
Abstract
In major modern reef regions, either in the Indo-Pacific or the Caribbean, scleractinian corals are described as the main reef framework builders, often associated with crustose coralline algae. We used underwater cores to investigate Late Holocene reef growth and characterise the main framework builders in the Abrolhos Shelf, the largest and richest modern tropical reef complex in the South Western Atlantic, a scientifically underexplored reef province. Rather than a typical coralgal reef, our results show a complex framework building system dominated by bryozoans. Bryozoans were major components in all cores and age intervals (2,000 yrs BP), accounting for up to 44% of the reef framework, while crustose coralline algae and coral accounted for less than 28 and 23%, respectively. Reef accretion rates varied from 2.7 to 0.9 mm yr−1, which are similar to typical coralgal reefs. Bryozoan functional groups encompassed 20 taxa and Celleporaria atlantica (Busk, 1884) dominated the framework at all cores. While the prevalent mesotrophic conditions may have driven suspension-feeders’ dominance over photoautotrophs and mixotrophs, we propose that a combination of historical factors with the low storm-disturbance regime of the tropical South Atlantic also contributed to the region’s low diversity, and underlies the unique mushroom shape of the Abrolhos pinnacles.
Similar content being viewed by others

Introduction
Shallow-water tropical and subtropical reefs are rigid carbonate structures built mainly by corals and crustose coralline algae (CCA) during the Cenozoic1. Such living coralgal reefs may reach thousands of square kilometers in shallow tropical and subtropical regions and encompass the world’s most biodiverse marine ecosystems2. Reef distribution, structure and morphology vary as a function of the geological, climatic and biogeographic histories of each ocean basin, coupled with modern climate-oceanographic conditions. For instance, benthic community structure, including the relative abundance of reef-builders and eroders, may vary sharply in small spatial scales of a few meters, being influenced by depth, light, nutrient, sedimentation, current and turbulence3,4. In addition, biodiversity patterns and community structure also respond to latitudinal and longitudinal gradients of thousands of kilometers, within and among ocean basins5.
In the tropical and subtropical portions of the Indo-Pacific and the Caribbean it is well established that reef frameworks are built by species-rich assemblages of scleractinian corals and coralline algae6,7. Reef structures develop as well-known morphological types that range from patch and fringing reefs to barrier reefs and atolls8. The tropical South-western Atlantic (SWA) represents one of the world’s smallest and poorest coral reef provinces5,9, with high endemism levels of up to 50%. In the SWA, the largest and richest coral reefs occur in the Abrolhos Shelf (>8,000 km2, 17–19.5°S)10,11. The unusual mushroom-shaped morphology of the Abrolhos reefs and the low coral richness of the SWA was noted in the 19th Century12,13, but framework building within this entire region remains poorly understood.
The outer Abrolhos Shelf is largely covered by rhodoliths (CCA-structured nodules) and the inner shelf is covered by siliciclastic-carbonate mixed soft sediments11,14. Emergent and quasi-emergent reefs developed during the Holocene and occur in a near-shore and a mid-shelf arc, 12 and 60 km offshore respectively15, while mesophotic reefs with erosive topographies occur in the mid-outer shelf and near the shelf edge11 (Fig. 1). The major reef builder of the Abrolhos reefs was considered to be the Brazilian-endemic coral Mussismilia braziliensis (Verrill, 1868), in association with other corals and CCA10,16.

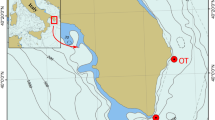
Study area and sampling sites. (A) Map showing the Abrolhos shelf physiography and the distribution of reefs in the near-shore and mid-shelf arcs (light blue) and drilling sites (Red dots). (B) Side Scan Sonar imagery of mid and outer shelf areas, showing complex submerged reef morphologies. (C) Aerial drone image of the “chapeirões” at the Parcel dos Abrolhos (research boat is 20 m long, for scale reference); (D) Satelite image (Ikonos 2, 4 m resolution) of Abrolhos Archipelago and its adjacent “chapeirão” reef system (indicated by yellow pixels); (E) Schematic view of the 11 cores sampled from the tops and walls of the three reef pinnacles at 4, 15 and 25 m water depth (not on scale). Aerial photo by Fernando Moraes/Rede Abrolhos.
