
Abstract
Deforestation associated with the initial settlement of New Zealand is a dramatic example of how humans can alter landscapes through fire. However, evidence linking early human presence and land-cover change is inferential in most continental sites. We employed a multi-proxy approach to reconstruct anthropogenic land use in New Zealand’s South Island over the last millennium using fecal and plant sterols as indicators of human activity and monosaccharide anhydrides, polycyclic aromatic hydrocarbons, charcoal and pollen as tracers of fire and vegetation change in lake-sediment cores. Our data provide a direct record of local human presence in Lake Kirkpatrick and Lake Diamond watersheds at the time of deforestation and a new and stronger case of human agency linked with forest clearance. The first detection of human presence matches charcoal and biomarker evidence for initial burning at c. AD 1350. Sterols decreased shortly after to values suggesting the sporadic presence of people and then rose to unprecedented levels after the European settlement. Our results confirm that initial human arrival in New Zealand was associated with brief and intense burning activities. Testing our approach in a context of well-established fire history provides a new tool for understanding cause-effect relationships in more complex continental reconstructions.
Similar content being viewed by others
Introduction
Humans have altered landscapes around the world for thousands of years through their use of fire1. However, the intensity and extent of anthropogenic burning in many regions remains unresolved2. Most reconstructions of past fire regimes focus on variations in the concentration of charcoal particles preserved in the sediments of lakes and other natural wetlands. Such records disclose the occurrence of fire and variations in the frequency and timing of biomass burning, but they are unable to discern whether ignitions originate from natural or human causes. Finding methods for disentangling naturally caused fires from those deliberately started by people remains one of the great challenges in paleofire science1,2. Times and places where past fire activity and vegetation depart from trends that can be explained by climate alone are often used as indirect evidence for anthropogenic burning. Identifying past human presence in particular watersheds has generally relied on archeological data, but few locations offer a clear archeological signal of local fire use. As a result, linking past changes in vegetation and fire with human activity is largely inferential3,4.
One of the most dramatic examples of prehistoric anthropogenic burning occurs in New Zealand, where charcoal and pollen records provide incontrovertible evidence of unprecedented fire activity and native forest decline coinciding with the arrival of people at c. AD 12805,6,7,8. Results from modeling studies that examine vegetation-fire feedbacks attribute the rapid environmental change to small-scale positive feedbacks between fire and the post-fire establishment of high flammability shrublands that enable subsequent burning9,10. Humans, as a new source of ignition, thus triggered a conversion of forest to a more fire-prone shrubland/grassland. This reconstruction, however, is based on charcoal and pollen data alone, and in the absence of archeological evidence of burning it is not possible to determine if humans were present in a particular watershed at the time of a fire event, or if the timing of their activities coincided with the loss of forest11,12. Here we provide direct evidence of brief but intense intervals of local human activity in two New Zealand watersheds at the same time as other proxies show fire and deforestation.
We utilize specific molecular markers to provide a continuous record of human presence, fire activity and land use in the South Island of New Zealand during the last millennium. We determined polycyclic aromatic hydrocarbons (PAHs), monosaccharide anhydrides (MAs), fecal and plant sterols fluxes in sediment cores from Lake Kirkpatrick (45.03° S, 168.57° E) and Lake Diamond (44.65° S, 168.96° E) (Fig. 1) to compare with pollen and charcoal records and paleoclimate information. The three MA compounds (levoglucosan, mannosan and galactosan) are formed from cellulose and hemicellulose combustion13 and the seventeen PAHs (Table S1) are tracers for incomplete combustion of organic matter14. MAs and PAHs can travel medium to long distances in association with the fine fraction of aerosol particles15,16 and ultimately reach lake sediments through wet and dry deposition17. Their signal in lakes complements the macroscopic charcoal (pieces > 125 μm) signal of local fires by providing fire information at regional scales. In Lake Kirkpatrick and Lake Diamond, we also employ two 5β-stanols originating from human feces (coprostanol and epi-coprostanol)18 to trace human presence in the catchments. Two Δ5-sterols (cholesterol and sitosterol) and two 5α-stanols (cholestanol and sitostanol) are used as additional land-use proxies, as these markers account for terrigenous input to the lake and the chemical conditions of sediments19,20 (Table S2).

