
Abstract
The biogeochemistry of hypersaline environments is strongly influenced by changes in biological processes and physicochemical parameters. Although massive evaporation events have occurred repeatedly throughout Earth history, their biogeochemical cycles and global impact remain poorly understood. Here, we provide the first nitrogen isotopic data for nutrients and chloropigments from modern shallow hypersaline environments (solar salterns, Trapani, Italy) and apply the obtained insights to δ15N signatures of the Messinian salinity crisis (MSC) in the late Miocene. Concentrations and δ15N of chlorophyll a, bacteriochlorophyll a, nitrate, and ammonium in benthic microbial mats indicate that inhibition of nitrification suppresses denitrification and anammox, resulting in efficient ammonium recycling within the mats and high primary productivity. We also suggest that the release of 15N-depleted NH3(gas) with increasing salinity enriches ammonium 15N in surface brine (≈34.0‰). Such elevated δ15N is also recorded in geoporphyrins isolated from sediments of the MSC peak (≈20‰), reflecting ammonium supply sufficient for sustaining phototrophic primary production. We propose that efficient nutrient supply combined with frequent bottom-water anoxia and capping of organic-rich sediments by evaporites of the Mediterranean MSC could have contributed to atmospheric CO2 reduction during the late Miocene.
Similar content being viewed by others

Introduction
Present-day and past hypersaline environments are unique and extremely important settings that affect biology, climate, and global geochemistry1,2,3. The biogeochemical cycle of hypersaline environments is highly distinct from that of freshwater and marine conditions because it is strongly affected by changes in the physicochemical parameters of the brine as well as in the state and composition of the biological community in response to salinity variations. In spite of the energy cost to overcome the salt stresses imposed on cellular systems4, hypersaline environments can be highly productive5. This is especially the case for shallow hypersaline environments, where densely populated microbial mats inhabited by a range of physiological groups of microorganisms are formed (e.g., cyanobacteria, phototrophic and chemotrophic sulfur-oxidizing bacteria, and sulfate-reducing bacteria). Constraining the biogeochemical cycle, which is responsible for the diversity of the niche space6 and the primary productivity5,7, is essential for understanding such ecosystems. Moreover, considering that hypersaline environments can become surprisingly productive, the massive evaporation events that occurred repeatedly throughout Earth’s history8 could have had a global impact. However, the biogeochemical cycles of the massive evaporation events remain poorly constrained.
Nitrogen has an active redox cycle comprised mainly of biological processes (e.g., nitrate and ammonium assimilation, N2-fixation, nitrification, denitrification, anaerobic ammonium oxidation [anammox]). It strongly influences photosynthetic primary productivity, and thus the entire biological community, by modifying the amount and chemical species of the bioavailable nitrogenous nutrients9. Because hypersaline microbial mats are often highly productive5, the nitrogen cycle in these systems is expected to operate such that the supply of bioavailable nitrogen is sufficient to maintain active photosynthesis. Various approaches have been applied to study nitrogen cycle, such as in situ measurements, culture and incubation experiments, and genetic analyses; some studies report the importance of N2-fixation10,11, whereas others have demonstrated that ammonium10,12 and dissolved organic nitrogen13 derived from the degradation of organic matter are crucial as nitrogen sources.
The nitrogen isotopic compositions (δ15N) of nitrogenous nutrients and organic compounds enable us to elucidate and quantify the relative importance of the processes controlling the flow of nitrogen14,15. Significantly, the isotopic signatures in modern hypersaline environments can be used for interpreting the δ15N signatures of the analogous ancient environments. In particular, the isotopic compositions of chloropigments (chlorophylls and bacteriochlorophylls) record primary signals of the physiology of phototrophs and the nitrogen substrates that they assimilate16,17,18. Moreover, the preservation of chloropigments as geoporphyrins in sediments on a geological timescale make them an ideal tool for reconstructing past nitrogen cycles19,20,21,22.
Here, we report the nitrogen cycle in a modern shallow hypersaline environment: the solar salterns in Sicily, Italy (Fig. 1A–F). The concentrations and nitrogen isotopic compositions of nitrate, ammonium, and chloropigments allowed us to trace the major processes occurring inside the microbial mats and surface brine. On the basis of insights obtained from modern solar salterns, we sought to reconstruct the nitrogen cycle during the peak of the massive evaporation event in the late Miocene, the Messinian salinity crisis, by measuring the nitrogen isotopic compositions of geoporphyrins from an ancient Sicilian deposit (Fig. 1G,H).

Description of samples analyzed. (A,B) Carbonate ponds are characterized by bottom deposits with a slimy layer several millimeters thick, composed of thin yellow, green, and pink layers on the surface and loose, black deposits underneath. (C,D) Gypsum ponds contain gypsum crusts with transparent yellowish, green, and pink layers (from the surface downward), and loose black deposits buried beneath. (E,F) Halite ponds are characterized by crusts of halite crystals several centimeters thick. (G,H) Halite–mud–anhydrite triplets of Unit C salts in the “Church section” of the Realmonte salt mine, and the rock sample analyzed. Modified from Isaji et al.23.
Distributions and δ15N Signatures of Nitrogenous Compounds
We investigated three commercial solar salterns at Trapani, western Sicily, Italy: the Sosalt (SS), Culcasi (CU), and Chiusicella (CH) salterns (Fig. S1). Three saltern pond types can be defined according to the evaporite minerals that progressively precipitate at the bottom: calcium carbonate (CaCO3) and gypsum (CaSO4·2H2O) ponds, where benthic microbial mats are present, and halite (NaCl) ponds, which lack a benthic ecosystem (Fig. 1A–F). The degree of evaporation of the surface brine in each pond is calculated from the molar concentrations of Mg2+ in the seawater and brine samples as follows (Table S1):
Normalization of the solute concentrations in the surface brine by using DEMg cancelled out the effect of the condensation of solutes during evaporation and thereby allowed us to examine the addition or removal of the solutes (Fig. 2A,B).