Here we examine the Late Holocene (~2000 yrs BP) framework of reef pinnacles with tops at 4, 15, and 25 m water depth in the Abrolhos Shelf, based on samples obtained from rotary drilling within the mid-shelf arc. We analyzed accretion rates and reef-builder assemblages, and compared our data with available information from Abrolhos near-shore reefs16 and elsewhere, within and outside the tropical SWA. We also compiled meteoceanographic historical data in order to support our interpretation of reef morphology, biodiversity and benthic assemblage structure, within the context of the low-disturbance regime of SWA reefs.
Results
Pinnacle distribution and morphology
Pinnacles were observed along the inner shelf, usually no deeper than 35 m (seabed water depth). These reefs occurred either in a shallow (seabed at 20–25 m, pinnacles tops at 2 to 6 m water depth) and in a deeper margin (seabed at 25–35 m, pinnacles tops at 15 to 25 m water depth) of the reef area. In general, pinnacle distribution showed a greater density in the shallower region, transitioning to more sparsely-distributed pinnacles in the deeper margin of the reef (Fig. 1). Pinnacles in the more central and shallower region are emergent or quasi-emergent and include some structures coalesced in the tops (Fig. 1B–D). These isolated structures consisted of a cylindrical basement from which laterally-expanded tops develop, resembling giant mushrooms (Figs 1E and 2) which are over 20 m high and 40 m across at the tops. Coral richness and cover was highest at the tops, with several important builders restricted to this well-illuminated and relatively flat and shallow habitat (0-10 m depth), including Mussismilia braziliensis, M. harttii (Verrill, 1868) and Millepora spp. (Fig. 2 and Supplementary Information Fig. S1). Conversely, pinnacles walls were steep and dim, and a single coral species, Montastraea cavernosa (Linnaeus, 1767) was the most conspicuous coral species. The marginal pinnacles with tops below 10 m depth lacked the mushroom-shaped morphology (Figs 1E and 2B) and had smaller coral cover and diversity.

Underwater photos of Abrolhos Bank deep pinnacle reefs in Parcel dos Abrolhos (PA) and California Reef (CR): (A) Side view of part of a mushroom shaped “chapeirão” in PA (15 m deep); (B) Lateral bottom-up view of a columnar biogenic pinnacle structure in CR (soft bottom at 33 m deep and reef top at 20 m deep); (C) Panoramic view of that pinnacle’s reef top in CR, revealing lateral expansion process by the plate coral Montastraea cavernosa (arrows). Photos: Áthila Bertoncini & Fernando Moraes/Rede Abrolhos.
Reef framework and accretion
Based on 14C AMS dating of coral, CCA and bryozoan components, reef sequences span 387–757 (4 m depth core), 463–1,199 (15 m depth core), and 224–1,897 cal years BP (25 m depth core) (Figs 3–5). Average vertical accretion rates increased as pinnacle depth decreased, ranging from 0.93 mm.yr−1 in the deeper pinnacles (tops at 15 and 25 m depths) to 2.7 mm.yr−1 in the shallowest pinnacle (top at 4 m) (Table S1; Supplementary Information Fig. S2 and Table S1). Average horizontal accretion rates varied between 1.4 (4 m), 1.3–2.3 (15 m) and 0.98 mm.yr−1 (25 m).

Simplified core log showing the main reef framework types, sedimentary facies, and 14C AMS age data collected from Abrolhos reef at 4 m water depth. Vertical (4V-05, 4V-06) and horizontal cores (4H-07, 4H-08) were sampled from the pinnacle’s top and wall, respectively. The stratigraphic distribution of the main reef framework building components was mapped, comprising mainly thickly encrusting cheilostome bryozoans with subordinate amounts of CCAs, corals and sediments (see Methods). The vertical and horizontal accretion rates of the pinnacles were assessed based on calibrated 14C AMS measurements from bryozoans and corals in growth position (Methods and Table 1). (A) Close-up image of a vertical core sample showing the reef framework dominated by multilayers of encrusting bryozoan colonies; (B) Petrographic image showing multilayers of the most abundant bryozoan species forming the reef framework (Celleporaria atlantica), including zooids partially filled by cementing matrix; (C) Stereomicroscope image of a multilayer colony of Reptadeonella bipartita, another important bryozoan species forming the reef framework.