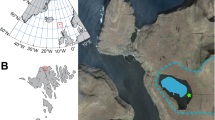
Map of the sampling locations. (a) Diamond Lake in the Lake Wanaka area and sampling location. (b) Lake Kirkpatrick in the Lake Wakatipu area and sampling location. Contains data sourced from the LINZ Data Service licensed for reuse under CC BY 3.0 (https://creativecommons.org/licenses/by/3.0/nz/).
Fire Record
Prior to initial human settlement c. AD 1280, fluxes of charcoal and biomarkers in Lake Kirkpatrick and Lake Diamond were nearly undetectable (Fig. 2). The lack of large charcoal particles (>125 µm in diameter) testifies to the near-absence of fire in the surrounding Lophozonia-podocarp forests which is typical of records from the region5. The presence of extremely low values of molecular fire tracers (PAH total flux < 1 ng cm−2 yr−1, MA total flux < 4 ng cm−2 yr−1) prior to human arrival is attributed to small fires in drier settings in New Zealand or to background atmospheric deposition from distant fires in southeastern Australia and post-depositional processes21,22,23.

Multi-proxy comparison. (a–c) Fecal sterols, total PAH and total MA fluxes (ng cm−2 yr−1) in Lake Kirkpatrick (this study). (d,e) Charcoal flux (pieces cm−2 yr−1) and pollen percentages in Lake Kirkpatrick (McWethy et al.6).
The fluxes of all MAs and PAHs abruptly and simultaneously increase in sediments at c. AD 1345–1365 and mark a period of intense or multiple fire events. The peaks occur shortly after Māori arrival and during the Initial Burning Period, which has been identified from charcoal records from Lake Kirkpatrick and other South Island lakes5,24,25. Levoglucosan, mannosan and galactosan records clearly indicate that combustion of plant biomass reached a maximum at c. AD 1350 with fluxes of 390, 278 and 66 ng cm−2 yr−1, respectively. Relative proportions of the three isomers are consistent with emission factors typical of conifer burning26.
The PAH pattern (Fig. S1) is consistent with typical PAH profiles obtained from the combustion of biomass including several types of hardwood27 and softwood28,29,30. Low molecular weight compounds (Table S1), such as naphthalene, acenaphthylene and acenaphthene (128–154 g mol−1), are poorly represented, which is not surprising as they are commonly present in the gaseous phase and relatively more water-soluble and prone to biodegradation than heavier PAHs, which absorb to atmospheric particles31. The PAH distribution is dominated by 3- and 4-ring molecules (166–228 g mol−1), in particular phenanthrene, fluorene, fluoranthene and pyrene (Fig. S1). These PAH compounds are produced during biomass burning and are involved in the formation of atmospheric particles, eventually incorporated into lake sediments as a result of aerial deposition and surface runoff17. At Lake Kirkpatrick, phenanthrene accounted for 43% of total PAHs on average, with concentrations ranging from a few nanograms per gram (dry weight) to a maximum of 212 ng g−1, corresponding to a flux of 180 ng cm−2 yr−1, during the IBP. Heavier compounds (252–278 g mol−1) were present only in small to negligible concentrations, and this was not surprising as they are mainly produced by higher temperature processes (e.g. fossil fuel combustion) and associated with coarse particles, less likely to travel far from the source area17,32,33.
The abundance and distribution of medium-weight PAHs during the Initial Burning Period is consistent with sustained fires characterized by low oxygen availability and a high flaming to smoldering combustion ratio34,35. Based on the scarcity of high molecular weight PAHs, on the thermal stability of detected compounds36 and on burning experiments37,38, we infer a maximum combustion temperature averaging 400–500 °C. This temperature range was found to maximize the production of 3–4 ring PAHs from biomass combustion32,39. Concentrations and trends detected for medium-weight PAHs thus suggest an infrequent low-intensity natural fire regime before the arrival of humans and high-intensity or high-frequency fires during the Initial Burning Period.
