
Abstract
Phospho flow is a powerful approach to detect cell signaling aberrations, identify biomarkers and assess pharmacodynamics, and can be performed using cryopreserved samples. The effects of cryopreservation on signaling responses and the reproducibility of phospho flow measurements are however unknown in many cell systems. Here, B lymphocytes were isolated from healthy donors and patients with the B cell malignancy chronic lymphocytic leukemia and analyzed by phospho flow using phospho-specific antibodies targeting 20 different protein epitopes. Cells were analyzed both at basal conditions and after activation of cluster of differentiation 40 (CD40) or the B cell receptor. Pharmacodynamics of the novel pathway inhibitor ibrutinib was also assessed. At all conditions, fresh cells were compared to cryopreserved cells. Minimal variation between fresh and frozen samples was detected. Reproducibility was tested by running samples from the same donors in different experiments. The results demonstrate reproducibility across different phospho flow runs and support the use of cryopreserved samples in future phospho flow studies of B lymphocytes.
Similar content being viewed by others

Introduction
Cancer represents an extremely heterogeneous disease underscored by high genetic variability, risk of clonal progression of resistant tumors after treatment with targeted therapies and diverse clinical outcomes. Much effort is invested in identification of biomarkers showing prognostic value, indicating treatment options or predicting disease outcome. Novel targeted therapies are often directed at signaling molecules, including the PI3Kδ inhibitor idelalisib and the Bruton tyrosine kinase (BTK) inhibitor ibrutinib, both used to treat B cell malignancies, including chronic lymphocytic leukemia (CLL)1. Since activation of mitogenic signaling pathways play a central role in cancer development and progression it is reasonable to assume that the activity level of central signaling molecules may serve as cancer biomarkers. For medium- to high-throughput screening of single cell phospho-protein levels, phospho flow cytometry is an attractive approach2. By applying this technique, signaling patterns have been characterized in diverse B cell malignancies3,4,5,6. We recently identified STAT3 (pY705) as aberrantly upregulated in a group of CLL patients and showed that STAT3 inhibitors potently kill the cancer cells4. Phospho flow analysis can thus be used to identify relevant drug targets.
Characterization of cell signaling is commonly performed on cryopreserved cells. Cryopreservation allows long-term storage of lymphocytes before immunological assays are performed. In this way, samples collected at multiple time points or from multiple patients can be analyzed simultaneously, reducing variation arising from different runs. Frozen samples can also easily be transported to a single laboratory for analysis, reducing variation introduced by different operators or laboratory conditions7. This is particularly useful in multi-center trials when fresh samples cannot be transported within an acceptable time frame to ensure intact samples. It is, however, important that freezing, storage and thawing do not have major negative impact on assay read-outs.
The effects of cryopreservation have been most extensively studied in T cells, but opinion and empirical evidence are divided. To varying degree cryopreservation has been shown to alter stability of surface marker expression8,9,10, cytokine production9 and cell proliferation11. Such discrepancies can, however, be minimized when an optimized protocol is followed10,12,13,14. Freezing of cells within 12 h of blood collection12, minimized temperature fluctuations during storage15 and resting of the thawed cells before staining for phenotypic analysis16 are factors which have been shown to improve preservation of cell function. In general, T cells have been shown to be more sensitive to cryopreservation than other immune cells16. Studies on the effect of cryopreservation on B-cell function by ELISpot have shown little variation between fresh and frozen cells and thus support the use of cryopreserved samples7,17.
The aim of this study was to compare signaling responses in B lymphocytes detected by phospho flow using both fresh and frozen samples and to test the reproducibility of the phospho flow assay.
Methods
Patient material and ethical considerations
Buffy coats from anonymized healthy blood donors and blood samples from CLL patients were received from the Blood Centre (Oslo University Hospital) and the Department of Haematology, Oslo University Hospital, respectively, following written informed consent from all donors. The study was approved by the Regional Committee for Medical and Health Research Ethics of South-East Norway, and the research on human blood was carried out in accordance with the Declaration of Helsinki18.
Reagents and antibodies
sCD40L (cat. no. 11343345) was from ImmunoTools, Germany and F(ab’)2 anti-human IgM (cat. no. 2022-01) was from Southern Biotechnology, CA, USA. Ibrutinib was purchased from Selleckchem (TX, USA). A list of the antibodies used in this study can be found in Supplementary Table 1.