Variations in the concentrations and δ15N values of nitrate and ammonium in the Trapani solar salterns. (A) Concentrations of surface-brine nitrate (blue circles) and ammonium (red squares) along the salinity gradient in the samples collected in 2015, (B) surface-brine nitrate and ammonium concentrations normalized to the degree of evaporation (DEMg), (C) δ15N of the surface-brine ammonium, and (D) pH23. (E) Cross plot of δ15N and [NH4+]/DEMg of the surface-brine ammonium. Ponds with similar DEMg values are averaged. Arrow a averages the entire evaporation path from the carbonate to the halite pond, and arrows b to d represent more detailed evaporation paths through each pond type. (F) Changes in the δ15N plotted against normalized changes in concentration of ammonium through the evaporation paths indicated in the arrows in (E). Lines represent expected pathways of isotopic composition assuming the Rayleigh distillation model for the different fractionation factors shown. The ε calculated for arrow d is probably an artifact derived from the variability of the ammonium concentrations in the different salterns.
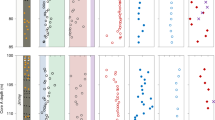
All microbial mats and gypsum crusts investigated had similar distributions of chloropigments, which are mostly derived from benthic rather than planktic phototrophs23. We determined δ15N values for the chloropigments derived from the main phototrophs inhabiting each layer of the microbial mats: chlorophyll a from cyanobacteria, which generally dominate the yellow and green layers, and bacteriochlorophyll a from the purple sulfur bacteria living in the pink layers24. In contrast to the bulk δ15N values that varied over a range from 0.5‰ to 4.8‰, those of chloropigments differed dynamically, with higher values for chlorophyll a (8.8‰ to 19.4‰) than for bacteriochlorophyll a (−8.1‰ to 0.1‰) in all ponds (Fig. 3, Table S2).

Depth profiles of δ15N in the microbial mats in the carbonate ponds (CU-1, -6, -7, -8) and the gypsum crusts in the gypsum ponds (SS-1, CH-1). The δ15N values of cyanobacteria (solid light- and dark-green circles) were calculated from chlorophyll a (open light- and dark-green circles), and those of the purple sulfur bacteria (solid red circles) from bacteriochlorophyll a (open red circles). Light- and dark-blue circles indicate δ15N of nitrate and ammonium, respectively, in the surface brine and porewater. Gray squares indicate δ15N values of total organic nitrogen. Also shown are rough estimates of the thickness of each layer. CU, Culcasi; SS, Sosalt; CH, Chiusicella.
To investigate the physiology of phototrophs and their role in the nitrogen cycle, the δ15N values of phototrophs must be estimated from those of the chloropigments. Laboratory culture experiments and field observations have yielded several relationships between the δ15N of chlorophyll a and cyanobacteria: 15N enrichment of 9.8‰ ± 1.8‰25,26, 15N depletion of 0.9‰ ± 1.3‰25, and 15N depletion of 4.8‰ ± 1.4‰27 in chlorophyll a relative to the cell. Here, we assumed 9.8‰ ± 1.8‰ 15N enrichment in chlorophyll a relative to the cell25, because the δ15N values of cyanobacteria estimated from the other relationships differed unrealistically from those of the bulk deposits (Table S2). On the other hand, the δ15N relationship between purple sulfur bacteria and their bacteriochlorophyll a is considered to be similar to that of the eukaryotic algae and their chlorophyll a25. Thus, we applied an 15N depletion of 4.8‰ ± 1.4‰ in bacteriochlorophyll a relative to the cell20,27. The resulting variations of δ15N values with depth in the phototrophs show similar trends in all ponds, with cyanobacteria in the green layers enriched in 15N relative to purple sulfur bacteria, except for pond CU-1 (Fig. 3).
The concentrations and δ15N patterns of nitrate and ammonium (NH4+ and NH3(aq)) also showed similar depth profiles in the studied ponds. Ammonium was the largest pool of dissolved inorganic nitrogen (334.3–558.7 µmol L−1); its concentrations were more than an order of magnitude higher than those of the surface-brine nitrate, ammonium, and porewater nitrate (Table S1). The porewater ammonium pools had a narrow δ15N range, between 6.5‰ and 7.2‰, whereas ammonium in the surface brine was more enriched in 15N and showed a progressive increase along the salinity gradient, to as high as 34.0‰ in the halite ponds (Fig. 2C). Although the concentrations and δ15N values of nitrate and ammonium in the surface brine varied interannually (Table S2), the similar depth variations of δ15N values of the phototrophs suggest that similar chemical conditions and similar biological processes occurred inside the mats. Variability in the surface brine has also been described previously12 and possibly reflect climate conditions during the previous several days to weeks, such as light availability and rainfall.
Nitrogen dynamics in microbial mats
The similar δ15N values for porewater ammonium and black layer deposits indicate that the ammonium was supplied by anaerobic degradation of organic matter in the black anoxic layer. The general decrease of ammonium concentrations toward the top of the mat10,11 suggests net upward diffusion and consumption of ammonium by processes such as ammonium assimilation, nitrification, or anammox.
The consumption of porewater ammonium by nitrification is of minor importance, considering the low nitrate concentrations (Table S1). Immediate utilization of the produced nitrite or nitrate could keep the concentrations low, but the relatively low δ15N of porewater nitrate suggests that assimilation, denitrification, and anammox, which fractionate against 15N-enriched nitrate14,28, are inactive. Because denitrification and anammox are the major pathways for removing bioavailable nitrogen from the system, their suppression results in accumulation of bioavailable nitrogen within the mat. The cause of the suppression of nitrification is attributable to the extreme energy cost of the hypersaline conditions4, photoinhibition29, or the presence of phototrophic bacteria, which out-compete nitrifying bacteria and archaea for ammonium30.
The δ15N values of the phototrophs indeed support the possibility of ammonium assimilation as a major consumption pathway. The δ15N values of purple sulfur bacteria cover a range where both the porewater nitrate and ammonium could be their nitrogen source. However, the low nitrate concentrations in the porewater, at only about 1/300 of the ammonium concentration (Table S1), as well as a general preference for ammonium over nitrate as a substrate31, suggest ammonium as their major nitrogen source. The assimilation of porewater ammonium by purple sulfur bacteria leaves 15N-enriched ammonium, which diffuses upwards to be assimilated by cyanobacteria in the green and yellow layers, resulting in distinctly higher δ15N values in cyanobacteria than in purple sulfur bacteria (Fig. 4). The lower δ15N of cyanobacteria in the yellowish transparent layer compared with that in the green layer in the gypsum crusts could be attributable to lower primary productivity in the former layer, which has lower chlorophyll a concentrations by at least an order of magnitude.

