the greater susceptibility of basalt to weathering, up to twenty times faster than granitic rocks40, the erosion of typical basaltic terrain might increase the flux of copper by two orders of magnitude relative to typical granitic terrain. The contrast would have been even greater in the Cryogenian, when the basalts were conspicuously mineralized by copper. Even unmineralized early Cryogenian basalts contain copper levels higher than normal. The model for accelerated weathering is based on basalts on the Laurentian continent8,38,40, where the Franklin Igneous Province (FIP) covers an area exceeding 2 million km2. Multiple data sets from Alaska to Greenland show mean copper levels in the FIP over twice those of average basalts (Fig. 2, Table 2), and FIP basalts also contain native copper. These statistics combine to indicate the liberation of copper by weathering of the FIP up to 300 times that of granitic terrain, independent of any additional enhancement due to rapid glacial weathering. Evidence from ɛNd values for an enhanced contribution of eroded magmatic rock to marine sediments from about 750 to 600 Ma40 suggests that this potential was realized.

The high flux of copper into the surface/near-surface environment immediately before the Sturtian glaciation is evident from the widespread copper mineralization of clastic sedimentary successions beneath the diamictite. It is very likely that these copper reservoirs were exposed during the extensive (global, long-lived) Cryogenian glaciation, which would have caused further weathering and erosion. Where glacial diamictites are preserved, there is no doubt about the supply of copper. Sturtian diamictites lie unconformably on copper-mineralized rocks in Australia, Africa, North America and Greenland. Clasts of copper ore in the diamictites, in Canada41 and Greenland42, convincingly demonstrate down-cutting into the underlying ores. FIP basalts interfinger with the diamictite in Alaska and contribute clasts to it43. Evidence of copper sulphide replacement of early pyrite in the Central African Copper Belt indicate that copper mineralization was still taking place during burial of the diamictites44. Similarly, copper mineralization in south Australia continued from pre-diamictite volcanic rocks into post-diamictite sediments45, indicating that copper cycling spanned the period of glaciation, and also left ore in periglacial breccias46. The African copper belt deposits, together with early Cryogenian copper deposits in western Canada, constitute most of the known resources of sediment-hosted copper32, implying the availability of exceptional amounts of copper in the upper crust at that time. In summary, a widespread copper-rich substrate to the diamictites, copper-rich basalt interfingering with diamictite, copper-rich detritus in diamictites, and the flush of copper-rich fluids through diamictites combine to indicate an unprecedented flux of copper to the Sturtian surface.
Globally, the enhanced availability of copper in the surface environment is evident from the chemistry of marine organic-rich shales47 and the chemistry of pyrite48 precipitated during the early burial of the shales, which give a measure of seawater composition. Copper levels in Neoproterozoic shales and the diagenetic pyrite are both higher than at any other phase of Earth’s history47,48. This records a higher supply of copper than hitherto, which was extracted from seawater by the precipitation of sulphides under locally anoxic conditions. Following the Neoproterozoic, as the oceans became predominantly oxic, more copper was retained in solution and thereby bioavailable. The surface chemistry in the immediate aftermath of the Sturtian glaciation can be inferred from the earliest diagenetic sulphides in the postglacial succession. The sulphur isotope composition of diagenetic pyrite in the glacial-postglacial succession indicates an origin through (low-temperature) microbial sulphate reduction49. Exceptionally, the postglacial sulphides include copper sulphides, with isotopic compositions similar to those of accompanying pyrite, and also attributable to microbial sulphate reduction49. The formation of discrete copper sulphides is strong evidence for a copper-rich environment during and after the glaciation49.
It is possible that episodes of copper mineralization are missing from the earlier Proterozoic record due to gaps in preservation, but the occurrence of multiple examples of both magmatic and sedimentary copper mineralization in the Neoproterozoic strongly suggests that this was an exceptional period of copper delivery to the upper crust. Basalts in general are commonly mineralized by copper, to the extent that they constitute a recognized ore type50, so the huge FIP represented a major reservoir of copper. The release of copper from basalts was enhanced under oxic conditions51. The anomalous availability of copper during the Sturtian glaciation does not mean that the glaciation was critical to the flux of copper. Rather, the glaciation and release of copper were both products of a single process, i.e. the weathering of the FIP basalts, which consumed carbon dioxide to cause cooling36 and liberated the metal. This is why the sediments below the Sturtian diamictite were already mineralized by copper-rich groundwaters in several parts of the world. The model for weathering-induced cooling emphasizes that weathering was enhanced shortly before the glaciation due to a combination of continental break-up and low latitude position of the continents36,37,40. Break-up triggered magmatism, and an equatorial setting provided a suitable tropical climate in which basalts yield a large proportion (>50%) of their copper52. Thus these factors would have accelerated the flux of copper to surface environments prior to, and during, the Sturtian glaciation.