The peak in retene, a tracer of combusted coniferous wood and the associated degradation of abietic acid30, in the L. Kirkpatrick core (101–112 cm depth) implies an abundance of softwood fuel (Fig. S2). However, the levoglucosan to mannosan ratio suggests an increased hardwood to softwood fuel ratio40 at ~AD 1348–1394 (101–112 cm), ~AD 1790–1805 (33–36 cm) and ~AD 1924–1949 (9–13 cm) (Fig. S2). The observed retene record is consistent with the combustion and/or the post-depositional reduction of diterpenoids from softwood species41.
After c. AD 1350, the pollen data5,6 suggest a shift in the composition of vegetation from native Lophozonia-podocarp forest towards more fire-adapted shrubs (e.g., Leptospermum), grasses and bracken fern (Pteridium esculentum). The biomarker reconstruction agrees with the new fuel types that replaced the podocarp-hardwood forests and the change in fire regime characterized by infrequent or smaller low-intensity fires based on low levels of macroscopic charcoal (Fig. 2)6. Such fires would result in lower temperatures and less oxygen depleted conditions, limiting the production of MAs and PAHs, which is strongly dependent on fuel and burning conditions30,35, as observed in the post-settlement record.
Anthropogenic Land Use Record
Unlike the fire tracers, sterols primarily reach lake sediments through runoff42, and thus describe inputs from the local watershed. The evidence of human presence within the Lake Kirkpatrick and Lake Diamond catchments is provided by coprostanol - the most abundant sterol in human feces18- and epi-coprostanol resulting from the epimerization of coprostanol19. In addition, cholesterol and cholestanol are produced by vertebrates, especially mammals including humans43, whereas sitosterol (the most abundant sterol in land plants44,45) and sitostanol come from the input of terrigenous organic matter to the lake. The degree of sterol to stanol conversion infers redox conditions of the basin46 (see supplementary information).
Prior to human settlement, fluxes of fecal sterols in both lakes were near zero, although we measured small background fluxes (at the sub-ng cm−2 yr−1 level) of coprostanol and epi-coprostanol in all pre-Maori samples. Small amounts of coprostanol were previously detected also in anaerobic soils devoid of a fecal input and related to the presence of microbial species capable of converting cholesterol to 5β-stanols47,48, which would provide an explanation for the presence of a natural background in anoxic sediments.
The substantial increase at c. AD 1345–1365 (106–116 cm depth) in Lake Kirkpatrick and at c. AD 1310–1380 (42–55 cm depth) in Lake Diamond (Fig. 3) likely originated from human waste.

Fire and human presence at Lake Kirkpatrick and Lake Diamond. (a–c) Sterols in Lake Kirkpatrick (this study). (d) charcoal flux in Lake Kirkpatrick (McWethy et al.6). (e–g) Sterols in Lake Diamond (this study). (h) charcoal flux in L. Diamond (McWethy et al.25). All sterols fluxes are shown as ng cm−2 yr−1 and charcoal fluxes as pieces cm−2 yr−1.
A short-lived peak in fecal sterols occurred with the onset of the Initial Burning Period in the Lake Kirkpatrick record, and slightly preceded it at Lake Diamond25. The brevity of the peaks at both sites suggests that human activity in the watershed lasted only a few decades. After a large increase in fire activity during the 1300 s, the charcoal influx and fecal sterols values decreased dramatically but remained at about twice the flux of the pre- Māori period. This decline between c. AD 1400 and 1800 likely marks a reduced presence of people in the watershed prior to European colonization, although humans may have been present sporadically or in low numbers.
An increase in sedimentation rate at Lake Kirkpatrick at ~AD 1340–1350 (108–118 cm depth) and Lake Diamond at ~AD 1300–1450 (35–56 cm depth) is consistent with increased erosion following forest clearance. In addition, the high levels of sitostanol imply greater terrigenous input to the lake during and after the Initial Burning Period, as a consequence of increased erosion, reworking of burnt trees on the landscape as they slowly decompose, and runoff. At Lake Kirkpatrick, sitosterol and cholesterol fluxes were replaced by a rise in the reduced compounds sitostanol and cholestanol (Fig. 4) immediately after the Initial Burning Period, suggesting a high level of organic input resulting in reducing chemical conditions.