Isolation of B lymphocytes
Normal B cells were isolated from buffy coats by negative selection using RosetteSep Human B Cell Enrichment Cocktail (20 μl/mL blood; Stemcell Technologies, Cambridge, United Kingdom) followed by gradient centrifugation with Lymphoprep (Alere Technologies AS, Oslo, Norway). CLL cells were isolated from whole blood by Lymphoprep according to the manufacturer’s protocol. Cell samples were either analyzed immediately after isolation or cryopreserved in liquid nitrogen. Cryopreserved cells were rested in RPMI 1640 GlutaMAX medium supplemented with 10% fetal calf serum at 37 °C and 5% CO2 for one hour after thawing.
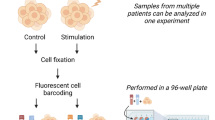
Phospho flow with fluorescent cell barcoding
Single cell signaling analysis was carried out using phospho flow with fluorescent cell barcoding as described19. Briefly, B cells were treated as described in the figure legends before fixation (BD Phosflow Fix Buffer I, BD Biosciences, Franklin Lakes, NJ, USA). The cells were then washed three times with PBS and stained with different concentrations of the barcoding reagents Alexa Fluor 488 5-TFP, Pacific Blue Succinimidyl Ester and Pacific Orange Succinimidyl Ester (all from ThermoFisher Scientific, Waltham, MA, USA) for 20 min at room temperature. The cells were then washed twice with flow wash (PBS, 10% FCS and 0.09% sodium azide), combined in one tube and permeabilized with BD Phosflow Perm Buffer III (BD Biosciences) which was pre-cooled at −20 °C, and kept at −80 °C until further processing. Before antibody staining, the cells were washed three times with flow wash. The antibodies used in this study are listed in Supplementary Table 1, including the surface marker anti-CD19. After 30 min incubation with the antibodies, the cells were washed once, resuspended in flow wash and analyzed with a BD FACSCanto II (4-2-2) cytometer equipped with 405 nm, 488 nm and 633 nm lasers. The data were analyzed in Cytobank (https://cellmass.cytobank.org/cytobank/) as described4.
Statistical analysis
Statistical analyses were performed in Prism 7 (GraphPad Software, CA, USA) using the statistical tests indicated in the figure legends.
Results and Discussion
Characterization of basal signaling patterns in freshly isolated and cryopreserved primary B cells
In order to investigate whether the basal phosphorylation level in primary B cells is affected by cryopreservation, a medium throughput single cell signaling analysis was carried out on freshly isolated and cryopreserved primary human B cells from 10 healthy donors by phospho flow (Fig. 1). The phosphorylation level of 20 signaling molecules relevant to B-cell signaling was analyzed. As many as 19 out of 20 phospho-proteins were not differently regulated in freshly isolated versus cryopreserved B cells (Fig. 1). This analysis demonstrates that basal signaling patterns in B cells are nearly unaffected by cryopreservation and suggests that cryopreserved cells can serve as a model for fresh cells in studies of basal signaling.

Characterization of basal signaling patterns in freshly isolated and cryopreserved primary B cells. B cells were isolated from healthy blood donors and analyzed immediately (n = 10; fresh) or after cryopreservation (n = 10; cryo). The cells were fixed, permeabilized and stained with anti-CD19 surface marker and phospho-specific antibodies as described in Methods. The fluorescence signals were detected by flow cytometry and analyzed in Cytobank. The basal fluorescence signals were measured relative to IgGκ isotype control and shown as arcsinh ratio (box-and-whisker; outliers are indicated separately). Statistical significance was calculated by a paired t-test with correction for multiple comparisons using the Holm-Sidak’s method, ****p < 0.00001. The plots to the right show raw flow cytometry data from one representative donor (fresh). Each plot corresponds to the protein shown in the aligned box-and-whisker plot, with phospho-protein (x-axis) plotted against FSC-A (y-axis).
Phospho flow measurements are highly reproducible
Characterization of signaling patterns by phospho flow has shown promising utility in cancer biomarker discovery20,21. When patient samples are analyzed in search of predictive biomarkers, small differences in signaling amplitude may distinguish patients classified as positive or negative for a given biomarker. It is therefore very important that the phospho flow assay produces reliable and reproducible data. In order to test the reproducibility of the method, B lymphocytes from four healthy donors were aliquoted and fixed, followed by antibody staining and analysis on three different days. The normal B cells were analyzed for STAT3 (pY705) phosphorylation level both before and after cryopreservation (Fig. 2). As shown, the triplicates presented highly similar, almost overlapping, phospho-signals, demonstrating the robustness of the assay (Fig. 2). A recent report from our group showed that the basal level of STAT3 (pY705) is upregulated in CLL patients4. Three cryopreserved CLL patient samples were therefore included as a control and analyzed in triplicate on three different days. The STAT3 (pY705) levels in the triplicate patient samples were strongly reproducible and, in agreement with our previous report, elevated compared to the level in normal B cells (Fig. 2). These results indicate that phospho flow is a reliable method for biomarker identification.