Proposed nitrogen cycle for the carbonate and gypsum ponds in the Trapani solar salterns. Ammonium (blue circles) produced by the anaerobic degradation of organic matter (grey circles) is assimilated by purple sulfur bacteria (red circles) and cyanobacteria (green and light-green circles) as it diffuses upward, leaving behind substantially 15N-enriched ammonium, of which some part possibly leaks into the surface brine. The increase in salinity causes the release of 15N-depleted NH3(gas) to the atmosphere, thereby increasing the δ15N of the surface-brine ammonium.
Among several mechanisms for ammonium transport through the cell membrane and subsequent biosynthetic pathways, the phototrophs in the mats actively transport NH4+ and assimilate it with glutamine synthase in the range of porewater ammonium concentrations found in this study, with associated nitrogen isotopic fractionation (ε) ranging from −4‰ to −20‰ in laboratory culture experiments under freshwater to marine conditions32,33,34. Our rough estimates of ε for phototrophic assimilation of ammonium inside microbial mats show ε values for purple sulfur bacteria (εPSB) of −10‰ to −5‰ for CU-6, −15‰ to −10‰ for CU-7, and −18‰ to −12‰ for CU-8 (Fig. S2). Although we could not constrain ε for cyanobacteria because of its strong dependence on the assumed εPSB, our results imply that the range of ε for phototrophic assimilation of ammonium under hypersaline conditions is consistent with that reported for freshwater to marine conditions32,33,34.
The δ15N values of chloropigments therefore suggest that the major source of nitrogen for phototrophs is ammonium. However, the large discrepancies of some samples between bulk δ15N values and the estimated cyanobacterial δ15N (SS-1 and CH-1; Table S2) indicate the presence of non-phototrophic biomass depleted in 15N. A possible candidate for this biomass is diazotrophic microorganisms, which have δ15N ranging between −2‰ and 0‰35. This is consistent with previous incubation experiments and genetic analyses indicating N2-fixation in hypersaline microbial mats. In particular, our results are consistent with the suggestion that the activity of non-phototrophic microorganisms such as chemoautotrophic or heterotrophic N2-fixers is important11,36,37. The abundant organic matter produced by photosynthesis probably supports the extreme energy expenditure of these diazotrophs on N2-fixation and adaptation to hypersaline conditions. Thus, the high primary productivity of the hypersaline microbial mats is supported by efficient recycling of ammonium within the mat in the absence of nitrification, denitrification, or anammox, as well as by the supply of new nitrogen from chemoautotrophic and/or heterotrophic diazotrophs.
Ammonium 15N enrichment under evaporative conditions
The δ15N of ammonium in the surface brine increased progressively along the salinity gradient (Fig. 2C). This could be due in part to the leakage of 15N-enriched ammonium from the mat, produced as a result of the assimilation of ammonium by benthic phototrophs. However, considering the net decrease in surface-brine ammonium with increasing salinity ([NH4+/DEMg]; Fig. 2B), there are likely other processes consuming 15N-depleted ammonium. Here, we estimated an apparent nitrogen isotopic fractionation factor assuming that the surface-brine ammonium in all investigated ponds behaved as a continuous system. The overall ε for the entire succession from the carbonate (CU-2) to the halite pond (CU-5) was around −22‰, with ε in the carbonate and gypsum ponds ranging from −5‰ to −20‰ (Fig. 2E,F). Although the extremely high ε calculated for the halite pond is probably an artifact of the variability of the ammonium concentrations in the different salterns, it seems likely that 15N-depleted ammonium was removed even after the saturation point of halite.
We suggest that the δ15N increase in surface-brine ammonium can be ascribed to the release of NH3(gas) to the atmosphere due to the decrease in gas solubility along the salinity gradient. A laboratory study has suggested that the release of NH3(gas) from fluids occurs episodically; between these release episodes the gas stays in the fluids and maintains isotopic equilibrium, with NH3(aq) depleted in 15N relative to NH4+ by 45.4‰ at 23 °C38. Although the pH decline from 8.5 in carbonate ponds to 7.0 in halite ponds (Fig. 2D) is associated with a decrease in the relative proportion of total ammonium as NH3(aq) from around 10% to less than 1%, continuous evaporation, inducing repeated degassing and subsequent dissociation under isotopic equilibrium, could result in a progressive δ15N increase in the remaining ammonium. We expect that 15N-enriched ammonium can be used as an indicator to constrain the nitrogen cycle of hypersaline environments, where ammonium is abundant and partially dissociates to NH3(aq).
Nitrogen cycle during the MSC peak
During the late Miocene (5.97–5.33 Ma), the Mediterranean Sea experienced a massive evaporation event referred to as the Messinian salinity crisis (MSC39,40). Its peak occurred during a short time window (stage 2; 5.60–5.55 Ma40) and was accompanied with the precipitation of huge volumes of primary halite in the deep Mediterranean settings. Here, we measured the δ15N of geoporphyrins purified from the mud–anhydrite layers between the halite layers of the Realmonte salt mine (Sicily, Italy), which were deposited in the Caltanissetta basin during the MSC peak (Fig. 1G,H). These layers are considered to have been deposited under density-stratified conditions during humid seasons of the year, formed as a result of the continental inflow that capped the relatively shallow brine water-body, which precipitated halite during the arid seasons41. The lack of a mat structure in the mud layer suggests that the phototrophs inhabited the water column rather than the bottom deposits.
Two mud–anhydrite-layer samples had identical Fe-porphyrin compositions (Fig. S3). The δ15N values for the combined geoporphyrins from each sample were 17.2‰ and 21.7‰ (Table S2), which we interpret to reflect the assimilation of 15N-enriched ammonium pools. We propose that the 15N-enriched ammonium was produced by mechanisms similar to those discovered for the solar salterns: the suppression of nitrification under extreme salinity and the release of NH3(gas) during the arid season (Fig. 5). The supply of this 15N-enriched ammonium to the less-saline surface layer during the following humid season resulted in 15N-enriched phototrophic biomass. During the humid season, nitrification in the less-saline surface layer and the associated denitrification may have also contributed to increased δ15N of nitrate and ammonium42. However, because subsurface ammonium is the starting material for these processes, its 15N-enrichment during the arid season should have acted to elevate the overall δ15N of the nitrogenous compounds in the system.

