
Abstract
The effect of feeding competition on foraging efficiency is an important link between ecological factors and the social organization of gregarious species. We examined the effects of group size on daily travel distances, activity budgets, and energy intake of mountain gorillas in Rwanda. We measured daily travel distances of five groups, activity budgets of 79 gorillas in nine groups, and energy intake data for 23 adult females in three groups over a 16-month period. Travel distances and the proportion of time spent traveling increased with size for most groups, which would be expected if their foraging efficiency is limited by intragroup feeding competition. However, travel distances and times decreased for the largest group, which also had higher energy intake rates than intermediate sized groups. The improved foraging efficiency of the largest group may be explained by advantages in intergroup contest competition. The largest group had much lower home range overlap than the other study groups which may be due to groups avoiding one another as a result of male mating competition. Collectively, our results indicate that intermediate sized groups had the lowest foraging efficiency and provide a new twist on the growing evidence of non-linear relationships between group size and foraging efficiency in primates.
Similar content being viewed by others
Introduction
Feeding competition has been considered one of the most important links between ecological factors and the social organization of gregarious species1,2. Socioecological models predict that ecological factors primarily determine the foraging efficiency, reproductive success, and distribution of females; which in turn will influence the distribution of males3,4. The relationships between foraging efficiency and group size can be influenced by both scramble competition and contest competition, which can occur both within groups and between groups5,6.
Within-group scramble competition (WGS) is predicted to create a positive correlation between group size versus daily travel times and distances7,8. If food is distributed into discrete patches, then large groups may need to visit more patches to obtain the same amount of food per individual9,10. When food is more evenly dispersed, individuals in larger groups may have a greater probability of encountering a food site where another group member has already eaten, so they may need to travel farther to find fresh sites11,12.
If foraging groups advance through their habitat by “pushing forward” the members in the front, then WGS and/or within-group contest competition (WGC) could also contribute to a positive correlation between group size and travel9,13,14. WGC can involve aggressive displacement by dominant individuals, or subordinates may move preemptively to avoid such aggression15,16. Such competition is typically associated with rank related differences in foraging efficiency and reproductive success17,18. WGC may be more intense when foods are small enough to be monopolized and take a long time to consume19,20. We will use the term “intragroup feeding competition” to encompass both WGS and WGC.
Between-group contest competition (BGC) may create a negative correlation between group size and travel7,8. Small groups may travel farther to avoid encounters with large groups, and they may be displaced from food patches where such encounters occur21,22. BGC is predicted when patches of food are large enough to accommodate an entire group, yet small enough that the group can exclude other rivals1,23. For species with sexual dimorphism, the quality and quantity of males may be more important for winning contests between groups than the total number of members in a group, and feeding competition may overlap with male mating competition24,25,26,27. We will use the term “intergroup contest competition” to refer to BGC while acknowledging a potential influence of mating competition.
The fourth permutation of feeding competition, between-group scramble competition (BGS), is not expected to involve correlations with group size, but may lead to lower foraging efficiency at higher population density5,28. Increasing population density can reduce the abundance of food, which may intensify the other types of feeding competition29. For all four types of competition, lower foraging efficiency may involve greater depletion of food sites, and individuals may settle for closer sites with lower quality, resulting in lower food intake rates8,30. To compensate for lower intake rates, as well as the energetic costs of greater travel, individuals may spend more time feeding28,31.
A combination of intragroup feeding competition and BGC has been evoked to explain why optimal foraging efficiency occurs at intermediate group sizes32,33. Intragroup feeding competition can reduce the foraging efficiency for large groups, while BGC can reduce the foraging efficiency of small groups. Furthermore, an upper limit for optimal group sizes has been attributed to infanticide, and a lower limit has been attributed to predation risks1,21,34. Nonetheless, group sizes can vary considerably both within and among species, and the consequences of those variations have not been well quantified14.
Here we investigate the effects of group size on the foraging efficiency of mountain gorillas in the Virunga volcano region. The Virunga mountain gorillas are expected to have weak feeding competition, primarily because their food is abundant35,36. Distances between patches are small, which limits the extra travel requirements if larger groups must visit more patches37,38. Food sites are numerous and densely distributed within those patches, so extra foraging costs are minimal if some sites have already been depleted, and individuals have little incentive to compete for any particular spot39,40,41. The Virunga mountain gorillas have been considered unlikely candidates for BGC because intergroup encounters have traditionally been infrequent and intergroup aggression is mainly limited to male mating competition42,43,44. Male mating competition may be more influential than BGC, as groups try to retain females by avoiding encounters with potential rivals, and daily travel distances can be longer after encounters occur35,45,46,47. Hypothetically, competition among males could also be based on resource defense, but significant evidence has not been found to support such a possibility for gorillas48,49.
Previous studies have supported expectations that group size has little influence on the foraging efficiency of this population. Inconsistent results have emerged from analyses of daily travel distance versus group size35,45. Time spent feeding was significantly longer in larger groups, but the increase was only three percentage points across a three-fold variation in group size39. Dominance rank did also not have a significant effect on the energy intake rates or the proportion of time spent traveling, and rank-related differences in female reproductive success might be due to female quality rather than contest competition41,50,51. Thus, both types of intragroup feeding competition (WGS and WGC) appear to be weak in this species44. Subsequent to most of the previous studies of group size, however, some groups have become larger and intergroup encounters have become more frequent, which increases the potential for both intragroup feeding competition and intergroup contest competition52.
