
Abstract
Rhizoctonia solani Kühn (teleomorph = Thanatephorus cucumeris (Frank) Donk) is one of the important soil-borne fungal pathogen, which infects tomato with typical symptoms of seedling damping-off and foot rot. During surveys (2014 and 2015 crop season) of nine tomato growing areas in Pothohar region of Pakistan, symptoms of foot rot were noted on approximately 33.4% of the plants observed at soil line level of the stem. Lesions on infected plant stems were irregular in shape, water-soaked, brown in colour manifesting sunken appearance. Fungal colonies isolated from stem portions of the diseased plants on malt extract agar medium were light grey to brown in colour with abundant mycelial growth and branched hyphae. A septum was always present in the branch of hyphae near the originating point with a slight constriction at the branch. No conidia or conidiophores were observed. All isolates were multinucleate when subjected to DAPI (4′,6-diamidino-2-phenylindole) stain. Based on morphological characteristics of fungal hyphae, isolates were identified as R. solani. Restriction analysis of PCR-amplified ribosomal DNA with four discriminant enzymes (MseI, AvaII, HincII, and MunI) and hyphal interactions with known tester strains confirmed these isolates belong to AG-3-PT (64.2%), AG-2-1 (14.2%), AG-2-2 (9.5%), AG-5 (7.1%) and AG-4-HGI (4.7%). AG-3-PT was widely distributed to major tomato growing areas while other groups were confined to distinct locations. Internal transcribed spacer (ITS) region of rDNA was amplified with the primers ITS1/ITS4 and sequenced which had 99–100% identity with the corresponding gene sequences of respective R. solani AGs. To confirm Koch’s postulates, four week old tomato plants were transplanted into 1.5 L plastic pots containing sterilized potting mixture i.e. sand: clay: farmyard manure, at the rate of 1:1:1. Soil inoculum containing 10 g of barley grains colonized with each isolate of R. solani for 14 days was mixed in the upper 2 cm layer of soil (Taheri and Tarighi, 2012). A set of uninoculated plants was used as a control. Ambient conditions were provided under the greenhouse. 21 days after inoculation, water-soaked greyish to brown lesions similar to the symptoms of the previous infection were observed on stem portions of all inoculated plants while control plants remained symptomless. Fungus re-isolated from infections was confirmed as R. solani by microscopic appearance of the hyphae. Present study is the first report of AG composition of R. solani infecting tomato in Pakistan which will be useful to breeding programs working on varietal evaluation.
Similar content being viewed by others

Introduction
Solanum lycopersicum L., formerly known as Lycopersicon esculentum Mill., is one of the dominant vegetable crop worldwide that is generally cultivated in warm or tropical climate. It is an adaptable crop used for both fresh market and processing in prepared foods as canned, ketchup, sauce, juice, paste, powder, puree, salad dressings, soups, vegetable and juice cocktails, frozen tomatoes, preserved or dried foods. It is now considered to be a part of the daily diet1. Tomato is the second most consumed vegetable after potato2. In Pakistan tomato is grown on an area of 62536 hectares with a production of about 587111 tons and yield 10.5 tons/ha (FAO, 2017). The Pothohar region contributes 7.58% of the total tomato production of the country. This local tomato yield is low as compared to other countries of the world including USA (97.9 tons/ha), China (58 tons/ha) and India (23.4 tons/ha)3.
Several biotic and abiotic factors contribute to this low tomato yield. Of all primary food crops, tomato crop also endures the utmost yield losses and a significant part of the produce is lost due to disease attack and pests4,5,6. Among the biological factors causing disease in tomato crop, fungal pathogens especially R. solani is the worst damaging that play a key role in reducing the yield7,8,9.
R. solani has a wide host range of more than 200 plant species especially Solanaceae family including eggplant, pepper, potato, tobacco and tomato cultivated under both, greenhouse and field conditions10,11,12. R. solani is a species complex of several anastomosis groups (AGs) based on the hyphal fusion of isolates which differ in genotypic and phenotypic characters13. To date, thirteen AGs designated as AG1–AG13 and AG-BI have been assigned on the basis of hyphal anastomosis interactions14,15. Although AGs of R. solani are identified on the basis of hyphal anastomosis reactions, however reproducibility of these interactions needs experiences, is a time consuming process and can be affected by factors including laboratory environment, nutritional conditions and genetic stability14,16. Molecular approaches including DNA based sequence homology, restriction analysis of ribosomal DNA have been confirmed as reliable tools to differentiate isolates of R. solani into distinct clades corresponding to different AGs and subgroups17,18. Different anastomosis groups cause infection on differential hosts. AG-3 is most widely distributed in Pakistan which causes black scurf on potato19 however, it has also been reported to infect other solanaceous vegetable crops20,21,22. Other AGs including AG-2-122, AG-2-223, AG-424 and AG-525 have also been reported to infect tomato. Although, foot rot of tomato caused by R. solani has already been reported from Pakistan however, AG group composition of R. solani responsible for this infection is not determined. The objective of the present study is to determine anastomosis groups (AGs) of R. solani causing foot rot of tomato using tester isolates and restriction analysis of ribosomal DNA.
Materials and Methods
Sampling and isolation of Rhizoctonia solani
Surveillance of different tomato production areas of Pothohar region which includes districts; Jhelum, Chakwal, Attock, Rawalpindi, and Islamabad Capital Territory was done to collect diseased plants showing typical symptoms of foot rot and damping off. Pothohar region is situated between latitude 32.5°00′N to 34°00′N and altitude 72°00′E to 74°00′E in the Asian sub-continent with an elevation of 517 m above sea level and experiences semi-arid to humid climate26. Tomato is grown in scattered locations in the selected districts so purposive sampling was done. Disease prevalence and incidence percentage was calculated using formula;
A total of 117 symptomatic plant samples were collected. Sections of the stem (ca. 5 mm cube) were surface sterilized with 1% sodium hypochlorite solution, washed twice in tap water and blotted on sterilized filter paper were placed on 9 cm diameter Petri-plates of PDA incubated at 25 °C for 4 days. Hyphae resembling Rhizoctonia27 were identified under a microscope and pure cultures were obtained using the hyphal tipping technique. All isolates were maintained on Malt Extract Agar (MEA) and were preserved on barley grains by the method described by28.