Proposed nitrogen cycle during the peak of the Messinian salinity crisis. (A) Nitrogen cycle in the arid season during the deposition of Unit C halite of the Realmonte salt mine. The suppression of nitrification and degassing of 15N-depleted NH3(gas) resulted in the accumulation of 15N-enriched ammonium in the brine. (B) Nitrogen cycle in the humid season during the deposition of the mud layer. 15N-enriched ammonium accumulated in the subsurface was assimilated by phototrophs, resulting in the elevated δ15N of the geoporphyrins.
The first insights into the nitrogen cycle of the MSC provided by our study suggests that ammonium accumulated in the subsurface anoxic brine was supplied to the surface phototrophs under density-stratified conditions at a rate sufficient to sustain phototrophic primary production (Fig. 5). It is reasonable to believe that other essential nutrients, such as phosphate, were also supplied from the subsurface to enhance the phototrophic primary productivity. Importantly, the preservation efficiency of the organic matter produced must have been increased because of the presence of the anoxic bottom brine and the capping of every shale layer by halites. Stratigraphic and geochemical investigations suggest that such an efficient carbon sequestration may actually have been a common feature of the MSC. For example, organic-rich shales, marls, and dolomites have been found widespread and repeatedly throughout the MSC sequence43,44, and there are descriptions of rapidly growing gypsum crystals effectively trapping filamentous organic material45,46. It is noteworthy that about 50% of the world’s largest oilfields are sealed by evaporites, although they constitute less than 2% of the world’s sedimentary rocks47.
The role of the Mediterranean MSC as a carbon sink needs to be explored because it has important implications for the remarkable shifts in the global carbon cycle during the late Miocene. This period is characterized by a decline in the atmospheric CO2 level, which occurred concurrently with intense tectonic activity and expansion of the Antarctic ice sheet48. The major decrease in the CO2 concentration, together with other regional factors, may have promoted substantial and irreversible changes in ecosystems from 7 to 5 Ma, such as the global expansion of C4 plants49,50 and modifications in carbon acquisition strategies of marine algae51. The extensive salt formations and the efficient organic carbon burial could also have contributed to the atmospheric CO2 decrease. Nevertheless, a precise, quantitative evaluation is still difficult at this stage because the MSC intermediate and deep basin deposits remain virtually unexplored. Future drilling of cores from the deep Mediterranean basin (e.g., MEDSALT initiative, https://medsalt.eu) would reveal whether efficient carbon sequestration in marginal basins is a local process or widespread in the Mediterranean, thereby clarifying the impact of the MSC on the global carbon cycle.
Conclusions
Our study highlights the dynamic variations of the nitrogen isotopic signatures in solar salterns, suggesting that the high primary productivity of the hypersaline microbial mats is supported by efficient recycling of ammonium resulting from the suppression of nitrification, denitrification, and anammox. This first report of the highly distinct δ15N signatures of chloropigments in the benthic microbial mats is expected to be important comparison data for elucidating the nitrogen cycle of ancient analogous environments such as the stromatolites, which strongly influenced Earth’s surface biogeochemistry3. Moreover, the substantially high δ15N of ammonium formed during evaporation is useful in clarifying the nitrogen cycle of hypersaline environments: we interpret the extremely high δ15N of geoporphyrins from the MSC peak in the Caltanissetta basin as reflecting a supply of ammonium from the subsurface anoxic brine sufficient to sustain phototrophic primary production. In combination with the frequent formation of bottom water anoxia and capping of organic-rich sediments by evaporites, our first insights into the nitrogen cycle during the MSC imply efficient carbon sequestration, which might have contributed to the decrease in atmospheric CO2 during the late Miocene.
Materials and Methods
Trapani solar salterns
Three commercial solar salterns at Trapani (western Sicily, Italy) were investigated: the Sosalt (SS), Culcasi (CU), and Chiusicella (CH) salterns (Fig. S1). These salterns, each consisting of multiple ponds with different salinities, differ in scale. Sosalt is the largest and Chiusicella is the smallest in both surface area and the number of ponds.
Three pond types can be defined in the solar salterns, according to the evaporite minerals that precipitate at their bottoms: calcium carbonate (CaCO3) ponds, gypsum (CaSO4·2H2O) ponds, and halite (NaCl) ponds. The carbonate ponds (salinity <150), in which calcium carbonate precipitates, are characterized by the formation of a dense benthic microbial mat (Fig. 1A,B) that consists of a slimy layer a few millimeters thick, which is composed of thin yellow, green, and pink layers on the surface, and black, loose deposits buried underneath. In the gypsum ponds (salinity 150–350), a thick layer of selenite gypsum crust grows up from the bottom (Fig. 1C,D), forming layers of different colors (yellowish transparent, green, and pink) from the surface to an average depth of ~5 cm, with loose black deposits below these. Large halite crystals form in the halite ponds (salinity > 350), in which benthic microbial communities are apparently absent (Fig. 1E,F).
Normal seawater, surface brine, porewater, and bottom deposits were collected from the ponds during the daytime in September 2015 and 2016 (Table S1). The surface brine samples and bottom deposits were collected from six carbonate ponds (SS-3, CU-1, -2, -6, -7, and -8), three gypsum ponds (SS-1, SS-2, and CH-1), and two halite ponds (SS-4 and CU-5). The brine samples were immediately passed through 0.45 µm filters to minimalize any biological activity before the nitrate and ammonium concentrations and their nitrogen isotopic compositions were measured. We also collected porewater from three carbonate ponds (CU-6, -7, and -8). Sediment cores, ~50 cm in length, were recovered from each pond, and a Rhizon sampler connected to a 0.45 µm filter was inserted into the black layer for sample collection, at 2.5 cm below the deposit surface in CU-6 and at 5 cm in CU-7 and CU-8. All samples were stored in a freezer until analysis.