Reduction of Δ5-sterols in sediments of Lake Kirkpatrick. (a) Sitosterol. (b) Sitostanol. (c) Cholesterol. (d) Cholestanol. All compounds are plotted as fluxes (ng cm−2 yr−1). The favored reaction pathway converts Δ5-sterols (Sit, Chl) to 5α corresponding stanols (5α-Sit, 5α-Chl).
These erosion-related biomarkers were not present in Lake Diamond sediments, where all sterols and stanols increased between AD 1310 and 1380 (Fig. 2). The differences in water depth and catchment steepness between sites may result in lower sedimentation rate and thus limited reducing conditions at Lake Diamond. All sterol markers in Lake Kirkpatrick increased dramatically during the decades of European settlement (c. AD 1800-present), despite few or no changes in local fire activity and sedimentation rate. This increase is therefore ascribed mostly to the intensification of land use for sheep grazing43 and associated growing human presence in the Lake Kirkpatrick watershed. The lack of a 19th century increase in sterols at Lake Diamond is consistent with the absence of sheep grazing and with the scarce anthropic pressure in this particular watershed.
Human Colonization and Land-Use Activity in the South Island of New Zealand
Archeological and paleoenvironmental records indicate that Māori rapidly traveled throughout the North and South Islands of New Zealand soon after their arrival ~AD 12806,7,8,9,22. North Island Māori populations cultivated a number of food crops, including kumara (Ipomoea batatas) and taro (Colocasia esculenta), which was likely associated with initial clearing with fire49. The inability to grow crops in the interior and southern South Island necessitated foraging for a wide array of forest, wetland and marine resources and the use of fire to promote key carbohydrate resources, such as fire-adapted bracken fern50.
Comparing the different proxies of humans, fire and vegetation reveals their relative source areas. The macroscopic charcoal and sterols data at Lake Kirkpatrick and Lake Diamond have a watershed source and jointly show the initial period of forest clearance (the Initial Burning Period) (Fig. 2) and the subsequent period of low-intensity or infrequent burning, and partial forest recovery c. AD 1500 to 1700 (Fig. 2a–e). The fire biomarkers – MAs and PAHs – show high levels of intense burning soon after human arrival, although their source area likely encompasses the whole South Island or beyond. The low charcoal influx and fire biomarker levels after the Initial Burning Period at L. Kirkpatrick suggest smaller and lower temperature fires until the arrival of Europeans. Not all peaks in fire intensity, recorded by PAH and levoglucosan (Fig. 2b,c), coincide with charcoal peaks (Fig. 1d) suggesting that the biomarkers may have a wider source area than the charcoal. Declines in fecal sterols flux (Fig. 2a) from c. AD 1400 to c. 1800 suggest reduced anthropogenic pressure in the watershed, even when forest clearance through fire was still occurring elsewhere on the island.
European settlement at the beginning of the 19th century led to further forest conversion to scrubland and pasture in the South Island. However, fire biomarker concentrations do not significantly increase at the same time as pollen evidence of land use (Fig. 2b–d). The low levels of fire biomarkers, as compared to changes in pollen assemblages, imply that land-use practices did not require burning (i.e., grazing), the type of fuel shifted from forest to seral vegetation (including Leptospermum, bracken and grasses), or that the two proxies have different source areas.
The initial burning of forests by early Māori settlers therefore led to a dramatic transformation of the vegetation that continued again in the 19th and 20th centuries by Europeans. Recent size estimates of the initial founding group suggest it was composed of approximately 500 individuals51. It is remarkable that these small populations were able to convert c. 40% of New Zealand’s forests to grass and shrubland within 1–2 centuries of their arrival7. Our records of sterols and multiple fire markers, so closely matching paleoecological evidence, show that initial Māori presence in particular watersheds was brief and transient, and open vegetation was maintained by subsequent low-intensity fires and sporadic human use of the catchments until the arrival of Europeans. The successful demonstration of biomarker detection in New Zealand sediments, where the anthropogenic nature of fires is undisputed, illustrates the power of this approach to resolve the role of humans in biomass burning, in regions where fire drivers and the timing of human arrival are still debated.