The high availability of copper was not a short-lived phenomenon. Following the Sturtian glaciation, the record of high copper flux continued in the later Cryogenian and Ediacaran, including for example copper-mineralized flood basalts across Eastern Europe and Ukraine53, and mineralized sediments in North Africa, Siberia and Australia45,54. The continuing record of copper mineralization, and recycling of older Proterozoic copper deposits, in the latter part of the Neoproterozoic, shows that the anomalous supply of copper persisted from the origin of the metazoans through to the Cambrian explosion of life. Following peak delivery of copper to the upper crust, continued access to the copper was facilitated by the higher oxygen content of the atmosphere from the late Neoproterozoic onwards. Increased oxygen allowed greater concentrations of dissolved copper in surface waters and seawater33. Prior to the late Neoproterozoic, when oceans were anoxic, lower levels of dissolved copper limited its potential use. Then in the late Neoproterozoic copper was readily available to support the development of the copper-dependent proteins that facilitated the early evolution and diversification of animals (Fig. 3).

Schematic cycling of copper in Neoproterozoic era. Early Neoproterozoic enrichment of the crust in copper, followed by glacial and other erosion, introduced copper to surface environments where it facilitated use of elevated oxygen by metazoans.
Methods
Copper contents in basalts in the Franklin province of the Laurentian continent were collated from published literature, and summarized as mean values for distinct regions. The composite value for the whole province is a weighted mean from constituent regions. Copper ore grades are most recent estimates available for deposits considered viable for mining, in published literature. Estimates are for either the largest deposit or ore province in each continent.
References
Canfield, D. E., Poulton, S. W. & Narbonne, G. M. Late-Neoproterozoic deep-ocean oxygenation and the rise of animal life. Science 315, 92–95 (2007).
Johnston, D. T. et al. Late Ediacaran redox stability and metazoan evolution. Earth and Planetary Science Letters 335-336, 25–35 (2012).
Peterson, K. J., Cotton, J. A., Gehling, J. G. & Pisani, D. The Ediacaran emergence of bilaterians: Congruence between the genetic and the geological fossil records. Philosophical Transactions of the Royal Society B: Biological Sciences 363, 1435–1443 (2008).
Reinhard, C. T., Planavsky, N. J., Olson, S. L., Lyons, T. W. & Erwin, D. H. Proceedings of the National Academy of Sciences, 113, 8933–8938 (2016).
Mills, D. B. & Canfield, D. E. Oxygen and animal evolution: Did a rise of atmospheric oxygen trigger the origin of animals? Bioessays 36, 1145–1155 (2014).
Lenton, T. M., Boyle, R. A., Poulton, S. W., Shields-Zhou, G. A. & Butterfield, N. J. Co-evolution of eukaryotes and ocean oxygenation in the Neoproterozoic era. Nature Geoscience 7, 257–265 (2014).
Sperling, E. A. et al. Statistical analysis of iron geochemical data suggests limited late Proterozoic oxygenation. Nature 523, 451–454 (2015).
Mills, D. B. et al. Oxygen requirements of the earliest animals. Proceedings of the National Academy of Sciences 111, 4168–4172 (2014).
Van Holde, K. E., Miller, K. I. & Decker, H. Hemocyanins and invertebrate evolution. Journal of Biological Chemistry 276, 15563–15566 (2001).
Jaenicke, E. & Decker, H. Functional changes in the family of Type 3 copper proteins during evolution. Chem Bio Chem 5, 163–169 (2004).
Vest, K. E., Hashemi, H. F. & Cobine, P. A. The copper metallome in eukaryotic cells. In: Banci, L. (ed.) Metallomics and the Cell. Metal Ions in Life Sciences 12, 451–478 (2013).
Andreini, C., Banci, L., Bertinin, I. & Rosato, A. Occurrence of copper proteins through the three domains of life: A bioinformatic approach. Journal of Proteome Research 7, 209–216 (2008).
Ridge, P. G., Zhang, Y. & Gladyshev, V. N. Comparative genomic analyses of copper transporters and cuproproteomes reveal evolutionary dynamics of copper utilization and its link to oxygen. Plos ONE 3, e1378, https://doi.org/10.1371/journal.pone.0001378 (2008).
Aguilera, F., McDougall, C. & Degnan, B. M. Origin, evolution and classification of type-3 copper proteins: lineage-specific gene expansions and losses across the Metazoa. BMC Evolutionary Biology 13, 96 (2013).