Cultural Characteristics and Microscopic studies of R. solani
Recovered isolates were initially identified as Rhizoctonia by culture characteristics on MEA as described by Sneh, et al.29. Hyphae of Rhizoctonia have right-angle branches, branches at the distal septae of cells and dolipore septa. Isolates were grown in 9 cm Petri plates containing 2% water agar (WA) at 25 °C for 4 days stained with 0.05% lactophenol blue and examined under a microscope to observe hyphal morphology. A number of nuclei per cell of R. solani were counted by staining hyphae with 1ug/ml of 4′-6 diamidino-2-phenylindole (DAPI stain). Petri plates were examined under a fluorescent microscope at 400X magnification to count a number of nuclei per cell.
Pathogenicity Testing
Pathogenicity tests were performed according to the method described by Misawa and Kuninaga22. Plastic cell trays (53.49 cm L × 26.82 cm W) having 32 cells/ tray were filled with sterilized potting mixture i.e. sand: clay: farmyard manure at the rate of 1:1:130. Four week old plants cv. Rio Grande were transplanted into the cells. The inoculum was prepared by colonizing isolates of R. solani on barley grains for 14 days. 10 g of ground barley grains colonized with each isolate of R. solani was mixed in the upper 2 cm layer of soil. Cells colonized only with ground barley grains were used as a control. Plants were grown at 25 ± 2 °C for 28 days. Infection on soil line level of the stem was categorized as −, no symptom; ±, brown lesion on part of the stem; +, brown lesion girdled the stem; ++, brown lesion girdled the stem and plants wilted. The trial was conducted with three replicates for 67 recovered isolates arranged in completely randomized design (CRD). The whole experiment was repeated twice.
Anastomosis group typing
Generally, isolates of R. solani are identified based on hyphal interaction reactions, however, resolution of this method at subgroup level is insufficient31,32,33. Recovered isolates were subjected to Restriction fragment length polymorphism (RFLP) analysis of ribosomal DNA (rDNA) sequences. The results of AG group composition were further confirmed by hyphal anastomosis interactions.
PCR–RFLP analysis
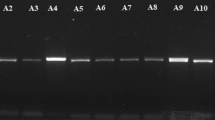
Isolates were maintained on malt extract broth (MEB) medium in 9 cm Petri plates incubated at 25 °C for 5 days. Mycelium mat for each isolate was harvested blot dried, lyophilized and ground to fine powder. DNA from each isolate was extracted using the standard protocol of Omniprep for fungi extraction kit (G-Biosciences) (Cat. # 786-399) and was subjected to PCR amplification of the ITS region with primers RS1 (5′-CCTGTGCACCTGTGAGACAG-3′) and RS4 (5′-TGTCCAAGTCAATGGACTAT-3′)34. PCR reaction mixture was prepared by adding 10 µL 5X Buffer, 1 µL dNTPs, 2.5 µL each forward and reverse primers, 1 µL MgCl2, 0.5 µL Taq Polymerase 2.5 µL DNA template in the final volume of 50 µL. A negative control (without DNA template) was always included in PCR reactions. Amplifications were performed in MJ Research Tetrad PTC-225 Thermal Cycler system (Bunker Lake Blvd. Ramsey, Minnesota, USA) with an initial denaturation at 94 °C for 2 min followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for ITS for 30 s, extension at 72 °C for 1 min, and a final extension at 72 °C for 5 min. Aliquots of the PCR products were analyzed in 2% agarose gel stained with ethidium bromide and viewed using a UV transilluminator. Each PCR product was cleaned up using Sephadex G-50 and amplifications were further characterized using discriminating enzymes; MseI, AvaII + HincII and MunI as determined by Guillemaut, et al.35 (Fig. 1).

Restriction patterns revealed by RFLP analysis of internal transcribed spacers sequences of Rhizoctonia solani with discriminating enzymes35.
Anastomosis interactions
Isolates were paired with tester strains of the respective AGs identified by PCR-RFLP. An agar disc (5 mm) was excised from the edge of the actively growing hyphae and placed on 1.5% WA coated clean glass slides having similar agar disc of tester strain of the known AG. After 48–72 hours when hyphae from each isolate overlapped, slides were stained with lactofuchsin and were examined under 400x magnification for hyphal anastomosis. Anastomosis reactions were classified from C0 to C3 where, C0 = no reaction, C1 = contact fusion, C2 = somatic fusion or perfect anastomosis and C3 = auto-anastomosis as described by Carling36. C3 type interactions; auto-anastomosis or self-pairing were used as positive control.
Twenty random locations on each glass slide were selected to observe hyphal interactions between unknown strain and the tester strain of respective AG and percentage fusion frequency (% FF) was determined as;
A = Sum of fusion locations (in C1, C2, C3) in 20 microscopic fields
B = Sum of contact points in 20 microscopic fields
Isolates pairing at more than 80% locations were confirmed as belonging to respective anastomosis group.