Simplified core log showing the main reef framework types, sedimentary facies, and 14C AMS age data collected from Abrolhos reef at 15 m water depth. Vertical (15V-01, 15V-02) and horizontal cores (15H-03, 15H-04) were sampled from the pinnacle’s top and wall, respectively. (A) Close-up image of a shallow/younger horizontal core sample showing the reef framework composed mainly by tick multilayers of encrusting bryozoan colonies interspersed with CCA; (B) Close-up image of a deeper/older horizontal core sample showing preponderance of bryozoans and CCA over corals in the reef framework building.

Simplified core log showing the main reef framework types, sedimentary facies, and 14C AMS age data collected from Abrolhos reef at 25 m water depth. Vertical (25V-09, 25V-10) and horizontal cores (25H-11) were sampled from the pinnacle’s top and wall, respectively. (A) Close-up image of a vertical core sample showing the recurrent pattern of interspersed bryozoans and CCAs in the reef framework. Note sediment fills overgrown by bryozoan layers and remarkable incorporation of fine grains inside the reef matrix.
Bryozoans were major components of the reef framework in all cores and age intervals (Figs 3–5; Supplementary Information Table S2). Considering only framework builders, bryozoans comprised 36, 60 and 44% of the reef framework (in 4, 15 and 25 m deep pinnacles, respectively). Bryozoans encompassed at least 20 taxa (11–15 species in each pinnacle), and species composition varied with depth (Table 1). Celleporaria atlantica (Busk, 1884) dominated the framework at all cores and depths, followed by two undescribed species, Parasmittina sp. nov. 1 and Stylopoma sp. nov. 2. Five species occurred at all depths (C. atlantica, Hippaliosina imperfecta (Canu & Bassler, 1928), Parasmittina sp. nov. 1, and Stylopoma sp. nov. 1 and 2), while three were restricted to the 25 m depth pinnacle (Steginoporella magnilabris (Busk, 1854), Gemelliporina glabra (Smitt, 1873) and Metrarabdotos sp.). Multilayer encrusting was the most common growth form, with 10 species growing as colonies with more than two consecutive layers, six of which with more than 10 layers. Bryozoans were frequently intergrown with CCA. The few erect bryozoan species (S. magnilabris (Busk, 1854) and Crisia sp.) occurred mainly at the 25 m depth pinnacle (Table 1). The former was consistently found inside the cores, while the latter was represented by scattered loose material.
CCA were the second most abundant framework component (15–28%) and encompassed 12 taxa. The cores from both the shallower and intermediate depth pinnacles’ tops comprised species typical of the shallow-water local assemblage17 (e.g. Porolithon onkodes (Foslie, 1909), whereas the cores from walls and deeper reefs were dominated by sciaphyllic taxa such as Sporolithon (Figs 3–5; Supplementary Information Tables S2 and S3).
Corals and hydrocorals reached 23% of the framework in the shallower pinnacle, decreasing to 9–12% in the deeper ones (Figs 3–5; Supplementary Information Tables S2 and S3). Scleractinians comprised five taxa of encrusting and fast-growing “weedy” corals (e.g. Favia sp.), and massive slower-growing “stress-tolerant” species. These latter include both shallow water (e.g. Mussismilia) and wider depth-ranging species (e.g. Madracis, Siderastrea). The only branching forms were fast-growing shallow-water hydrocorals (Millepora spp.).
Discussion
The reef framework of the Abrolhos mid-shelf pinnacles deviates from the archetypal coralgal reefs of the Indo-Pacific and the Caribbean, being dominated by suspension-feeding bryozoans interlayered with CCA, with minor contributions from corals. Instead of being a mere local outlier, this particular framework-building system may be more widespread than previously acknowledged in marginal coralline provinces such as the SWA. For instance, the single coring study from the Abrolhos nearshore reef arc18 described a reef framework “dominated by corals”, with bryozoans being “very common and sometimes abundant”. The first study of the drowned reefs off the Amazon river19 also described a mixed-framework composed by polychaetes, foraminiferans, bryozoans and mollusks.
Bryozoan contribution on recent reefs generally encompasses low-biomass assemblages of encrusting and cavity-dwelling species that strengthen the structures built by corals and CCA20. Encrusting bryozoans are rare on the surface of the Abrolhos pinnacles tops, where CCA and corals dominate the benthic assemblage, being more frequent on the dim walls21. Experimental work with colonization plates22 revealed that bryozoans were the second most abundant group, following CCA. They were particularly fast-growing on the shaded surfaces of22, especially C. atlantica (Supplementary Information Fig. S1C) and four Stylopoma spp. Our cores show that encrusting bryozoans add volume to the framework, going beyond the mere “filling-cavity” role that is widely reported from most modern tropical reefs.