Methods
Two sediment cores were retrieved at Lake Kirkpatrick (195 cm) and Lake Diamond (160 cm) in 2009 using 7 cm polycarbonate tubes (Klein corer)5,6. The cores were split for archival purposes and the working half was sectioned into 1 cm thick slices. Three aliquots from each sample were processed separately for the analyses of charcoal, pollen (ref.5,6) and organic tracers (this study), respectively. Chronology was obtained by Accelerated Mass Spectrometry (AMS) 14C dates based on twig charcoal and plant macrofossils, and calibrated with BChron5,6,52.
72 samples were obtained in the 6–135 cm section of the Lake Kirkpatrick (0–191 cm) core and 49 samples were obtained for Lake Diamond (0–160 cm) in the 5–147 cm section. Wet samples were dried in the desiccator with silica gel until they maintained a constant weight and were then hand milled and homogenized in a ceramic mortar. Samples were stored at room temperature in sealed vials until extraction, which was performed with an ASE 200 (Accelerated Solvent Extraction, Dionex Thermo Fisher Scientific). Each sample was dispersed with diatomaceous earth and spiked with a known amount of a 13C-labeled internal standard solution for the quantification of the analytes (13C6-Cholesterol, 13C6-Acenaphtylene, 13C6-Phenanthrene, 13C4-Benzo(a)pyrene, 13C6-Levoglucosan) and extracted twice at 150 °C and 1500 psi with dichloromethane (L. Diamond) or with a dichloromethane:methanol = 9:1 (v/v) mixture (L. Kirkpatrick).
Extracts were concentrated in a centrifugal evaporator (Genevac EZ-2 Solvent Evaporator) up to ~0.5 mL and purified onto disposable solid phase extraction silica tubes (Supelco DSC-Si 12 mL, 2 g bed weight), previously conditioned with 40 mL of dichloromethane (DCM). The clean-up and fractionation of samples were achieved by eluting the samples with 70 mL of DCM followed by 20 mL of methanol (MeOH), adapting previously published procedures24,46,53. PAH and sterols were collected in the DCM fraction (F1), while MA were contained in the polar MeOH fraction (F2) and treated separately. F1 was concentrated to 100–200 μL and PAHs were analyzed through gas chromatography-mass spectrometry (GC-MS). After the analysis, 100 μL of BSTFA + 1% TMCS (N,O- Bis(trimethylsilyl)trifluoroacetamide + 1% Trimethylchlorosilane) were added to the samples to allow derivatization at 70 °C for 1 h. After a stabilization period of 24 h at room temperature, sterols were analyzed by GC-MS. F2 was evaporated to dryness, redissolved in 0.5 mL of ultrapure water and centrifuged before analysis by ion chromatography-mass spectrometry (IC-MS)24. The GC-MS and IC-MS methods, along with instrumental setup and target and qualifier mass-to-charge ratios employed are reported elsewhere24,53,54. Analytes were quantified based on internal standards through the isotope dilution technique. Results were calculated and corrected by the instrumental response factors, obtained by the repeated analysis of standard solutions containing all analytes and 13C internal standards at 100 pg µL−1 for PAH and 1 ng µL−1 for MA and sterols. Fluxes were calculated using the accumulation rate and the dry density following Menounos (1997)55.
Several procedural blanks were also analyzed in order to quantify possible contamination from the laboratory equipment. Absolute quantities were corrected for the blank plus three times the standard deviation and divided by the dry weight of samples to obtain concentrations. The average accuracy was 89 ± 16% for PAH, 115 ± 17% for sterols and 99 ± 11% for MA. Precision ranged between 2 and 23% for PAH, 12–24% for sterols 19–33% for MA.
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Bowman, D. M. J. S. et al. The human dimension of fire regimes on Earth. J. Biogeogr. 38, 2223–2236 (2011).
Williams, A. N., Mooney, S. D., Sisson, S. A. & Marlon, J. Exploring the relationship between Aboriginal population indices and fire in Australia over the last 20,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 432, 49–57 (2015).
Marlon, J. R. et al. Climate and human influences on global biomass burning over the past two millennia. Nat. Geosci 1, 697–702 (2008).