Sequencing of ITS-5.8S rDNA and phylogeny
Molecular identification of the type isolates belonging to different AGs was accomplished by amplification of their ITS region using universal sense ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) encoding ITS-1-5.8S-ITS-239. PCR mixtures were prepared in a total volume of 50 µL containing 2.5 µL of the total DNA, 2.5 µL each forward and reverse primers, 10 µL 5X Buffer, 1 µL MgCl2, 1 µL dNTPs, and 0.5 µL of Taq DNA polymerase. A negative control (without DNA template) was always included in PCR reactions. PCR conditions were same as described in the PCR-RFLP analysis. The PCR amplified products were analyzed in 2% agarose gel (high resolution agarose, Q-BIOgen) in TAE buffer containing 40 mmol/L Tris–HCl (pH 7.9), 4 mmol/L sodium acetate, and 1 mmol/L EDTA (pH 7.9). Aliquots of the PCR products were analyzed in 2% agarose gel stained with ethidium bromide and viewed using a UV transilluminator. PCR products were purified using Sephadex G-50 and were sequenced in both directions (GenBank Accession; MG548644-48, MG844369-81) given in Table 1. The ITS sequence data was compared with those of related genera available in the National Center for Biotechnology Information (NCBI) GenBank. Sequence data was aligned using BioEdit software40 with Clustal W programme41. Phylogenetic and molecular evolutionary analyses were accomplished by constructing Maximum Likelihood tree with the ITS sequences for R. solani obtained from GenBank using Mega42 and MrBayes software43. The positions containing gaps and missing data were eliminated. Bootstrapping was performed at 1000 replications of the data being analyzed.
Statistical analysis
Data pertaining to the pathogenicity testing was statistically analyzed using Genstat 6th edition44. Analysis of variance was used to test differences between variables and means were separated by means of Fisher’s protected least significant differences (LSD).
Results
Sampling and isolation of Rhizoctonia solani
R. solani infection was 100% prevalent to all the visited locations. District wise, maximum mean disease incidence was observed in Islamabad (38.7%) followed by district Attock (36.3%), district Rawalpindi (34.9%) and district Chakwal (29.6%) while minimum mean disease incidence was recorded in district Jhelum (27.5%). Lesions on infected plant stems were irregular in shape, water-soaked, brown in colour, and sunken in appearance. The disease was observed in patches of 6–12 plants. Wilting of diseased plants and stem damage with brown canker near soil line level of the stem was observed in mature plants, however, roots remained healthy. A total of 67 isolates of R. solani recovered on water agar (WA) medium started the hyphal growth from the second day of incubation. The hyphal tips of the actively growing mycelium were cultured on Malt Extract Agar (MEA) medium. The hyphal growth on MEA medium started on the second day, however, the growth was more vigorous than WA medium. Isolates incubated on MEA medium were light grey or medium to dark brown with abundant mycelial growths.
Microscopic studies and morphology of R. solani isolates
Isolates were morphologically characterized according to the descriptions of R. solani by Ogoshi37 and Sneh, et al.29. With a considerable variation, all isolates exhibited typical R. solani colony and cultural characteristics (Fig. 2). Fungal hyphae were branched at right angles and a septum was always present in the branch of hyphae near the originating point with a slight constriction at the branch (Fig. 3). No conidia or conidiophores were observed. The hyphal distance between two septa varied from 67.6 to 149.8 μm (average 109.5 μm). The hyphal diameter of the isolates ranged between 5.1 to 8.1 μm (average 6.40 μm) as shown in Fig. 4. DAPI (4′-6 diamidino-2-phenylindole) stain was used to count a number of nuclei per cell of R. solani. Microscopic studies under a fluorescent light microscope revealed all isolates were multinucleate (Fig. 5). Seven days after incubation on MEA medium 89% of the isolates produced sclerotia, however, 11% of the isolates failed to produce sclerotia. The sclerotia developed from the middle to the edges of the colonies and were light to dark brown in the start and later turned dark brown to black in colour. The sclerotia were either rough or smooth. Most of the isolates produced rough sclerotia that were superficially available on the hyphal mass. Formation of the dark brown to black exudates was also observed in some of the recovered isolates. A detailed morphological description of the isolates is given in the table as Supplementary Information.

Colony morphology of R. solani isolates recovered from tomato.

Branched hyphae of R. solani at right angle to each other with septum under light microscope.

Hyphal diameter of R. solani isolates recovered from tomato.

Nuclear number testing of R. solani isolates stained with DAPI (4′,6-diamidino-2-phenylindole).
Pathogenicity testing
All isolates varied in virulence to stem infections on tomato seedlings leading to wilting and/or plant death. Infection on soil line level of the stem was categorized as −, no symptom (avirulent); ±, brown lesion on part of the stem (Moderately virulent); +, brown lesion girdled the stem (Virulent); ++, brown lesion girdled the stem and plants wilted (Highly virulent). A wide variation in aggressiveness toward tomato was observed among R. solani isolates, as reflected in DI ranging from 0 to 51% for stem damage (Table 2). Out of sixty-seven isolates, 8% of the isolates showed an avirulent response, 10% showed moderately virulent, 19% showed virulent while 63% of the isolates showed a highly virulent response (Table 3).
PCR–RFLP analysis
Discriminating enzymes (MseI, AvaII + HincII and MunI) restricted each fragment to multiple locations for each marker as shown in Fig. 6. The combination of these markers was used to designate specific anastomosis group to each isolate (Table 4). A total of twenty-seven isolates were assigned AG-3 PT as they shared BNAN and FNAN RFLP type. Twenty-three isolates shared BNAN and four isolates shared FNAN RFLP type. MseI restricted the fragments at two locations; 106–119, 188–233 bp for marker B and 194, 327 bp for marker F. The restriction patterns for combination of AvaII and HincII were 189–203 and 306–337 bp for marker NA, while the restriction patterns corresponding to MunI were 22 and 475–550 bp for marker N. This indicates majority of the recovered isolates belong to AG-3 PT as expected strains for crop types.

PCR-RFLP restriction patterns revealed by discriminating enzymes (MseI, AvaII + HincII and MunI).
Six isolates were assigned AG-2-1 and four isolates to AG-2-2 based on the restriction patterns. The restriction patterns corresponding to MseI were 106–109 and 188–233 bp for marker B, 44–57, 58–71 and 193–219 for marker C while 15–23, 84–94 and 192–233 for marker D. The restriction patterns for combination of AvaII and HincII were 20, 63–71, 175–224 and 255–272 bp for marker AA, 71, 448–475 bp for marker AN while 20 b, 70 b, 149 and 287–295 bp for marker BN. MunI restricted the fragments at two locations; 184–215, 306–340 bp for marker A and 22,475–550 bp for marker N. Out of six isolates belonging to AG-2-1, four isolates shared BBNA while two isolates shared DANA RFLP type. Isolates assigned AG-2-2 shared BAAN RFLP type.