Dupont, C. L., Butcher, A., Valas, R. E., Bourne, P. E. & Caetano-Anollés, G. History of biological metal utilization inferred through phylogenomic analysis of protein structures. Proceedings of the National Academy of Sciences 107, 10567–10572 (2010).
Ji, H. F., Chen, L., Jiang, Y. Y. & Zhang, H. Y. Evolutionary formation of new protein folds is linked to metallic cofactor recruitment. BioEssays 31, 975–980 (2009).
Wang, M. et al. A universal molecular clock of protein folds and its power in tracing the early history of aerobic metabolism and planet oxygenation. Molecular Biology and Evolution 28, 567–582 (2011).
Gupta, A. & Lutsenko, S. Evolution of copper transporting ATPases in eukaryotic organisms. Current Genomics 13, 124–133 (2012).
Babcock, G. T. & Wikstrom, M. Oxygen activation and the conservation of energy in cell respiration. Nature 356, 301–309 (1992).
Horn, D. & Barrientos, A. Mitochondrial copper metabolism and delivery to cytochrome c oxidase. IUBMB Life 60, 421–429 (2008).
Gérard, C., Bordeleau, L. J., Barralet, J. & Doillon, C. J. The stimulation of angiogenesis and collagen deposition by copper. Biomaterials 31, 824–831 (2010).
Grau-Bové, X., Ruiz-Trillo, I. & Rodriguez-Pascual, F. Origin and evolution of lysyl oxidases. Scientific Reports, 5, https://doi.org/10.1038/srep10568 (2015).
Lieb, B. & Marml, J. Evolution of molluscan hemocyanins as deduced from DNA sequencing. Micron 35, 117–119 (2004).
Pushie, M. J., Pratt, B. R., Macdonald, T. C., George, G. N. & Pickering, I. J. Evidence for biogenic copper (hemocyanin) in the Middle Cambrian Arthropod Marrella from the Burgess Shale. Palaios 29, 512–524 (2014).
Maloof, A. C. et al. Possible animal-body fossils in pre-Marinoan limestones from South Australia. Nature Geoscience 3, 653–659 (2010).
Love, G. D. et al. Fossil steroids record the appearance of Demospongiae during the Cryogenian period. Nature 457, 718–721 (2009).
Martín-Durán, J. M., de Mendoza, A., Sebé-Pedrós, A., Ruiz-Trillo, I. & Hejnol, A. A broad genomic survey reveals multiple origins and frequent losses in the evolution of respiratory hemerythins and hemocyanins. Genome Biology and Evolution 5, 1435–1442 (2013).
Rodriguez-Pascual, F. & Slatter, D. A. Collagen cross-linking: insights on the evolution of metazoan extracellular matrix. Scientific Reports, 6, https://doi.org/10.1038/srep37374 (2016).
Hynes, R. O. The evolution of metazoan extracellular matrix. Journal of Cell Biology 196, 671–679 (2012).
Towe, K. M. Oxygen-collagen priority and the early metazoan fossil record. Proceedings of the National Academy of Science 65, 781–788 (1970).
Saul, J. M. Did detoxification processes cause complex life to emerge? Lethaia 42, 179–184 (2009).
Hitzman, M. W., Selley, D. & Bull, S. Formation of sedimentary rock-hosted stratiform copper deposits through Earth history. Economic Geology 105, 627–639 (2010).
Brown, A. C. Constraints on conceptual and quantitative modelling of early diagenetic sediment-hosted stratiform copper mineralization. Minerals, 7, https://doi.org/10.3390/min7100192 (2017).
Anderson, S. P. Biogeochemistry of glacial landscape systems. Annual Review of Earth and Planetary Sciences 35, 375–399 (2007).
Li, Z. X. et al. Assembly, configuration, and break-up history of Rodinia: A synthesis. Precambrian Research 160, 179–210 (2008).
Goddéris, Y. et al. The Sturtian ‘snowball’ glaciation: fire and ice. Earth and Planetary Science Letters 211, 1–12 (2003).
Cox, G. M. et al. Continental flood basalt weathering as a trigger for Neoproterozoic Snowball Earth. Earth and Planetary Science Letters 446, 89–99 (2016).
Donnadieu, Y., Goddéris, Y., Ramstein, G., Nédélec, A. & Meert, J. A ‘snowball Earth’ climate triggered by continental break-up through changes in runoff. Nature 428, 303–306 (2004).
Wedepohl, K. H. Handbook of Geochemistry. Springer, Berlin (1978).