Caldararo, N. Human ecological intervention and the role of forest fires in human ecology. Sci. Total Environ. 292, 141–65 (2002).
McWethy, D. B. et al. Rapid landscape transformation in South Island, New Zealand, following initial Polynesian settlement. Proc. Natl. Acad. Sci. 107, 21343–21348 (2010).
McWethy, D. B., Wilmshurst, J. M., Whitlock, C., Wood, J. R. & McGlone, M. S. A High-Resolution Chronology of Rapid Forest Transitions following Polynesian Arrival in New Zealand. PLoS One 9, e111328 (2014).
Perry, G. L. W., Wilmshurst, J. M., McGlone, M. S., McWethy, D. B. & Whitlock, C. Explaining fire-driven landscape transformation during the Initial Burning Period of New Zealand’s prehistory. Glob. Chang. Biol. 18, 1609–1621 (2012).
Wilmshurst, J. M., Anderson, A. J., Higham, T. F. G. & Worthy, T. H. Dating the late prehistoric dispersal of Polynesians to New Zealand using the commensal Pacific rat. Proc. Natl. Acad. Sci. USA 105, 7676–80 (2008).
Tepley, A. J., Veblen, T. T., Perry, G. L. W., Stewart, G. H. & Naficy, C. E. Positive Feedbacks to Fire-Driven Deforestation Following Human Colonization of the South Island of New Zealand. Ecosystems 19, 1325–1344 (2016).
Kitzberger, T. et al. Fire–vegetation feedbacks and alternative states: common mechanisms of temperate forest vulnerability to fire in southern South America and New Zealand. New Zeal. J. Bot. 54, 247–272 (2016).
Butler, K. Interpreting charcoal in New Zealand’s palaeoenvironment—What do those charcoal fragments really tell us? Quat. Int. 184, 122–128 (2008).
McGlone, M. S. & Wilmshurst, J. M. Dating initial Maori environmental impact in New Zealand. Quat. Int. 59, 5–16 (1999).
Simoneit, B. R. T. Biomass burning - a review of organic tracers for smoke from incomplete combustion. Appl. Geochemistry 17, 129–162 (2002).
Abdel-Shafy, H. I. & Mansour, M. S. M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 25, 107–123 (2016).
Yang, H.-H., Tsai, C.-H., Chao, M.-R., Su, Y.-L. & Chien, S.-M. Source identification and size distribution of atmospheric polycyclic aromatic hydrocarbons during rice straw burning period. Atmos. Environ. 40, 1266–1274 (2006).
Zangrando, R. et al. Levoglucosan and phenols in Antarctic marine, coastal and plateau aerosols. Sci. Total Environ. 544, 606–616 (2016).
Stogiannidis, E. & Laane, R. In Reviews of Environmental Contamination and Toxicology 234, 38 (2015).
Bull, I. D., Lockheart, M. J., Elhmmali, M. M., Roberts, D. J. & Evershed, R. P. The origin of faeces by means of biomarker detection. Environ. Int. 27, 647–654 (2002).
Birk, J. J., Teixeira, W. G., Neves, E. G. & Glaser, B. Faeces deposition on Amazonian Anthrosols as assessed from 5β-stanols. J. Archaeol. Sci. 38, 1209–1220 (2011).
Derrien, M., Yang, L. & Hur, J. Lipid biomarkers and spectroscopic indices for identifying organic matter sources in aquatic environments: A review. Water Res. 112, 58–71 (2017).
Marx, S. K., Kamber, B. S. & McGowan, H. A. Estimates of Australian dust flux into New Zealand: Quantifying the eastern Australian dust plume pathway using trace element calibrated 210Pb as a monitor. Earth Planet. Sci. Lett. 239, 336–351 (2005).
McGowan, H. A., Kamber, B., McTainsh, G. H. & Marx, S. K. High resolution provenancing of long travelled dust deposited on the Southern Alps, New Zealand. Geomorphology 69, 208–221 (2005).
Marx, S. K., Kamber, B. S. & McGowan, H. A. Provenance of long-travelled dust determined with ultra-trace-element composition: A pilot study with samples from New Zealand glaciers. Earth Surf. Process. Landforms 30, 699–716 (2005).