Three isolates belong to AG-5 as they shared HAAC types. The restriction patterns corresponding to MseI were 44, 192 and 272 bp for marker H. The restriction patterns for a combination of AvaII and HincII were 20, 63–71, 175–224 and 255–272 bp for marker AA. MunI restricted the fragments at two locations; 51, 186–188, 271–275 bp for marker C.
Two isolates were designated as AG-4 HGI based on the RFLP type; IEAA they shared. The restriction patterns corresponding to MseI were 21–26, 34, 53–72, 71–90, 126–149 and 213–219 bp for marker I while patterns conforming combination of AvaII and HincII were 19, 40–50, 199 and 258–273 for marker EA. MunI restricted the fragments at two locations; 184–215, 306–340 bp for marker A.
Hyphal Anastomosis Interaction
A considerable variation in the hyphal interactions as; C0 (no reaction), C1 (only contact fusion), C2 (somatic fusion or perfect anastomosis) and C3 (auto anastomosis) were observed. A total of twenty-seven isolates formed C2 = somatic fusion of perfect anastomosis interactions with the tester strain AG-3. C3 type or self anastomosis interactions were not taken into consideration as they represented the interactions between hyphae of the same isolates. The C2 type hyphal fusion frequency among these isolates was more than 80%. Selected isolates were identified as members of AG-3. Eleven isolates showed perfect fusion with the tester strain of AG-2. Five isolates showed strong somatic fusion with the tester strains of AG-5 while two isolates fused with the tester strain of AG-4. All these isolates confirmed the identity of PCR-RFLP analysis (Table 4). Two isolates RWPT-14 and JHET15 showed somatic fusion with the tester strains of AG-3, AG-2, AG-4 and AG-6 at 45–69% fusion frequency. The identity of these two isolates was subjected to molecular characterization.
Sequencing of ITS-5.8S rDNA and phylogeny
DNA fragments of eighteen type isolates representing anastomosis groups identified by PCR-RFLP and hyphal anastomosis interactions together with two isolates showing hyphal interactions with more than one tester strain R. solani AGs were subjected to PCR amplification with a set of universal sense primers; ITS1 and ITS4 encoding ITS-1-5.8S-ITS-239. The amplicons generated a fragment of approximately 700 bp on an agarose gel. The ITS region (ITS1, 5.8S rDNA, and ITS2) of each isolate was sequenced in both sense and antisense directions. BLAST analysis of these sequences with the known sequences of R. solani AGs from NCBI GenBank confirmed the identity of respective AGs (99–100% sequence identities) previously revealed by PCR-RFLP and hyphal interactions.
DNA sequences of the isolates representing AG-3 PT, AG-2-1, AG-5, AG-2-2 and AG-4 HGI formed different clades with 99, 90, 93, 99 and 99% bootstrap support respectively (Fig. 7). The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach and then selecting the topology with superior log likelihood value. Each tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). The analysis involved 24 nucleotide sequences. Codon positions included were 1st + 2nd + 3rd + Noncoding. All positions containing gaps and missing data were eliminated. There was a total of 514 positions in the final dataset. Evolutionary analyses were conducted in MEGA745.

Phylogenetic analysis of Rhizoctonia solani isolates infecting tomato.
Isolates; CHKT5, RWPT, CHKT1, ATKT9, JHET8 and ISBT4 shared the clade of reference isolate AG-3 PT. Within this clade, two isolates; CHKT5 and RWPT5 made a distinct cluster with a bootstrap support of 49% from all other isolates. Isolate JHET4 formed a different cluster at 97% bootstrap value from isolates; ATKT10, RWPT14, ISBT5 while all shared the clade of reference isolate of AG-2-1. Isolates; JHET14, CHKT4 and ATKT6 formed a clade with reference isolate of AG-5 while isolate CHKT4 formed different cluster from other isolates with 93% bootstrap support within this clade. Isolates; RWPT8, CHKT8 and ATKT17 shared the same clade with reference isolate AG-2-2.
Discussion
Since the climate of Pothohar region has considerable temperature and moisture variations including semiarid and sub-humid regions46, R. solani can survive under both cool and warm soils. It can remain active at a range of temperatures47, and is well adapted to survive unfavourable conditions as it remains dormant as sclerotia48. The optimum temperature ranges 24–31 °C for vegetative growth and the emergence of tomato seedlings also provide optimum temperatures for R. solani disease development; 24–32 °C49. Pothohar region receives an average of 1,249 mm rainfall of which more than 65% is received in monsoon. Soil moisture greatly influences the amount of R. solani inoculum in the soil50 that ultimately favour the disease development30,51,52. Multicropping and the intercropping are the common practices adopted by the farmers of the region. The use of noncertified seeds is also commonly practised as the same germplasm of the few local varieties is used for cultivation year after years. The most commonly used tomato varieties are Money Maker, Roma Rio Grande and Tropic53. In present studies, Rio Grande was the most susceptible to R. solani infection. It is well accepted that the occurrence of soil-borne pathogens including R. solani is greatly influenced by intensive cropping54,55,56. Solanaceous vegetable cultivation on the same fields also helps in the inoculum multiplication, however, this pathogen is also well adapted for life outside the host plants47,57. A substantial variation in disease incidence among different locations may be attributed to prevailing environmental conditions and different levels of susceptibility in the growing cultivars.
In the present study, a total of 67 isolates of R. solani were recovered from diseased tomato plant samples on malt extract agar (MEA) medium. With considerable variations, all recovered isolates exhibit typical R. solani colony and cultural characteristics. All isolates were multinucleate with 3–8 nuclei per cell and had hyphal branching at a right angle to the constriction found at the point of branching mycelium, a known feature for R. solani described by Sneh, et al.29. A septum, that is of immense taxonomical importance was always present near the branching junction. All 67 isolates were morphologically differentiated and classified on the basis of septal distance, hyphal diameter, no. of sclerotia, texture and topography of sclerotia.