Phospho flow measurements are highly reproducible. Basal STAT3 (pY705) signals were analyzed in normal B cells from four healthy blood donors (N1-N4) before (fresh) and after (cryo) cryopreservation, and in cryopreserved B cells from three CLL patients (CLL105, CLL135, CLL136). Symbols with a cross represent CLL patients with unmutated IgVH status. The analysis was done on three samples from each donor on three different days, following the procedure described in Methods.
In addition to the low intra-donor variability in phospho-protein levels detected by phospho flow, we here show that the variability between healthy donors, as well as between different studies performed by different operators, is low. Ten cryopreserved healthy samples from an earlier study4 were compared with the ten cryopreserved healthy samples from this study. As shown in Supplementary Fig. 1, the basal phosphorylation levels of all 20 proteins analyzed were similar between the two groups. This suggests that a relatively low number of healthy donors is needed to control patient studies and that historical data can be used.
sCD40L induced signaling is not significantly affected by cryopreservation
When cells are located in their natural microenvironment they will receive signals from the microenvironment which will induce signal transduction and phosphorylation events. We have shown that activation of the CD40 co-receptor induces distinct signaling patterns in normal B cells and CLL cells6. To test if protein phosphorylation induced by microenvironmental stimuli is affected by cryopreservation, phospho flow was performed on fresh and cryopreserved normal B cells after exposure to sCD40L for 5–10 min. Twenty signaling molecules (see Fig. 1) were analyzed, of which 12 that displayed significantly induced phosphorylation levels upon sCD40L stimulation are presented in Fig. 3. Of these, six proteins did not show any significant variation in regulation between fresh and cryopreserved cells. Five proteins showed statistically different phosphorylation level at only one of the time-points tested, whereas Akt (pS473) as the only protein was significantly different in fresh and cryopreserved cells at both time-points (Fig. 3). Interestingly, the proteins that displayed modified signaling response as a result of cryopreservation primarily belonged to the MAPK and PI3K pathways. In summary, little variation is observed between fresh and cryopreserved cells after sCD40L stimulation.

sCD40L induced signaling is not significantly affected by cryopreservation. B cells from the same two healthy donors were analyzed before (fresh) and after (cryo) cryopreservation. The cells were stimulated with sCD40L (400 ng/mL) for 5 and 10 min before fixation and phospho flow analysis as described in Methods. Results are shown as mean ± standard deviation. Statistical significance was calculated between the two groups (fresh, cryo) by an ordinary two-way ANOVA with Sidak’s multiple comparisons test. **p < 0.01, *p < 0.05.
Signaling downstream of the B-cell receptor is mostly unaffected by cryopreservation
The B-cell receptor (BCR) plays a central role in B cell signaling and development of B cell malignancies. Over the last few years, management of hematological diseases has focused on novel therapies that target the BCR signaling pathway, including the Btk inhibitor ibrutinib1. BCR induced signaling is therefore very relevant in biomarker discovery studies. To test if BCR signaling is affected by cryopreservation, normal B cells were stimulated with anti-IgM for up to 30 min. The effects on 20 signaling molecules were monitored (Fig. 4). For the majority of the analyzed proteins the signaling responses in fresh and cryopreserved cells were practically identical (Fig. 4). Only four proteins [BLNK (pY84), Histone H3 (pS10), Lck (pY505) and MEK1 (pS298)] showed statistically significant responses in fresh versus cryopreserved cells. In conclusion, BCR induced signaling is mostly unaffected by cryopreservation.

Signaling downstream of the BCR is mostly unaffected by cryopreservation. B cells were isolated from healthy blood donors and analyzed immediately (n = 3; fresh) or after cryopreservation (n = 3; cryo). The cells were stimulated with anti-IgM (10 μg/mL) for the indicated time-course before fixation and phospho flow analysis as described in Methods. Results are shown as mean ± standard error of the mean (SEM). Statistical significance was calculated between the two groups (fresh, cryo) by a paired t-test with excluded time parameter. Significant p-values are indicated.