To examine the effects of group size on foraging efficiency, we looked at the daily travel distances, activity budgets, and energy intake rates of the Virunga mountain gorillas. If intragroup feeding competition is the main influence on foraging efficiency, then we expect larger groups to have longer travel distances and times, as well as lower energy intake rates. We predict the opposite patterns if intergroup contest competition predominates. If both types of competition are determining the optimal group size for this population, then foraging efficiency should be highest at intermediate sizes, with lower values for smaller and larger groups (if the probability of winning is based on group size and male number). We discuss our results within the context of the socioecological theories about foraging efficiency and social organization.
Results
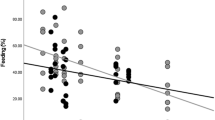
Daily travel distances averaged 712.5 ± 339.6 meters (n = 448 group-days in five groups). The daily travel distances showed a significant quadratic relationship with group size (Table 1a). The linear mixed model that we used for statistical analysis predicted longer travel distances at intermediate group sizes, however, so the curvature is in the opposite direction of expectations (Fig. 1). Visual inspection suggests that the quadratic term might be excessively sensitive to the data from the largest or smallest group. The p-value remained significant when we removed the largest group from the analysis, however, which suggests that the effects of group size were already leveling off for the other groups. The p-value also remained significant when we removed the smallest group (or any of the other groups). The linear term for group size was also significant in the original model, but not when we removed the quadratic term (p = 0.10).

Group size versus the daily travel distance (meters). Each data point represents one day that the group travel was measured. The line is based on a bivariate regression between daily travel versus group size and size-squared. Detailed results from a more complete model are shown in Table 1a.
Using a larger dataset that included four more groups and 1000 more group-days, the results for the proportion of time spent traveling were similar to the daily travel distances (Fig. 2a). The quadratic term for group size was significant, and the model again predicted greater travel for intermediate group sizes (Table 1b). The observed pattern remained significant when we removed the smallest group, but not when we removed the largest group.

Group size versus the proportion of time that gorillas spent traveling (a) or feeding (b). For purposes of clarity, the results are aggregated into one data point for each group in each habitat, and data point with less than 100 scans are omitted. The size of each symbol reflects the sample size. The legend at the bottom of (b) indicates the group ID for each data point.
The gorillas spent an average of 45.4% ± 3.2% of their time feeding (Table 2). The proportion of time spent feeding showed a significant negative correlation with group size, and the quadratic term was not significant (Table 1c, Fig. 2b). The linear term remained significant if we removed group BWE, KUY, or UGE; it became a statistical trend (0.05 < p < 0.10) if we removed group INS, ISA, NTA, TIT, or URU; and the p-value increased to 0.57 if we removed the largest group.
Adult females in the largest group had an average energy intake rate of 116.0 ± 128.2 per minute, which is significantly higher than 81.7 ± 63.9 kJ per minute for two intermediate sized groups (Table 1d). Adult females in the largest group also obtained a significantly higher amount of energy per food site (246.3 ± 286.0 kJ versus 176.8 ± 181.2 kJ for the intermediate sized groups). Adult females in the largest group traveled an average of 4.3 ± 4.7 meters between food sites, which is not significantly different from 4.9 ± 6.8 meters for the intermediate sized groups (Table 1f). The latter two results indicate that the largest group could visit fewer food sites to obtain the same amount of energy as the intermediate size groups, and it did not need to travel farther between those sites, which again suggests that its travel requirements should be lower.
Hypothetically, the higher energy intake rates of the largest group could be distributed across all plant species, or the group could have greater access to more favorable species. To separate those two hypotheses, we ran a post hoc analysis with a subset of the food sites in which a single plant species accounted for at least 80% of the total energy intake. The post hoc analysis was similar to our original model of energy intake rates, except that we added a random effect variable to control for the main plant species of the food site. The category variable for group size was no longer significant in the post hoc analysis (p = 0.22), so we did not find support for the hypothesis that the higher energy intake rates of largest group are distributed across all plant species. Instead, the largest group may have greater access to more favorable species. For example, gorillas had an average energy intake rate of 226.7 ± 196.3 kJ per minute while feeding on Rubus spp. versus only 85.1 ± 74.9 for the other plant species. Rubus accounted for 30.1% of the total energy intake by the largest group, versus only 6.3% for the other two groups.
Discussion
Our analyses of daily travel distances and travel times showed a significant quadratic relationship with group size. Foraging efficiency decreased with size among most groups, but then began to increase for the largest group. The largest group also had significantly higher energy intake rates than the intermediate sized groups, which is consistent with the quadratic pattern for our other measures of foraging efficiency. One potential interpretation of these results is that foraging efficiency of most groups was limited by intragroup feeding competition, which the larger group may have been able to mitigate through intergroup competition23,53.
Non-linear relationships between foraging efficiency versus group size have also been reported for four groups of woolly monkeys (Lagothrix lagothricha) and five groups of savanna baboons (Papio cynocephalus), but their patterns were generally in the opposite direction of our study, because foraging efficiency was highest for their intermediate sized groups32,33. From a mathematical perspective, the contrasting patterns are represented by the sign of the quadratic term in the statistical models. The significant quadratic term was negative in our analyses of travel times and daily travel distances, but it was positive in four of five multivariate analyses for savanna baboons32. A literature review of seventeen studies showed only one significant (positive) quadratic term, and it merely reflected a slight curvature rather than a full U-shaped pattern7,54. The rarity of significant quadratic patterns has been attributed to insufficient sample sizes (particularly for smaller groups) as well as weak socioecological influences32.