Morphological variations between isolates from different geographical regions have previously been studied by Parmeter, et al.58, Sharma, et al.59 and Goswami, et al.60. Sunder, et al.61 reported colony colour ranged from brown, light brown, dark brown and yellowish brown. Neeraja, et al.62 and Vineeta, et al.63 reported the significant importance of the mycelial and sclerotial characteristics in categorizing R. solani isolates into distinct groups. The septal distance ranged between 67.6 to 149.8 μm. The hyphal diameter ranged between 5.1 to 8.1 μm. Hansen64 also found that hyphal diameter ranged from 4.3–8.0. These findings were also in line with the findings of Vijayan and Nair65. Meyer, et al.66 found some R. solani isolates may not produce sclerotia under certain cultural conditions. Therefore, the absence of sclerotia may not be criteria for the mycelium to be excluded from R. solani. Location of the sclerotial production as superficial or immersed was also supported by the findings of Vineeta, et al.63. Anderson67 and Hoa68 also differentiated sclerotia from different isolates on the basis of colour. The findings of Sinha and Ghufran69 also supported the variations in colony colour, number size and the colour of sclerotia formed. Variations in cultural characteristics were in line with Kuiry, et al.70. Categorizing isolates based on the cultural and morphological features showed the diversity among the isolates was not correlated with their origin of the collection as supported by Baird, et al.71. The morphological classes based on the present studies were however, conservative since only MEA medium was used for this study.
The isolates were subjected to pathogenicity determination under greenhouse experiments on tomato. Distinct variations were observed in the pathogenicity of these isolates. The pathogenicity results showed that isolates within the same AG had variability in pathogenicity and virulence, which may be isolate dependent rather than AG dependent. Among sixty-seven isolates tested for pathogenicity on tomato cv. Rio Grande 63% of the isolates showed a highly virulent response. There was no correlation between mycelial growth and virulence. This was supported by the results of Basu et al. (2004).
Isolates purified using hyphal tipping on PDA medium were preserved by colonizing on hulled barley grains maintained at 4 °C. Sneh, et al.72 and Webb, et al.73 also used the cryogenic storage method for long-term preservation of R. solani isolates.
Hyphal anastomosis interactions are considered to be a more accurate method for accommodating isolates of R. solani into AGs however, reproducibility of this method for a large number of populations is difficult and its reliability at subgroup identification is unsatisfactory17,32. A total of 42 highly virulent isolates were subjected to PCR-RFLP analysis with four discriminating enzymes (MseI, AvaII + HincII and MunI) to categorize them into different at AGs. Results of the PCR-RFLP analysis revealed 27 isolates belonged to AG-3 PT while 6, 4, 3 and 2 isolates belonged to AG-2-1, AG-2-2, AG-5 and AG-4 HGI respectively.
Each isolate was paired with the tester strain of respective AG identified by PCR-RFLP. Environmental factors including temperature variations and nutritional stress may greatly influence the vegetative compatibility of the isolates74. In present studies, compatibility of the isolates with tester strains was tested on MEA medium with optimum growth conditions. Among four types of anastomosis reactions; C0 to C3 only C2 reactions were considered as somatic fusion or perfect anastomosis as described by Carling36. All isolates except two confirmed the AG group identity revealed by PCR-RFLP while the identity of two isolates was not confirmed as they produced hyphal interactions with more than one tester strains of the known AGs. The identity of these two isolates was unknown and they were further subjected to sequence analysis of their ITS-5.8S rDNA.
Type isolates representing anastomosis groups identified by PCR-RFLP and confirmed by hyphal anastomosis interactions together with two unknown isolates were subjected to PCR amplification with a set of universal sense primers; ITS1 and ITS4 encoding ITS-1-5.8S-ITS-2. BLAST analysis of the obtained sequences with the known sequences of R. solani AGs from NCBI GenBank confirmed the identity of respective AGs (99–100% sequence identities). All the isolates showed heterogeneity in their ITS sequences. Sequence analysis also confirmed the identity of unknown isolates. DNA sequences of R. solani isolates from Pakistan infecting tomato crop are now available at NCBI.
AG group composition of the virulent isolates revealed 64.2% isolated belonged to AG-3 PT and they were predominant to all tomato production areas of Pothohar region concordant with the previous findings by Rauf, et al.19 on potato. The frequency of the other AGs was far less than AG-3 as AG-2-1 14.2%, AG-2-2 9.5%, AG-5 7.1% and AG-4 HGI 4.7%. The association of AG-3 PT and AG-5 with foot rot of tomato was also supported by the findings of Muzhinji, et al.32 however the association of AG-2-1 was in contradiction. AG-2-1 and AG-2-2 have also been reported to cause foot rot of tomato22. The relative proportion of AG-3 PT, AG-5 and AG-2-1 in the present study is in line with the findings of Campion, et al.75 and Chand and Logan76. Majority of the AG-2-1, AG-2-2 and AG-4 HGI isolates were found in the three districts; Attock, Rawalpindi, and Jhelum. The localized occurrence of these two AGs could be attributed to the susceptible preceding crops like potato and chilli.
Conclusion and Recommendations
Study reports status of R. solani infection with reference to Pakistan along with the occurrence of five anastomosis groups of R. solani on tomato with various levels of intensities. AG-3 PT was found to be the most prevalent and aggressive compared to other AGs in all tomato growing areas. Stringent surveillance for the occurrence of R. solani AGs on vegetable crops is needed. While evolving new varieties of tomato, R. solani isolates belonging to the reported anastomosis groups (AGs) may be used in breeding program.
References
Onyambus, G. K., Maranga, R. O., Gitonga, L. M. & Knapp, M. Host plant resistance among tomato accessions to the spider mite Tetranychus evansi in Kenya. Experimental and Applied Acarology 54, 385–393 (2011).