Kirchgeorg, T., Schüpbach, S., Kehrwald, N., McWethy, D. B. & Barbante, C. Method for the determination of specific molecular markers of biomass burning in lake sediments. Org. Geochem. 71, 1–6 (2014).
McWethy, D. B., Whitlock, C., Wilmshurst, J. M., McGlone, M. S. & Li, X. Rapid deforestation of South Island, New Zealand, by early Polynesian fires. The Holocene 19, 883–897 (2009).
Alves, C. A. et al. Particulate organic compounds emitted from experimental wildland fires in a Mediterranean ecosystem. Atmos. Environ. 44, 2750–2759 (2010).
Oros, D. R. & Simoneit, B. R. T. Identification and emission factors of molecular tracers in organic aerosols from biomass burning Part 2. Deciduous trees. Appl. Geochemistry 16, 1545–1565 (2001).
Jimenez, J., Farias, O., Quiroz, R. & Yañez, J. Emission factors of particulate matter, polycyclic aromatic hydrocarbons, and levoglucosan from wood combustion in south-central Chile. J. Air Waste Manage. Assoc. 67, 806–813 (2017).
Bari, M. A., Baumbach, G., Kuch, B. & Scheffknecht, G. Wood smoke as a source of particle-phase organic compounds in residential areas. Atmos. Environ. 43, 4722–4732 (2009).
Oros, D. R. & Simoneit, B. R. Identification and emission factors of molecular tracers in organic aerosols from biomass burning Part 1. Temperate climate conifers. Appl. Geochemistry 16, 1513–1544 (2001).
Ravindra, K., Sokhi, R., Vangrieken, R. & Van Grieken, R. Atmospheric polycyclic aromatic hydrocarbons: source attribution, emission factors and regulation. Atmos. Environ. 42, 2895–2921 (2008).
Wang, C., Wang, Y. & Herath, H. M. S. K. Polycyclic aromatic hydrocarbons (PAHs) in biochar – Their formation, occurrence and analysis: A review. Org. Geochem. 114, 1–11 (2017).
Tobiszewski, M. & Namieśnik, J. PAH diagnostic ratios for the identification of pollution emission sources. Environ. Pollut. 162, 110–119 (2012).
Andreae, M. O. & Merlet, P. Emissions of trace gases and aerosols from biomass burning. Global Biogeochem. Cycles 15, 955–966 (2001).
Reid, J. S., Koppmann, R., Eck, T. F. & Eleuterio, D. P. A review of biomass burning emissions part II: intensive physical properties of biomass burning particles. Atmos. Chem. Phys 5, 799–825 (2005).
Marynowski, L., Kubik, R., Uhl, D. & Simoneit, B. R. T. Molecular composition of fossil charcoal and relationship with incomplete combustion of wood. Org. Geochem. 77, 22–31 (2014).
McGrath, T. E., Chan, W. G. & Hajaligol, M. R. Low temperature mechanism for the formation of polycyclic aromatic hydrocarbons from the pyrolysis of cellulose. J. Anal. Appl. Pyrolysis 66, 51–70 (2003).
Kislov, V. V., Sadovnikov, A. I. & Mebel, A. M. Formation Mechanism of Polycyclic Aromatic Hydrocarbons beyond the Second Aromatic Ring. J. Phys. Chem. A 117, 4794–4816 (2013).
Schellekens, J. et al. Molecular characterization of biochar from five Brazilian agricultural residues obtained at different charring temperatures. J. Anal. Appl. Pyrolysis 130, 249–255 (2018).
Schmidl, C. et al. Chemical characterisation of fine particle emissions from wood stove combustion of common woods growing in mid-European Alpine regions. Atmos. Environ. 42, 126–141 (2008).
Achten, C., Beer, F. T., Stader, C. & Brinkhaus, S. G. Wood-Specific Polycyclic Aromatic Hydrocarbon (PAH) Patterns in Soot Using Gas Chromatography-Atmospheric Pressure Laser Ionization-Mass Spectrometry (GC-APLI-MS). Environ. Forensics 16, 42–50 (2015).