Concentrations and nitrogen isotopic compositions of NO3 − and NH4 +
Our experimental procedures largely followed those reported previously52,53. The concentration and nitrogen isotopic composition of NO3− were measured with the microbial denitrifier method54. Pseudomonas chlororaphis subsp. aureofaciens ATCC 13985 T, a denitrifying bacterium lacking the capacity to reduce N2O55, was used to convert NO3− to N2O in a glass vial. The N2O produced was purged from the vial with ultrapure helium and trapped cryogenically with liquid nitrogen (PreCon, Thermo Finnigan, Bremen, Germany). The purged N2O was then introduced into a gas chromatograph (Agilent 6890, Agilent Technologies, Santa Clara, California, USA) equipped with a PoraPLOT column (25 m × 0.32 mm, Agilent Technologies) and a GC III Interface (Thermo Finnigan) to purify the N2O. The nitrogen isotopic composition of the purified N2O was measured with an isotope ratio mass spectrometer (IRMS: Finnigan DELTAplus XP, Thermo Finnigan). The isotopic data was calibrated with three isotopic nitrate standards (USGS32, USGS34, and IAEA-NO3). The isotopic compositions are expressed in conventional δ notation relative to AIR. The analytical precision was within 2.0‰ (±1σ).
We used the diffusion method to recover the total ammonium (NH4+ and NH3(aq)) as (NH4)2SO42− 52,56. The (NH4)2SO42− was then converted to NO3− with the persulfate oxidation method52,57. The nitrogen isotopic composition of the NO3− produced was measured with the microbial denitrifier method. The measured δ15N of N2O was calibrated to that of total ammonium using three ammonium isotopic standards (IAEA-N2, USGS25, and USGS26) that were analyzed concurrently with the samples. The analytical precision was within 1.0‰ (±1σ). The concentration of total ammonium was measured with a fluorometric or colorimetric technique; i.e., the o-phthalaldehyde method58 for the surface brine of CU-6, -7, and -8, and the indophenol blue method59 for all other samples.
Compound-specific nitrogen isotopic compositions of chloropigments
For the bulk δ15N measurements, the microbial mat samples were separated into two parts: the upper slimy layer and the underlying loose black deposits, and the gypsum crusts were separated into four parts: the yellowish transparent, green, and pink gypsum layers, and the underlying loose black deposits. The subsampled deposits were freeze-dried, powdered, and treated with 0.1 M HCl to remove CaCO3 in precleaned smooth-wall tin capsules before analysis. The δ15N measurements were conducted using a modified Flash EA1112 automatic elemental analyzer (EA) connected to a Thermo Finnigan Delta plus XP IRMS via a ConFlo III Interface60. Based on repeated measurements of our laboratory standards, the analytical error was estimated to be within 0.6‰ (±1σ).
The analytical procedure used for the purification of chloropigments followed that described previously23. In brief, the surface deposits of the microbial mats (CU-1, -6, -7, and -8), and the yellowish transparent, green, and pink layers of the gypsum crusts (SS-1 and CH-1) were each freeze-dried and ground to powder. The samples were extracted three times with acetone by sonication, and then extracted with n-hexane. The n-hexane fraction was dried completely under N2 gas and dissolved in 100 µL of N,N-dimethylformamide for HPLC analysis. All procedures were performed in a dark room.
The pigments were isolated and purified with dual-step HPLC using an Agilent Zorbax Eclipse XDB-C18 column (4.6 mm × 250 mm; 5 μm silica particle size). The pigments were eluted isocratically with 75% acetonitrile: pyridine (100:0.5, v/v) and 25% ethyl acetate: pyridine (100:0.5, v/v) for 5 min, followed by a linear gradient of ethyl acetate: pyridine to 50% over 50 min. The flow rate was set to 1 mL min−1 and the column temperature to 30 °C. The collected chlorophyll a and bacteriochlorophyll a were dissolved in 1.5 mL of hexane and reacted with 2 M HCl to convert them to pheophytin a and bacteriopheophytin a, respectively for the second purification step with HPLC. The column used for the second purification step was an Agilent Zorbax Eclipse PAH column (4.6 mm × 250 mm; 5 μm particle size). The pigments were eluted isocratically with 80% acetonitrile: pyridine (100:0.5, v/v) and 20% ethyl acetate: pyridine (100:0.5, v/v) for 5 min, followed by a linear gradient of ethyl acetate: pyridine to 60% over 25 min, and a linear gradient of ethyl acetate: pyridine to 100% over 10 min. The flow rate was set to 1 mL min−1 and the column temperature to 15 °C.
The stable nitrogen isotopic compositions of the chloropigments were measured using a modified EA/IRMS59. The purified chloropigments were dissolved in dichloromethane, transferred to precleaned smooth-wall tin capsules, and dried before analysis. Based on repeated measurements of our laboratory standards, the analytical error was estimated to be within 0.7‰ (±1σ).
Realmonte salt mine
The Realmonte salt mine is composed of a salt unit in the Caltanissetta Basin in Sicily (Fig. S1), deposited during the peak (stage 2) of the MSC at 5.60–5.55 Ma61,62. The salt unit has been divided into four main lithological units from base to top: A–D63,64. Unit C, investigated in this study, was deposited after the basin had undergone desiccation64 and is composed of more than 120 lithological cycles, each one represented by the superposition of three facies: (i) a millimeter-thick mud layer; (ii) a millimeter–centimeter-thick anhydrite (CaSO4) or polyhalite (K2MgCa2(SO4)4·2H2O) layer; and (iii) a decimeter-thick halite layer with white patchy polyhalite grains (Fig. 1G)64,65. The highly pure halite cumulates of skeletal hoppers with chevron overgrowths indicate that the precipitation occurred in a relatively shallow, nonstratified water body, whereas the mud layers are considered to have been deposited under a bottom-anoxic stratified water body41,64. The triplets are interpreted as reflecting annual cyclicity, with anhydrite–halite layers representing the dry season and the mud layers the wet season with increased terrestrial runoff41.