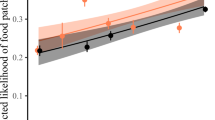
From a socioecological perspective, the sign of the quadratic term may reflect the ways that group size affects intragroup feeding competition versus intergroup contest competition (Fig. 3). For example, it could become increasingly difficult for larger groups to avoid the effects of intragroup competition (Fig. 3a), if their ability to increase group spread is limited by the distribution of food23,35. Alternatively, groups might not adjust to such competition until it approaches a threshold level (Fig. 3b), if the benefits of subgrouping must be balanced against the risk of predation55,56. Similarly, increases in group size might reduce the costs of intergroup contest competition most dramatically for small groups (Fig. 3a), or those cost reductions might not accelerate until groups become larger than average (Fig. 3b). The relationship between group size and intergroup contest competition could even form an inverted U-shaped pattern, with the greatest costs incurred by mid-sized groups57. Further study is needed to disentangle these nuances of each type of competition5,29.

Hypothetical effects of group size on the overall costs of feeding competition (thick line), which equals the combined impact of competition within groups (circles) versus between groups (triangles). The overall costs have a U-shaped pattern if the second derivative is positive for competition both within groups and between groups (a). An inverted U-shape arises if the second derivative is negative for both types of competition (b). If the two types of competition have opposite curvatures, then the overall pattern will depend on which curvature is stronger (not shown). If both types of competition have no curvature, then the combined effect will also be linear.
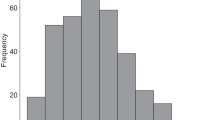
If foraging efficiency is the primary influence on individual fitness and behavior, then the aforementioned U-shaped patterns for woolly monkeys and savanna baboons should lead to narrower group size distributions, which would be analogous to stabilizing selection for genetic traits58,59. Small groups would benefit from getting larger; and large groups would benefit from getting smaller. In contrast, the inverted-U pattern in this study should lead to a broader (and ultimately bimodal) group size distribution, which would be analogous to disruptive stabilization60,61. A comparative study of group size distributions has not been reported for primates, so it is unclear whether the variance is especially broad for Virunga mountain gorillas (Fig. 4). Their group size distribution is not bimodal, however, which suggests that our results are not typical of the overall population, or that foraging efficiency is less influential than other factors such as the quality and quantity of adult males44,62,63,64. The breadth of group size distributions can also depend on variability in ecological conditions, as well as the flexibility of individuals to adjust to such conditions, particularly via patterns of dispersal and group fissioning32,58.

Group size distribution of the Virunga mountain gorillas. The y-axis indicates the number of groups that were recorded within each size category during eight censuses of the entire population from 1971–201074,88,89,90,91,92,93,94,95. The numbers along the x-axis represent the largest group size that was included in each category.
Previous studies of this population did not find the inverted-U pattern, perhaps because they did not encompass the size of our largest group35,45. Nonetheless, over the past 50 years of research and ecotourism in the Virunga Volcano region, seven of the 39 (18%) habituated groups have contained at least 25 gorillas, which is more than two standard deviations beyond the average value for the other habituated groups65. Thus the tail in Fig. 4 does not represent an isolated incident, suggesting that large groups may be beneficial but limited in occurrence by other factors such as the ability of a single male to attract and retain a large number of females (mate competition). Group size was not significantly correlated with female reproductive success in a study that included five of those seven large groups, which supports our finding that such groups are not constrained by foraging efficiency44. The probability of female emigration was significantly lower in larger groups, which may indicate that females prefer to reproduce in such groups64. The latter result was merely interpreted as further evidence that intragroup feeding competition is minimal, yet it could also suggest that larger groups have an advantage in intergroup competition.
The largest group had only two known encounters with other groups throughout the study, so any advantage in intergroup contest competition did not arise by routinely displacing its competitors from patches of food. Instead, the largest group had far less home range overlap than the other study groups (~20% versus 90%), which is more consistent with an avoidance mechanism of intergroup contest competition52,66. Among the other groups, greater home range overlap likely caused more BGS, which may have exacerbated WGS. The home range comparisons do not include any overlap with non-study groups, but the largest group had no known encounters with those groups during this study, so its overlap with their home ranges may have been minor. Other large groups have also had low home range overlap and/or low rates of intergroup encounters, but statistical correlations have not been reported between those parameters and group size52,67. Groups may avoid each other by travelling farther after encounters, by using long-distance signaling mechanisms (e.g. chest beats), and by moving away from areas where foraging has already occurred68,69,70.
The avoidance mechanism is probably driven by male mating competition, which predominates the behavioral interactions when mountain gorilla groups meet42,47,70. Such avoidance may help males to retain their females, who disperse only during encounters with other adult males43,71. In some cases, a large group may even avoid smaller groups, because it has more females at stake during such encounters35,45. Indeed, the home range of the largest group gradually shifted farther from the other study groups between 2005–2011, which may indicate that it was avoiding an area where group density was increasing52. The smaller groups have not expanded into the same area as the largest group, however, perhaps because they wanted to avoid intergroup competition with it. Thus, the largest group generally had the lowest degree of home range overlap throughout the ten years prior to our study52. Analysis that would reveal if larger or smaller groups are avoiding each other in this population remains to be done. Just as dominance hierarchies among individuals are used to evaluate intragroup contest competition, evidence of a dominance hierarchy among groups could help to clarify the effects of intergroup contest competition on ranging patterns, but previous studies of mountain gorillas do not indicate an obvious winner of most encounters42,47.
Interestingly, time spent feeding showed a significant negative correlation with group size. Similarly, the recent study of baboons showed a linear relationship between group size and time spent foraging (feeding and moving), despite a quadratic relationship for other measures of foraging efficiency32. Again, however, their results were in the opposite direction of ours, because the foraging time of baboons increased with group size32. Their positive correlation could indicate an influence of group size on intragroup feeding competition, whereas our negative correlation is more consistent with theoretical predictions for intergroup contest competition7,8. The feeding time of mountain gorillas seems to be more sensitive to differences in energy intake rates than differences in the energy requirements from travel36,41,72. If so, then our negative correlation could be interpreted to suggest that larger groups had greater access to areas that facilitate higher energy intake rates.