Meyers, P. A. Organic geochemical proxies of paleoceanographic, paleolimnologic, and paleoclimatic processes. Org. Geochem. 27, 213–250 (1997).
Leeming, R., Ball, A., Ashbolt, N. & Nichols, P. Using faecal sterols from humans and animals to distinguish faecal pollution in receiving waters. Water Res. 30, 2893–2900 (1996).
Volkman, J. K. A review of sterol markers for marine and terrigenous organic matter. Org. Geochem. 9, 83–99 (1986).
Meyers, Pa & Ishiwatari, R. Lacustrine organic geochemistry—an overview of indicators of organic matter sources and diagenesis in lake sediments. Org. Geochem. 20, 867–900 (1993).
Battistel, D. et al. Fire and human record at Lake Victoria, East Africa, during the EarlyIron Age: Did humans or climate cause massive ecosystem changes? The Holocene, https://doi.org/10.1177/0959683616678466 (2016).
Bethell, P. H., Goad, L. J., Evershed, R. P. & Ottaway, J. The Study of Molecular Markers of Human Activity: The Use of Coprostanol in the Soil as an Indicator of Human Faecal Material. J. Archaeol. Sci. 21, 619–632 (1994).
Prost, K., Birk, J. J., Lehndorff, E., Gerlach, R. & Amelung, W. Steroid biomarkers revisited - Improved source identification of faecal remains in archaeological soil material. PLoS One 12, 1–30 (2017).
Furey, L. Maori Gardening - An archaeological perspective. (Department of Conservation, 2006).
McGlone, M. S., Wilmshurst, J. M. & Leach, H. M. An ecological and historical review of bracken (Pteridium esculentum) in New Zealand, and its cultural significance. N. Z. J. Ecol. 29, 165–184 (2005).
Walter, R., Buckley, H., Jacomb, C. & Matisoo-Smith, E. Mass Migration and the Polynesian Settlement of New Zealand. J. World Prehistory 30, 351–376 (2017).
Haslett, J. & Parnell, A. A simple monotone process with application to radiocarbon-dated depth chronologies. J. R. Stat. Soc. Ser. C (Applied Stat. 57, 399–418 (2008).
Battistel, D. et al. GC-MS method for determining faecal sterols as biomarkers of human and pastoral animal presence in freshwater sediments. Anal. Bioanal. Chem. 407, 8505–8514 (2015).
Piazza, R. et al. Development of a method for simultaneous analysis of PCDDs, PCDFs, PCBs, PBDEs, PCNs and PAHs in Antarctic air. Anal. Bioanal. Chem. 405, 917–932 (2013).
Menounos, B. The water content of lake sediments and its relationship to other physical parameters: an alpine case study. The Holocene 7, 207–212 (1997).
Acknowledgements
The research leading to these results has received funding from the European Research Council under the European Union’s Seventh Framework Programme (FP7/2007–2013)/ERC Grant agreement n° 267696 – EARLYhumanIMPACT. The research was also supported by grants from the US National Science Foundation (0966472 and 1024413). Elga Lab water, High Wycombe UK, supplied the pure water system used in this study. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Contributions
E.A., T.K. and M.V. analyzed samples; E.A. analyzed data, designed figures and wrote the paper; E.A., D.B. and N.M.K. developed analytical tools; D.B.M., C.W. and J.M.W. conducted field work, provided samples and pollen and charcoal data. C.B. and C.W. supervised the projects. All authors collaborated on manuscript editing at all stages.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Argiriadis, E., Battistel, D., McWethy, D.B. et al. Lake sediment fecal and biomass burning biomarkers provide direct evidence for prehistoric human-lit fires in New Zealand. Sci Rep 8, 12113 (2018). https://doi.org/10.1038/s41598-018-30606-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-30606-3
This article is cited by
-
Fecal steroids as a potential tool for conservation paleobiology in East Africa
Biodiversity and Conservation (2022)
-
Paleolimnology in support of archeology: a review of past investigations and a proposed framework for future study design
Journal of Paleolimnology (2021)
-
Geochemical Evidence for the Control of Fire by Middle Palaeolithic Hominins
Scientific Reports (2019)
-
Anhydrosugars as tracers in the Earth system
Biogeochemistry (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