Bryozoan dominance in Late Holocene-modern tropical reefs have rarely been reported. To our knowledge, small patchy reefs built by bryozoans, especially Celleporaria albirostris (Smitt, 1873), are only recorded in Joulters Reefs, Bahamas23. These reefs thrive in shallow water (4 m deep), with a succession of cheilostome bryozoans (9 species) dominating the base (60–70% of the framework) and corals and CCA in the upper portions.
When bryozoans are considered in the context of longer geological time-scales, the present is not a reliable key to the past20,24. Shallow-water reefs dominated by bryozoans were widespread in the Paleozoic25, with scleractinian reefs becoming increasingly important since the Late Triassic26,27. Concurrently, paleolatitudinal patterns of sediment-producing bryozoans shifted from a pan-tropical distribution in the Paleozoic to a more extratropical range during the Post-Paleozoic28. Giant Eocene shelf edge bryozoan mounds are described along the Great Australian Bight, the largest Cenozoic cold-water carbonate province29,30. Although temperate and deep assemblages are not a perfect analogue to Abrolhos, increased bryozoan diversity and biomass occurred during Pleistocene lowstands in southern Australia30, associated with increased subtropical convergence and upwelling. In Miocene reefs in the Kutai Basin (Indonesia), bryozoans are restricted to coral sheetstone facies and diverse bryozoan assemblage occur encrusting the undersides of thin platy corals embedded in a muddy matrix31,32. These species-rich coral communities formed low-relief buildups lacking rigid frameworks in nearshore turbid environments31,33. By contrast, bryozoans are absent in deposits with high siliciclastic input, and in rigid frameworks of tabulate and branching corals and coralline algae, which formed under lower terrestrial influx31,32.
Modern bryozoans occur at all latitudes, but tend to form significant accumulations only in eutrophic/turbid and deep/cold waters20,34,35. The Abrolhos reefs are within 15–30 m water depths and under 21.7–29.6 °C, i.e. within the ranges where coralgal reef growth predominates circutropically36. Bryozoan dominance in the reef framework seems to be primarily associated to the region high nutrient and turbidity. The eastern coast of tropical South America receives substantial riverine input and terrigenous sedimentation10,37. Dissolved inorganic nitrogen and phosphorous concentrations in Abrolhos are above commonly assumed eutrophication thresholds38, and phytoplankton productivity is significantly high39. Remote sensing-derived chlorophyll a annual values range from 0.38 to 0.79 mg.m−3 40, corresponding to a persistent mesotrophic condition41 that is enhanced by shelf-edge upwelling during the wet season (spring/summer). Turbidity is overall high and increases seasonally due to re-suspension of biogenic sediments42. Therefore, oceanographic conditions deviate sharply from those of the archetypal Indo-Pacific and Caribbean coralgal reefs, determining the unique reef-building assemblages. With the exception of oceanic islands43,44, Brazilian reefs develop under similar high turbidity and nutrient levels15,19, comprising foremost examples of turbid zone reefs with low diversity but significant coral cover45,46. Nevertheless, modern and ancient examples include turbid zone reefs that can present high coral diversity3,46, and even high bryozoan diversity31,32, but the latter largely occur at low biomasses and encrusting underneath coral surfaces31, which deviates from the high bryozoan abundance recorded in the Abrolhos mid-shelf reefs.
Another potential driver of reef growth and diversity along the eastern Brazilian margin is the region’s low disturbance regime, with very rare tropical storms and hurricanes (Fig. 6). Akin to tropical rainforests, disturbance from extreme wave and wind energy events may play an important role in coral reef diversity47,48, creating large habitat patches under different succession stages. Such extreme events change reef morphology and also preclude dominance by slow-growing “stress-tolerant” species. While historical factors (e.g. isolation, glacial compression) may underlie the low species richness and functional diversity of SWA coral assemblages, the marginal conditions (e.g. turbidity) and low disturbance regime may be major drivers of the region’s low coral diversity. In addition, the low disturbance regime does not impose significant constraints (e.g., physical erosion) to horizontal accretion of carbonate framework, preventing the breakage of the huge lateral expansions of the oddly shaped pinnacles. Previous mechanisms explaining the mushroom-shaped morphology of the Abrolhos reefs accounted only for the limitations in upward accretion due to the sea level drop during the last 4,000 years10. However, while sea level plays a major role in reef growth, it does not seem to be the single driver of the peculiar morphology of the Abrolhos reefs. Flat pinnacles tops occur in the subsurface, at depths of up to 10 m, and our results indicate ongoing vertical growth. For instance, a depth vs age analysis from the shallower pinnacle (4 m deep x 910–1310 Cal yrs BP) does not support a sea-level control of its shape10.