The higher energy intake rate of the largest group arose mainly from the consumption of Rubus plants, which are especially concentrated on the western edge of our study site. Thus, for the largest group, shifting its range slightly to the west, perhaps to avoid an area of greatly increased group density, also exposed it to high quality habitat. In which case, its higher energy intake rate could be a result of being in the right place at the right time rather than outcompeting smaller groups. Rubus has increased in biomass over the last 20 years (from being present in 1% to 26% of sampled plots), which may also help to explain why our results do not match previous studies35,45,73. Rubus does not seem to entirely explain our quadratic relationships, however, because the results for daily travel distance remained significant even when we excluded the largest group.
The population of the Virunga mountain gorillas has doubled in size over the past few decades, which provides an opportunity to evaluate the impact of between-group scramble competition upon their foraging patterns and food availability74. The average daily travel distance in this study was approximately 25–40% longer than one group in the late 1970s, and the proportion of time spent traveling was 50–70% greater than previous data from four groups35,39,75. Home range sizes have remained unchanged45,52. The biomass density declined by more than 50% for two of their most frequently consumed foods, but it increased for three of their other top-five foods73. Some of these apparent changes could be due to differences in methodology, and collectively the results do not indicate an immediate threat to continued population growth, but they highlight areas that warrant continued monitoring for this critically endangered species52,65,73. The changes in population density and food density could also intensify both intragroup feeding competition and intergroup contest competition, thereby contributing to the quadratic relationships that were detected in this study.
Methods
Study population
We studied nine groups of habituated mountain gorillas that are monitored by the Dian Fossey Gorilla Fund’s Karisoke Research Center in the Volcanoes National Park of Rwanda (Table 2). The Virunga Volcano region contains a range of habitats that vary along an altitudinal gradient from the bamboo zone below 2800 meters to afro-montane vegetation above 3600 meters36,39,45,73. Between those two extremes, the additional habitats are the mixed species montane forest with an understory of Mimulopsis spp. at 2500–2700 m, the Hagenia- Hypericum woodland found in the saddle between two volcanoes at 2800–3300 m, the Hypericum woodland (a.k.a. brush ridge) on the volcano slopes at 3000–3300 m, the herbaceous vegetation zone containing dense tall herbs with no tree cover at 2800–3300 m, and the subalpine zone containing stands of Lobelia stuhlmannii and thickets of Rubus spp. at 3300–3600 m. The Virunga mountain gorillas primarily feed on herbaceous vegetation that is available year-round, except for bamboo shoots which were consumed from October through December during this study76,77,78.
Data collection
We measured the daily travel distance of five groups from September 2009 to September 2010 (n = 7.5 ± 1.8 measurements per group-month, Table 2). Each adult gorilla makes a nest at night and the group creates a trail of trampled vegetation throughout the day. We followed the main trail of the group, and took readings every 30 seconds using the “track log” function of a GPS (accurate to within ca. five meters). The daily travel distance was calculated as the sum of distances between each set of coordinates in ArcGIS®. Some nest-to-nest distances covered several altitudinal zones and were corrected for terrain using a Digital Elevation Model with 50 m contours (courtesy of M. Gray). Daily travel may be influenced by attempts to avoid other groups and/or solitary males68, so we excluded eight days when the focal group was involved in an intergroup encounter.
We measured the activity budgets of 79 gorillas in nine groups from October 2009 through December 2010. During 50 minute focal sessions of each individual, the protocol involved taking an instantaneous scan every 10 minutes to record the activity (feeding, traveling, resting, grooming, or playing). The analyses were limited to gorillas who were at least eight years old, which is when females are typically considered adults. Males are not considered adults until age twelve, but by age eight they are already as large as adult females, and activity budgets are similar among age-sex classifications39. As required by the Rwanda Development Board, observations of the gorillas were limited to four hours per day to minimize anthropogenic disturbance.
Energy intake data was taken from Grueter (2016), which analyzed food sites for 23 adult females in three groups from October 2009 through December 2010. The energy intake rate for each food site equaled the total energy intake, divided by the food site residence time (FSRT). The FSRT was defined as the elapsed time from when a female settled at a food site and commenced eating, until she stopped eating and/or moved more than one meter72,79. The energy intake for each food site equaled the number of food “units” consumed during the FSRT (e.g., one leaf or stem of a specific plant species), multiplied by the average energy content for each type of unit80,81. The energy content for 33 types of food units from 25 species was calculated from nutritional analyses of crude protein, lipids, neutral detergent fiber, and total ash; as conducted by the Leibniz Institute for Zoo and Wildlife Research in Berlin. For the 23 adult females in the three study groups, we collected 657 hours of data with the energy intake protocol and 931 focal hours with the activity budget protocol, which represent 28.5 and 39.9 hours per female, respectively.
Statistical analyses
To analyze the daily travel distances, we ran a linear mixed model with one data point for each day that each group was measured (n = 448 data points). The response variable was the distance that the group traveled (meters). The predictor variables were the group size (the number of weaned individuals) and group size squared. A positive term for group size squared would represent a U-shaped pattern, with the shortest daily travel distances occurring at intermediate group sizes, and longer travel distances for the smallest and largest groups. A negative term for group size squared would represent an inverted U-shaped pattern, with the longest daily travel distances occurring at intermediate group sizes, and shorter travel distances for the smallest and largest groups. We included the daily rainfall as a control variable because the gorillas typically stop travelling when it is raining35,82. We also added a control variable to account for potential temporal autocorrelation among the data points within each group83. The random effect variables were the group ID and the habitat ID. The latter random effect variable may help to control for spatial variations in food density11,35,39. To account for the seasonality of bamboo shoots, the random effect variable of habitat ID included one category for its habitat during the bamboo season, and another category for its habitat during the rest of the year.