A similar study of anti-IgM induced signaling responses was previously performed on B cells from 10 healthy donors4. Since the data analysis was performed in the same way, and the data are presented as arcsinh ratio in both studies, the effects can easily be compared. As shown in Supplementary Fig. 2, for most of the 20 phospho-proteins analyzed, the signals of the donors from the two studies lie within the same range. This confirms low variability in phospho flow experiments over time and between operators, as observed in other assays on primary B cells7.
Signaling responses induced by ibrutinib treatment are unaffected by cryopreservation
Assessment of pharmacodynamics of drugs that interfere with signaling is one application of phospho flow and a central topic in precision medicine. As mentioned above, novel cancer therapies include BCR inhibitors such as ibrutinib. Here, the effect of cryopreservation on pharmacodynamics of ibrutinib was investigated as an example. Normal B cells were exposed to a range of ibrutinib concentrations for 20 min before stimulation with anti-IgM for three min. Ibrutinib treatment reduced the phosphorylation level of several proteins downstream of the BCR (Fig. 5). However, some of the effects were only observed at very high concentrations and are probably not clinically relevant. Importantly, no statistically significant differences in signaling responses were detected between fresh and cryopreserved cells (Fig. 5).

Signaling responses induced by ibrutinib treatment is unaffected by cryopreservation. B cells were isolated from healthy blood donors and analyzed immediately (n = 3; fresh) or after cryopreservation (n = 3; cryo). The cells were treated with the indicated concentrations of ibrutinib for 20 min before stimulation with anti-IgM (10 μg/mL) for three min. The cells were then fixed and analyzed by phospho flow as described in Methods. Results are shown as mean ± standard deviation. Statistics were performed as in Fig. 4. No significant p-values were achieved.
Conclusion
Minimal variation was observed between fresh and cryopreserved primary B cells in phospho flow assays, which is in agreement with earlier reports in which other methods were applied7,17. Importantly, the presented protocol was optimized to improve preservation of cell function after cryopreservation, i.e. the cells were frozen within 12 h of blood collection12, temperature fluctuations were minimal during storage15 and the thawed cells were rested before signaling analysis16. In conclusion, the present study supports the use of cryopreserved cells in biomarker discovery and assessment of pharmacodynamics in B cell malignancies.
References
Arnason, J. E. & Brown, J. R. Targeting B Cell Signaling in Chronic Lymphocytic Leukemia. Curr. Oncol. Rep 19, 61, https://doi.org/10.1007/s11912-017-0620-7 (2017).
Krutzik, P. O., Irish, J. M., Nolan, G. P. & Perez, O. D. Analysis of protein phosphorylation and cellular signaling events by flow cytometry: techniques and clinical applications. Clin. Immunol 110, 206–221, https://doi.org/10.1016/j.clim.2003.11.009 (2004).
Blix, E. S. et al. Phospho-specific flow cytometry identifies aberrant signaling in indolent B-cell lymphoma. BMC. Cancer 12, 478, https://doi.org/10.1186/1471-2407-12-478 (2012).
Myhrvold, I. K. et al. Single cell profiling of phospho-protein levels in chronic lymphocytic leukemia. Oncotarget 9, 9273–9284, https://doi.org/10.18632/oncotarget.23949 (2018).
Myklebust, J. H. et al. Distinct patterns of B-cell receptor signaling in non-Hodgkin lymphomas identified by single-cell profiling. Blood 129, 759–770, https://doi.org/10.1182/blood-2016-05-718494 (2017).
Parente-Ribes, A. et al. Spleen tyrosine kinase inhibitors reduce CD40L-induced proliferation of chronic lymphocytic leukemia cells but not normal B cells. Haematologica 101, e59–e62, https://doi.org/10.3324/haematol.2015.135590 (2016).
Truck, J. et al. Effect of cryopreservation of peripheral blood mononuclear cells (PBMCs) on the variability of an antigen-specific memory B cell ELISpot. Hum. Vaccin. Immunother 10, 2490–2496, https://doi.org/10.4161/hv.29318 (2014).
Costantini, A. et al. Effects of cryopreservation on lymphocyte immunophenotype and function. J. Immunol. Methods 278, 145–155, S0022175903002023 (2003).
Owen, R. E. et al. Loss of T cell responses following long-term cryopreservation. J. Immunol. Methods 326, 93–115, https://doi.org/10.1016/j.jim.2007.07.012 (2007).
Weinberg, A. et al. Optimization and limitations of use of cryopreserved peripheral blood mononuclear cells for functional and phenotypic T-cell characterization. Clin. Vaccine Immunol 16, 1176–1186, https://doi.org/10.1128/CVI.00342-08 (2009).