(A) World occurrence of storm track records from 1985–2015 superposed on coral richness of all reef provinces, (B) Graph showing the relationship between coral species richness and number of tropical storms/hurricanes.
Despite being relatively small and species-poor, tropical SWA reefs provide relevant insights on global patterns of reef building and on the meso-scale drivers of reef biodiversity. Our results challenge the coralgal-framework paradigm of modern shallow water tropical reef building. The Abrolhos mid-shelf reefs deviate from coral reefs in which mixo- and autotrophic organisms (scleractinian corals and CCA) are the main builders. Instead, at least during the last 2,000 yr., these reefs have been largely built by bryozoans, which are heterotrophic suspension-feeders, with a secondary role of CCA and a species-poor coral assemblage. These findings suggest that the role played by bryozoans in reef building may be underestimated in other shallow water tropical reef systems under mesotrophic conditions.
Methods
Study region
Detailed descriptions of the Abrolhos Shelf reefs (Fig. 1) are provided by11,14, including the side scan images in Fig. 1B. Besides using literature data to describe reef morphology and benthic assemblage structure, we carried out a photographic assessment, both underwater (by SCUBA diving) and aerial (by drone) (Fig. 1D). Reefs in the mid-shelf arc are locally called “chapeirões” (=large hats), accounting for their unique “mushroom-like” morphology that was remarked since the first scientific surveys12,13. The mid-shelf chapeirões consist of 2–20 m height pinnacles, with diameters of 2–50 m and laterally expanded tops near the surface. Near-shore, pinnacles are shorter but similarly shaped, often coalescing to form extensive shallow banks and patch reefs15.
Field sampling and sample processing
Coring was carried out using a diver-operated, submarine hydraulic rotary drill. Cores were taken in three depths (4, 15 and 25 m) and habitats (pinnacles’ tops and walls) along the mid-shelf arc. For each depth and habitat, two along side cores were taken, with the exception of the 25 m deep pinnacle wall, where only one core was recovered. All cores were sliced in two halves and photographed in high resolution. The main reef framework types, sedimentary facies and sample context in the cores were logged using a combination of sample material, petrographic thin sections and digital images following procedures laid out in49. Core recoveries and depths were calculated by moving the recovered samples to the top of each core run, following standard International Ocean Discovery Program (IODP) protocols50. The high resolution core images were displayed in Corelyzer 2.0.250 and the percent surface area of each of the main lithologies and reef framework types calculated.
Drilled subsamples of specific lithologies (corals, bryozoans or coralline algae) were radiocarbon dated at Beta Analytics-Florida (USA) via Accelerator Mass Spectrometry (AMS). Dates are reported as calendar years BP (“present” = 1950 CE) using the 2 sigma confidence interval. Calibration was carried out using Calib 7.1 available at http://calib.qub.ac.uk/calib. The Marine13 calibration curve was applied assuming a global marine reservoir effect of 400 years and a Delta R of 85 ± 25 (average value for the two closest localities) for regional correction. Only samples interpreted as in situ (i.e. original growth position) were chosen for dating purposes. Vertical accretion rates were calculated using mean ages and the stacked core sample depths. The maximum potential depth error of each dated core sample was directly related to the actual core recovery of each core run (recovery error in Supplementary Table 1). Only dated samples that were outside of their respective ages errors were used to calculate accretion rates.
All the processed data, including drilling sites coordinates, C14 results, core composition and taxa information are provided as tables in the Supplementary Information file.