To examine the proportion of time that gorillas spent traveling, we ran a linear mixed model with one data point for each habitat that each focal gorilla used on each day (n = 3866 data points). For example, if the group used two habitats during focal observations of one gorilla, and only one habitat during the focal observations of another gorilla, then the model would have three data points for that day. For each data point, the response variable equaled the duration of time that the gorilla spent traveling, with an offset control variable for the total duration of their focal time (traveling plus all other activities). Our approach is similar to using proportion of time spent traveling as the response variable, but the control variable helps to avoid excessive influence from data points that are based on brief observations84. The predictor variables were the group size and group size squared. The control variables were the daily rainfall and the term for temporal autocorrelation. The random effect variables were the gorilla ID, group ID, and the habitat ID. We ran a similar model to examine the proportion of time that gorillas spent feeding.
To analyze the energy intake rates, we ran a linear mixed model with one data point for each food site (n = 3342 data points). The response variable was the total energy intake at the food site (kJ), with an offset control variable for the time spent feeding at the site (minutes). The number of groups was insufficient to look for nonlinear effects of group size (or even a strong test of linear effects), so the predictor variable merely tested whether the largest group (PAB) was significantly different from two intermediate sized groups (NTA and BWE). The random effect variables were the gorilla ID, group ID, and the habitat ID; and we added a control variable for temporal autocorrelation. We ran a similar model to analyze the energy intake per food site. The response variable again was the total energy intake at the food site (kJ), but we omitted the control variable for the time spent feeding at the site. We ran a third model in which the response variable was the distance travelled between consecutive food sites. For all three models, the random effect for gorilla ID can help to control for female rank, which did not show a significant effect on these response variables in the previous study of this data41.
All linear mixed models were run with a Gaussian error structure and identity link while using the “lmer” function of the “lme4” package in R85. We used log or exponential transformations of the response variables as needed to obtain more normally distributed residuals. The predictor variables for group size were log transformed to provide a more uniform distribution of values, and then were also standardized so they each had a mean value of “0” and a standard deviation of “1”86. We obtained p-values for each predictor variable by using the “drop1” function, which performs likelihood ratio tests to compare the full model versus a set of reduced models that exclude one predictor at a time87.
Data Availability
The datasets generated during and analyzed during the current study are available from the corresponding author on reasonable request.
References
van Schaik, C. P. The ecology of social relationships amongst female primates in Comparative socioecology: the behavioural ecology of humans and other mammals (eds Standon, V. & Foley, R. A.) 195–218 (Blackwell Scientific Publications, Oxford, 1989).
Clutton-Brock, T. & Janson, C. Primate socioecology at the crossroads: Past, present, and future. Evol. Anthropol. 21, 136–150 (2012).
Kappeler, P. M. & van Schaik, C. P. Evolution of primate social systems. Int. J. Primatol. 23, 707–740 (2002).
Koenig, A., Scarry, C. J., Wheeler, B. C. & Borries, C. Variation in grouping patterns, mating systems and social structure: what socio-ecological models attempt to explain. Phil. Trans. Roy. Soc. B 368 (2013).
Janson, C. H. & van Schaik, C. P. Recognizing the many faces of primate food competition - methods. Behaviour 105, 165–186 (1988).
Koenig, A. Competition for resources and its behavioral consequences among female primates. Int. J. Primatol. 23, 759–783 (2002).
Janson, C. H. & Goldsmith, M. L. Predicting group size in primates - foraging costs and predation risks. Behav. Ecol. 6, 326–336 (1995).
Majolo, B., Vizioli, A. D. & Schino, G. Costs and benefits of group living in primates: group size effects on behaviour and demography. Anim. Behav. 76, 1235–1247 (2008).
Chapman, C. A. & Chapman, L. J. Constraints on group size in red colobus and red-tailed guenons: Examining the generality of the ecological constraints model. Int. J. Primatol. 21, 565–585 (2000).
Snaith, T. V. & Chapman, C. A. Red colobus monkeys display alternative behavioral responses to the costs of scramble competition. Behav. Ecol. 19, 1289–1296 (2008).
Gillespie, T. R. & Chapman, C. A. Determinants of group size in the red colobus monkey (Procolobus badius): an evaluation of the generality of the ecological-constraints model. Behav. Ecol. Sociobiol. 50, 329–338 (2001).
Teichroeb, J. A. & Sicotte, P. Test of the ecological constraints model on ursine colobus monkeys (Colobus vellerosus) in Ghana. Am. J. Primatol. 71, 49–59 (2009).
van Schaik, C. P., van Noordwijk, M. A., Deboer, R. J. & Dentonkelaar, I. The effect of group-size on time budgets and social behavior in wild long-tailed macaques (Macaca-Fascicularis). Behav. Ecol. Sociobiol. 13, 173–181 (1983).
Chapman, C. A. & Chapman, L. J. Determinants of group size in primates: the importance of travel costs in On the move - how and why animals travel in groups (eds Boinski, S. & Garber, P.) 24–42 (University of Chicago Press, Chicago, 2000).
Wheeler, B. C., Scarry, C. J. & Koenig, A. Rates of agonism among female primates: a cross-taxon perspective. Behav. Ecol. (2013).