Weinberg, A., Betensky, R. A., Zhang, L. & Ray, G. Effect of shipment, storage, anticoagulant, and cell separation on lymphocyte proliferation assays for human immunodeficiency virus-infected patients. Clin. Diagn. Lab Immunol 5, 804–807 (1998).
Kierstead, L. S. et al. Enhanced rates and magnitude of immune responses detected against an HIV vaccine: effect of using an optimized process for isolating PBMC. AIDS Res. Hum. Retroviruses 23, 86–92, https://doi.org/10.1089/aid.2006.0129 (2007).
Kreher, C. R., Dittrich, M. T., Guerkov, R., Boehm, B. O. & Tary-Lehmann, M. CD4+ and CD8+ cells in cryopreserved human PBMC maintain full functionality in cytokine ELISPOT assays. J. Immunol. Methods 278, 79–93 (2003).
Maecker, H. T. et al. Impact of cryopreservation on tetramer, cytokine flow cytometry, and ELISPOT. BMC. Immunol 6, 17, https://doi.org/10.1186/1471-2172-6-17 (2005).
Germann, A. et al. Temperature fluctuations during deep temperature cryopreservation reduce PBMC recovery, viability and T-cell function. Cryobiology 67, 193–200, https://doi.org/10.1016/j.cryobiol.2013.06.012 (2013).
Lemieux, J., Jobin, C., Simard, C. & Neron, S. A global look into human T cell subsets before and after cryopreservation using multiparametric flow cytometry and two-dimensional visualization analysis. J. Immunol. Methods 434, 73–82, https://doi.org/10.1016/j.jim.2016.04.010 (2016).
Kyu, S. Y. et al. Frequencies of human influenza-specific antibody secreting cells or plasmablasts post vaccination from fresh and frozen peripheral blood mononuclear cells. J. Immunol. Methods 340, 42–47, https://doi.org/10.1016/j.jim.2008.09.025 (2009).
World Medical Association Declaration of Helsinki: ethical principles for medical research involving human subjects. JAMA 310, 2191–2194, https://doi.org/10.1001/jama.2013.281053 (2013).
Skånland, S. S. Phospho Flow Cytometry with Fluorescent Cell Barcoding for Single Cell Signaling Analysis and Biomarker Discovery. J. Vis. Exp 140, e58386 (2018).
Friedman, A. A., Letai, A., Fisher, D. E. & Flaherty, K. T. Precision medicine for cancer with next-generation functional diagnostics. Nat. Rev. Cancer 15, 747–756, https://doi.org/10.1038/nrc4015 (2015).
Kotecha, N. et al. Single-cell profiling identifies aberrant STAT5 activation in myeloid malignancies with specific clinical and biologic correlates. Cancer Cell 14, 335–343, https://doi.org/10.1016/j.ccr.2008.08.014 (2008).
Acknowledgements
The authors are grateful to all the study participants, without whom this study would not have been possible. Martine Schrøder, Marianne Enger and Gladys Marie Tjørhom are acknowledged for their expert technical assistance. This work was supported by the Regional Health Authority for South-Eastern Norway, the Research Council of Norway, the Norwegian Cancer Society and Stiftelsen Kristian Gerhard Jebsen. S.S.S. was funded by a Career Development Research Fellowship from the Norwegian Cancer Society.
Author information
Authors and Affiliations
Contributions
J.U.H., K.T. and S.S.S. designed the research. J.U.H. and S.S.S. performed the experiments and analyzed the data. G.E.T. provided patient samples. J.U.H. and S.S.S. wrote the article. G.E.T., K.T. and L.A.M. contributed to data analysis and writing the paper. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hermansen, J.U., Tjønnfjord, G.E., Munthe, L.A. et al. Cryopreservation of primary B cells minimally influences their signaling responses. Sci Rep 8, 17651 (2018). https://doi.org/10.1038/s41598-018-36121-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-36121-9
Keywords
This article is cited by
-
Extracellular vesicles from type-2 macrophages increase the survival of chronic lymphocytic leukemia cells ex vivo
Cancer Gene Therapy (2024)
-
Immunophenotyping with (phospho)protein profiling and fluorescent cell barcoding for single-cell signaling analysis and biomarker discovery
npj Precision Oncology (2024)
-
Shedding Light on Intracellular Proteins using Flow Cytometry
Cell Biochemistry and Biophysics (2024)
-
A tumor microenvironment model of chronic lymphocytic leukemia enables drug sensitivity testing to guide precision medicine
Cell Death Discovery (2023)
-
Standardized assays to monitor drug sensitivity in hematologic cancers
Cell Death Discovery (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