Horton, F. Did phosphorus derived from the weathering of large igneous provinces fertilize the Neoproterozoic ocean? Geochemistry, Geophysics, Geosystems 16, 1723–1738 (2015).
Helmstaedt, H., Eisbacher, G. H. & McGregor, J. A. Copper mineralization near an intra-Rapitan unconformity, Nite copper prospect, Mackenzie Mountains, Northwest Territories, Canada. Canadian Journal of Earth Sciences 16, 50–59 (1979).
Harpøth, O., Pedersen, J. L., Schønwandt, H. K. & Thomassen, B. The mineral occurrences of central East Greenland. Meddelelser om Grønland, Geoscience 17, 1–139 (1986).
Macdonald, F. A. The Hula-Hula Diamictite and Katakturuk Dolomite, Arctic Alaska. In: Arnaud, E., Halverson, G. P. & Shields-Zhou, G. (eds) The Geological Record of Neoproterozoic Glaciations. Geological Society, London, Memoirs, 36, 379–387 (2011).
Schmandt, D. et al. The Kamoa copper deposit, Democratic Republic of Congo: Stratigraphy, Diagenetic and Hydrothermal Alteration, and Mineralization. Economic Geology 108, 1301–1324 (2013).
Selley, D. Geological framework and copper mineralisation in South Australia. In: Proterozoic sediment-hosted copper deposits. Centre for Ore Deposit research, University of Tasmania (2000).
Williams, G. E., Schmidt, P. W. & Young, G. M. Strongly seasonal Proterozoic glacial climate in low palaeolatitudes: Radically different climate system on the pre-Ediacaran Earth. Geoscience Frontiers 7, 555–571 (2016).
Fru, E. C. et al. Cu isotopes in marine black shales record the Great Oxidation Event. Proceedings of the National Academy of Sciences 113, 4941–4946 (2016).
Gregory, D. D. et al. Trace element content of sedimentary pyrite in black shales. Economic Geology 110, 1389–1410 (2015).
Parnell, J. & Boyce, A. J. Microbial sulphate reduction during Neoproterozoic glaciation, Port Askaig Formation, UK. Journal of the Geological Society, London, https://doi.org/10.1144/jgs2016-147 (2017).
Cox, D. P. Descriptive model of basaltic Cu. United State Geological Survey Bulletin 1693, 130 (1996).
Neaman, A., Chorover, J. & Brantley, S. L. Implications of the evolution of organic acid moieties for basalt weathering over geological time. American Journal of Science 305, 147–185 (2005).
Liu, S. A. et al. Copper and iron isotope fractionation during weathering and pedogenesis: Insights from saprolite profiles. Geochimica et Cosmochimica Acta 146, 59–75 (2014).
Emetz, A., Piestrzyński, A. & Zagnitko, V. Geological framework of the Volhyn copper fields with a review of the Volhyn flood basalt province (western margin of the East European Craton). Annales Societatis Geologorum Poloniae 74, 257–265 (2004).
Oummouch, A. et al. Geology and metallogenesis of the sediment-hosted Cu-Ag deposit of Tizert (Igherm Inlier, Anti-Atlas Copperbelt, Morocco). Geofluids, https://doi.org/10.1155/2017/7508484 (2017).
Jefferson, C. W., Scoates, R. F. J. & Smith, D. R. Evaluation of the Regional Non-renewable Resource Potential of Banks and Northwestern Victoria Islands, Arctic Canada. Geological Survey of Canada Open File 1695 (1988).
Chen, H. et al. A combined fluid inclusion and Pb isotope study of the Neoproterozoic Pingshui volcanogenic massive sulphide Cu-Zn deposit, Southeast China. Ore Geology Reviews 66, 388–402 (2014).
Chai, G. & Naldrett, A. J. Characteristics of Ni-Cu-PGE mineralization and genesis of the Jinchuan deposit, northwest China. Economic Geology 87, 1475–1495 (1992).
Li, X. H., Su, L., Chung, S. L., Li, Z. X. & Liu, Y. Formation of the Jinchuan ultramafic intrusion and the world’s third largest Ni-Cu sulfide deposit, associated with the 825 Ma south China mantle plume? Geochemistry Geophysics Geosystems 6(11), 1–6 (2005).
Kislov, E. V. Ni-Cu-PGE mineralization in the Upper Proterozoic Ioko-Dovyren mafic-ultramafic massif, Russia. In: Mao, J. W. & Bierlein, F. P. (eds) Mineral Deposit Research: Meeting the Global Challenge. Springer, Berlin, 413–416 (2005).