Thouless, C. R. Feeding competition between grazing red deer hinds. Anim. Behav. 40, 105–111 (1990).
Vogel, E. R. Rank differences in energy intake rates in white-faced capuchin monkeys, Cebus capucinus: the effects of contest competition. Behav. Ecol. Sociobiol. 58, 333–344 (2005).
Schülke, O. & Ostner, J. Ecological and social influences on sociality in The evolution of primate societies (eds Mitani, J., Call, J., Kappeler, P. M., Palombit, R. A. & Silk, J. B.) 195–219 (University of Chicago Press, Chicago, 2012).
Hirsch, B. T. Costs and benefits of within-group spatial position: A feeding competition model. Quart. Rev. Biol. 82, 9–27 (2007).
Isbell, L. A., Pruetz, J. D. & Young, T. P. Movements of vervets (Cercopithecus aethiops) and patas monkeys (Erythrocebus patas) as estimators of food resource size, density, and distribution. Behav. Ecol. Sociobiol. 42, 123–133 (1998).
Wrangham, R. W. An ecological model of female-bonded primate groups. Behaviour 75, 262–300 (1980).
Markham, A. C., Guttal, V., Alberts, S. C. & Altmann, J. When good neighbors don’t need fences: temporal landscape partitioning among baboon social groups. Behav. Ecol. Sociobiol. 67, 875–884 (2013).
Sterck, E. H. M., Watts, D. P. & van Schaik, C. P. The evolution of female social relationships in nonhuman primates. Behav. Ecol. Sociobiol. 41, 291–309 (1997).
Scarry, C. J. Between-group contest competition among tufted capuchin monkeys, Sapajus nigritus, and the role of male resource defence. Anim. Behav. 85, 931–939 (2013).
Harris, T. R., Chapman, C. A. & Monfort, S. L. Small folivorous primate groups exhibit behavioral and physiological effects of food scarcity. Behav. Ecol. 21, 46–56 (2010).
Cowlishaw, G. Behavioral patterns in baboon group encounters - the role of resource competition and male reproductive strategies. Behaviour 132, 75–86 (1995).
Richter, C., Heesen, M., Nenadic, O., Ostner, J. & Schülke, O. Males matter: increased home range size is associated with the number of resident males after controlling for ecological factors in wild assamese macaques. Am. J. Phys. Anthropol. 159, 52–62 (2016).
Snaith, T. V. & Chapman, C. A. Primate group size and interpreting socioecological models: do folivores really play by different rules? Evol. Anthropol. 16, 94–106 (2007).
Isbell, L. A. Contest and scramble competition: patterns of female aggression and ranging behavior in primates. Behav. Ecol. 2, 143–155 (1991).
Charnov, E. L. Optimal foraging, marginal value theorem. Theor. Pop. Biol. 9, 129–136 (1976).
Saj, T. L. & Sicotte, P. Scramble competition among Colobus vellerosus at Boabeng-Fiema, Ghana. Int. J. Primatol. 28, 337–355 (2007).
Markham, A. C., Gesquiere, L. R., Alberts, S. C. & Altmann, J. Optimal group size in a highly social mammal. Proc. Natl. Acad. Sci. 112, 14882–14887 (2015).
Stevenson, P. R. & Castellanos, M. C. Feeding rates and daily path range of the Colombian woolly monkeys as evidence for between- and within-group competition. Folia Primatol. 71, 399–408 (2000).
Steenbeek, R. & van Schaik, C. P. Competition and group size in Thomas’s langurs (Presbytis thomasi): the folivore paradox revisited. Behav. Ecol. Sociobiol. 49, 100–110 (2001).
Watts, D. P. Strategies of habitat use by mountain gorillas. Folia Primatol. 56, 1–16 (1991).
Wright, E. et al. Energetic responses to variation in food availability in the two mountain gorilla populations (Gorilla beringei beringei). Am. J. Phys. Anthropol. 158, 487–500 (2015).
Watts, D. P. Relations between group size and composition and feeding competition in mountain gorilla groups. Anim. Behav. 33, 72–85 (1985).
Altmann, S. A. Baboons, Space, Time, and Energy. Am. Zool. 14, 221–248 (1974).
Watts, D. P. Environmental influences on mountain gorilla time budgets. Am. J. Primatol. 15, 195–211 (1988).
Watts, D. P. Agonistic relationships between female mountain gorillas (Gorilla gorilla beringei). Behav. Ecol. Sociobiol. 34, 347–358 (1994).
Grueter, C. C. et al. Causes, mechanisms, and consequences of contest competition among female mountain gorillas in Rwanda. Behav. Ecol. 27, 766–776 (2016).
Sicotte, P. Intergroup encounters and female transfer in mountain gorillas - influence of group composition on male behavior. Am. J. Primatol. 30, 21–36 (1993).
Sicotte, P. Female mate choice in mountain gorillas in Mountain gorillas: three decades of research at Karisoke (eds Robbins, M., Sicotte, P. & Stewart, K.) 59–88 (Cambridge University Press, Cambridge, 2001).
Robbins, M. M., Robbins, A. M., Gerald-Steklis, N. & Steklis, H. D. Socioecological influences on the reproductive success of female mountain gorillas (Gorilla beringei beringei). Behav. Ecol. Sociobiol. 61, 919–931 (2007).
Watts, D. P. Long term habitat use by mountain gorillas (Gorilla gorilla beringei). 1. consistency, variation, and home range size and stability. Int. J. Primatol. 19, 651–680 (1998).
Fossey, D. Observations on the home range of one group of mountain gorillas (Gorilla gorilla beringei). Anim. Behav. 22, 568–581 (1974).