Secondary datasets and statistical analyzes
The storm dataset was compiled from NOAA National Climatic Data Center, World Data Centre for Meteorology - https://www.ncdc.noaa.gov/ibtracs/index.php?name=wmo-data, spanning from 1985 and 2015. Global level scleractinian coral richness data was compiled from3 and digitalized as shape files representing layers with <50, 51–100, 101–200, 201–300 and >400 species. Satellite image is an Ikonos - 2, with spatial resolution of 4 m. A supervised classification was carried out using Mahalanobis Distance algorithm51. All explicit spatially data were introduced and processed in a Geographic Information System (GIS).
Data Availability
All data generated or analysed during this study are included in this published article and its Supplementary Information files.
References
Kiessling, W. Geologic and Biologic Controls on the Evolution of Reefs. Annual Review of Ecology, Evolution, and Systematics 40, 19, https://doi.org/10.1146/annurev.ecolsys.110308.120251 (2009).
Riding, R. Structure and composition of organic reefs and carbonate mud mounds: concepts and categories. Earth-Science Reviews 58, 163–231, https://doi.org/10.1016/S0012-8252(01)00089-7 (2002).
Veron, J. E. N. Corals in Space and Time: the Biogeography and Evolution of the Scleractinia. (University of New South Wales. Press, 1995).
Goldberg, W. N. The Biology of Reefs and Reef Organisms (The University of Chicago Press, 2013).
Spalding, M. D. et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. BioScience 57, 573–583, https://doi.org/10.1641/B570707 (2007).
Montaggioni, L. F. History of Indo-Pacific coral reef systems since the last glaciation: Development patterns and controlling factors. Earth-Science Reviews 71, 75, https://doi.org/10.1016/j.earscirev.2005.01.002 (2005).
Gischler, E. Quaternary reef response to sea-level and environmental change in the western Atlantic. Sedimentology 62, 36, https://doi.org/10.1111/sed.12174 (2015).
Chappell, J. Coral morphology, diversity and reef growth. Nature 286, 249, https://doi.org/10.1038/286249a0 (1980).
Pinheiro Hudson, T. et al. South‐western Atlantic reef fishes: Zoogeographical patterns and ecological drivers reveal a secondary biodiversity centre in the Atlantic Ocean. Diversity and Distributions 0, https://doi.org/10.1111/ddi.12729 (2018).
Leão, Z. M. A. N., Kikuchi, R. K. P. & Testa, V. in Latin America Coral Reefs (ed Jorge Cortés) 9–52 (Elsevier Science., 2003).
Moura, R. L. et al. Spatial patterns of benthic megahabitats and conservation planning in the Abrolhos Bank. Continental Shelf Research 70, 109–117, https://doi.org/10.1016/j.csr.2013.04.036 (2013).
Darwin, C. R. The struture and distribution of coral reefs. (Smith, Elder & Company, 1842).
Hartt, C. F. Geology and physical geography of Brazil., (Fields, Osgood & Co, 1870).
Bastos, A. C. et al. Shelf morphology as an indicator of sedimentary regimes: A synthesis from a mixed siliciclastic–carbonate shelf on the eastern Brazilian margin. Journal of South American Earth Sciences 63, 125–136, https://doi.org/10.1016/j.jsames.2015.07.003 (2015).
Laborel, J. L. Madreporaires et hydrocoralliaires recifaux des cotes brésiliennes. Systematique, ecologie, repartition verticale et geographie. Ann. Inst. Oceanogr 47, 171–229 (1969).
Leão, Z. M. A. N. & Ginsburg, R. N. In Proceedings of the 8th International Coral Reef Symosium 1767–1772.
Jesionek, M. B. et al. A taxonomic account of non-geniculate coralline algae (Croallinophycidae, Rhodophyta) from shallow reefs of the Abrolhos Bank, Brazil. Algae 31, 1–24 (2016).
Leão, Z. M. A. N. Morphology, Geology and Developmental History of the southernmost coral reefs of WesternAtlantic, Abrolhos Bank, Brazil PhD thesis, University of Miami (1982).
Moura, R. L. et al. An extensive reef system at the Amazon River mouth. Science Advances Vol. 2, https://doi.org/10.1126/sciadv.1501252 (2016).
Taylor, P. D. Bryozoans and palaeoenvironmental interpretation. Journal of The Palaeontological Society of India 50, 1–11 (2005).
Francini-Filho, R. B. et al. Dynamics of coral reef benthic assemblages of the Abrolhos Bank, eastern Brazil: inferences on natural and anthropogenic drivers. PLoS One 8, e54260, https://doi.org/10.1371/journal.pone.0054260 (2013).