Georgé, S. et al. Changes in the contents of carotenoids, phenolic compounds and vitamin C during technical processing and lyophilisation of red and yellow tomatoes. Food Chemistry 124, 1603–1611, https://doi.org/10.1016/j.foodchem.2010.08.024 (2011).
FAO. (United Nations, New York, United States, 2016).
Roux, F. et al. Resistance to phytopathogense tutti quanti: placing plant quantitative disease resistance on the map. Molecular Plant Pathology 15, 427–432, https://doi.org/10.1111/mpp.12138 (2014).
Mickelbart, M. V., Hasegawa, P. M. & Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nature Reviews Genetics 16, 237–251, https://doi.org/10.1038/nrg3901 (2015).
Agrios, G. N. In Plant Pathology Vol. 4 Ch. 952, 385–614 (Elsevier, 2005).
Morsy, E. M., Abdel-Kawi, K. A. & Khalil, M. N. A. Efficiency of Trichoderma viride and Bacillus subtilis as biocontrol agents against Fusarium solani on tomato plants. Egyptian Journal of Phytopathology 37, 47–57 (2009).
Newman, S. E., Brown, W. M. & Ozbay, N. In XXVI International Horticultural Congress: Managing Soil-Borne Pathogens: A Sound Rhizosphere to Improve Productivity in Intensive Horticultural Systems. 131–135 (ISHS Acta Horticulturae).
Nikraftar, F., Taheri, P., Falahati Rastegar, M. & Tarighi, S. Tomato partial resistance to Rhizoctonia solani involves antioxidative defense mechanisms. Physiological and Molecular Plant Pathology 81, 74–83, https://doi.org/10.1016/j.pmpp.2012.11.004 (2013).
Adams, G. C. Thanatephorus cucumeris (Rhizoctonia solani), a species complex of wide host range. Advances in Plant Pathology 6, 535–552 (1988).
Sneh, B. Rhizoctonia Species: taxonomy, molecular biology, ecology, pathology and disease control. (Springer, 1996).
Prabha, S. B., Vignesh, A. & Murugesan, K. Biological control of damping off and stem rot of tomato (Lycopersicon esculentum Mill.) using an antagonistic actinomycete, Saccharopolyspora sp. International Journal of Agricultural Science and Research 4, 55–65 (2014).
Carling, D. E., Baird, R. E., Gitaitis, R. D., Brainard, K. A. & Kuninaga, S. Characterization of AG-13, a newly reported anastomosis group of Rhizoctonia solani. Phytopathology 92, 893–899 (2002).
Carling, D. E., Kuninaga, S. & Brainard, K. A. Hyphal anastomosis reactions, rDNA-internal transcribed spacer sequences, and virulence levels among subsets of Rhizoctonia solani anastomosis group-2 (AG-2) and AG-BI. Phytopathology 92, 43–50 (2002).
Gónzalez, D. et al. Phylogenetic relationships of Rhizoctonia fungi within the Cantharellales. Fungal Biology 120(4), 603–619 (2016).
Stodart, B. J., Harvey, P. R., Neate, S. M., Melanson, D. L. & Scott, E. S. Genetic variation and pathogenicity of anastomosis group 2 isolates of Rhizoctonia solani in Australia. Mycological Research 111, 891–900, https://doi.org/10.1016/j.mycres.2007.05.008 (2007).
Fang, X., Finnegan, P. M. & Barbetti, M. J. Wide Variation in Virulence and Genetic Diversity of Binucleate Rhizoctonia Isolates Associated with Root Rot of Strawberry in Western Australia. PLOS One 8, 55877, https://doi.org/10.1371/journal.pone.0055877 (2013).
Sharon, M., Sneh, B., Kuninaga, S., Hyakumachi, M. & Naito, S. Classification of Rhizoctonia spp. using rDNA-ITS sequence analysis supports the genetic basis of the classical anastomosis grouping. Mycoscience 49, 93–114, https://doi.org/10.1007/s10267-007-0394-0 (2008).
Rauf, C., Ahmad, I. & Ashraf, M. Anastomosis groups of Rhizoctonia solani Kuhn isolates from potato in Pakistan. Pakistan Journal of Botany 39, 1335–1340 (2007).
Woodhall, J. W., Lees, A. K., Edwards, S. G. & Jenkinson, P. Infection of potato by Rhizoctonia solani: Effect of anastomosis group. Plant Pathology 57, 897–905, https://doi.org/10.1111/j.1365-3059.2008.01889.x (2008).
Kodama, T., Horimoto, K. & Ogoshi, A. On the brown spot of eggplant caused by Thanatephorus cucumeris (Frank) Donk (Rhizoctonia solani) AG3. Annals of the Phytopathological Society of Japan 48, 237–243 (1982).
Misawa, T. & Kuninaga, S. The first report of tomato foot rot caused by Rhizoctonia solani AG-3 PT and AG-2-Nt and its host range and molecular characterization. Journal of General Plant Pathology 76, 310–319, https://doi.org/10.1007/s10327-010-0261-2 (2010).
Mikhail, M. S., Sabet, K. K., Omar, M. R., Asran, A. A. & Kasem, K. K. Current Rhizoctonia solani anastomosis groups in Egypt and their pathogenic relation to cotton seedlings. African Journal of Microbiology Research 4, 386–395 (2010).
Kuramae, E. E., Buzeto, A. L., Ciampi, M. B. & Souza, N. L. Identification of Rhizoctonia solani AG 1-IB in lettuce, AG 4 HG-I in tomato and melon, and AG 4 HG-III in broccoli and spinach, in Brazil. European Journal of Plant Pathology 109, 391–395 (2003).
Yildiz, A. & Döken, M. T. Anastomosis group determination of Rhizoctonia solani Kühn (telemorph: Thanatephorus cucumeris) isolates from tomatoes grown in Aydin, Turkey and their disease reaction on various tomato cultivars. Journal of Phytopathology 150, 526–528 (2002).