Robbins, M. M. & Sawyer, S. C. Intergroup encounters in mountain gorillas of Bwindi Impenetrable National Park, Uganda. Behaviour 144, 1497–1519 (2007).
Breuer, T. et al. Variance in the male reproductive success of western gorillas: acquiring females is just the beginning. Behav. Ecol. Sociobiol. 64, 515–528 (2010).
Mirville, M. O. The causes and consequences of intergroup interactions in mountain gorillas (Gorilla beringei beringei), PhD Thesis, University of Western Australia, Perth, Australia (2018).
Robbins, A. M., Robbins, M. M. & Fawcett, K. Maternal investment of the Virunga mountain gorillas. Ethology 113, 235–245 (2007).
Robbins, A. M., Stoinski, T., Fawcett, K. & Robbins, M. M. Lifetime reproductive success of female mountain gorillas. Am. J. Phys. Anthropol. 146, 582–593 (2011).
Caillaud, D., Ndagijimana, F., Giarrusso, A. J., Vecellio, V. & Stoinski, T. S. Mountain gorilla ranging patterns: Influence of group size and group dynamics. Am. J. Primatol. 76, 730–746 (2014).
Janson, C. H. Intra-specific food competition and primate social-structure - a synthesis. Behaviour 105, 1–17 (1988).
Waser, P. Feeding, ranging, and group size in the mangabey Cercocebus albigena in Primate ecology: studies of feeding and ranging behavior in lemurs, monkeys, and apes (ed. Clutton-Brock, T.) 183–222 (Academic Press, London, 1977).
Chancellor, R. L. & Isbell, L. A. Female grooming markets in a population of gray-cheeked mangabeys (Lophocebus albigena). Behav. Ecol. 20, 79–86 (2009).
Arlet, M. E. & Isbell, L. A. Variation in behavioral and hormonal responses of adult male gray-cheeked mangabeys (Lophocebus albigena) to crowned eagles (Stephanoaetus coronatus) in Kibale National Park, Uganda. Behav. Ecol. Sociobiol. 63, 491–499 (2009).
Teichroeb, J. A. & Sicotte, P. Cascading competition: the seasonal strength of scramble influences between-group contest in a folivorous primate. Behav. Ecol. Sociobiol. 72 (2018).
Brown, C. R., Brown, M. B., Roche, E. A., O’Brien, V. A. & Page, C. E. Fluctuating survival selection explains variation in avian group size. Proc. Natl. Acad. Sci. 113, 5113–5118 (2016).
Kingsolver, J. G. & Pfennig, D. W. Patterns and power of phenotypic selection in nature. Bioscience 57, 561–572 (2007).
Rueffler, C., Van Dooren, T. J. M., Leimar, O. & Abrams, P. A. Disruptive selection and then what? TREE 21, 238–245 (2006).
Massol, F., Duputie, A., David, P. & Jarne, P. Asymmetric patch size distribution leads to disruptive selection on dispersal. Evolution 65, 490–500 (2011).
Caillaud, D., Levrero, F., Gatti, S., Menard, N. & Raymond, M. Influence of male morphology on male mating status and behavior during interunit encounters in western lowland gorillas. Am. J. Phys. Anthropol. 135, 379–388 (2008).
Breuer, T., Robbins, A. M., Boesch, C. & Robbins, M. M. Phenotypic correlates of male reproductive success in western gorillas. J. Hum. Evol. 62, 466–472 (2012).
Robbins, A. M., Stoinski, T. S., Fawcett, K. A. & Robbins, M. M. Socioecological influences on the dispersal of female mountain gorillas - evidence of a second folivore paradox. Behav. Ecol. Sociobiol. 63, 477–489 (2009).
Robbins, M. M. et al. Extreme conservation leads to recovery of the Virunga mountain gorillas. Plos One 6, e19788 (2011).
Waser, P. M. Cercocebus albigena Site attachment, avoidance, and intergroup spacing. Am. Nat. 110, 911–935 (1976).
Robbins, A. M., Stoinski, T. S., Fawcett, K. A. & Robbins, M. M. Does dispersal cause reproductive delays in female mountain gorillas? Behaviour 146, 525–549 (2009).
Watts, D. P. The influence of male mating tactics on habitat use in mountain gorillas (Gorilla gorilla beringei). Primates 35, 35–47 (1994).
Crofoot, M. C. The cost of defeat: Capuchin groups travel further, faster and later after losing conflicts with neighbors. Am. J. Phys. Anthropol. 152, 79–85 (2013).
Seiler, N., Boesch, C., Mundry, R., Stephens, C. & Robbins, M. M. Space partitioning in wild, non-territorial mountain gorillas: the impact of food and neighbours. Roy. Soc. Op. Sci. 4 (2017).
Harcourt, A. H., Stewart, K. S. & Fossey, D. Male emigration and female transfer in wild mountain gorilla. Nature 263, 226–227 (1976).
Wright, E., Robbins, A. M. & Robbins, M. M. Dominance rank differences in the energy intake and expenditure of female Bwindi mountain gorillas. Behav. Ecol. Sociobiol. 68, 957–970 (2014).
Grueter, C. C. et al. Long-term temporal and spatial dynamics of food availability for endangered mountain gorillas in Volcanoes National Park, Rwanda. Am. J. Primatol. 75, 267–280 (2013).
Gray, M. et al. Genetic census reveals increased but uneven growth of a critically endangered mountain gorilla population. Biol. Cons. 158, 230–238 (2013).
Watts, D. Foraging strategy and socioecology of mountain gorillas (Pan gorilla beringei), PhD thesis, University of Chicago, Chicago (1983).