Fallen rocks consisting of a mud–anhydrite layer sandwiched between halite layers (Fig. 1H) were collected at “Church section” (approximately at 10th cycle) and “Rosone section” (towards the bottom of Unit C)41, where Unit C mud–anhydrite–halite triplets are observed. Before the analysis, the halite layer of the rock sample was largely removed with a chisel and hammer. The remaining mud–anhydrite layer of the rock sample was washed in Milli-Q water, methanol, and dichloromethane for 1 min each with sonication to eliminate any possible contamination on the surface of the rock.
Compound-specific isotopic compositions of geoporphyrins
The analytical procedure for the analysis of geoporphyrins was modified from a previously reported procedure66. The geoporphyrins contained in the pulverized sediment samples were extracted with dichloromethane: methanol (7:3, v/v). The extract was then separated with silica gel column chromatography and the low-polarity fraction was collected with dichloromethane. This was further separated into six subfractions with another round of silica gel column chromatography; the reddish-colored band was collected with dichloromethane: methanol (95:5, v/v).
The individual porphyrins were isolated and purified with dual-step HPLC. A reverse-phase HPLC analysis was performed with an Agilent Zorbax SB-C18 column (4.6 mm × 500 mm; 5 μm silica particle size). The porphyrins were eluted isocratically with acetonitrile: N,N-dimethylformamide: pyridine: acetic acid (80:20:0.5:0.5, v/v/v/v) at a flow rate of 1 mL min−1 and a column temperature of 20 °C. For the second purification step, a normal-phase HPLC analysis was conducted with an Agilent Zorbax SIL column (4.6 mm × 500 mm; 5 μm silica particle size). The mobile phase for the isocratic analysis was n-hexane: dichloromethane: N,N-dimethylformamide: pyridine: acetic acid (87:10:3:0.5:0.5, v/v/v/v/v) at a flow rate of 1 mL min−1 and a column temperature of 40 °C
The measurement of the stable nitrogen isotopic compositions of the porphyrins was similar to the measurement of the chloropigments. To obtain enough samples for isotopic measurements, the purified peaks were combined for each sample. Based on repeated measurements of our laboratory standards, the analytical error was estimated to be within 0.5‰ (±1σ).
The metal chelated in the porphyrins was determined with a quadrupole ICP-MS (iCAP Qc, Thermo Scientific, Bremen, Germany) using the metals standard solution VII (Kanto Chemical Co. Inc., Tokyo, Japan). The purified porphyrins were combined and reacted with 3 M HNO3 (high-purity TAMAPURE AA-100 reagents; Tama Chemical, Kanagawa, Japan) for 2 h under 130 °C to release the chelated metal. The relative standard deviations based on the replicate measurements of the sample were within 2.5% for the monitored elements (±2σ).
Data Availability
All the data reported in this article are available from the corresponding author.
References
Sternai, P. et al. Magmatic pulse driven by sea-level changes associated with the Messinian salinity crisis. Nat. Geosci. 10, 783–789 (2017).
Daffonchio, D. et al. Stratified prokaryote network in the oxic–anoxic transition of a deep-sea halocline. Nature 440, 203–207 (2006).
Hoehler, T. M., Bebout, B. M. & Des Marais, D. J. The role of microbial mats in the production of reduced gases on the early Earth. Nature 412, 324–327 (2001).
Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 63, 334–348 (1999).
Canfield, D. E. & Des Marais, D. J. Biogeochemical cycles of carbon, sulfur, and free oxygen in a microbial mat. Geochim. Cosmochim. Acta 57, 3971–3984 (1993).
Ley, R. E. et al. Unexpected diversity and complexity of the Guerrero Negro hypersaline microbial mat. Appl. Environ. Microbiol. 72, 3685–3695 (2006).
Wieland, A., Zopfi, J., Benthien, M. & Kühl, M. Biogeochemistry of an iron-rich hypersaline microbial mat (Camargue, France). Microb. Ecol. 49, 34–49 (2005).
Warren, J. K. Evaporites through time: Tectonic, climatic and eustatic controls in marine and nonmarine deposits. Earth-Sci. Rev. 98, 217–268 (2010).
Capone, D. G., Bronk, D. A., Mulholland, M. R. & Carpenter, E. J. (eds) Nitrogen in the marine environment (Academic Press, New York, 2008).
Bebout, B. M., Paerl, H. W., Bauer, J. E., Canfield, D. E. & Des Marais, D. J. In Microbial Mats: Structure, Development and Environmental Significance, 265–271 (Springer, Berlin, 1994).
Omoregie, E. O., Crumbliss, L. L., Bebout, B. M. & Zehr, J. P. Comparison of diazotroph community structure in Lyngbya sp. and Microcoleus chthonoplastes dominated microbial mats from Guerrero Negro, Baja, Mexico. FEMS Microbiol. Ecol. 47, 305–308 (2004).
Joint, I., Henriksen, P., Garde, K. & Riemann, B. Primary production, nutrient assimilation and microzooplankton grazing along a hypersaline gradient. FEMS Microbiol. Ecol. 39, 245–257 (2002).
Yannarell, A. C. & Paerl, H. W. Effects of salinity and light on organic carbon and nitrogen uptake in a hypersaline microbial mat. FEMS Microbiol. Ecol. 62, 345–353 (2007).
Sigman, D., Karsh, K. & Casciotti, K. In Encyclopedia of Ocean Sciences, 40–54 (Academic Press, New York, 2009).
Ohkouchi, N., Ogawa, N. O., Chikaraishi, Y., Tanaka, H. & Wada, E. Biochemical and physiological bases for the use of carbon and nitrogen isotopes in environmental and ecological studies. Prog. Earth Planet. Sci. 2, 1 (2015).
Ohkouchi, N. et al. Biogeochemical processes in the saline meromictic Lake Kaiike, Japan: implications from molecular isotopic evidences of photosynthetic pigments. Environ. Microbiol. 7, (1009–1016 (2005).
Higgins, M. B., Robinson, R. S., Carter, S. J. & Pearson, A. Evidence from chlorin nitrogen isotopes for alternating nutrient regimes in the Eastern Mediterranean Sea. Earth. Planet. Sci. Lett. 290, 102–107 (2010).
Isaji, Y. et al. Varying responses to Indian monsoons during the past 220 kyr recorded in deep‐sea sediments in inner and outer regions of the Gulf of Aden. J. Geophys. Res. Oceans 120, 7253–7270 (2015).