Knight, J. et al. The Khetri Copper Belt, Rajasthan: Iron oxide copper-gold terrane in the Proterozoic of NW India, in Porter, T. N. (ed.) Hydrothermal Iron Oxide Copper-Gold and Related Deposits: A Global Perspective, volume 2, 321–341. PGC Publishing, Adelaide (2002).
Agar, R. A. The tectono-metallogenic evolution of the Arabian Shield. Precambrian Research 58, 169–194 (1992).
Barrie, C. T., Nielsen, F. W. & Aussant, C. H. The Bisha volcanic-associated massive sulphide deposit, Western Nafka Terrane, Eritrea. Economic Geology 102, 717–738 (2007).
Leblanc, M. & Billaud, P. Zoned and recurrent deposition of Na-Mg-Fe-Si exhalites and Cu-Fe sulphides along synsedimentary faults (Bleida, Morocco). Economic Geology 85, 1759–1769 (1990).
Thomas, R. J. et al. A new lithostratigraphic framework for the Anti-Atlas Orogen, Morocco. Journal of African Earth Sciences 39, 217–226 (2004).
Kamona, A. F. & Günzel, A. Stratigraphy and base metal mineralization in the Otavi Mountain Land, Northern Namibia – a review and regional interpretation. Gondwana Research 11, 396–413 (2007).
Borg, G. Metallogenesis of Neoproterozoic basins in Namibia and Botswana. Communications Geological Survey Namibia 10, 109–119 (1995).
De Oliveira, C. G., De Queiroz, C. L. & Pimentel, M. M. The Arenopólis-Mara Rosa gold-copper belt, Neoproterozoic Goiás magmatic arc. Revista Brasileira Geociências 30, 219–221 (2000).
de Oliveira, C. G., Pimentel, M. M., de Melo, L. V. & Fuck, R. A. The copper-gold and gold deposits of the Neoproterozoic Mara Rosa magmatic arc, central Brazil. Ore Geology Reviews 25, 285–299 (2004).
Sánchez-Bettucci, L. et al. Mineralizations of the Lavalleja Group (Uruguay), a probable Neoproterozoic volcano-sedimentary sequence. Gondwana Research 7, 745–751 (2004).
Drexel, J. F. Review of the Burra Mine Project, 1980–2008 – a progress report. Report, South Australia Department of Mines and Energy (2009).
Cox, G. M. et al. Kikiktat volcanics of Arctic Alaska – Melting of harzburgitic mantle associated with the Franklin large igneous province. Lithosphere 7, 275–295 (2015).
Shellnutt, J. G., Dostal, J. & Keppie, J. D. Petrogenesis of the 723 Ma Coronation sills, Amundsen basin, Arctic Canada: implications for the break-up of Rodinia. Precambrian Research 129, 309–324 (2004).
Williamson, N. M. B., Ootes, L., Rainbird, R. H., Bédard, J. H. & Cousens, B. Initiation and early evolution of the Franklin magmatic event preserved in the 720 Ma Natkusiak Formation, Victoria Island, Canadian Arctic. Bulletin Volcanology, 78, 19, https://doi.org/10.1007/s00445-016-1012-9 (2016).
Jefferson, C. W., Nelson, W. E., Kirkham, R. V., Reedman, J. H. & Scoates, R. F. J. Geology and copper occurrences of the Natkusiak basalts, Victoria Island, District of Franklin. Geological Survey of Canada, Current Research, Part A, 85-1A, 203–214 (1985).
Bédard, J. H. et al. Geochemical database of Franklin sills, Natkusiak Basalts and Shaler Supergroup rocks, Victoria Island, Northwest Territories, and correlatives from Nunavut and the mainland; Geological Survey of Canada, Open File 8009, 1.zip file, https://doi.org/10.4095/297842 (2016).
Kettanah, Y., Zentilli, M., Hanley, J. & Tweedale, F. Geological setting and fluid inclusion characteristics of a lead-copper-barium occurrence hosted in a Neoproterozoic mafic sill at Kiatak, Northumberland Island, Northwestern Greenland. Ore Geology Reviews 79, 268–287 (2016).
Acknowledgements
We thank J. Johnston for skilled technical support. NERC provides funding for the Isotope Community Support Facility. C. Scott and an anonymous reviewer provided invaluable criticism on the manuscript. This work was supported by the NERC under Grant NE/M010953/1.
Author information
Authors and Affiliations
Contributions
J.P. led the project and interpretation. A.B. undertook geochemical analysis and interpretation.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Parnell, J., Boyce, A.J. Neoproterozoic copper cycling, and the rise of metazoans. Sci Rep 9, 3638 (2019). https://doi.org/10.1038/s41598-019-40484-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-40484-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