Chaudhry, Q. Z. & Rasul, G. Agro-climatic classification of Pakistan. Science Vision 9, 59 (2004).
Ogoshi, A. Ecology And Pathogenicity of Anastomosis And Intraspecific Groups of Rhizoctonia Solani Kuhn. Annual Review of Phytopathology 25, 125–143, https://doi.org/10.1146/annurev.phyto.25.1.125 (1987).
Naito, S. A simple method for the long-time culture storage of Rhizoctonia spp. on barley grains 20–23 (Japan, 1993).
Sneh, B., Burpee, L. & Ogoshi, A. Identification of Rhizoctonia species. (APS press, 1991).
Naz, F., Rauf, C. A., Abbasi, N. A., Haque, I. & Ahmad, I. Influence of inoculum levels of Rhizoctonia solani and susceptibility on new potato germplasm. Pakistan Journal of Botany 40, 2199–2209 (2008).
Muzhinji, N., Woodhall, J. W., Truter, M. & Van der Waals, J. E. Elephant hide and growth cracking on potato tubers caused by Rhizoctonia solani AG3-PT in South Africa. Plant Disease 98, 570–570 (2014).
Muzhinji, N., Truter, M., Woodhall, J. W. & Van der Waals, J. E. Anastomosis groups and pathogenicity of Rhizoctonia solani and binucleate Rhizoctonia from potato in South Africa. Plant Disease 99, 1790–1802 (2015).
Sharon, M., Kuninaga, S., Hyakumachi, M. & Sneh, B. The advancing identification and classification of Rhizoctonia spp. using molecular and biotechnological methods compared with the classical anastomosis grouping. Mycoscience 47, 299–316 (2006).
Camporota, P., Soulas, M. L. & Perrin, R. In 4th Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party: Disease and Insects in Forest Nurseries. 95–106.
Guillemaut, C. et al. Typing of anastomosis groups of Rhizoctonia solani by restriction analysis of ribosomal DNA. Canadian Journal of Microbiology 49, 556–568 (2003).
Carling, D. E. In Rhizoctonia species: taxonomy, molecular biology, ecology, pathology and disease control 37–47 (Springer, 1996).
Ogoshi, A. Grouping of Rhizoctonia solani Kühn and their perfect stages. Review of Plant Protection Research 8, 98–103 (1975).
Carling, D. E. & Sumner, D. R. In Methods for Research on Soilborne Phytopathogenic Fungi (eds Singleton. L. L., Mihail, J. D. & Rush. C. M.) 157–165 (American Phytopathological Society, 1992).
White, T. J., Bruns, T., Lee, S. J. W. T. & Taylor, J. L. In In: PCR protocols: a guide to methods and applications Vol. 18 315–322 (Academic Press, Inc., 1990).
Hall, T. A. In Nucleic acids symposium series (Ibis Therapeutics; A division of Isis Pharmaceuticals, Carlsbad, CA, USA, 1999).
Thompson, J. D., Higgins, D. G. & Gibson, T. J. Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research 22, 4673–4680 (1994).
Tamura, K. & Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology and Evolution 10, 512–526 (1993).
Ronquist, F. & Huelsenbeck, J. P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19, 1572–1574 (2003).
GenStat-Committee. (ed. Lawes Agricultural Trust) (Rothamsted Research, Harpenden, United Kingdom, 2000).
Kumar, S., Stecher, G. & Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33, 1870–1874 (2016).
Shamshad, K. M. (no paging) (Royal Book Agency, Karachi, Pakistan, 1988).
Olsen, M. W. & Young, G. J. Damping Off. 1–3 (Cooperative Extension, College of Agriculture, The University of Arizona, 2011).
Ceresini, P. C., Shew, H. D., Vilgalys, R. J. & Cubeta, M. A. Genetic diversity of Rhizoctonia solani AG-3 from potato and tobacco in North Carolina. Mycologia 94, 437–449 (2002).
Harikrishnan, R. & Yang, X. Recovery of anastomosis groups of Rhizoctonia solani from different latitudinal positions and influence of temperatures on their growth and survival. Plant Disease 88, 817–823 (2004).
Frank, J. A. The Rhizoctonia disease of potatoes in Maine. American Potato Journal 55, 59–60 (1978).
Kyritsis, P. & Wale, S. J. In British Crop Protection Conference: Pests & Diseases. 761–764 (British Crop Protection Enterprises).
Shehata, M. A., Davis, D. W. & Anderson, N. A. Resistance to Rhizoctonia stem rot in peas as influenced by temperature, watering method, and period of disease development (Rhizoctonia solani). Plant. Diseases 68, 22–24 (1984).
Sajjad, M., Ashfaq, M., Suhail, A. & Akhtar, S. Screening of tomato genotypes for resistance to tomato fruit borer (Helicoverpa armiger Hubner) in Pakistan. Pakistan Journal of Agricultural Sciences 48, 59–62 (2011).
Gilligan, C. A., Simons, S. A. & Hide, G. A. Inoculum density and spatial pattern of Rhizoctonia solani in field plots of Solanum tuberosum: effects of cropping frequency. Plant Pathology 45, 232–244 (1996).
Hooker, W. J. Compendium of potato diseases. 59 (International Potato Center, 1981).
Kluth, C., Buhre, C. & Varrelmann, M. Susceptibility of intercrops to infection with Rhizoctonia solani AG 2‐2 IIIB and influence on subsequently cultivated sugar beet. Plant Pathology 59, 683–692 (2010).
Keijer, J. et al. In vitro analysis of host plant specificity in Rhizoctonia solani. Plant Pathology 46, 659–669 (1997).
Parmeter, J. R., Sherwood, R. T. & Platt, W. D. Anastomosis grouping among isolates of Thanatephorus cucumeris. Phytopathology 9, 1270–1278 (1969).