Reis, V. M. et al. Carbonate Production by Benthic Communities on Shallow Coralgal Reefs of Abrolhos Bank, Brazil. PLoS One 11, e0154417, https://doi.org/10.1371/journal.pone.0154417 (2016).
Cuffey, R. J. et al. In Proceedings of Third International Coral Reef Symposium. 339–345.
Taylor, P. D. & James, N. P. Secular changes in colony-forms and bryozoan carbonate sediments through geological history. Sedimentology 60, 1184–1212, https://doi.org/10.1111/sed.12032 (2013).
Ernst, A. & Königshof, P. The role of bryozoans in fossil reefs—an example from the Middle Devonian of the Western Sahara. Facies 54, 613–620, https://doi.org/10.1007/s10347-008-0149-1 (2008).
Stanley, G. D. Jr. The history of early mesozoic reef communities: A three-step process. Palaios 3, 170–183, https://doi.org/10.2307/3514528 (1988).
Kiessling, W. Reef expansion during the Triassic: Spread of photosymbiosis balancing climatic cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 290, 11–19, https://doi.org/10.1016/j.palaeo.2009.03.020 (2010).
Taylor, P. D. & Allison, A. P. Bryozoan carbonates through time and space. Geology 26, 459–462, https://doi.org/10.1130/0091-761 (1998).
Sharples, A. G. W. D., Huuse, M., Hollis, C., J. M., T. & Taylor, P. D. Giant middle Eocene bryozoan reef mounds in the Great Australian Bight. Geology 42, 4, https://doi.org/10.1130/G35704.1 (2014).
James, N. P. et al. Origin of Late Pleistocene bryozoan reef mounds; Great Australian Bight. Journal of Sedimentary Research 74, 28 (2004).
Novak, V. et al. Environmental reconstruction of a late Burdigalian (Miocene) patch reef in deltaic deposits (East Kalimantan, Indonesia). Palaeogeography, Palaeoclimatology, Palaeoecology 374, 110–122, https://doi.org/10.1016/j.palaeo.2013.01.009 (2013).
Santodomingo, N. et al. A Diverse Patch Reef from Turbid Habitats in the MIddle Miocene (East Kalimantan, Indonesia). Palaios 30, 128–149, https://doi.org/10.2110/palo.2013.047 (2015).
Wilson, M. E. J. Development of Equatorial Delta-Front Patch Reefs During the Neogene, Borneo. Journal of Sedimentary Research 75, 114–133, https://doi.org/10.2110/jsr.2005.010 (2005).
Klicpera, A., Taylor, P. D. & Westphal, H. Bryozoans on the move: adaptations to hard substrate-limiting tropical heterozoan carbonates (Banc d’Arguin, Mauritania). Marine Biodiversity 45, 601–602, https://doi.org/10.1007/s12526-014-0279-3 (2015).
Kuklinski, P. & Bader, B. Diversity, structure and interactions of encrusting lithophyllic macrofaunal assemblages from Belgica Bank, East Greenland. Polar Biology 30, 709–717, https://doi.org/10.1007/s00300-006-0228-0 (2006).
Guan, Y., Hohn, S. & Merico, A. Suitable Environmental Ranges for Potential Coral Reef Habitats in the Tropical Ocean. Plos One 10, e0128831, https://doi.org/10.1371/journal.pone.0128831 (2015).
Dominguez, J. M. L. In Geology and Geomorphology of Holocene Coastal Barriers of Brazil Lecture Notes in Earth Sciences (eds Dillenburg Sergio, R. & Patrick, A. Hesp) Ch. 2, XVI, 380 (Springer-Verlag Berlin Heidelberg, 2009).
Bruce, T. et al. Abrolhos bank reef health evaluated by means of water quality, microbial diversity, benthic cover, and fish biomass data. PLoS One 7, e36687, https://doi.org/10.1371/journal.pone.0036687 (2012).
Silveira, C. B. et al. Microbial and sponge loops modify fish production in phase-shifting coral reefs. Environ Microbiol 17, 3832–3846, https://doi.org/10.1111/1462-2920.12851 (2015).