Watts, D. P. Composition and variability of mountain gorilla diets in the central Virungas. Am. J. Primatol. 7, 323–356 (1984).
McNeilage, A. Diet and habitat use of two mountain gorilla groups in contrasting habitats in the Virungas in Mountain gorillas: three decades of research at Karisoke (eds Robbins, M. M., Sicotte, P. & Stewart, K. J.) 265–292 (Cambridge University Press, Cambridge, 2001).
Grueter, C. C., Deschner, T., Behringer, V., Fawcett, K. & Robbins, M. M. Socioecological correlates of energy balance using urinary C-peptide measurements in wild female mountain gorillas. Physiol. Behav. 127, 13–19 (2014).
Chancellor, R. L. & Isbell, L. A. Food site residence time and female competitive relationships in wild gray-cheeked mangabeys (Lophocebus albigena). Behav. Ecol. Sociobiol. 63, 1447–1458 (2009).
Nakagawa, N. Feeding rate as valuable information in primate feeding ecology. Primates 50, 131–141 (2009).
Rothman, J. M., Plumptre, A. J., Dierenfeld, E. S. & Pell, A. N. Nutritional composition of the diet of the gorilla (Gorilla beringei): a comparison between two montane habitats. J. Trop. Ecol. 23, 673–682 (2007).
Ganas, J. & Robbins, M. M. Ranging behavior of the mountain gorillas (Gorilla beringei beringei) in Bwindi Impenetrable National Park, Uganda: a test of the ecological constraints model. Behav. Ecol. Sociobiol. 58, 277–288 (2005).
Furtbauer, I., Mundry, R., Heistermann, M., Schulke, O. & Ostner, J. You mate, I mate: Macaque females synchronize sex not cycles. Plos One 6 (2011).
McCullagh, P. & Nelder, J. Generalized linear models (Chapman & Hall, London, 2008).
Bates, D., Machler, M., Bolker, B. M. & Walker, S. C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 (2015).
Schielzeth, H. & Forstmeier, W. Conclusions beyond support: overconfident estimates in mixed models. Behav. Ecol. 20, 416–420 (2009).
Barr, D. J., Levy, R., Scheepers, C. & Tily, H. J. Random effects structure for confirmatory hypothesis testing: Keep it maximal. J. Mem. Lang. 68, 255–278 (2013).
Gray, M. et al. Census of the Virunga mountain gorillas: complete sweep method versus monitoring. J. Afr. Ecol. 48, 588–599 (2009).
Kalpers, J. et al. Gorillas in the crossfire: population dynamics of the Virunga mountain gorillas over the past three decades. Oryx 37, 326–337 (2003).
Weber, B. & Vedder, A. Population dynamics of the Virunga gorillas: 1959–1978. Biol. Cons. 26, 341–366 (1983).
Vedder, A. Virunga mountain gorilla census, coordinators report. Wildlife Conservation Society, New York (1986).
Aveling, C. & Harcourt, A. H. A census of the Virunga gorillas. Oryx 18, 8–13 (1984).
Harcourt, A. H. & Groom, A. F. G. Gorilla census. Oryx 11, 355–363 (1972).
Groom, A. F. G. Squeezing out the mountain gorillas. Oryx 2, 207–215 (1973).
Sholley, C. Census of mountain gorillas in the Virungas of central Africa, Mountain Gorilla Project (1989).
Acknowledgements
We thank Tony Mudakikwa, Rosette Chantal Rugamba, and Prosper Uwingeli at Rwanda Development Board for permission to conduct research at Volcanoes National Park, Christophe Boesch for endorsing this collaborative project, the Karisoke Research Center data technicians and trackers for facilitating our field work and collecting behavioural data, Avit Nkurikiyumukiza for leading the ‘nest-to-nest’ team, Katie Fawcett, Gaetan Nsengiyumva, John Ndayambaye, Fundi Jean Damascene Hategekimana, and Joel Glick for administrative and logistical support, Ferdinand Ndamiyabo for assistance in collecting plant samples, Sylvia Ortmann and Heidrun Barleben for plant sample analyses, and Roger Mundry for statistical advice. Funding for this study was provided by the Max Planck Society, the Dian Fossey Gorilla Fund International, and the Swiss National Science Foundation/Schweizerischer Nationalfonds (PBZHP3-128152). The research conducted adhered to the legal requirements of Rwanda and complied with all Rwanda Development Board regulations.
Author information
Authors and Affiliations
Contributions
C.C.G. and D.A. were primarily responsible for collecting data, V.V. and F.N. contributed to data collection, A.M.R. and C.C.G. analyzed data, A.M.R., C.C.G., M.M.R. and T.S.S. wrote the manuscript, and A.M.R. prepared figures. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Grueter, C.C., Robbins, A.M., Abavandimwe, D. et al. Quadratic relationships between group size and foraging efficiency in a herbivorous primate. Sci Rep 8, 16718 (2018). https://doi.org/10.1038/s41598-018-35255-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-35255-0
Keywords
This article is cited by
-
Tight quarters: ranging and feeding competition in a Colobus angolensis ruwenzorii multilevel society occupying a fragmented habitat
Behavioral Ecology and Sociobiology (2022)
-
Dissecting the two mechanisms of scramble competition among the Virunga mountain gorillas
Behavioral Ecology and Sociobiology (2021)
-
Heterogeneity in patterns of helminth infections across populations of mountain gorillas (Gorilla beringei beringei)
Scientific Reports (2021)
-
Using long-term ranging patterns to assess within-group and between-group competition in wild mountain gorillas
BMC Ecology (2020)
-
Female dispersal patterns influenced by male tenure duration and group size in western lowland gorillas
Behavioral Ecology and Sociobiology (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