Chicarelli, M. I., Hayes, J. M., Popp, B. N., Eckardt, C. B. & Maxwell, J. R. Carbon and nitrogen isotopic compositions of alkyl porphyrins from the Triassic Serpiano oil shale. Geochim. Cosmochim. Acta 57, 1307–1311 (1993).
Ohkouchi, N., Kashiyama, Y., Kuroda, J., Ogawa, N. O. & Kitazato, H. The importance of diazotrophic cyanobacteria as primary producers during Cretaceous Oceanic Anoxic Event 2. Biogeosciences 3, 467–478 (2006).
Junium, C. K., Freeman, K. H. & Arthur, M. A. Controls on the stratigraphic distribution and nitrogen isotopic composition of zinc, vanadyl and free base porphyrins through Oceanic Anoxic Event 2 at Demerara Rise. Org. Geochem. 80, 60–71 (2015).
Higgins, M. B., Robinson, R. S., Husson, J. M., Carter, S. J. & Pearson, A. Dominant eukaryotic export production during ocean anoxic events reflects the importance of recycled NH4 +. Proc. Natl. Acad. Sci. USA 109, 2269–2274 (2012).
Isaji, Y. et al. Biological and physical modification of carbonate system parameters along the salinity gradient in shallow hypersaline solar salterns in Trapani, Italy. Geochim. Cosmochim. Acta 208, 354–367 (2017).
Oren, A. Diversity of halophilic microorganisms: environments, phylogeny, physiology, and applications. J. Ind. Microbiol. Biotechnol. 28, 56–63 (2002).
Higgins, M. B. et al. Paleoenvironmental implications of taxonomic variation among δ15N values of chloropigments. Geochim. Cosmochim. Acta 75, 7351–7363 (2011).
Beaumont, V. I., Jahnke, L. L. & Des Marais, D. J. Nitrogen isotopic fractionation in the synthesis of photosynthetic pigments in Rhodobacter capsulatus and Anabaena cylindrica. Org. Geochem. 31, 1075–1085 (2000).
Sachs, J. P., Repeta, D. J. & Goericke, R. Nitrogen and carbon isotopic ratios of chlorophyll from marine phytoplankton. Geochim. Cosmochim. Acta 63, 1431–1441 (1999).
Brunner, B. et al. Nitrogen isotope effects induced by anammox bacteria. Proc. Natl. Acad. Sci. USA 110, 18994–18999 (2013).
Guerrero, M. A. & Jones, R. D. Photoinhibition of marine nitrifying bacteria. I. Wavelength-dependent response. Mar. Ecol. Prog. Ser., 183–192 (1996).
Smith, J. M., Chavez, F. P. & Francis, C. A. Ammonium uptake by phytoplankton regulates nitrification in the sunlit ocean. PLoS One 9, e108173 (2014).
Zehr, J. P. & Ward, B. B. Nitrogen cycling in the ocean: new perspectives on processes and paradigms. Appl. Environ. Microbiol. 68, 1015–1024 (2002).
Hoch, M. P., Fogel, M. L. & Kirchman, D. L. Isotope fractionation associated with ammonium uptake by a marine bacterium. Limnol. Oceanogr. 37, 1447–1459 (1992).
Vo, J., Inwood, W., Hayes, J. M. & Kustu, S. Mechanism for nitrogen isotope fractionation during ammonium assimilation by Escherichia coli K12. Proc. Natl. Acad. Sci. USA 110, 8696–8701 (2013).
Liu, K.-K. et al. Concentration dependent nitrogen isotope fractionation during ammonium uptake by phytoplankton under an algal bloom condition in the Danshuei estuary, northern Taiwan. Mar. Chem. 157, 242–252 (2013).
Wada, E. Nitrogen isotope fractionation and its significance in biogeochemical processes occurring in marine environments. Isotope marine chemistry, 375–398 (1980).
Pinckney, J. L. & Paerl, H. W. Anoxygenic photosynthesis and nitrogen fixation by a microbial mat community in a bahamian hypersaline lagoon. Appl. Environ. Microbiol. 63, 420–426 (1997).
de Wit, R., I. Falcón, L. & Charpy-Roubaud, C. Heterotrophic dinitrogen fixation (acetylene reduction) in phosphate-fertilised Microcoleus chthonoplastes microbial mat from the hypersaline inland lake ‘la Salada de Chiprana’ (NE Spain). Hydrobiologia 534, 245–253 (2005).
Li, L., Lollar, B. S., Li, H., Wortmann, U. G. & Lacrampe-Couloume, G. Ammonium stability and nitrogen isotope fractionations for–NH3(aq)–NH3(gas) systems at 20–70 °C and pH of 2–13: Applications to habitability and nitrogen cycling in low-temperature hydrothermal systems. Geochim. Cosmochim. Acta 84, 280–296 (2012).
Hsü, K., Ryan, W. & Cita, M. Late Miocene desiccation of the Mediterranean. Nature 242, 240–244 (1973).
Roveri, M. et al. The Messinian Salinity Crisis: Past and future of a great challenge for marine sciences. Mar. Geol. 352, 25–58 (2014).
Manzi, V. et al. High-frequency cyclicity in the Mediterranean Messinian evaporites: Evidence for solar-lunar climate forcing. J. Sed. Res. 82, 991–1005 (2012).
Collister, J. W. & Hayes, J. M. In Geochemical, Biogeochemical, and Sedimentological Studies of the Green River Formation, Wyoming, Utah and Colorado., C1–C16 (U.S. Geological Survey Bulletin, Washington, 1991).
Lugli, S., Manzi, V., Roveri, M. & Schreiber, B. C. The Primary Lower Gypsum in the Mediterranean: A new facies interpretation for the first stage of the Messinian salinity crisis. Palaeogeogr., Palaeoclimatol., Palaeoecol. 297, 83–99 (2010).
Natalicchio, M. et al. An archaeal biomarker record of paleoenvironmental change across the onset of the Messinian salinity crisis in the absence of evaporites (Piedmont Basin, Italy). Org. Geochem. 113, 242–253 (2017).
Dela Pierre, F. et al. Are the large filamentous microfossils preserved in Messinian gypsum colorless sulfide-oxidizing bacteria? Geology 43, 855–858 (2015).