Ghisolfi, R. D. et al. Physical forcing mechanisms controlling the variability of chlorophyll-a over the Royal-Charlotte and Abrolhos Banks-Eastern Brazilian Shelf. PLoS One 10, e0117082, https://doi.org/10.1371/journal.pone.0117082 (2015).
Hallock, P. In The history and sedimentology of ancient reef systems (ed G.D. Stanley) 387–427 (Kluwer Academic and Plenum Publishers, 2001).
Segal, B. et al. Potential impacts of polar fronts on sedimentation processes at Abrolhos coral reef (South-West Atlantic Ocean/Brazil). Continental Shelf Research 28, 533–544, https://doi.org/10.1016/j.csr.2007.11.003 (2008).
Amado-Filho, G. M. et al. Mesophotic ecosystems of the unique South Atlantic atoll are composed by rhodolith beds and scattered consolidated reefs. Marine Biodiversity 46, 933–936, https://doi.org/10.1007/s12526-015-0441-6 (2016).
Gherardi, D. F. M. & Bosence, D. W. J. Composition and community structure of the coralline algal reefs from Atol das Rocas, South Atlantic, Brazil. Coral Reefs 19, 205–219, https://doi.org/10.1007/s003380000100 (2001).
Morgan, K. M., Perry, C. T., Smithers, S. G., Johnson, J. A. & Daniell, J. J. Evidence of extensive reef development and high coral cover in nearshore environments: implications for understanding coral adaptation in turbid settings. Scientific Reports 6, 29616, :https://doi.org/10.1038/srep29616 https://www.nature.com/articles/srep29616#supplementary-information (2016).
Browne, N. K., Smithers, S. G. & Perry, C. T. Coral reefs of the turbid inner-shelf of the Great Barrier Reef, Australia: An environmental and geomorphic perspective on their occurrence, composition and growth. Earth-Science Reviews 115, 1–20, https://doi.org/10.1016/j.earscirev.2012.06.006 (2012).
Rogers, C. S. Hurricanes and coral reefs: The intermediate disturbance hypothesis revisited. Coral Reefs 12, 127–137, https://doi.org/10.1007/BF00334471 (1993).
Connell, J. H. Diversity in Tropical Rain Forests and Coral Reefs. Science 199, 1302–1310, https://doi.org/10.1126/science.199.4335.1302 (1978).
Dechnik, B., Webster, J. M., Davies, P. J., Braga, J. C. & Reimer, P. J. Holocene “turn-on” and evolution of the Southern Great Barrier Reef: Revisiting reef cores from the Capricorn Bunker Group. Marine Geology 363, 174–190, https://doi.org/10.1016/j.margeo.2015.02.014 (2015).
Webster, J. M., Yokoyama, Y., Cotterill, C. & Expedition 325 Scientists. In Proceedings of the Integrated Ocean Drilling Program (Integrated Ocean Drilling Program Management International, Inc., for the Integrated Ocean Drilling Program, 2011).
Richards, J. A. Remote Sensing Digital Image Analysis. (Springer-Verlag, 1999).
Acknowledgements
We thank CNPq/FAPES-Sisbiota/PELD, CAPES/IODP, CAPES/Ciências do Mar, and ANP/Brasoil for long term project funding. We also thank ICMBio for research permits and field logistic support, and Conservation International for providing and authorizing the use of the IKONOS image. JMW and JCB are International Visiting Researcher at UFES and JBRJ, supported by the Science Without Borders program. Zá Cajueiro provided invaluable field support and Ronaldo Francini, Carlos Janovitch and Lucio Engler helped in the drilling operations. This is a contribution from the Rede Abrolhos (abrolhos.org).
Author information
Authors and Affiliations
Contributions
A.C.B., R.L.M. and G.M.A.F. designed the experiment. A.C.B., L.S.V., R.L.M. and U.R.M. carried out the statistical and geospatial analyses. A.C.B., R.L.M., L.S.V., G.M.A.F., J.C.B. and J.M.W. analyzed the cores. A.C.B., R.L.M., F.C.M. and G.M.A.F. obtained the samples and compiled secondary data. L.V.R. identified the bryozoans. All authors reviewed the manuscript and provided relevant inputs to its final version.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bastos, A.C., Moura, R.L., Moraes, F.C. et al. Bryozoans are Major Modern Builders of South Atlantic Oddly Shaped Reefs. Sci Rep 8, 9638 (2018). https://doi.org/10.1038/s41598-018-27961-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-27961-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