Sharma, M., Gupta, S. K. & Sharma, T. R. Characterization of variability in Rhizoctonia solani by using morphological and molecular markers. Journal of Phytopathology 153, 449–456 (2005).
Goswami, B. K., Bhuiyan, K. A. & Mian, I. H. Morphological and pathogenic variations in the isolates of Rhizoctonia solani in Bangladesh. Bangladesh Journal of Agricultural Research 35, 375–380 (2010).
Sunder, S., Singh, R. & Dodan, D. S. Standardization of inoculation methods and management of sheath blight of rice. Indian Journal of Plant Pathology 21, 92–96 (2003).
Neeraja, C. N., Vijayabhanu, N., Shenoy, V. V., Reddy, C. S. & Sarma, N. P. RAPD analysis of Indian isolates of rice sheath blight fungus Rhizoctonia solani. Journal of Plant Biochemistry and Biotechnology 11, 43–48 (2002).
Vineeta, S., Singh, U. S., Singh, K. P., Major, S. & Anil, K. Genetic diversity of Rhizoctonia solani isolates from rice: differentiation by morphological characteristics, pathogenicity, anastomosis behaviour and RAPD fingerprinting. Journal of Mycology and Plant Pathology 32, 332–344 (2002).
Hansen, L. R. Sharp eye spot on grain parachute by Rhizoctonia solani Kühn. Report Norway Landbrttøjsk 42, 1–12 (1963).
Vijayan, M. & Nair, M. C. Anastomosis grouping of isolates of Rhizoctonia solani Kühn (Thanatephorus cucumeris (Frank) Donk) causing sheath blight of rice. Current Science 54, 289–291 (1985).
Meyer, L., Wehner, F. C., Nel, L. H. & Carling, D. E. Characterization of the crater disease strain of Rhizoctonia solani. Phytopathology 88, 366–371 (1998).
Anderson, N. A. The Genetics and Pathology of Rhizoctonia Solani. Annual Review of Phytopathology 20, 329–347, https://doi.org/10.1146/annurev.py.20.090182.001553 (1982).
Hoa, T. T. C. Characterization and Pathogenicity of Rhizoctonia Solani Kuhn Isolates from Different Rice Zones and Management of Sheath Blight of Rice M. Phil. thesis, IARI, Division of Mycology and Plant Pathology, New Delhi (1994).
Sinha, B. B. P. & Ghufran, S. M. Physiological studies on five isolates of sheath blight of rice caused by Rhizoctonia solani Kuhn. Journal of Research-Rajendra Agricultural University 6, 61–67 (1988).
Kuiry, S. P., Mondal, A., Banerjee, S. & Dutta, S. Morphological variability in Rhizoctonia solani isolates from different agro-ecological zones of West Bengal, India. Archives of Phytopathology and Plant Protection 47, 728–736 (2014).
Baird, R. E., Carling, D. E. & Mullinix, B. G. Characterization and comparison of isolates of Rhizoctonia solani AG-7 from Arkansas, Indiana, and Japan, and select AG-4 isolates. Plant Disease 80, 1421–1424 (1996).
Sneh, B., Zeidan, M., Ichielevich-Auster, M., Barash, I. & Koltin, Y. Increased growth responses induced by a nonpathogenic Rhizoctonia solani. Canadian Journal of Botany 64, 2372–2378 (1986).
Webb, K. M., Hill, A. L., Laufman, J., Hanson, L. E. & Panella, L. Long‐term preservation of a collection of Rhizoctonia solani using cryogenic storage. Annals of Applied Biology 158, 297–304 (2011).
Julián, M. C., Debets, F. & Keijer, J. Independence of sexual and vegetative incompatibility mechanisms of Thanatephorus cucumeris (Rhizoctonia solani) anastomosis group 1. Phytopathology 86, 566–574 (1996).
Campion, C., Chatot, C., Perraton, B. & Andrivon, D. Anastomosis groups, pathogenicity and sensitivity to fungicides of Rhizoctonia solani isolates collected on potato crops in France. European Journal of Plant Pathology 109, 983–992 (2003).
Chand, T. & Logan, C. Cultural and pathogenic variation in potato isolates of Rhizoctonia solani in Northern Ireland. Transactions of the British Mycological Society 81, 585–589, https://doi.org/10.1016/s0007-1536(83)80129-6 (1983).
Acknowledgements
Financial assistance received from Higher Education Commission Islamabad, Pakistan under HEC Indigenous PhD Fellowships for 5000 Scholars (112-34183-2AV1-067) and Department of Plant, Soil and Microbial Sciences, Michigan State University for hosting scholar under HEC International Research Support Initiative Program is gratefully acknowledged.
Author information
Authors and Affiliations
Contributions
Amjad Shahzad Gondal Conducted research experiment and wrote the paper. Abdul Rauf Helped author during research as major supervisor. Farah Naz Helped author during research as supervisory committee member.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gondal, A.S., Rauf, A. & Naz, F. Anastomosis Groups of Rhizoctonia solani associated with tomato foot rot in Pothohar Region of Pakistan. Sci Rep 9, 3910 (2019). https://doi.org/10.1038/s41598-019-40043-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-40043-5
This article is cited by
-
Diversity and Pathogenicity of Anastomosis Groups of Rhizoctonia solani Isolates Associated with Potato Diseases in Northern Sinaloa, Mexico
Potato Research (2024)
-
Selection of reference genes for RT-qPCR analysis of rice with Rhizoctonia solani infection and biocontrol PGPR/KSi application
Molecular Biology Reports (2023)
-
Research progress of volatile organic compounds produced by plant endophytic bacteria in control of postharvest diseases of fruits and vegetables
World Journal of Microbiology and Biotechnology (2023)
-
Molecular variability and host range of Rhizoctonia solani AG1-IA in rice-vegetable based cropping systems in West Bengal, India
Journal of Plant Pathology (2022)
-
Anastomosis groups and sensitivity to fungicides of Rhizoctonia solani strains isolated from potato in Russia
Journal of Plant Diseases and Protection (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