Panieri, G. et al. Ribosomal RNA gene fragments from fossilized cyanobacteria identified in primary gypsum from the late Miocene, Italy. Geobiology 8, 101–111 (2010).
Warren, J. K. Evaporites: A geological compendium (Springer, New York, 2016).
Ohneiser, C. et al. Antarctic glacio-eustatic contributions to late Miocene Mediterranean desiccation and reflooding. Nat. Commun 6, 8765 (2015).
Cerling, T. E. et al. Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153–158 (1997).
Herbert, T. D. et al. Late Miocene global cooling and the rise of modern ecosystems. Nat. Geosci. 9, 843–849 (2016).
Bolton, C. T. & Stoll, H. M. Late Miocene threshold response of marine algae to carbon dioxide limitation. Nature 500, 558–562 (2013).
Isobe, K. et al. Analytical techniques for quantifying 15N/14N of nitrate, nitrite, total dissolved nitrogen and ammonium in environmental samples using a gas chromatograph equipped with a quadrupole mass spectrometer. Microbes Environ. 26, 46–53 (2011).
Nishizawa, M. et al. Nitrification-driven forms of nitrogen metabolism in microbial mat communities thriving along an ammonium-enriched subsurface geothermal stream. Geochim. Cosmochim. Acta 113, 152–173 (2013).
Sigman, D. M. et al. A bacterial method for the nitrogen isotopic analysis of nitrate in seawater and freshwater. Anal. Chem. 73, 4145–4153 (2001).
Casciotti, K. L., Sigman, D. M., Hastings, M. G., Böhlke, J. & Hilkert, A. Measurement of the oxygen isotopic composition of nitrate in seawater and freshwater using the denitrifier method. Anal. Chem 74, 4905–4912 (2002).
Holmes, R. M., McClelland, J. W., Sigman, D. M., Fry, B. & Peterson, B. J. Measuring 15N–NH4 + in marine, estuarine and fresh waters: An adaptation of the ammonia diffusion method for samples with low ammonium concentrations. Mar. Chem. 60, 235–243 (1998).
Stark, J. M. & Hart, S. C. Diffusion technique for preparing salt solutions, Kjeldahl digests, and persulfate digests for nitrogen-15 analysis. Soil Sci. Soc. Am. J. 60, 1846–1855 (1996).
Holmes, R. M., Aminot, A., Kérouel, R., Hooker, B. A. & Peterson, B. J. A simple and precise method for measuring ammonium in marine and freshwater ecosystems. Can. J. Fish. Aquat. Sci. 56, 1801–1808 (1999).
Solorzano, L. Determination of ammonia in natural waters by the phenolhypochlorite method. Limnol. Oceanogr. 14, 799–801 (1969).
Ogawa, N. O., Nagata, T., Kitazato, H. & Ohkouchi, N. In Earth, Life, and Isotopes, 339–353 (Kyoto University Press, Kyoto, 2010).
Manzi, V., Lugli, S., Roveri, M. & Schreiber, B. C. A new facies model for the Upper Gypsum of Sicily (Italy): chronological and palaeoenvironmental constraints for the Messinian salinity crisis in the Mediterranean. Sedimentology 56, 1937–1960 (2009).
Krijgsman, W., Hilgen, F. J., Raffi, I., Sierro, F. J. & Wilson, D. S. Chronology, causes and progression of the Messinian salinity crisis. Nature 400, 652–655 (1999).
Decima, A. & Wezel, F. C. Late Miocene evaporites of the central Sicilian basin, Italy. Initial Reports of the Deep Sea Drilling Project 13, 1234–1241 (1973).
Lugli, S., Schreiber, B. C. & Triberti, B. Giant polygons in the Realmonte mine (Agrigento, Sicily): evidence for the desiccation of a Messinian halite basin. J. Sed. Res. 69, 764–771 (1999).
Yoshimura, T. et al. An X-ray spectroscopic perspective on Messinian evaporite from Sicily: Sedimentary fabrics, element distributions, and chemical environments of S and Mg. Geochem. Geophys. Geosyst. 17, 1383–1400 (2016).
Kashiyama, Y., Kitazato, H. & Ohkouchi, N. An improved method for isolation and purification of sedimentary porphyrins by high-performance liquid chromatography for compound-specific isotopic analysis. J. Chromatogr. A 1138, 73–83 (2007).
Acknowledgements
We are grateful to SoSalt Spa and R.N.O Saline di Trapani e Paceco for permission to sample the Trapani solar works. We are also grateful to A. Suzuki of AIST for the measurements of salinity. We thank H. Suga of JAMSTEC for assistance with laboratory work. This study was partly supported by a Japan Society for the Promotion of Science (JSPS) Research Fellowship (16J07844) to YI, Grants-in-Aid (16H02236) and the JAMSTEC President Fund to NO.
Author information
Authors and Affiliations
Contributions
Y.I., H.K., J.K., T.Y. and N.O. designed the study. Y.I., H.K., J.K., T.Y., F.J.J.E., S.L., A.S., V.M. and M.R. collected the samples. Y.I., N.O.O., T.Y., A.M. and T.S. conducted the experimental work and data analysis. Y.I. wrote the paper with contributions from N.O. and F.J.J.E. and all authors edited and revised the paper.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Isaji, Y., Kawahata, H., Ogawa, N.O. et al. Efficient recycling of nutrients in modern and past hypersaline environments. Sci Rep 9, 3718 (2019). https://doi.org/10.1038/s41598-019-40174-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-40174-9
This article is cited by
-
Lake desiccation drives carbon and nitrogen biogeochemistry of a sub-tropical hypersaline lake
Hydrobiologia (2023)
-
Integrated isotopic and organic geochemical constraints on the depositional controls and source rock quality of the Neogene Kalamaki sedimentary successions (Zakynthos Island, Ionian Sea)
Mediterranean Geoscience Reviews (2021)
-
Nutrient dynamics and budgeting in a semi-enclosed coastal hypersaline lagoon
Environmental Science and Pollution Research (2021)
-
In silico determination of nitrogen metabolism in microbes from extreme conditions using metagenomics
Archives of Microbiology (2021)
-
Depthwise microbiome and isotopic profiling of a moderately saline microbial mat in a solar saltern
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
